Back to Journals » Cancer Management and Research » Volume 12
Long Non-Coding RNA DARS-AS1 Contributes to Prostate Cancer Progression Through Regulating the MicroRNA-628-5p/MTDH Axis
Authors Fan H, Hou J, Liu S, Xiao Z, Cui J
Received 6 July 2020
Accepted for publication 19 August 2020
Published 10 September 2020 Volume 2020:12 Pages 8363—8377
DOI https://doi.org/10.2147/CMAR.S271021
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
This paper has been retracted.
Haitao Fan,1 Junhui Hou,2 Siqing Liu,3 Zuomin Xiao,4 Jia Cui5
1Department of Urology, The Second Hospital of Jilin University, Changchun, Jilin 130041, People’s Republic of China; 2Department of Oncology & Radiotherapy, Qingdao Central Medical Group, Qingdao, Shandong 266000, People’s Republic of China; 3Department of Outpatient, Qingdao Special Service Sanatorium of PLA Navy, Qingdao, Shandong 266071, People’s Republic of China; 4Department of Clinical Laboratory, Jinan Jigang Hospital, Jinan, Shandong 250101, People’s Republic of China; 5Department of Oncology, The Second Hospital of Dalian Medical University, Dalian, Liaoning 116023, People’s Republic of China
Correspondence: Jia Cui
Department of Oncology, The Second Hospital of Dalian Medical University, Dalian, Liaoning 116023, People’s Republic of China
Email [email protected]
Purpose: DARS antisense RNA 1 (DARS-AS1) is a long non-coding RNA that has been validated as a critical regulator in several human cancer types. Our study aimed to determine the expression profile of DARS-AS1 in prostate cancer (PCa) tissues and cell lines. Functional experiments were conducted to explore the detailed roles of DARS-AS1 in regulating PCa carcinogenesis. Furthermore, the detailed mechanisms by which DARS-AS1 regulates the oncogenicity of PCa cells were uncovered.
Methods: Reverse transcription quantitative polymerase chain reaction was performed to analyze DARS-AS1 expression in PCa tissues and cell lines. Cell Counting Kit-8 assays, flow cytometry analyses, Transwell assays, and tumor xenograft experiments were conducted to determine the regulatory effects of DARS-AS1 knockdown on the malignant phenotype of PCa cells. Bioinformatics analysis was performed to identify putative microRNAs (miRNAs) targeting DARS-AS1, and the direct interaction between DARS-AS1 and miR-628-5p was verified using RNA immunoprecipitation and luciferase reporter assays.
Results: DARS-AS1 was highly expressed in PCa tissues and cell lines. In vitro functional experiments demonstrated that DARS-AS1 depletion suppressed PCa cell proliferation, promoted cell apoptosis, and restricted cell migration and invasion. In vivo studies revealed that the downregulation of DARS-AS1 inhibited PCa tumor growth in nude mice. Mechanistic investigation verified that DARS-AS1 functioned as an endogenous miR-628-5p sponge in PCa cells and consequently promoted the expression of metadherin (MTDH). Furthermore, the involvement of miR-628-5p/MTDH axis in DARS-AS1-mediated regulatory actions in PCa cells was verified using rescue experiments.
Conclusion: DARS-AS1 functioned as a competing endogenous RNA in PCa by adsorbing miR-628-5p and thereby increasing the expression of MTDH, resulting in enhanced PCa progression. The identification of a novel DARS-AS1/miR-628-5p/MTDH regulatory network in PCa cells may offer a new theoretical basis for the development of promising therapeutic targets.
Keywords: DARS antisense RNA 1, non-coding RNA, ceRNA theory, target therapy
Introduction
Prostate cancer (PCa) is the second most frequent human cancer among men.1 It is also the seventh leading cause of tumor-related deaths worldwide, and an estimated 359,000 patients died of PCa in 2018 globally.2,3 The primary treatment alternatives, including surgical resection, radiotherapy, chemotherapy, and hormone therapy, have achieved significant progress and can effectively treat most patients with PCa diagnosed at an early stage.4 However, a large number of patients die within 5 years after their initial diagnosis.5 The poor clinical outcomes of patients with PCa are largely caused by the fact that more than 30% of cases experience biochemical recurrence and are diagnosed at advanced stages.6 Furthermore, approximately 10% of newly diagnosed PCa cases show evidence of a locally advanced stage, and 5% develop distant metastases, which reduces the opportunity for further therapy.7 Although great efforts have been put to study this tumor, the mechanisms underlying PCa formation and progression remain elusive and poorly understood.8 Hence, the investigation of genes contributing to the molecular pathogenesis in PCa may allow the identification of novel clinical markers and therapeutic targets.
In the past decades, the critical functions of long non-coding RNAs (lncRNAs) in human diseases have received increasing attention.9,10 They are a type of transcript without protein-coding ability characterized by a length of over 200 nucleotides, but some debate exists.11 Several studies have revealed the involvement of lncRNAs in a wide range of physiological and pathological processes.12–14 Of note, the altered expression of lncRNAs is closely correlated with the pathogenesis of PCa, in which they participate in modulating the pathological processes of PCa cells.15 As such, several lncRNAs are dysregulated in PCa and play pro-oncogenic or anti-oncogenic roles during PCa oncogenesis and progression.16–18
MicroRNAs (miRNAs) are a class of short non-coding regulatory RNAs with 17–24 nucleotides.19 miRNAs directly bind to the 3′-untranslated (UTR) region of their target genes through complementary base pairing, triggering target mRNA degradation and/or suppressing translation.20 In recent years, a competing endogenous RNA (ceRNA) theory has been proposed and has gained widespread acceptance. According to this theory, lncRNAs function as endogenous molecular sponges for certain miRNAs and consequently attenuate the ability of miRNAs to suppress their target genes.21,22 Altogether, the above evidence highlights the important roles of lncRNAs and miRNAs and suggests their potential as molecular targets for anticancer treatments in PCa.
The long non-coding RNA DARS antisense RNA 1 (DARS-AS1) has been identified as a crucial regulator in thyroid cancer,23 ovarian cancer,24 non-small-cell lung cancer,25 and renal cell carcinoma.26 However, the expression profile and roles of DARS-AS1 in PCa has not been reported. Here, we determined the expression of DARS-AS1 in PCa tissues and cell lines. The detailed roles of DARS-AS1 in regulating PCa carcinogenesis were explored using a series of functional experiments. Furthermore, the mechanisms underlying the regulation of the oncogenicity of PCa cells by DARS-AS1 were revealed in detail. To our knowledge, this is the first study on the role of DARS-AS1 in PCa pathogenesis and the mechanisms involved. Our findings may provide a scientific basis for the search of promising therapeutic targets.
Materials and Methods
Human Tissue Samples
This study was approved by the Ethics Committee of the Second Hospital of Dalian Medical University (2017SHDLMU-0617) and complied with the Declaration of Helsinki. All participants signed informed consent forms prior to their enrollment in this study. Fresh PCa tissues and adjacent normal tissues were obtained from 53 patients in the Second Hospital of Dalian Medical University, none of which had been diagnosed with other types of human cancer or received preoperative anticancer treatments. After resection, all fresh tissues were immediately frozen in liquid nitrogen and then immersed in liquid nitrogen until RNA extraction. The clinical and demographic characteristics of all subjects enrolled in the study are summarized in Table 1.
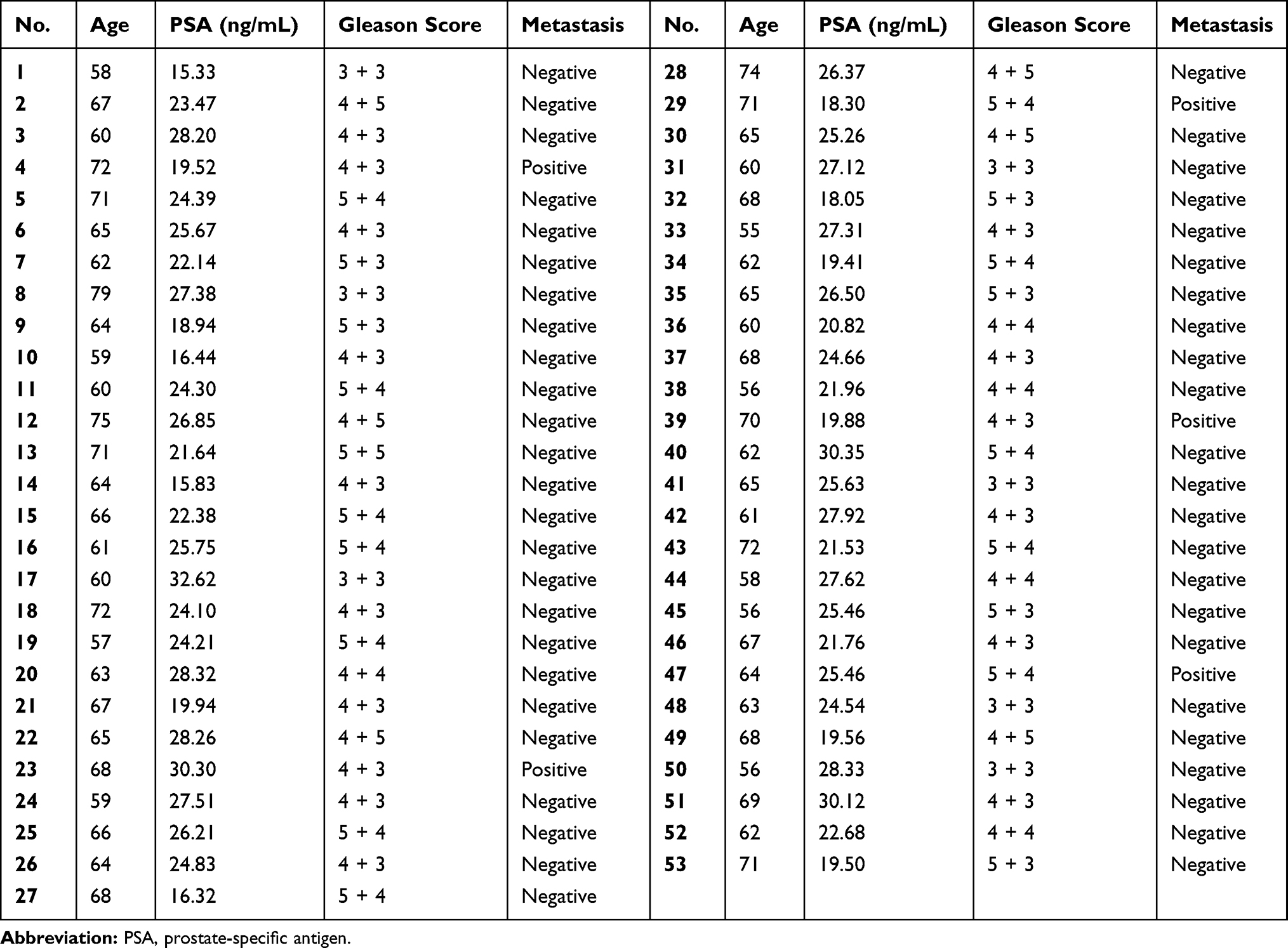 |
Table 1 The Clinical and Demographic Characteristics of All Subjects Enrolled in the Study |
Cell Culture
The human normal prostate epithelial cell line RWPE-1 was obtained from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China) and grown in Keratinocyte-SFM medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with a gentamicin/amphotericin solution (Gibco; Thermo Fisher Scientific, Inc.,). RPMI Medium 1640 containing 10% fetal bovine serum (FBS) and 1% sodium pyruvate (all from Gibco; Thermo Fisher Scientific, Inc.,) was used for the culture of LNCaP and 22RV1 PCa cell lines (the Type Culture Collection of the Chinese Academy of Sciences). The culture medium of LNCaP cells contained 1% Glutamax (Gibco; Thermo Fisher Scientific, Inc.,). The other two PCa cell lines PC-3 and DU145 were respectively grown in F-12 culture medium (Gibco; Thermo Fisher Scientific, Inc.,) and Dulbecco’s modified Eagle’s medium (Gibco; Thermo Fisher Scientific, Inc.,), respectively, both of which were supplemented with 10% FBS. All cells were maintained in a sterilized incubator at 37°C with 5% CO2.
Plasmid, Oligonucleotide, and siRNA Transfections
The small interfering RNAs (siRNAs) targeting DARS-AS1 (si-DARS-AS1) and scramble negative control siRNA (si-NC) were obtained from GenePharma Company (Shanghai, China). The metadherin (MTDH)-overexpressing plasmid (pcDNA3.1-MTDH) and empty pcDNA3.1 plasmid were designed and chemically synthesized by Genechem Company (Shanghai, China). miR-628-5p mimic (RiboBio; Guangzhou, China) was transfected into PCa cells to increase endogenous miR-628-5p expression, with mimic control used as the control. miR-628-5p inhibitor (RiboBio) was used for silencing miR-628-5p expression, and inhibitor control acted as the control.
For cell transfection, PCa cells were seeded in 6-well plates, and transfections with plasmids, oligonucleotides, or siRNAs were performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) as the carrier. After 6-h incubation, the medium was replaced with fresh culture medium.
Nucleus-Cytoplasm Fractionation Assay
The separation and purification of cytoplasmic and nuclear RNA were conducted using the Cytoplasmic and Nuclear RNA Purification Kit (Norgen, Belmont, CA, USA). The isolated RNA was analyzed by reverse transcription quantitative polymerase chain reaction (RT-qPCR). Glycerol 3-phosphate dehydrogenase (GAPDH) and U6 small nuclear RNA expression levels were measured as the cytoplasmic and nuclear references, respectively.
RNA Extraction and RT-qPCR
Total RNA was isolated with the TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,). A Thermo NanoDrop 2000 Spectrophotometer was used to determine the concentration and quality of total RNA, which was reverse transcribed into complementary DNA (cDNA) using a QuantiTect Reverse Transcription Kit (Qiagen GmbH, Hilden, Germany). The expression levels of DARS-AS1 and MTDH were detected by quantitative PCR with the QuantiTect SYBR Green PCR Kit (Qiagen GmbH). To analyze miR-628-5p expression, reverse transcription was conducted using the miScript Reverse Transcription Kit (Qiagen GmbH). The miScript SYBR Green PCR Kit (Qiagen GmbH) was used for quantitative PCR. Using the 2−ΔΔCt method, miR-628-5p expression was normalized to that of U6 small nuclear RNA, and GAPDH served as the control for DARS-AS1 and MTDH mRNA expression.
Cell Counting Kit-8 (CCK-8) Assay
Transfected cells were collected after 24-h incubation and seeded into 96-well plates. Each well was covered with a 100 μL cell suspension containing 2 × 103 cells. Cell proliferation was examined by incubating cells with 10 μL of the CCK-8 solution (Dojindo Laboratories, Kumamoto, Japan). Two hours later, the optical density at a wavelength of 450 nm (OD450) was measured using a SUNRISE Microplate Reader (Tecan Group, Ltd., Mannedorf, Switzerland). All assays were performed with six duplications and repeated three times.
Flow Cytometry
An annexin V–fluorescein Isothiocyanate (FITC) Apoptosis Detection Kit (BioLegend, Inc., San Diego, CA, USA) was used to quantify cell apoptosis. Twenty-four hours later, transfected cells were harvested and rinsed with cold phosphate-buffered saline, followed by centrifugation and removal of the supernatant. The resultant cells were then resuspended in 100 µL of 1× binding buffer and double stained with 5 µL of annexin V–FITC and 5 µL of propidium iodide. Following a 15 min incubation in the dark, the fraction of apoptotic cells was obtained using a flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Transwell Migration and Invasion Assays
For migration assays, cells were collected with trypsin at 24-h post-transfection and used to prepare single-cell suspensions in FBS-free basal medium. The upper compartment of Transwell chambers (BD Biosciences) was filled with a 100 µL cell suspension containing 5 × 104 cells, and 600 µL of culture medium supplemented with 20% FBS was added into the lower compartments. After 24 h, the upper surface of the membranes was cleaned with a cotton bud, and the migrated cells were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet. Following three washes with phosphate-buffered saline, the migrated cells were photographed under a light microscope (Olympus Corporation, Tokyo, Japan), and five randomly selected fields were analyzed to determine the number of migrated cells. For invasion assays, the Transwell chambers were precoated with Matrigel (BD Biosciences), and the remainder experimental steps were the same as those of migration assays.
Tumor Xenograft Experiments
The animal-related experimental procedures were approved by the Animal Research Ethics Committee of the Second Hospital of Dalian Medical University (2019SHDLMU-0308) and conducted in accordance with NIH guidelines for the care and use of laboratory animals. The lentiviral viruses harboring DARS-AS1 short hairpin (sh) RNA (sh-DARS-AS1) or scramble negative control shRNA (sh-NC) were purchased from Genechem Company and transfected into DU145 cells to obtain the DARS-AS1 stably depleted cell line. For in vivo assays, male BALB/c nude mice (4–6 weeks old) were bought from the Shanghai Experimental Animal Center of the Chinese Academy of Sciences (Shanghai, China) and subcutaneously injected with 1 × 107 DU145 cells stably overexpressing sh-DARS-AS1 or sh-NC. Tumor sizes were recorded weekly using a caliper, and the volume of subcutaneous tumors was calculated using the formula: Volume = (length × width2)/2. All mice were euthanized at week 5, and subcutaneous tumors were resected and photographed. After weighing the tumors, total RNA and protein samples were extracted and used for molecular analyses.
Bioinformatics Analysis
The identification of putative miRNAs targeting DARS-AS1 was conducted using StarBase version 3.0 (http://starbase.sysu.edu.cn/). Three bioinformatics tools, including StarBase 3.0, TargetScan (http://www.targetscan.org/), and miRDB (http://mirdb.org/), were employed to determine the candidate target genes of miR-628-5p.
RNA Immunoprecipitation (RIP) Assay
RIP assays were performed using the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore, Bedford, MA, USA). PCa cells in the logarithmic growth phase were treated with RIP lysis buffer. Afterward, a 100-μL aliquot of the cell lysate was probed with RIP buffer containing magnetic beads conjugated with a human anti-Ago2 antibody (Millipore) or negative control IgG (Millipore). The magnetic beads were collected after overnight incubation at 4°C, followed by the addition of Proteinase K to digest the protein. The immune precipitated RNA was extracted, and the relative enrichment of DARS-AS1 and miR-628-5p was measured by RT-qPCR.
Luciferase Reporter Assay
The fragments of DARS-AS1 and the MTDH 3′-UTR containing miR-628-5p binding sites were chemically synthesized and inserted into the pmirGLO luciferase vector (Promega Corporation, Madison, WI, USA), and the generated luciferase reporter vectors were named DARS-AS1-wild-type (WT) and MTDH-WT, respectively. The DARS-AS1 and MTDH 3ʹ-UTR mutated fragments were obtained with a GeneTailor™ Site-Directed Mutagenesis System (Invitrogen, Carlsbad, CA, USA), and the synthesized reporter vectors were termed DARS-AS1-mutant (MUT) and MTDH-MUT, respectively. The miR-628-5p mimic or mimic control alongside the WT or MUT reporter vector was transfected into PCa cells using Lipofectamine 2000. At 48 h following transfection, the Dual-Luciferase Reporter Assay System (Promega) was used to measure luciferase activity.
Western Blot Analysis
Total protein from cells or tumor xenografts were extracted using RIPA lysate buffer (Beyotime Biotechnology; Shanghai, China). The BCA Protein Assay Kit (Beyotime Biotechnology) was used for total protein quantification. Equal quantities of proteins were resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto polyvinylidene difluoride membranes. After being blocked with 5% skim milk powder in Tris-buffered saline containing 0.1% Tween-20 (TBST) at room temperature for 2 h, membranes were incubated with a 1:1000 dilution of primary antibodies against MTDH (ab124789; Abcam, Cambridge, USA) or GAPDH (ab181602; Abcam) overnight at 4°C. The membranes were washed with TBST three times and incubated with a horseradish peroxidase-conjugated secondary antibody (ab205718; Abcam) for 1 h at room temperature. Protein signals were detected using an Enhanced Chemiluminescence Western Blotting Detection Reagent (GE Healthcare Life Sciences).
Statistical Analysis
All results were obtained from at least three biological replicates and expressed as the mean ± standard error. One-way analysis of variance followed by Tukey’s post hoc test was used in the comparisons among multiple groups. The differences between two groups were analyzed with Student’s t-test. Pearson’s correlation coefficient was determined for correlation analysis between DARS-AS1 and miR-628-5p expressions. All statistical analyses were performed with SPSS 17.0 software (SPSS Inc., Chicago, USA), and P-value of <0.05 was considered statistically significant.
Results
DARS-AS1 Knockdown Inhibits PCa Cell Proliferation, Migration, and Invasion and Induces Cell Apoptosis in vitro
To address the functions of DARS-AS1 in PCa, its expression profile was first analyzed in TCGA and GTEx databases using GEPIA (http://gepia.cancer-pku.cn/index.html). As presented in Figure 1A, DARS-AS1 was found to be highly expressed in PCa tissues (n = 492) compared with that in normal prostate tissues (n = 152). Next, RT-qPCR analysis was conducted to determine DARS-AS1 expression in 53 pairs of PCa tissues and adjacent normal tissues. The results showed that the expression level of DARS-AS1 was approximately three times higher in PCa tissues than that in adjacent normal tissues (Figure 1B, P = 0.00008). Similarly, DARS-AS1 overexpression was confirmed in PCa cell lines (Figure 1C). Furthermore, the clinical relevance of DARS-AS1 in patients with PCa was examined using GEPIA. No correlation was identified between DARS-AS1 expression and overall survival (Figure 1D) or disease-free survival (Figure 1E) in patients with PCa.
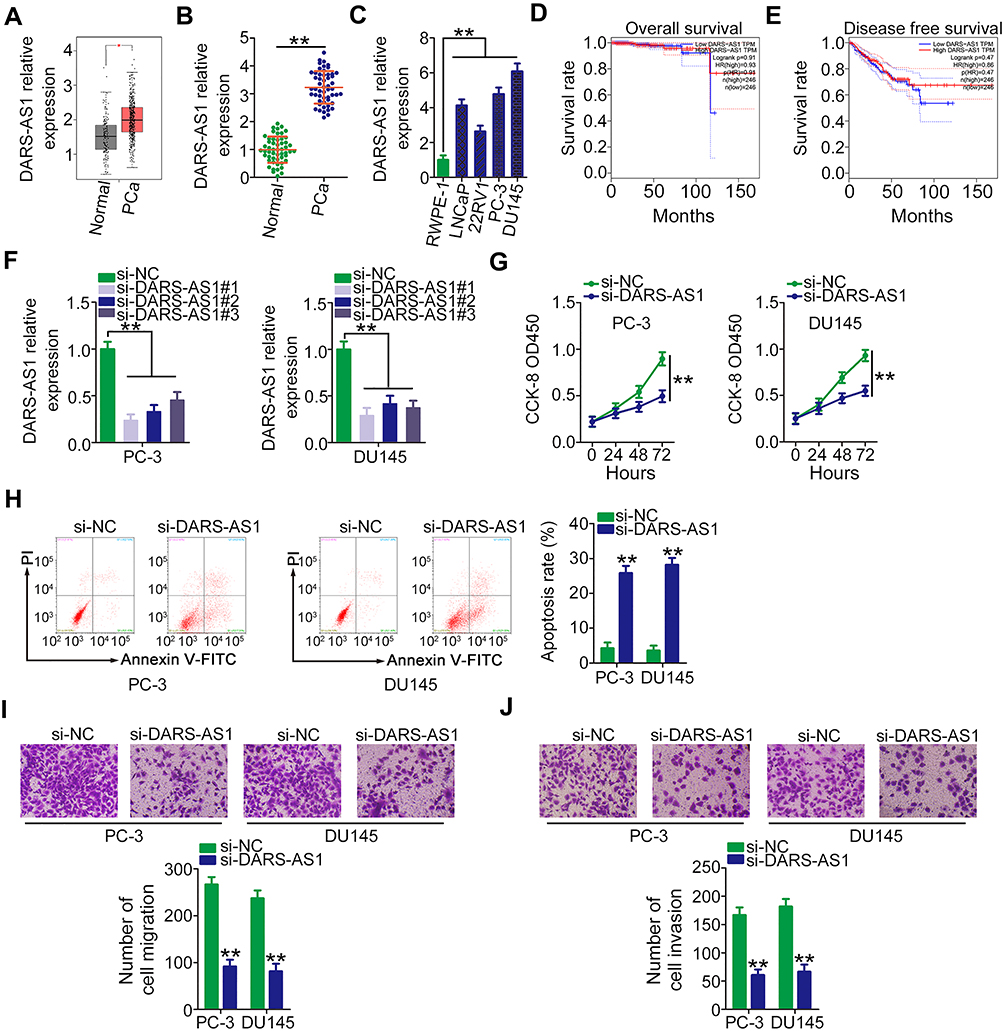 |
Figure 1 DARS-AS1 depletion suppresses PCa cell proliferation, migration, and invasion and promotes cell apoptosis. (A) DARS-AS1 expression in PCa tissues and normal tissues was determined using TCGA and GTEx databases. (B) RT-qPCR analysis was performed to detect DARS-AS1 expression in 53 pairs of PCa tissues and adjacent normal tissues. (C) DARS-AS1 expression in four PCa cell lines and the human normal prostate epithelial cell line RWPE-1. (D, E) GEPIA was used to examine the correlation between DARS-AS1 expression and overall survival and disease-free survival in PCa. (F) The efficiency of DARS-AS1 silencing in PC-3 and DU145 cells by si-DARS-AS1 was evaluated by RT-qPCR. (G) CCK-8 assay was performed to determine the proliferation of PC-3 and DU145 cells after DARS-AS1 downregulation. (H) Flow cytometry analysis revealed the effect of DARS-AS1 silencing on the apoptosis of PC-3 and DU145 cells. (I, J) The migratory and invasive capacities of DARS-AS1-silenced PC-3 and DU145 cells were measured via Transwell assays. **P < 0.01. |
Three siRNAs targeting DARS-AS1 (si-DARS-AS1) were used to silence DARS-AS1 expression in PC-3 and DU145 cells, and the silencing efficiency was evaluated by RT-qPCR. si-DARS-AS1#1 presented the highest efficiency (Figure 1F) and was selected for subsequent experiments. CCK-8 assay was conducted to test whether DARS-AS1 regulates cell proliferation. The results confirmed that the proliferation of PC-3 and DU145 cells was obviously suppressed by DARS-AS1 silencing (Figure 1G). Additionally, knocking down DARS-AS1 enhanced the apoptotic proportion of PC-3 and DU145 cells, as evidenced by flow cytometry analyses (Figure 1H). Furthermore, Transwell assays were carried out to investigate the effects of DARS-AS1 depletion on the migration and invasion of PCa cells. The migratory (Figure 1I) and invasive (Figure 1J) capacities of PC-3 and DU145 cells were substantially impaired by the loss of DARS-AS1. Taken together, our results indicated that DARS-AS1 was highly expressed in PCa and performed pro-oncogenic actions to promote cancer progression.
DARS-AS1 Functions as a ceRNA of miR-628-5p in PCa Cells
To investigate the mechanisms associated with the oncogenic roles of DARS-AS1 in PCa, the lncRNA subcellular localization predictor lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/) was used to identify the localization of DARS-AS1. The results showed that DARS-AS1 was predominantly distributed in the cytoplasm (Figure 2A). The nucleus-cytoplasm fractionation assay results further confirmed the cytoplasmic localization of DARS-AS1 in PC-3 and DU145 cells (Figure 2B), suggesting the post-transcriptional modulation of DARS-AS1. Based on the ceRNA theory, lncRNAs are involved in post-transcriptional processing; thus, StarBase 3.0 was used to predict the potential miRNAs targeting DARS-AS1. In total, seven miRNAs (miR-552-3p, miR-628-5p, miR-188-5p, miR-6866-3p, miR-3200-5p, miR-370-3p, and miR-6893-3p) were found to harbor a complementary sequence to DARS-AS1.
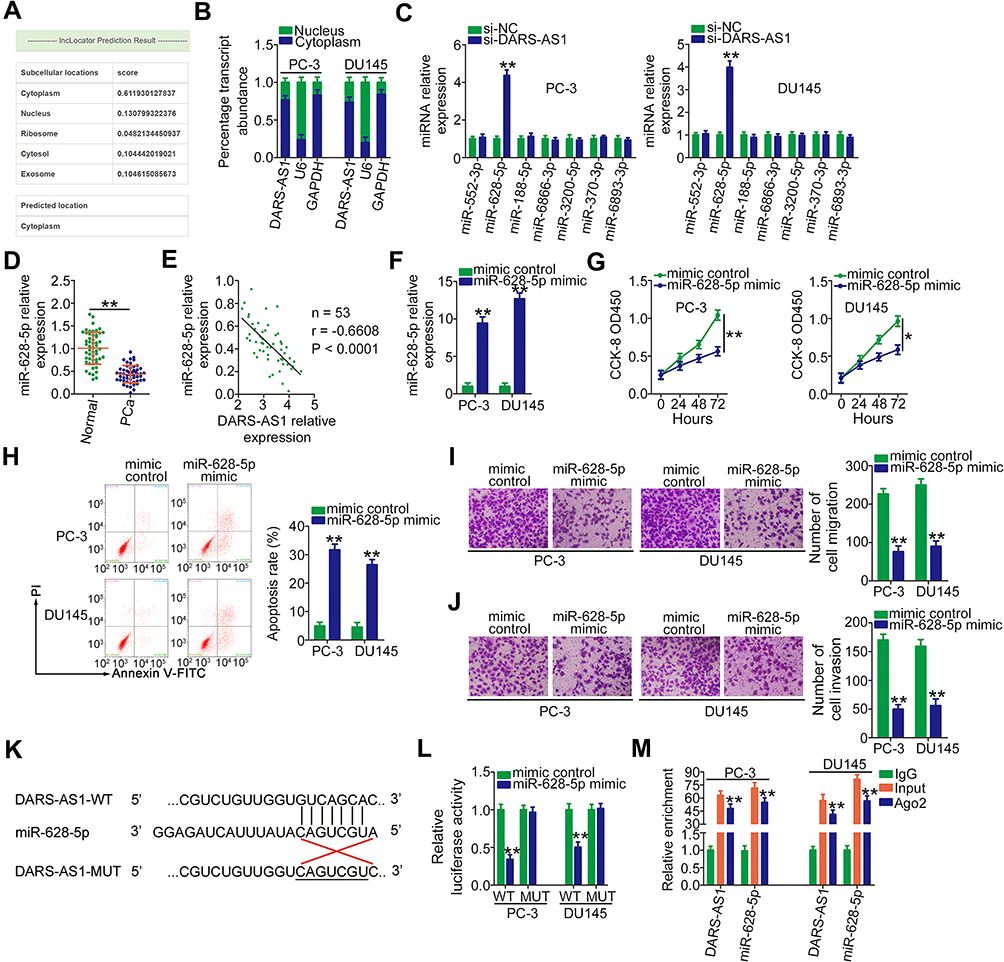 |
Figure 2 DARS-AS1 acts as a miR-628-5p sponge in PCa cells. (A) lncLocator predicted the cytoplasmic localization of DARS-AS1. (B) Relative amounts of DARS-AS1 in the cytoplasm and nucleus of PC-3 and DU145 cells were analyzed via RT-qPCR. (C) RT-qPCR was performed to detect miRNA (miR-552-3p, miR-628-5p, miR-188-5p, miR-6866-3p, miR-3200-5p, miR-370-3p, and miR-6893-3p) expression in PC-3 and DU145 cells after si-DARS-AS1 or si-NC transfection. (D) The expression level of miR-628-5p was detected by RT-qPCR in 53 pairs of PCa tissues and adjacent normal tissues. (E) Pearson’s correlation coefficient was applied to determine the correlation between the levels of DARS-AS1 and miR-628-5p in PCa tissues. (F) miR-628-5p expression in miR-628-5p mimic-transfected or mimic control-transfected PC-3 and DU145 cells was measured by RT-qPCR analysis. (G, H) CCK-8 assay and flow cytometry analysis were performed to evaluate the proliferation and apoptosis of miR-628-5p-overexpressing PC-3 and DU145 cells. (I, J) The impacts of miR-628-5p upregulation on the migration and invasion of PC-3 and DU145 cells were assessed by Transwell assays. (K) The predicted binding site between DARS-AS1 and miR-628-5p. The mutant sequences were also shown. (L) Luciferase reporter assays were performed in PC-3 and DU145 cells transfected with miR-628-5p mimic or mimic control to detect the luciferase activity of reporter DARS-AS1-WT and DARS-AS1-MUT. (M) The enrichment of miR-628-5p and DARS-AS1 in Ago2-containing immune precipitated RNA was quantified by RT-qPCR. *P < 0.05 and **P < 0.01. |
RT-qPCR was conducted to screen the miRNAs that may be sequestered by DARS-AS1 in PCa cells. Interestingly, miR-628-5p expression was increased following the interference of DARS-AS1 expression in PC-3 and DU145 cells, whereas the expression of the other six miRNAs was unaffected (Figure 2C). Next, miR-628-5p expression was determined in 53 pairs of PCa tissues and adjacent normal tissues. The results revealed that the expression of miR-628-5p was downregulated in PCa tissues to approximately 45% of that in normal tissues (Figure 2D, P = 0.0002). Interestingly, an inverse correlation between the DARS-AS1 and miR-628-5p levels in PCa tissues was validated through Pearson’s correlation coefficient analysis (Figure 2E; r = −0.6608, P < 0.0001). Then, the detailed roles of miR-628-5p in PCa cells were investigated. First, miR-628-5p expression was increased in PC-3 and DU145 cells transfected with miR-628-5p mimic (Figure 2F). The influences of miR-628-5p upregulation on PCa cell proliferation and apoptosis were tested by CCK-8 assays and flow cytometry analyses. The increased expression of miR-628-5p effectively suppressed cell proliferation (Figure 2G) and promoted cell apoptosis (Figure 2H) in PC-3 and DU145 cells. Furthermore, the migration (Figure 2I) and invasion (Figure 2J) of PC-3 and DU145 cells were clearly impaired by miR-628-5p overexpression.
Next, luciferase reporter assays were conducted in two PCa cell lines to examine the binding between miR-628-5p and DARS-AS1. The WT and mutant binding sites were shown in Figure 2K. The results of the luciferase reporter assay indicated that miR-628-5p upregulation reduced the luciferase activity of DARS-AS1-WT in PC-3 and DU145 cells (Figure 2L). When the binding sequences were mutated, miR-628-5p mimic did not affect the luciferase activity of DARS-AS1-MUT, indicating the direct binding of miR-628-5p to DARS-AS1. The RIP assay results further confirmed that miR-628-5p and DARS-AS1 were significantly enriched in the Ago2-containing beads compared with the levels in control IgG beads (Figure 2M). Collectively, these results demonstrated that DARS-AS1 functioned as a ceRNA in PCa cells by sequestering miR-628-5p.
DARS-AS1 Regulates MTDH Expression in PCa Cells by Sequestering miR-628-5p
The direct target recognition sequence of miR-628-5p was predicted via bioinformatics analysis, and MTDH was selected for further analysis due to its well-established as a critical regulator of PCa oncogenicity.27,28 The binding sequence of miR-628-5p to the 3ʹ-UTR of MTDH was presented in Figure 3A. Luciferase reporter assays were performed to validate the binding of miR-628-5p and the MTDH 3′-UTR in PCa cells. The data uncovered that miR-628-5p mimic efficiently lowered the luciferase activity of MTDH-WT in PC-3 and DU145 cells but had no effect on MTDH-MUT (Figure 3B). Subsequently, the role of miR-628-5p in regulating MTDH expression in PCa cells was determined by RT-qPCR and Western blotting. Unsurprisingly, miR-628-5p overexpression decreased MTDH mRNA (Figure 3C) and protein (Figure 3D) levels in PC-3 and DU145 cells. Furthermore, MTDH mRNA was relatively highly expressed in PCa tissues (Figure 3E) and inversely correlated with miR-628-5p expression (Figure 3F; r = −0.7207, P < 0.0001).
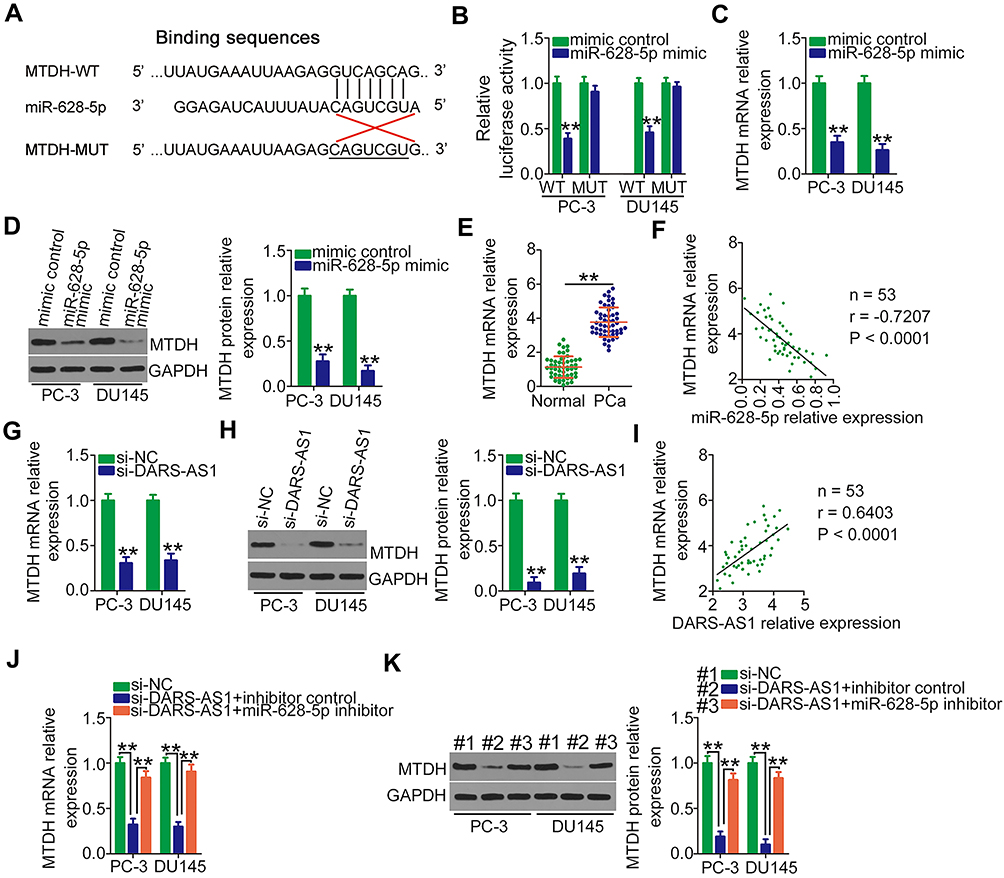 |
Figure 3 MTDH is a direct target of miR-628-5p and under the regulation of DARS-AS1 via the sequestering of miR-628-5p. (A) The wild-type and mutant binding sites of miR-628-5p to the MTDH 3′-UTR. (B) Luciferase activity of reporter MTDH-WT and MTDH-MUT was detected in PC-3 and DU145 cells transfected with miR-628-5p mimic or mimic control. (C, D) Relative mRNA and protein expressions of MTDH were detected in miR-628-5p-overexpressing PC-3 and DU145 cells by RT-qPCR and Western blotting, respectively. (E) RT-qPCR analysis was performed to detect MTDH mRNA expression in 53 pairs of PCa tissues and adjacent normal tissues. (F) Correlation between miR-628-5p and MTDH mRNA expression in PCa tissues was assessed by Pearson’s correlation coefficient. (G, H) RT-qPCR and Western blotting were conducted to measure MTDH mRNA and protein expressions in PC-3 and DU145 cells when DARS-AS1 was silenced. (I) Pearson’s correlation coefficient was determined to examine the association between DARS-AS1 and MTDH mRNA expression in PCa tissues. (J, K) PC-3 and DU145 cells were cotransfected with si-DARS-AS1 and miR-628-5p inhibitor or inhibitor control. The level of MTDH mRNA and protein expression was tested by RT-qPCR and Western blotting, respectively. **P < 0.01. |
Given the above results, we hypothesized that DARS-AS1 upregulates MTDH expression in PCa cells by sponging miR-628-5p. To test this hypothesis, the mRNA and protein levels of MTDH in PC-3 and DU145 cells transfected with si-DARS-AS1 or si-NC were measured. The results showed that both the levels of MTDH mRNA (Figure 3G) and protein (Figure 3H) were reduced by DARS-AS1 depletion. Notably, Pearson’s correlation coefficient analysis revealed a positive correlation between the expression of DARS-AS1 and MTDH mRNA in PCa tissues (Figure 3I; r = 0.6403, P < 0.0001). Moreover, the downregulation of miR-628-5p in PC-3 and DU145 cells restored the MTDH mRNA (Figure 3J) and protein (Figure 3K) expression suppressed by DARS-AS1 knockdown. Altogether, these results suggested that DARS-AS1 participated in a ceRNA pathway in PCa cells by sponging miR-628-5p and consequently promoting MTDH expression.
The miR-628-5p/MTDH Axis Mediates the Pro-Oncogenic Activities of DARS-AS1 in PCa Cells
Rescue experiments were performed to elucidate whether DARS-AS1 executed its oncogenic roles in PCa cells via the miR-628-5p/MTDH axis. First, RT-qPCR was used to evaluate the transfection efficiency of miR-628-5p inhibitor, and the data confirmed that miR-628-5p inhibitor effectively reduced endogenous miR-628-5p expression in PC-3 and DU145 cells (Figure 4A). CCK-8 assay showed that the decreased cell proliferation induced by si-DARS-AS1 was restored in PC-3 and DU145 cells after miR-628-5p inhibitor cotransfection (Figure 4B). Similarly, loss of DARS-AS1 promoted the apoptosis of PC-3 and DU145 cells, which was reversed by miR-628-5p inhibition (Figure 4C). In addition, the migratory (Figure 4D) and invasive (Figure 4E) abilities hindered by silencing DARS-AS1 were rescued via miR-628-5p inhibitor cotransfection. The overexpression of MTDH in PC-3 and DU145 cells was achieved via transfection with pcDNA3.1-MTDH (Figure 5A). The influences of DARS-AS1 interference on PC-3 and DU145 cell proliferation (Figure 5B), apoptosis (Figure 5C), migration (Figure 5D), and invasion (Figure 5E) were reversed by MTDH overexpression. Hence, the DARS-AS1/miR-628-5p/MTDH network was a modulatory of PCa progression.
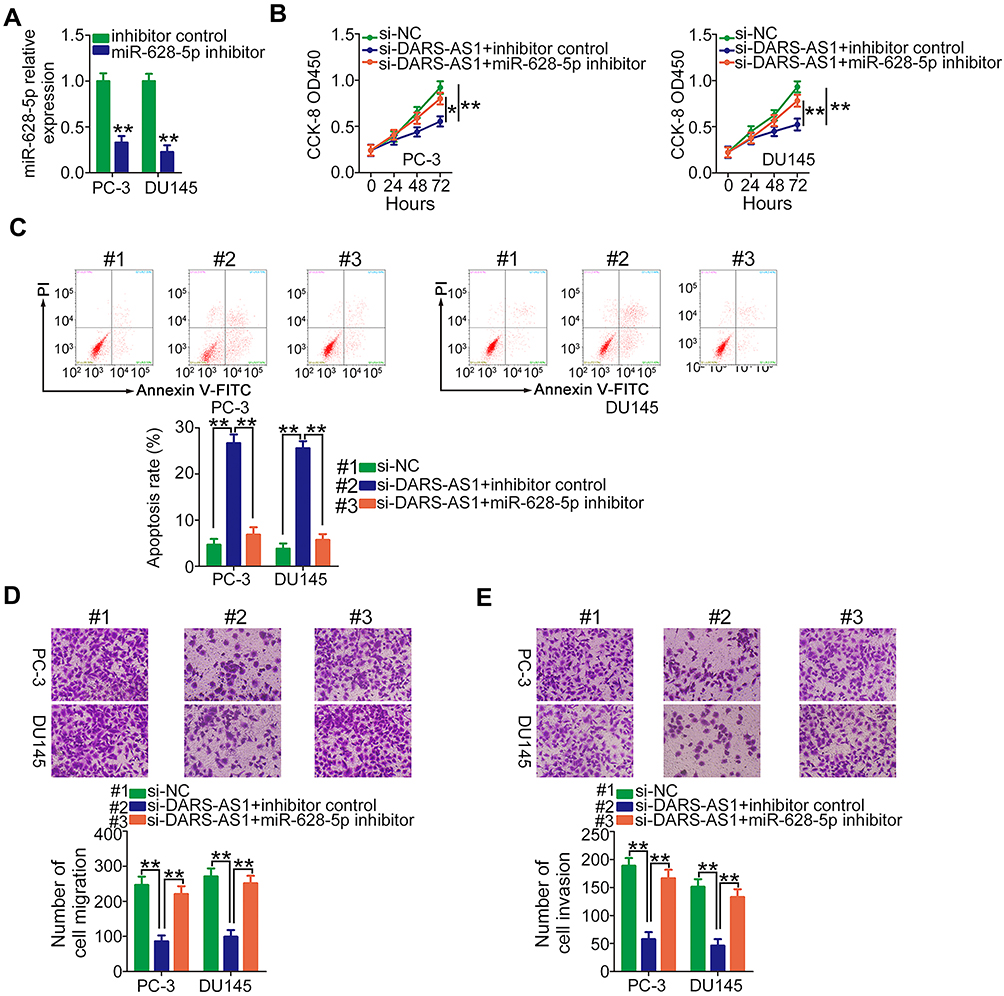 |
Figure 4 miR-628-5p inhibitor rescues the influences of si-DARS-AS1 on PCa cells. (A) RT-qPCR analysis of the transfection efficiency of miR-628-5p inhibitor in PC-3 and DU145 cells. (B–E) si-DARS-AS1, in combination with miR-628-5p inhibitor or inhibitor control, was transfected into PC-3 and DU145 cells. The transfected cells were subjected to CCK-8 assay, flow cytometry analysis, and Transwell assays for the determination of cell proliferation, apoptosis, and migration and invasion, respectively. *P<0.05 and **P < 0.01. |
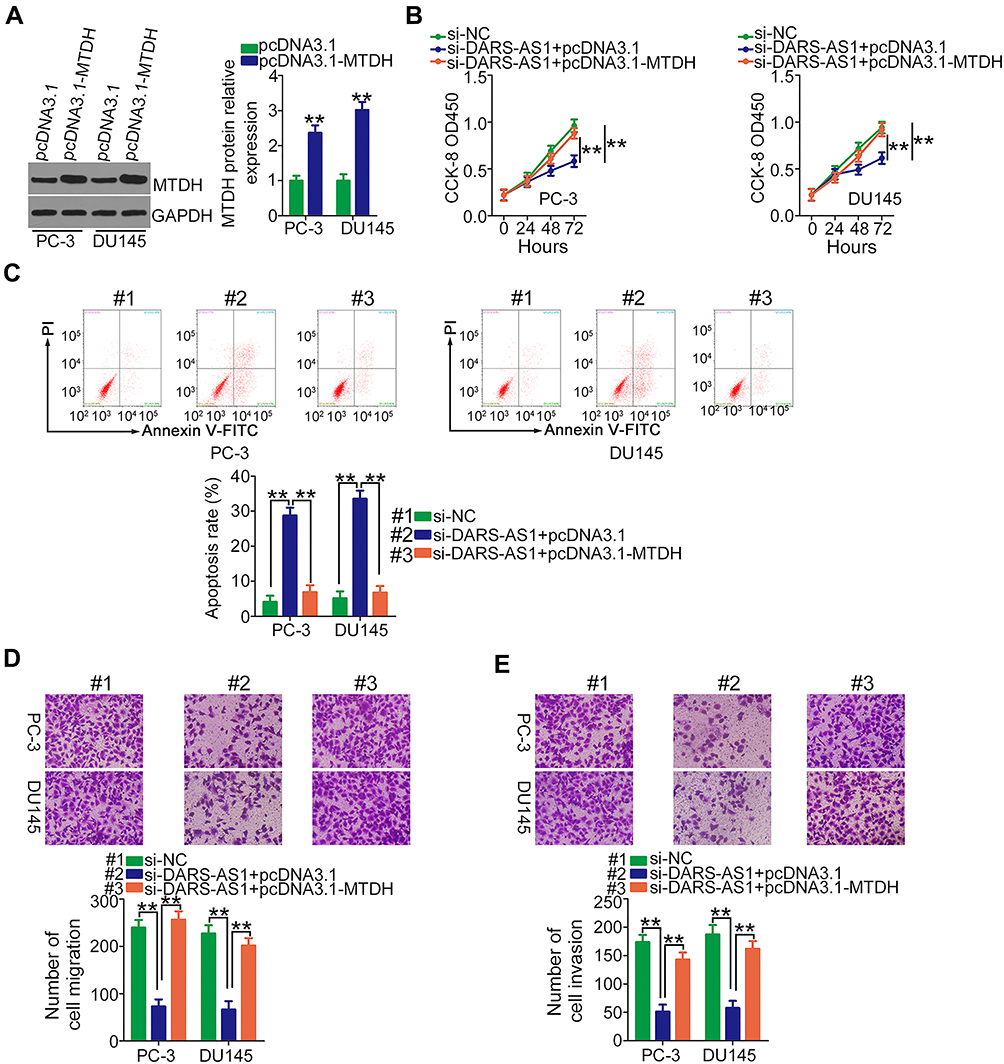 |
Figure 5 Restoring MTDH abrogates the inhibiting actions of DARS-AS1 knockdown on PCa cells. (A) The protein level of MTDH in pcDNA3.1-MTDH or pcDNA3.1-transfected PC-3 and DU145 cells was evaluated by Western blotting. (B, C) PC-3 and DU145 cells were transfected with si-DARS-AS1 in the presence of pcDNA3.1-MTDH or pcDNA3.1. Proliferation and apoptosis were respectively determined by CCK-8 assays and flow cytometry analyses. (D, E) Transwell assays were conducted to detect the migration and invasion of the cells described above. **P < 0.01. |
DARS-AS1 Represses PCa Tumor Growth in vivo
Finally, the impact of DARS-AS1 knockdown on PCa tumor growth in vivo was determined using tumor xenograft experiments. DU145 cells stably overexpressing sh-DARS-AS1 or sh-NC were subcutaneously inoculated into nude mice. The growth of tumor xenografts in the sh-DARS-AS1 group was significantly slower compared with the sh-NC group (Figure 6A and B). In addition, the weights of tumor xenografts were measured at the endpoint of the experiments. The tumor weight was striking decreased in the tumor xenografts originating from DARS-AS1-stably silenced DU145 cells (Figure 6C). Furthermore, the expression of DARS-AS1 was downregulated (Figure 6D), whereas miR-628-5p was upregulated (Figure 6E) in the tumors obtained from the sh-DARS-AS1 group. Western blotting analyses revealed that the protein level of MTDH was decreased in DARS-AS1-knockdown tumors (Figure 6F). These results indicated that loss of DARS-AS1 suppressed PCa tumor growth.
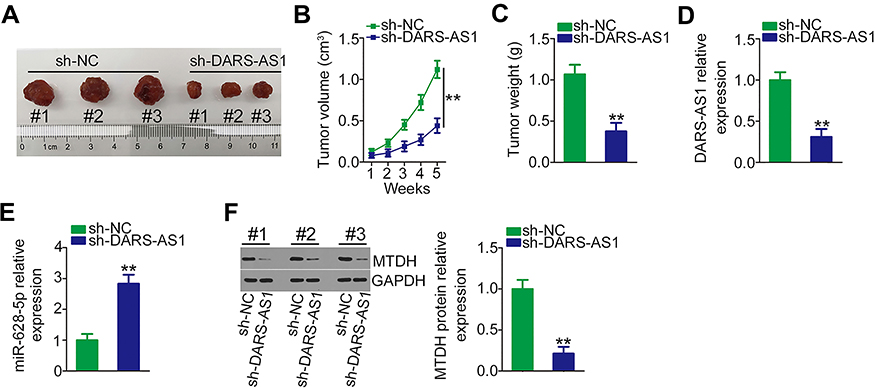 |
Figure 6 Depletion of DARS-AS1 attenuates PCa tumor growth in vivo. (A) Image of excised tumor xenografts from nude mice in the sh-DARS-AS1 and sh-NC groups. (B) The volumes of tumor xenografts in the sh-DARS-AS1 and sh-NC groups were monitored weekly. (C) The weights of tumor xenografts originating from DU145 cells stably overexpressing sh-DARS-AS1 or sh-NC. (D, E) The expression levels of DARS-AS1 and miR-628-5p in tumor xenografts collected from sh-DARS-AS1 and sh-NC groups were analyzed via RT-qPCR. (F) Western blotting was used for MTDH protein quantification in the tumor xenografts collected from the sh-DARS-AS1 and sh-NC groups. **P < 0.01. |
Discussion
An increasing number of studies have confirmed the critical roles of lncRNAs in various pathophysiological processes.29–31 Several lncRNAs are aberrantly expressed in PCa, and their expression is closely related to the overall survival of patients with PCa.32–34 In this regard, exploring the detailed roles of novel lncRNAs in PCa may offer promising strategies for cancer therapies. In total, 548 640 lncRNAs have been identified in the human genome;35 however, most lncRNAs in PCa have not been studied.36,37 In this work, our results identified a close relationship among DARS-AS1, miR-628-5p, and MTDH in PCa, indicating that the DARS-AS1/miR-628-5p/MTDH axis may contribute to PCa oncogenesis.
DARS-AS1 is highly expressed in thyroid cancer and significantly correlated with tumor stage and distant metastasis. Thyroid cancer patients with high DARS-AS1 expression have a lower overall survival rate than those with low DARS-AS1 expression.23 The overexpression of DARS-AS1 has also been verified in ovarian cancer,24 non-small-cell lung cancer,25 and renal cell carcinoma.26 To date, the expression profile of DARS-AS1 in PCa remains unknown, and further study is needed to elucidate whether DARS-AS1 critically contributes to PCa progression. In this study, DARS-AS1 expression in PCa was first analyzed in TCGA and GTEx databases using GEPIA. DARS-AS1 was found to be upregulated in PCa. RT-qPCR analysis further confirmed that DARS-AS1 levels were higher in PCa tissues than in adjacent normal tissues. The results of in vitro functional assays uncovered that DARS-AS1 silencing inhibited PCa cell proliferation, restricted cell migration and invasion, and promoted cell apoptosis. Furthermore, in vivo studies exhibited that downregulation of DARS-AS1 impaired tumor growth in nude mice.
The molecular events involved in DARS-AS1-triggered cellular progress in PCa were comprehensively elucidated. To this end, the subcellular localization of DARS-AS1 was predicted using lncLocator. DARS-AS1 was predominantly located in the cytoplasm, which was further verified by nucleus-cytoplasm fractionation assays. Thus, DARS-AS1 may function in post-transcriptional regulation in PCa. Recently, ceRNA regulatory networks have drawn great attention.38–40 In this network, lncRNAs function as ceRNAs by sponging certain miRNAs and thereby inhibiting the miRNA-mediated suppression of target mRNAs.41
In this study, the putative miRNAs that bind to DARS-AS1 were first identified. miR-628-5p was found to harbor a complementary sequence to DARS-AS1, and the binding interaction between miR-628-5p and DARS-AS1 in PCa cells was validated by luciferase reporter and RIP assays. Additionally, the expression of miR-628-5p was increased in PCa cells by transfecting with si-DARS-AS1. Furthermore, miR-628-5p exhibited weak expression in PCa tissues and was negatively correlated with DARS-AS1. These results identified DARS-AS1 as a novel miR-628-5p sponge in PCa.
Multiple studies have illustrated the expression and functions of miR-628-5p in human cancers.42–44 miR-628-5p has been reported to be downregulated in PCa.45 Herein, miR-628-5p was demonstrated to play a tumor-inhibiting role in PCa cells. Next, the molecular mechanisms by which miR-628-5p controls PCa progression were explored in detail. MTDH, also known as AEG-1,46 was identified as a direct target of miR-628-5p in PCa cells. In ceRNA networks, lncRNAs increase the expression of mRNAs by sequestering certain miRNAs. Hence, we next evaluated whether DARS-AS1 upregulates MTDH expression in PCa cells by interacting with miR-628-5p. DARS-AS1 interference led to a decrease in MTDH mRNA and protein levels in PCa cells, whereas these regulatory actions were abolished by cotransfecting with miR-628-5p inhibitor. Furthermore, MTDH was upregulated in PCa tissues and positively associated with DARS-AS1 expression. Altogether, the above results clearly identified a novel ceRNA regulatory network in PCa cells involving DARS-AS1, miR-628-5p, and MTDH.
MTDH, the first discovered in human fetal astrocytes in 2002,47 is overexpressed in PCa.48 MTDH performs a pro-oncogenic role in the regulation of PCa genesis and progression and is involved in the control of numerous malignant phenotypes.27,28 In the present study, our results indicated that MTDH was regulated by the DARS-AS1/miR-628-5p axis in PCa cells. DARS-AS1 can affect post-transcriptional processes and interfere with miR-628-5p in PCa cells by competing for shared miRNA response elements, consequently increasing the mRNA and protein levels of MTDH. Our rescue experiments further showed that the effects of DARS-AS1 silencing on PCa cell proliferation, apoptosis, migration, and invasion were reversed by miR-628-5p downregulation. Additionally, restored MTDH expression reversed the si-DARS-AS1-mediated anti-oncogenic actions in PCa cells. Overall, our findings suggest the implication of the miR-628-5p/MTDH axis in DARS-AS1-triggered biological processes.
Previous studies affirmed that the growth of PCa is androgen-dependent49 and that androgen receptor amplification has been demonstrated in PCa.50,51 Androgen receptor, a nuclear transcription factor and a steroid hormone receptor, was identified as the driver of PCa oncogeneis and progression.52 Androgen receptor signaling exerts important roles in the aggressive properties of PCa, and targeting of this signaling provides promising therapeutic benefit.53 Regarding the interaction between DARS-AS1 and androgen receptor, DARS-AS1 may be implicated in the control of androgen receptor signaling via a ceRNA way. The miRNAs targeting androgen receptor may be sponged by DARS-AS1, thereby forming a DARS-AS1/miRNAs/androgen receptor pathway.
Our study has two limitations. First, the sample size was small. Second, the effect of DARS-AS1 on androgen receptor signaling in PCa was not examined. These limitations will be addressed in future studies.
Conclusion
DARS-AS1 was upregulated in PCa and facilitated cancer progression. Mechanistic studies revealed that DARS-AS1 functioned as a ceRNA in PCa by adsorbing miR-628-5p and consequently increasing the expression of MTDH. As a result, the DARS-AS1/miR-628-5p/MTDH pathway (Figure 7) may have an important implication for the development of PCa treatments.
 |
Figure 7 Schematic diagram of the proposed mechanism. DARS-AS1 executes its pro-oncogenic roles in PCa cells by sponging miR-628-5p and consequently increasing MTDH expression. |
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Barsouk A, Padala SA, Vakiti A, et al. Epidemiology, Staging and Management of Prostate Cancer. Med Sci. 2020;8(3):3. doi:10.3390/medsci8030028
3. Whitaker H, Tam JO, Connor MJ, Grey A. Prostate cancer biology & genomics. Transl Androl Urol. 2020;9(3):1481–1491. doi:10.21037/tau.2019.07.17
4. Xie L, Li J, Wang X. Updates in prostate cancer detections and treatments - Messages from 2017 EAU and AUA. Asian j Urol. 2018;5(1):3–7. doi:10.1016/j.ajur.2017.11.004
5. Shepard DR, Raghavan D. Innovations in the systemic therapy of prostate cancer. Nat Rev Clin Oncol. 2010;7(1):13–21. doi:10.1038/nrclinonc.2009.187
6. Cornford P, Bellmunt J, Bolla M, et al. EAU-ESTRO-SIOG Guidelines on Prostate Cancer. Part II: treatment of Relapsing, Metastatic, and Castration-Resistant Prostate Cancer. Eur Urol. 2017;71(4):630–642. doi:10.1016/j.eururo.2016.08.002
7. May EJ, Viers LD, Viers BR, et al. Prostate cancer post-treatment follow-up and recurrence evaluation. Abdominal Radiol. 2016;41(5):862–876. doi:10.1007/s00261-015-0562-1
8. Nelson WG, De Marzo AM, DeWeese TL. The molecular pathogenesis of prostate cancer: implications for prostate cancer prevention. Urology. 2001;57(4 Suppl 1):39–45. doi:10.1016/S0090-4295(00)00939-0
9. Harries LW. Long non-coding RNAs and human disease. Biochem Soc Trans. 2012;40(4):902–906. doi:10.1042/BST20120020
10. Xiao Y, Xu J, Yin W. Aberrant Epigenetic Modifications of Non-coding RNAs in Human Disease. Adv Exp Med Biol. 2018;1094:65–75.
11. Fatica A, Bozzoni I. Long non-coding RNAs: new players in cell differentiation and development. Nat Rev Genet. 2014;15(1):7–21. doi:10.1038/nrg3606
12. Kim ED, Sung S. Long noncoding RNA: unveiling hidden layer of gene regulatory networks. Trends Plant Sci. 2012;17(1):16–21. doi:10.1016/j.tplants.2011.10.008
13. Nath A, Huang RS. Emerging role of long non-coding RNAs in cancer precision medicine. Mo Cell Oncol. 2020;7(1):1684130. doi:10.1080/23723556.2019.1684130
14. Ren X. Genome-wide analysis reveals the emerging roles of long non-coding RNAs in cancer. Oncol Lett. 2020;19(1):588–594. doi:10.3892/ol.2019.11141
15. Misawa A, Takayama KI, Inoue S. Long non-coding RNAs and prostate cancer. Cancer Sci. 2017;108(11):2107–2114. doi:10.1111/cas.13352
16. Xu YH, Deng JL, Wang G, Zhu YS. Long non-coding RNAs in prostate cancer: functional roles and clinical implications. Cancer Lett. 2019;464:37–55. doi:10.1016/j.canlet.2019.08.010
17. Smolle MA, Bauernhofer T, Pummer K, Calin GA, Pichler M. Current Insights into Long Non-Coding RNAs (LncRNAs) in Prostate Cancer. Int J Mol Sci. 2017;18(2):2. doi:10.3390/ijms18020473
18. Mitobe Y, Takayama KI, Horie-Inoue K, Inoue S. Prostate cancer-associated lncRNAs. Cancer Lett. 2018;418:159–166. doi:10.1016/j.canlet.2018.01.012
19. Mohr AM, Mott JL. Overview of microRNA biology. Semin Liver Dis. 2015;35(1):3–11. doi:10.1055/s-0034-1397344
20. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
21. Kartha RV, Subramanian S. Competing endogenous RNAs (ceRNAs): new entrants to the intricacies of gene regulation. Front Genet. 2014;5:8. doi:10.3389/fgene.2014.00008
22. Tay Y, Rinn J, Pandolfi PP. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–352. doi:10.1038/nature12986
23. Zheng W, Tian X, Cai L, et al. LncRNA DARS-AS1 regulates microRNA-129 to promote malignant progression of thyroid cancer. Eur Rev Med Pharmacol Sci. 2019;23(23):10443–10452. doi:10.26355/eurrev_201912_19683
24. Huang K, Fan WS, Fu XY, Li YL, Meng YG. Long noncoding RNA DARS-AS1 acts as an oncogene by targeting miR-532-3p in ovarian cancer. Eur Rev Med Pharmacol Sci. 2019;23(6):2353–2359. doi:10.26355/eurrev_201903_17379
25. Liu D, Liu H, Jiang Z, Chen M, Gao S. Long non-coding RNA DARS-AS1 promotes tumorigenesis of non-small cell lung cancer via targeting miR-532-3p. Minerva Med. 2019;1.
26. Jiao M, Guo H, Chen Y, Li L, Zhang L. DARS-AS1 promotes clear cell renal cell carcinoma by sequestering miR-194-5p to up-regulate DARS. Biomed Pharmacother. 2020;128:110323. doi:10.1016/j.biopha.2020.110323
27. Hu G, Wei Y, Kang Y. The multifaceted role of MTDH/AEG-1 in cancer progression. Clin Cancer Res. 2009;15(18):5615–5620. doi:10.1158/1078-0432.CCR-09-0049
28. Wang Z, Wei YB, Gao YL, Yan B, Yang JR, Guo Q. Metadherin in prostate, bladder, and kidney cancer: A systematic review. Mol clin oncol. 2014;2(6):1139–1144. doi:10.3892/mco.2014.392
29. Gutschner T, Hammerle M, Diederichs S. MALAT1 – a paradigm for long noncoding RNA function in cancer. J Mol Med. 2013;91(7):791–801. doi:10.1007/s00109-013-1028-y
30. Serviss JT, Johnsson P, Grander D. An emerging role for long non-coding RNAs in cancer metastasis. Front Genet. 2014;5:234. doi:10.3389/fgene.2014.00234
31. Chen X, Tang FR, Arfuso F, et al. The Emerging Role of Long Non-Coding RNAs in the Metastasis of Hepatocellular Carcinoma. Biomolecules. 2019;10:1. doi:10.3390/biom10010066
32. Ronnau CG, Verhaegh GW, Luna-Velez MV, Schalken JA. Noncoding RNAs as novel biomarkers in prostate cancer. Biomed Res Int. 2014;2014:591703. doi:10.1155/2014/591703
33. Li C, Yang L, Lin C. Long noncoding RNAs in prostate cancer: mechanisms and applications. Mo Cell Oncol. 2014;1(3):e963469.
34. Bolton EM, Tuzova AV, Walsh AL, Lynch T, Perry AS. Noncoding RNAs in prostate cancer: the long and the short of it. Clin Cancer Res. 2014;20(1):35–43. doi:10.1158/1078-0432.CCR-13-1989
35. Luo H, Bu D, Sun L, Fang S, Liu Z, Zhao Y. Identification and function annotation of long intervening noncoding RNAs. Brief Bioinform. 2017;18(5):789–797. doi:10.1093/bib/bbw046
36. Nanni S, Bacci L, Aiello A, et al. Signaling through estrogen receptors modulates long non-coding RNAs in prostate cancer. Mol Cell Endocrinol. 2020;511:110864. doi:10.1016/j.mce.2020.110864
37. Hua JT, Chen S, He HH. Landscape of Noncoding RNA in Prostate Cancer. Trends Gene. 2019;35(11):840–851. doi:10.1016/j.tig.2019.08.004
38. Yin J, Zeng X, Ai Z, Yu M, Wu Y, Li S. Construction and analysis of a lncRNA-miRNA-mRNA network based on competitive endogenous RNA reveal functional lncRNAs in oral cancer. BMC Med Genomics. 2020;13(1):84. doi:10.1186/s12920-020-00741-w
39. Tao L, Yang L, Huang X, Hua F, Yang X. Reconstruction and Analysis of the lncRNA-miRNA-mRNA Network Based on Competitive Endogenous RNA Reveal Functional lncRNAs in Dilated Cardiomyopathy. Front Genet. 2019;10:1149. doi:10.3389/fgene.2019.01149
40. Pan H, Guo C, Pan J, et al. Construction of a Competitive Endogenous RNA Network and Identification of Potential Regulatory Axis in Gastric Cancer. Front Oncol. 2019;9:912. doi:10.3389/fonc.2019.00912
41. Abdollahzadeh R, Daraei A, Mansoori Y, Sepahvand M, Amoli MM, Tavakkoly-Bazzaz J. Competing endogenous RNA (ceRNA) cross talk and language in ceRNA regulatory networks: A new look at hallmarks of breast cancer. J Cell Physiol. 2019;234(7):10080–10100. doi:10.1002/jcp.27941
42. Li M, Qian Z, Ma X, et al. MiR-628-5p decreases the tumorigenicity of epithelial ovarian cancer cells by targeting at FGFR2. Biochem Biophys Res Commun. 2018;495(2):2085–2091. doi:10.1016/j.bbrc.2017.12.049
43. Xie P, Wang Y, Liao Y, et al. MicroRNA-628-5p inhibits cell proliferation in glioma by targeting DDX59. J Cell Biochem. 2019;120(10):17293–17302. doi:10.1002/jcb.28991
44. Zhou L, Jiao X, Peng X, Yao X, Liu L, Zhang L. MicroRNA-628-5p inhibits invasion and migration of human pancreatic ductal adenocarcinoma via suppression of the AKT/NF-kappa B pathway. J Cell Physiol. 2020;2:12.
45. Srivastava A, Goldberger H, Dimtchev A, et al. Circulatory miR-628-5p is downregulated in prostate cancer patients. Tumour Biol. 2014;35(5):4867–4873. doi:10.1007/s13277-014-1638-1
46. Anttila V, Stefansson H, Kallela M, et al. Genome-wide association study of migraine implicates a common susceptibility variant on 8q22.1. Nat Genet. 2010;42(10):869–873.
47. Su ZZ, Kang DC, Chen Y, et al. Identification and cloning of human astrocyte genes displaying elevated expression after infection with HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid subtraction hybridization, RaSH. Oncogene. 2002;21(22):3592–3602. doi:10.1038/sj.onc.1205445
48. Bhatnagar A, Wang Y, Mease RC, et al. AEG-1 promoter-mediated imaging of prostate cancer. Cancer Res. 2014;74(20):5772–5781. doi:10.1158/0008-5472.CAN-14-0018
49. Huggins C. Effect of Orchiectomy and Irradiation on Cancer of the Prostate. Ann Surg. 1942;115(6):1192–1200. doi:10.1097/00000658-194206000-00030
50. Leversha MA, Han J, Asgari Z, et al. Fluorescence in situ hybridization analysis of circulating tumor cells in metastatic prostate cancer. Clin Cancer Res. 2009;15(6):2091–2097. doi:10.1158/1078-0432.CCR-08-2036
51. Senapati D, Kumari S, Heemers HV. Androgen receptor co-regulation in prostate cancer. Asian j Urol. 2020;7(3):219–232. doi:10.1016/j.ajur.2019.09.005
52. Narayanan R. Therapeutic targeting of the androgen receptor (AR) and AR variants in prostate cancer. Asian j Urol. 2020;7(3):271–283. doi:10.1016/j.ajur.2020.03.002
53. Saranyutanon S, Srivastava SK, Pai S, Singh S, Singh AP. Therapies Targeted to Androgen Receptor Signaling Axis in Prostate Cancer: progress, Challenges, and Hope. Cancers. 2019;12(1):1. doi:10.3390/cancers12010051
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.