Back to Journals » OncoTargets and Therapy » Volume 13
Long Non-Coding RNA 691 Regulated PTEN/PI3K/AKT Signaling Pathway in Osteosarcoma Through miRNA-9-5p
Received 14 February 2020
Accepted for publication 18 April 2020
Published 22 May 2020 Volume 2020:13 Pages 4597—4606
DOI https://doi.org/10.2147/OTT.S249827
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Pengju Yao,1 Yangming Ni,1 Changlu Liu2
1Department of Joint Surgery, Jiaozuo People’s Hospital, Henan Province, People’s Republic of China; 2Department of Joint Surgery, The Second Affiliated Hospital of Inner Mongolia Medical University, Hohhot, Inner Mongolia Autonomous Region, People’s Republic of China
Correspondence: Yangming Ni
Department of Joint Surgery, Jiaozuo People’s Hospital Email [email protected]
Background: Large amounts of researches indicate that non-coding RNAs play a crucial role in many malignancies. However, the potential mechanisms of non-coding RNAs involved in osteosarcoma tumorigenesis remain elusive.
Materials and Methods: The expression of long non-protein coding RNA 691 (lncRNA 691) in cell lines and paired osteosarcoma tissues was compared by qRT-PCR assay. Then, we explored the tumor suppressor function of lncRNA 691 with MTS and colony formation assay. Flow cytometry results showed lncRNA 691 can enhance cell apoptosis. Then, we predicted and verified the negative regulation relationship with miRNA and the miRNA’s target gene. Lastly, we revealed the tumorigenesis function of lncRNA-691/miRNA/target gene axis in osteosarcoma.
Results: In our study, we disclosed that lncRNA 691 had low expression levels in osteosarcoma cell lines and tissues. Overexpression of lncRNA 691 could suppress the cell proliferation and induce cell apoptosis in MG-63 cell line. Then, bioinformatics analyses were performed and miR-9-5p was found to negatively regulate the lncRNA 691 expression and promote the osteosarcoma tumorigenesis in vitro. PTEN was predicted as the target gene of miR-9-5p. Luciferase reporter assay and RIP assay demonstrated the regulatory network of lncRNA 691/miR-9-5p/PTEN. We revealed that PTEN was downregulated by the overexpression of miR-9-5p and upregulated by the overexpression of lncRNA 691. At last, the apoptosis-associated protein of the lncRNA 691/miR-9-5p/PTEN/PI3K/AKT was further demonstrated.
Conclusion: LncRNA 691/miR-9-5p could regulate the tumorigenesis by regulating the PTEN/PI3K/AKT signal pathway in osteosarcoma.
Keywords: osteosarcoma, lncRNA 691, miRNA-9-5p, PTEN/PI3K/AKT, apoptosis
Introduction
Osteosarcoma (OS) is the most common type of bone malignant tumor derived from the interstitial cell line.1–3 The rapid growth of OS is due to the direct or indirect formation of tumor bone-like tissue and bone tissue throughout the cartilage stage. The mutations of weight-bearing bone of the lower limb cells which related to the formation of OS can be attributed to the influence of external factors (such as virus).4
Non-coding RNAs (ncRNA) constitute a class of molecules that function as post-transcriptional gene regulators and have been recognized as oncogenes or anti-oncogenes.5 A variety of ncRNAs have been involved in the regulation of multiple processes of tumorigenesis.6 Long non-coding RNA (lncRNA) is a class of non-coding RNA with length more than 200 nt. Studies have shown that a number of lncRNAs were abnormally expressed in osteosarcoma and participated in the regulation of osteosarcoma-related genes.7 MicroRNA (miRNA) is a type of non-coding small included 22–28 nt in length. miRNA can affect cell proliferation, differentiation and apoptosis by binding to its target genes. Salmena et al proposed that competing endogenous RNA (ceRNA) mechanisms that lncRNAs could sponge miRNAs and bind proteins to exert the oncogene or anti-oncogene function.8 For example, DANCR working as a competitive endogenous RNA can promote ROCK1-mediated proliferation via miR-335-5p and miR-1972 in OS.9
lncRNA 691 is a novel lncRNA, which has never been studied in human cancer. Previous studies have found that miRNA-9-5p could promote proliferation and invasion of cervical cancer and renal clear cell carcinoma. Phosphatase and tensin homolog (PTEN) is a tumor suppressor gene associated with the occurrence and development of a variety of malignant tumors.10,11 PTEN is the only anti-tumor protein with dual phosphatase activities and it plays a wide regulatory role in physiological processes such as cell proliferation and apoptosis.7,12,13 Studies have shown that the function of PTEN in apoptosis regulation may be related to PI3K/AKT signaling pathway.14,15
In the current study, we firstly revealed the crucial function of lncRNA-691 in the tumorigenesis of OS regulated by miR-9-5p and PTEN/PI3K/AKT pathway.
Methods
Patients and Tissues
Seventeen osteosarcoma patients’ tumor tissues and adjacent normal tissues were collected in our center during 2015–2018. All the patients were diagnosed as osteosarcoma without radiotherapy or chemotherapy before surgery. The study was undertaken with the approval of the Second Affiliated Hospital of Inner Mongolia Medical University Institutional Ethics Committee Board. All patients have signed the written consent form.
Cell Culture
MG-63, U2OS, 143B human osteosarcoma cell lines and human osteoblast cell line hFOB 1.19 were purchased from American Tissue Culture Collection (Manassas, VA, USA). All the cells have been cultured in DMEM (Gibco, NY, USA) mixed with 10% fetal bovine serum (Gibco, NY, USA) and 100 µg/mL penicillin and 100 µg/mL streptomycin solution (Solarbio, Beijing, China). All cells were incubated at 37 °C in a humidified 5% CO2 atmosphere.
pcDNA empty vector (mock), pcDNA-lncRNA 691 overexpression vector were synthesized by GenePharma (Shanghai, China) and cloned into MG-63 cell line with the recombinant lentiviral ZsGreen Puro vector (Inovogen Tech. Co). MiR-9-5p mimic and NC mimic were purchased from GenePharma (Shanghai, China).
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Assay
The extraction of total RNA was performed with TRIzol reagent (Invitrogen, Carlsbad, CA) and cDNAs were synthesized by PrimeScriptTM RT reagent kit (Tiangen, Beijing, China). The quantitative real-time PCR was performed using an RT-PCR system (TransGen Biotech, Beijing, China) on the CFX96 Touch Real-Time PCR system (Biorad, Hercules, USA). All the procedures were performed following the manufacturer’s protocols. The sequences of the primers were as follows:
lncRNA-691
Forward: GCAGTCTGTGAGCTGGATGT
Reverse:GCAGTCTGTGAGCTGGATGT
miR-9-5p
Forward: AGCTTGCTGCACCTTAGTCT
Reverse: AACGCTTCACGAATTTGCGT
U6
Forward: CTCGCTTCGGCAGCACA
Reverse: AACGCTTCACGAATTTGCGT
PTEN
Forward: ACCACCACAGCTAGAACTT
Reverse:ATCTAGGGCCTTGTGTGCCT
β-Actin
Forward: CCACTGGCATCGTGATGGA
Reverse: CGCT CGGTGAGGATCTTCAT
Cell Proliferation (MTS) Assay
Cell viability was measured by CellTiter 96® AQueous One Solution Cell Proliferation Assay Kit (Promega, USA). Cells were divided into the lncRNA 691 overexpression group and MOCK group. All cells (2×103) were plated in triplet 96-well plates and cultured for 24, 48, 72, and 96 hrs. Then, 20 μL reagent was added into each well and the plates were incubated at 37°C for another 2 hrs. Lastly, absorbance was detected at 490 nm with an SpectraMax iD3 (Molecular Devices, USA).
Colony Formation Assay
Overexpression lncRNA 691 MG-63 cells and NC MG-63 cells (5 × 102 cells/well) were plated into six-well plates and cultured for 2 weeks. After fixation with 4% paraformaldehyde, the colonies were stained with 1% crystal violet in methanol and then counted to evaluate cell proliferation. The experiments were conducted in triplicate.
Cell Apoptosis Analysis by Flow Cytometry
For detecting cell apoptosis level, flow cytometry methods were performed to analyze in accordance with the Annexin V-APC/7-aminoactinomycin D (7-AAD) Apoptosis Detection Kit (KeyGen, China). After cultured cells were seeded, digested, harvested, washed and precooled, we added the Annexin V-APC and 7-AAD into the samples. The samples were incubated at room temperature in a dark place and the apoptosis rate was detected using flow cytometry analysis (BD Accuri™ C6, USA).
Western Blot
Total protein was extracted using RIPA lysis buffer (Solarbio, Beijing, China) with the Protease Inhibitor Cocktail (Sigma) and qualified using BCA detecting kit (Tiangen, Beijing, China). All procedures were performed according to the manufacture’s protocol. Then, equal amounts of proteins were loaded in sodium dodecyl sulfate-polyacrylamide gels and transferred to a polyvinylidene fluoride membrane. After blocked with tris buffered saline-tween with 5% nonfat milk at room temperature (BD Bioscience, USA) for 0.5 hrs, the membranes were incubated with PTEN, t-PI3K, p-PI3K, t-AKT, p-AKT, Caspase3, Bcl-2, BAX and β-actin antibodies (dilution rates of 1:500) (Abcam, Cambridge, MA, USA) at 4 °C overnight. Then, the membranes were incubated with anti-rabbit or anti-mouse secondary antibodies (1:5000) (Solarbio, Beijing, China) after washed. Finally, the signals were detected by SuperSignal West Femto Trial Kit (Thermo Fisher Scientific, USA).
Luciferase Reporter Assay
PGL3 control vector (Promega Corporation, Madison, WI) is used as a firefly luciferase reporter vector. The lncRNA 691 wild-type or mutant and PTEN 3’UTR wild-type or mutant with miR-9-5p binding sequence were cloned into downstream of pGL3-control luciferase reporter vectors (Genewiz, Suzhou, China) in 3’UTR to synthesize lncRNA 691–wt or –mut and PTEN –wt or –mut luciferase reporter vector, respectively. MG-63 cells were co-transfected with 100 ng of the corresponding luciferase reporter vector and miR-9-5p mimics or mimic control. Then, the luciferase activity was detected with the luciferase reporter assay system (Promega Corporation, Madison, WI) and the SpectraMax iD3 (Molecular Devices, USA). Then, the fluorescence intensity with the ratios of firefly luciferase (hLuc) activity/Renilla luciferase (hRluc) activity was calculated (Figures S1and S2).
RNA Immunoprecipitation Assay
The RNA immunoprecipitation (RIP) assay was performed using a Magna RIP™ RNA Binding Protein Immunoprecipitation Kit (Millipore, USA). MG-63 cells transfected with miR-9-5p or miR-NC were lysed by RIP buffer containing magnetic beads conjugated with human anti-Ago2 antibody or IgG antibody as negative control at 4 °C. Then, the immunoprecipitated RNA was washed and extracted with TRIzol and analyzed by qRT-PCR to detect the expression level.
Statistical Analysis
All the statistical analyses were performed in the R software environment (version 3.6.2; http://r-project.org/). Graphs were presented by using GraphPad Prism 7 Software (GraphPad, San Diego, CA). The significance of differences between groups was assessed by Student’s t-test, one-way ANOVAs or χ2 test. p < 0.05 was considered as statistically significant in all statistical analyses.
Results
lncRNA 691 Expression Level Is Low in OS Tissues and Cell Lines
The lncRNA 691 expression level in OS tissues and normal tissues was evaluated by qRT-PCR. We compared the expression of lncRNA 691 in 17 OS patients’ tumor tissues and their paired adjacent tissues. We found that the expression level of lncRNA 691 in OS tissues decreased significantly along with the T stage rise (p < 0.0001) (Figure 1A). Then, we compared the lncRNA 691 expression level in the OS tissues and their adjacent tissues. As the results presented in Figure 1B, lncRNA 691 was decreased in OS tissues compared with adjacent normal tissues (p < 0.001). At last, we compared the lncRNA 691 expression level in normal and OS cell lines. The expression level of lncRNA 691 in hFOB 1.19 cell line was higher than MG-63, U2OS, 143B (p < 0.0001) (Figure 1C), which is consistent with the result of the clinical samples.
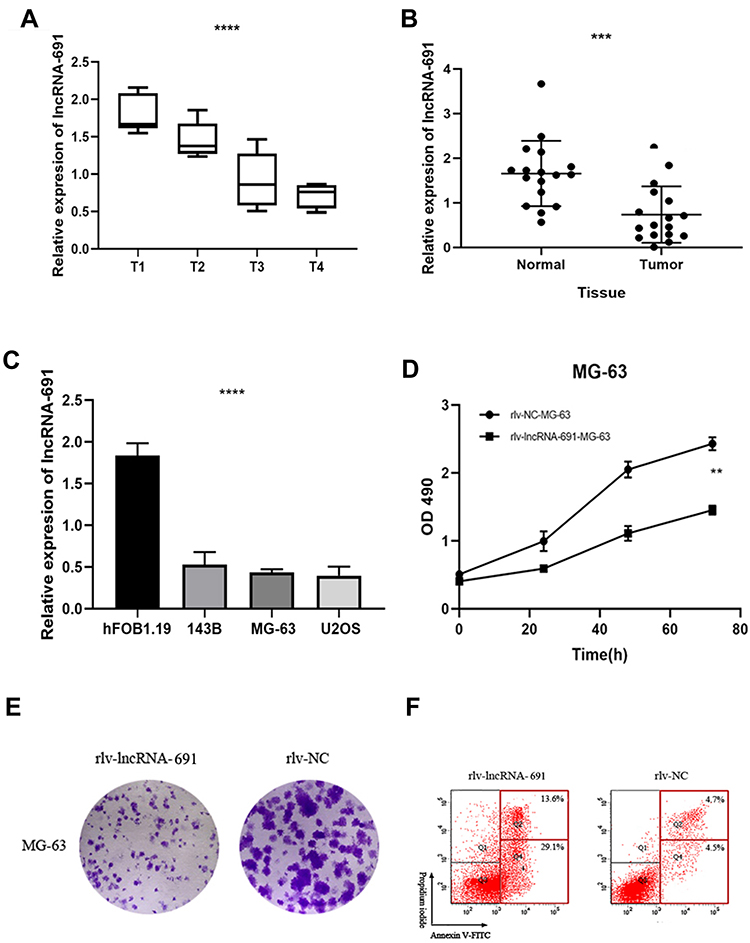 |
Figure 1 (A) The expression of lncRNA 691 in OS tissues was significantly reduced with the T stage rise. (B) The expression of lncRNA 691 in OS tissue was lower than adjacent normal tissue. (C) The expression of lncRNA 691 in hFOB1.19 was significantly higher than OS cell lines. (D) The OD values of overexpression lncRNA 691 group were lower than MOCK group. (E) The colony formation assay results showed that the number of MG-63 cell colonies were decreased significantly in lncRNA 691 overexpression group compared with the NC group. (F) The apoptotic cells percent of lncRNA 691 overexpression group was elevated. ***p < 0.001 or ****p < 0.0001 indicates a significant difference compared with the control group. |
Overexpression lncRNA 691 Could Inhibit Cell Viability, Proliferation and Promotes Apoptosis in OS Cell Lines
To evaluate the cell viability between the overexpression lncRNA 691 group and the MOCK group, we used the MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) assay. The results (Figure 1D) showed that the optical density (OD) values of MOCK group were higher than the lncRNA 691 overexpression group (p = 0.0312), which indicated that lncRNA 691 can suppress OS cell viability. The colony formation assay results (Figure 1E) were consistent with the MTS assay. The number of MG-63 cell colonies was decreased significantly in the lncRNA 691 overexpression group compared with the MOCK group, which demonstrated that upregulated lncRNA 691 expression can suppress OS cell proliferation. We also conducted the flow cytometry to detect the cell apoptosis between these two groups. The percentage of apoptotic cellsof lncRNA 691 overexpression group was reduced by (33.5±4.23)%. This phenomenon demonstrated that lncRNA 691 overexpression can enhance cell apoptosis in MG-63 cell line (Figure 1F).
miR-9-5p Expression Level in OS Cell Lines Is Higher Than Normal Cell Line and miR-9-5p Could Be Downregulated by lncRNA 691
According to the prediction results of DIANA tools (http://carolina.imis.athena-innovation.gr/)16 and starBase v2.0, 42 miRNAs were predicted have the potential ability to bind with lncRNA 691 (Figure S3). Combined with preliminary experiment results, we finally selected miR-9-5p as it show the significant difference. We compared the miR-9-5p level in normal cell line and OS cell lines, overexpression lncRNA 691 MG-63 and MOCK MG-63, respectively. The result (Figure 2A) showed that the expression level of miR-9-5p in hFOB 1.19 was lower than MG-63, U2OS, 143B (p < 0.0001) (Figure 1D) and miR-9-5p is downregulated in lncRNA 691 overexpression MG-63 cell line (Figure 2B).
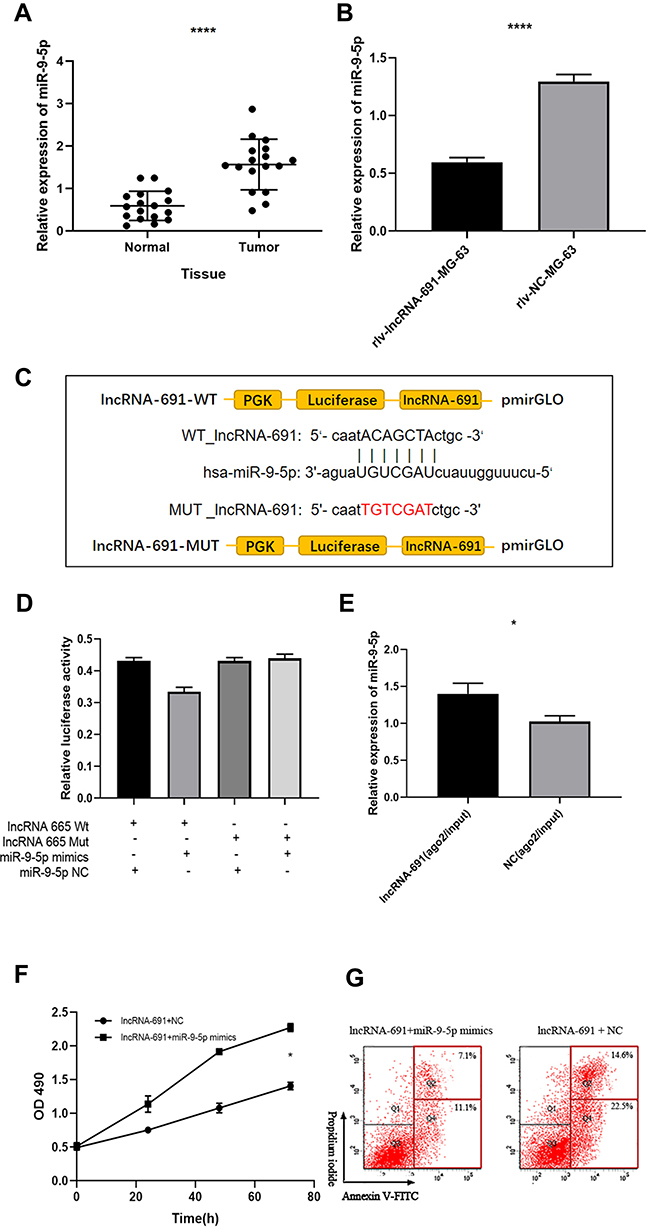 |
Figure 2 (A) The expression of miR-9-5p was significant higher in OS tissue than normal tissue. (B) The expression of miR-9-5p was significantly downregulated by lncRNA 691. (C) Luciferase reporter assay result showed lncRNA 691 can bind to miR-9-5p. (D) The luciferase activity of WT lncRNA 691 reporter vector was significantly reduced by miR-9‐5p mimics, compared with the empty vector and mutant reporter vector. (E) The expression of miR‐9‐5p in the lncRNA 691 overexpression group was remarkale higher than the NC group. (F) The OD values of overexpression lncRNA 691 and miR-9-5p mimics group were higher than overexpression lncRNA 691 and NC group. (G) The overexpression lncRNA 691 cells transfected with miR-9‐5p mimics can inhibit cell apoptosis.*p < 0.05 or ****p < 0.0001 indicates a significant difference compared with the control group. |
lncRNA 691 Could Bind to miR-9-5p: Luciferase Reporter Assay and RIP
To validate the bioinformatics tools’ prediction results, luciferase reporter assay was conducted. Wild-type (WT) and mutant pmirGLO-lncRNA 691 vector was inserted with the binding sites of miR-9-5p (Figure 2C). The data showed that the luciferase activity was significantly reduced in MG-63 cells co-transfected with lncRNA 691 and miR-9-5p (Figure 2D), which demonstrated that miR-9-5p could bind to lncRNA 691. Then, we conducted an anti-AGO2 RIP assay to further validate the regulatory relationship of lncRNA 691 and miR-9-5p. After pulling down, the RNA immunoprecipitates were detected by qRT-PCR. The results confirmed the regulatory relationship as the expression of miR-9-5p is enrichment in the lncRNA 691 overexpression group than the NC group (p=0.0251; Figure 2E).
miR-9-5p Could Offset the Effect Induced by lncRNA 691
To verify the correlation of miR-9-5p and lncRNA 691, we investigated whether the tumor suppressor effect of lncRNA 691 could be offset by miR-9-5p. MiR-9-5p mimics or miR-NC were transfected into the lncRNA 691 stable overexpression MG-63 cell line. MTS assay and flow cytometry were conducted again. As shown in Figure 2F and G, the group transfected with miR-9-5p mimics could enhance cell proliferation and inhibit cell apoptosis, offset the effect induced by lncRNA 691.
PTEN Is the Target Gene Regulated by miR-9-5p
To investigate the underlying mechanism of miR-9-5p biological function in OS, we predicted the mRNA binding sites in DIANA-micro-CDS software and Targetscan Human7.2 and starBase (Figure S4). Among the 31 genes of the intersection of three databases, only PTEN consisted of previous studies and our preliminary experiment result. We predicted that PTEN was a target gene of miR-9-5p with the binging sites (Figure 3A) and qRT-PCT result is in accordance with the hypothesis (Figure 3B). Luciferase reporter assay was conducted to determine whether miR-9-5p could directly bind with PTEN. The results supported our hypothesis as only the fluorescence intensity of Wt-PTEN–3ʹ-UTR reporter vector with miR-9-5p mimics group was significantly reduced in all four groups (p <0.0001; Figure 3C). Then, we compared the PTEN expression level in the miR-9-5p mimics group with the NC mimics group and we found that the PTEN in miR-9-5p mimics group was remarkably reduced (Figure 3D). These findings demonstrated that PTEN expression could be regulated by lncRNA 691/miR-9-5p as ceRNA manner.
 |
Figure 3 (A) Luciferase reporter assay showed the miR-9-5p can bind to PTEN. (B) PTEN expression was low in OS tissues. (C) The luciferase activity of group (pmirGLO‐PTEN–3ʹ‐UTR Wt reporter vector and miR‐9‐5p mimics) was dramatically reduced comparing with the other three groups. (D) The expression of PTEN in miR-9-5p mimics group was significantly lower than miR-NC group. *p < 0.05, **p<0.01, ***p<0.001 or ****p < 0.0001 indicates a significant difference compared with the control group. |
lncRNA 691/miRNA-9-5p Could Regulate the Apoptosis of MG-63 Cell Line by Targeting PTEN/PI3K/AKT Signal Pathway
Then, we detected the expression level of PTEN/PI3K/AKT signal pathway and apoptosis-associated proteins by Western blot in lncRNA 691 overexpression group, MOCK group, miR-9-5p mimics group and miR-NC group. The results revealed that PTEN expression level was prominently higher in lncRNA 691 overexpression cell and miR-9-5p could prominently reduce the level of PTEN. t-PI3K and t-AKT expression showed no difference but the p-PI3K and p-AKT expression were strengthened by miR-9-5p mimics and inhibited by lncRNA 691 overexpression. Earlier results manifested that the overexpression of lncRNA 691 could restrain PTEN/PI3K/AKT signaling pathways, while miR-9-5p mimic could reverse the effect (Figure 4). For the apoptosis-associated proteins, Bax and Cleaved Caspase-3 were upregulated by lncRNA 691 and downregulated by miR-9-5p, while BCL-2 was downregulated by lncRNA 691 and upregulated by miR-9-5p (Figure 4).
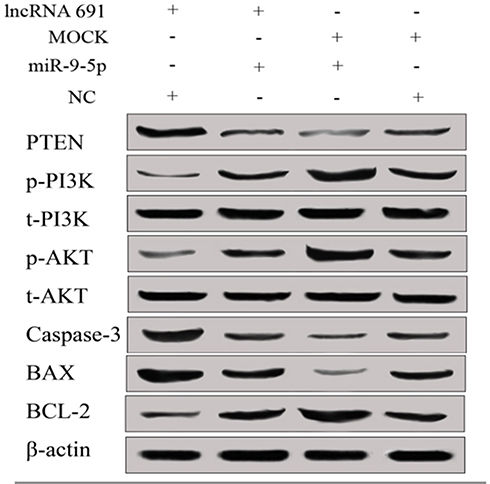 |
Figure 4 Overexpression of lncRNA 691 can upregulate PTEN expression and downregulate p-PI3K and p-AKT expression, miR-9-5p can downregulate PTEN and upregulate p-PI3K and p-AKT expression. t-PI3K and t-AKT expression showed no difference in all four groups. Caspase-3 and BAX were upregulated in overexpression lncRNA 691 group while Bcl-2 was downregulated, miR-9-5p mimics can reverse the effect. |
Discussion
Recent years have witnessed that lncRNAs serve as an oncogene or anti-oncogene and play crucial roles in the development of various cancers including OS.17–19 In addition, studies demonstrated that the lncRNAs could function as competing endogenous RNA (ceRNAs) to regulate target gene expression though competitively bind to miRNA response elements.20 A few lncRNAs involved in the tumorigenesis of osteosarcoma have been identified.21–24 For instance, lncRNA PCAT6 could sponge miR-185-5p to promote tumor progression of osteosarcoma through TGF-βpathway,25 LINC01278 could favor the progression of osteosarcoma via miR-133a-3p/PTHR1 pathway.26 In this study, we focused on the specific tumorigenesis mechanism induced by lncRNA 691 and its ceRNA, miR-9-5p in osteosarcoma.
lncRNA 691, first reported by Navarini in 2014,27 is a novel lncRNA which has never been studied in the human cancer progression. Emerging researches have identified that miR-9-5p was involved in the progression of many diseases, such as kidney fibrosis,28 tuberous sclerosis complex angiomyolipoma29 and different tumors. It could promote malignant biological progression in cervical cancer,30 prostate cancer.31 However, in pancreatic cancer and papillary thyroid cancer, miR-9-5p functions as a tumor suppressor.32,33 It could also promote angiogenesis by target CXCR4.34 PTEN is a classic anti-oncogene in human beings. Its loss or deactivation has been reported in the development of osteosarcoma.35,36 Many non-coding RNAs have been identified, which can bind to PTEN, such as circ-ORC2 and miR-19a37 and miR-29.38 An increasing number of studies have shown that PTEN/PI3K/Akt pathway is involved in the tumorigenesis of osteosarcoma as the loss of function of PTEN may prevent the phosphorylation of PIP2 and decrease the PIP3 and eventually activate the PI3K-Akt pathway to inhibit cell proliferation.39 Meanwhile, PTEN has been recognised as a PD-L1 promoter in OS immunotherapy as PTEN can mediate the PD-L1 expression level.40,41
In this research, we identified that lncRNA 691 expression level is low in OS tissues and cell lines compared with normal tissues and cell line, respectively. What is more, as the T stage improves, the expression level of lncRNA 691 decreases. On the basis of this phenomenon, we conducted a series of experiment in vitro to detect the function of lncRBA 691 in MG-63 cell line. We found that overexpression of lncRNA 691 could inhibit cell viability, restrain cell proliferation ability and strengthen cell apoptosis. These results revealed that lncRNA 691 acts as a tumor suppressor gene in the tumorigenesis of OS. Then, we established the lncRNA–miRNA–mRNA regulatory networks of lncRNA 691/miR-9-5p/PTEN.
A majority of lncRNAs and miRNAs have been demonstrated as ceRNA in regulating the process of tumorigenesis. Some sites of miRNAs’ 3ʹ-UTR can bind to lncRNAs and then compose the RNA-induced silencing complex. This mechanism could explain the broad biological process. In this article, firstly we predicted the binding miRNAs of lncRNA 691 with different bioinformatics prediction tools and then we validated the binding relationship of lncRNA 691 and miR-9-5p with luciferase report assay and RIP immunoprecipitation. Secondly, we explored the biological function of miR-9-5p in MG-63 cell line too. Then, we further predicted the target gene of miR-9-5p and validated it. Finally, we set the relationship between lncRNA 691/miR-9-5p and classic tumor suppressor gene PTEN.
To explore the function of lncRNA 691/miR-9-5p/PTEN axis, we detected the PI3K/AKT pathway proteins and apoptosis-associated proteins. We found that PI3K/AKT pathway was upregulated by overexpression of lncRNA 691 by suppressing cell apoptosis. These phenomena demonstrated our hypothesis that lncRNA 691 can competitively bind to miR-9-5p in a ceRNA model.
Conclusion
LncRNA 691 can regulate tumorigenesis by regulating the PTEN/PI3K/AKT signal pathway through miR-9-5p in osteosarcoma.
Acknowledgments
This work was supported by the Natural Science Foundation of Inner Mongolia Autonomous Region (2016MS08129).
Disclosure
The authors have declared that no competing interest exists.
References
1. Baumhoer D, Zillmer S, Unger K, et al. MicroRNA profiling with correlation to gene expression revealed the oncogenic miR-17-92 cluster to be up-regulated in osteosarcoma. Cancer Genet. 2012;205:212–219. doi:10.1016/j.cancergen.2012.03.001
2. Sahin Y, Altan Z, Arman K, Bozgeyik E, Ozer MK, Arslan A. Inhibition of miR-664a interferes with the migration of osteosarcoma cells via modulation of MEG3. Biochem Biophys Res Commun. 2017;490(3):1100–1105. doi:10.1016/j.bbrc.2017.06.174
3. Song Y-Z, Li J-F. Circular RNA hsa_circ_0001564 regulates osteosarcoma proliferation and apoptosis by acting miRNA sponge. Biochem Biophys Res Commun. 2018;495:2369–2375. doi:10.1016/j.bbrc.2017.12.050
4. Ouyang L, Liu P, Yang S, Ye S, Xu W, Liu X. A three-plasma miRNA signature serves as novel biomarkers for osteosarcoma. Med Oncol. 2013;30:340. doi:10.1007/s12032-012-0340-7
5. Lin Y, Liang X, Yao Y, Xiao H, Shi Y, Yang J. Osthole attenuates APP-induced Alzheimer’s disease through up-regulating miRNA-101a-3p. Life Sci. 2019;225:117–131. doi:10.1016/j.lfs.2019.04.004
6. Alevizos I, Illei GG. MicroRNAs as biomarkers in rheumatic diseases. Nat Rev Rheumatol. 2010;6:391. doi:10.1038/nrrheum.2010.81
7. Andersen GB, Knudsen A, Hager H, Hansen LL, Tost J. mi RNA profiling identifies deregulated mi RNA s associated with osteosarcoma development and time to metastasis in two large cohorts. Mol Oncol. 2018;12:114–131. doi:10.1002/1878-0261.12154
8. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146:353–358. doi:10.1016/j.cell.2011.07.014
9. Wang Y, Zeng X, Wang N, et al. Long noncoding RNA DANCR, working as a competitive endogenous RNA, promotes ROCK1-mediated proliferation and metastasis via decoying of miR-335-5p and miR-1972 in osteosarcoma. Mol Cancer. 2018;17:89.
10. Yan M, Ni J, Song D, Ding M, Huang J. Activation of unfolded protein response protects osteosarcoma cells from cisplatin-induced apoptosis through NF-κB pathway. Int J Clin Exp Pathol. 2015;8:10204.
11. Xia M, Tong J, Ji N, Duan M, Tan Y, Xu J. Tramadol regulates proliferation, migration and invasion via PTEN/PI3K/AKT signaling in lung adenocarcinoma cells. Eur Rev Med Pharmacol Sci. 2016;20:2573–2580.
12. Wang Y, Qin N, Zhao C, et al. The correlation between the methylation of PTEN gene and the apoptosis of osteosarcoma cells mediated by SeHA nanoparticles. Colloids Surf B Biointerfaces. 2019;184:110499. doi:10.1016/j.colsurfb.2019.110499
13. Zhang L, Hu J, Hao M, Bu L. Long noncoding RNA Linc01296 promotes hepatocellular carcinoma development through regulation of the miR-26a/PTEN axis. Biol Chem. 2019.
14. Zheng T, Shi Y, Zhang J, et al. MiR-130a exerts neuroprotective effects against ischemic stroke through PTEN/PI3K/AKT pathway. Biomed Pharmacother. 2019;117:109117. doi:10.1016/j.biopha.2019.109117
15. Huang Z, Xing S, Liu M, et al. MiR-26a-5p enhances cells proliferation, invasion, and apoptosis resistance of fibroblast-like synoviocytes in rheumatoid arthritis by regulating PTEN/PI3K/AKT pathway. Biosci Rep. 2019;39. doi:10.1042/BSR20182192
16. Karagkouni D, Paraskevopoulou MD, Chatzopoulos S, et al. DIANA-TarBase v8: a decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018;46:D239–D245. doi:10.1093/nar/gkx1141
17. Renganathan A, Felley-Bosco E. Long noncoding RNAs in cancer and therapeutic potential. In: Long Non Coding RNA Biology. Springer; 2017:199–222. doi:10.1007/978-981-10-5203-3_7
18. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
19. Zhou S, Yu L, Xiong M, Dai G. LncRNA SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by upregulating Notch2 by sponging miR-195-5p. Biochem Biophys Res Commun. 2018;495(2):1822–1832. doi:10.1016/j.bbrc.2017.12.047
20. Cesana M, Cacchiarelli D, Legnini I, et al. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell. 2011;147(2):358–369. doi:10.1016/j.cell.2011.09.028
21. Yang D, Liu K, Fan L, et al. LncRNA RP11-361F15. 2 promotes osteosarcoma tumorigenesis by inhibiting M2-like polarization of tumor-associated macrophages of CPEB4. Cancer Lett. 2020;473:33–49. doi:10.1016/j.canlet.2019.12.041
22. Yang G, He F, Duan H, Shen J, Dong Q. lncRNA FLVCR-AS1 promotes osteosarcoma growth by targeting miR381-3p/CCND1. Oncol Targets Ther. 2020;13:163. doi:10.2147/OTT.S214813
23. Han Q, Li C, Cao Y, et al. CBX2 is a functional target of miRNA let-7a and acts as a tumor promoter in osteosarcoma. Cancer Med. 2019;8:3981–3991. doi:10.1002/cam4.2320
24. Li S, Wu X, Pei Y, et al. PTHR1 may be involved in progression of osteosarcoma by regulating miR-124-3p-AR-Tgfb1i1, miR-27a-3p-PPARG-Abca1, and miR-103/590-3p-AXIN2 Axes. DNA Cell Biol. 2019;38:1323–1337. doi:10.1089/dna.2019.4880
25. Zhu C, Huang L, Xu F, Li P, Li P, Hu F. LncRNA PCAT6 promotes tumor progression in osteosarcoma via activation of TGF-β pathway by sponging miR-185-5p. Biochem Biophys Res Commun. 2020;521:463–470. doi:10.1016/j.bbrc.2019.10.136
26. Qu Z, Li S. Long noncoding RNA LINC01278 favors the progression of osteosarcoma via modulating miR-133a-3p/PTHR1 signaling. J Cell Physiol. 2020. doi:10.1002/jcp.29582
27. Navarini AA, Simpson MA, Weale M, et al. Genome-wide association study identifies three novel susceptibility loci for severe Acne vulgaris. Nat Commun. 2014;5:1–6. doi:10.1038/ncomms5020
28. Fierro-Fernández M, Miguel V, Márquez-Expósito L, et al. MiR-9-5p protects from kidney fibrosis by metabolic reprogramming. FASEB J. 2020;34:410–431. doi:10.1096/fj.201901599RR
29. Cai Y, Wang W, Guo H, Li H, Xiao Y, Zhang Y. miR-9-5p, miR-124-3p, and miR-132-3p regulate BCL2L11 in tuberous sclerosis complex angiomyolipoma. Lab Invest. 2018;98:856–870. doi:10.1038/s41374-018-0051-6
30. Xie Q, Lin S, Zheng M, Cai Q, Tu Y. Long noncoding RNA NEAT1 promotes the growth of cervical cancer cells via sponging miR-9-5p. Biochem Cell Biol. 2019;97:100–108. doi:10.1139/bcb-2018-0111
31. Chen L, Hu W, Li G, Guo Y, Wan Z, Yu J. Inhibition of miR-9-5p suppresses prostate cancer progress by targeting StarD13. Cell Mol Biol Lett. 2019;24:20. doi:10.1186/s11658-019-0145-1
32. Wang J, Wang B, Ren H, Chen W. miR-9-5p inhibits pancreatic cancer cell proliferation, invasion and glutamine metabolism by targeting GOT1. Biochem Biophys Res Commun. 2019;509:241–248. doi:10.1016/j.bbrc.2018.12.114
33. Guo F, Hou X, Sun Q. MicroRNA‑9‑5p functions as a tumor suppressor in papillary thyroid cancer via targeting BRAF. Oncol Lett. 2018;16:6815–6821. doi:10.3892/ol.2018.9423
34. Yi J, Gao Z-F. MicroRNA-9-5p promotes angiogenesis but inhibits apoptosis and inflammation of high glucose-induced injury in human umbilical vascular endothelial cells by targeting CXCR4. Int J Biol Macromol. 2019;130:1–9. doi:10.1016/j.ijbiomac.2019.02.003
35. Xi Y, Qi Z, Ma J, Chen Y. PTEN loss activates a functional AKT/CXCR4 signaling axis to potentiate tumor growth and lung metastasis in human osteosarcoma cells. Clin Exp Metastasis. 2019:1–13.
36. Zhou J, Xiao X, Wang W, Luo Y. Association between PTEN and clinical-pathological features of osteosarcoma. Biosci Rep. 2019;39(7). doi:10.1042/BSR20190954
37. Li X, Sun X, Xu H, Pan H, Liu Y, He L. Circ_ORC2 enhances the regulatory effect of miR-19a on its target gene PTEN to affect osteosarcoma cell growth. Biochem Biophys Res Commun. 2019;514(4):1172–1178. doi:10.1016/j.bbrc.2019.04.188
38. Liu Q, Geng P, Shi L, Wang Q, Wang P. miR-29 promotes osteosarcoma cell proliferation and migration by targeting PTEN. Oncol Lett. 2019;17:883–890. doi:10.3892/ol.2018.9646
39. Ma Z, Yang J, Yang Y, et al. Rosmarinic acid exerts an anticancer effect on osteosarcoma cells by inhibiting DJ-1 via regulation of the PTEN-PI3K-Akt signaling pathway. Phytomedicine. 2020;68:153186.
40. Liu Z, Wen J, Wu C, et al. MicroRNA-200a induces immunosuppression by promoting PTEN-mediated PD-L1 upregulation in osteosarcoma. Aging (Albany NY). 2020;12:1213–1236. doi:10.18632/aging.102679
41. Wu W, Jing D, Meng Z, et al. FGD1 promotes tumor progression and regulates tumor immune response in osteosarcoma via inhibiting PTEN activity. Theranostics. 2020;10:2859–2871. doi:10.7150/thno.41279
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.