")
Back to Journals » International Journal of General Medicine » Volume 16
Lnc-SNHG5 Promoted Hepatocellular Carcinoma Progression Through the RPS3-NFκB Pathway
Authors Hao J, Liu Y, Guo F, Fu G, Ning J, Ruan X, Zheng X, Gao M
Received 3 October 2023
Accepted for publication 27 November 2023
Published 1 December 2023 Volume 2023:16 Pages 5651—5664
DOI https://doi.org/10.2147/IJGM.S442937
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Jie Hao,1– 3,* Yu Liu,1,* Fengli Guo,1,4 Guiming Fu,1,5 Junya Ning,2,3 Xianhui Ruan,1 Xiangqian Zheng,1 Ming Gao1– 3
1Department of Thyroid and Neck Tumor, Tianjin Medical University Cancer Institute and Hospital, National Clinical Research Center for Cancer, Key Laboratory of Cancer Prevention and Therapy, Tianjin’s Clinical Research Center for Cancer, Tianjin, People’s Republic of China; 2Department of Breast and Thyroid Surgery, Tianjin Union Medical Center of Nankai University, Tianjin, People’s Republic of China; 3Tianjin Key Laboratory of General Surgery in Construction, Tianjin Union Medical Center Nankai University, Tianjin, People’s Republic of China; 4Department of Thyroid and Breast Surgery, Binzhou Medical University Hospital, Binzhou, People’s Republic of China; 5Thyroid-Otolaryngology Department, Sichuan Cancer Hospital & Institute, Sichuan Cancer Center School of Medicine, University of Electronic Science and Technology of China, Chengdu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiangqian Zheng; Ming Gao, Email [email protected]; [email protected]
Background: We planned to explore the underlying mechanism and clinical significance of lnc-SNHG5 and RPS3 in hepatocellular carcinoma in this current study.
Methods: The expression of Lnc-SNHG5 and RPS3 in HCC tissues and several cell lines were affirmed, respectively, using UALCAN, TIMER, TCGA and RT-qPCR assay. Cell proliferation ability was detected by colony formation assay and CCK8 assay. Cell apoptosis was monitored by flow cytometry assay. Next, the RPS3 expression levels and the related proteins in NFκB pathway were examined using Western blot analysis. The role of lnc-SNHG5 and RPS3 in vivo was identified by subcutaneous tumor bearing experiment.
Results: Lnc-SNHG5 was significantly increased in hepatocellular carcinoma tissues and in hepatocellular carcinoma cells. Further investigation showed that up-regulated lnc-SNHG5 promoted cell viability and cell proliferation ability of SMMC-7721 cells by regulating the cell apoptosis, while down-regulation of lnc-SNHG5 revealed opposite results in QGY-7703 cells. In terms of mechanism, we found that lnc-SNHG5 interacted with RPS3. Lnc-SNHG5 regulated the NFκB pathway through RPS3 in vitro and in vivo.
Conclusion: This study suggested that lnc-SNHG5 expression was signally up-regulated in hepatocellular carcinoma, and lnc-SNHG5 promoted the malignant phenotypes in vitro and in vivo via directly regulating RPS3-NFκB pathway. Lnc-SNHG5 might be a target for molecular targeted therapy, a potential and novel diagnostic marker for HCC patients.
Keywords: long non-coding RNA, SNHG5, RPS3, HCC, NFκB pathway
Introduction
As one of malignant tumors, hepatocellular carcinoma (HCC) is a common and fatal tumor., accounting for approximately nine hundred thousand new cases and eight hundred and thirty thousand deaths in 2020.1,2 Almost 75% of all liver cancers were caused by HCC.3 Up to now, increasing therapies have been used to cure HCC, such as chemotherapy, radiotherapy, gene therapy, immunotherapy, liver transplantation.4,5 However, the five years survival rate of HCC patients remains very poor due to the high rate of recurrence and distant metastasis. Hence, an in-depth understanding of the mechanism of hepatocellular carcinoma development will promote the development of treatments.
Long non-coding RNA (lncRNA) is a kind of non-coding RNA with around 200 nucleotides in length that cannot encode proteins.6 Many studies have indicated that lncRNAs play a vital role in regulating various biological functions through the different mechanisms in many cancers.7–9 For example, HBx induced MAFG-AS1 upregulation promoted cell proliferation and migration in HCC cell by regulating the MAFG-E2F1 pathway.10 Yukimoto A et al reported that lncRNA RMRP induced apoptosis in HCC.11 In addition, LncRNA SNHG6 promoted G1/S-phase transition through the miR-204-5p-E2F1 axis in HCC.12 Luo et al reported that quercetin activates the HIF-1α/lncRNA NEAT1/HMGB1 signal pathway and then improves the CI-AKI.13 Recently, it was proved that FAM225A activated the cGAS-STING signal pathway by controlling the CENP-N expression and then promoted cell migration and proliferation in nasopharyngeal carcinoma.14 In addition, a number of SNHGs have been found to play important roles in cancers.15 Xiao et al reported that SNHG5 expression was significantly higher in lung cancer tissues from smokers compared with those from nonsmokers. They also found that SNHG5 overexpression due to nicotine treatment was associated with TRAIL resistance in lung cancer.16 However, the character of lnc-SNHG5 in HCC and its underlying mechanism is still unclear.
NFkB was described in 1986 by Sen and Baltimore as a nuclear transcription factor in B-cells that bound to the B-site of the immunoglobulin k enhancer.17 The nuclear factor kappa B (NFκB) contributes to malignancy through impacting cell senescence, apoptosis, metabolism, stress responses, and tumorigenesis.18,19 In HCC, Leslie et al reported that hepatocyte c-Rel signaling limits genotoxic injury and subsequent HCC burden. Inhibiting c-Rel as an adjuvant therapy increased the effectiveness of DNA damaging agents and reduced HCC growth.20
In this current study, we confirmed that the SNHG5 expression level was obviously up-regulated in HCC, and SNHG5 promoted the malignant biological behaviors in HCC cells and in vivo by directly regulating RPS3-NFκB pathway. This present research may provide a potential target for treating HCC.
Materials and Methods
Samples
30 HCC tissues and the normal liver tissues from patients were collected from January 2019 to January 2022. The current study was conducted with the approval of the Ethics Committee of Tianjin Medical University Cancer Institute and Hospital (Ek2019034) and conducted in terms of the Declaration of Helsinki Principles, and all participants gave their written consent.
Bioinformatic Analysis
Tumor IMmune Estimation Resource (TIMER, https://cistrome.shinyapps.io/timer/) and The University of ALabama at Birmingham CANcer data analysis Portal (UALCAN, https://ualcan.path.uab.edu/index.html) was used to demonstrate the level of lnc-SNHG5 and RPS3. The Kaplan Meier plotter (http://kmplot.com/analysis/) was used to analyze the patient survival. The LnCeVar (http://www.bio-bigdata.net/LnCeVar/) was used to show the different cancer hallmark of lnc-SNHG5 involved. The LNCipedia (https://lncipedia.org/) was used to show the interaction of lnc-SNHG5 and the indicated proteins. The IntaRNA (http://www.bioinf.uni-freiburg.de/Software/) database was used to show the interactive sites of lnc-SNHG5 and RPS3.
Cell Culture and Transfection
THLE-2, HepG2, Huh7, QGY-7703 and SMMC-7721 cells were purchased from the iCell Bioscience Inc. (Shanghai, China), which were cultured in DMEM (Invitrogen, Carlsbad, CA, USA) with 10% FBS (Gibco, Carlsbad, CA, USA), 100 μg/mL streptomycin (Solarbio, Beijing, China), 100 U/mL penicillin (Solarbio, Beijing, China) in 5% CO2 at 37°C. The date of the last STR verification is in July 8, 2021. lnc-SNHG5, shR-SNHG5, shR-RPS3 and the empty vector or the negative control were transferred into cells using Lipofectamine 3000 (Life Technologies, Calif, USA).
RT-qPCR
Total RNAs were extracted from tissues and cells using TRIzol Reagent (Invitrogen). Reverse transcription was carried out using the RT-PCR Quick Master Mix Kit (TOYOBO CO., LTD. Life Science Department OSAKA JAPAN). The resultant cDNA was amplified using a SYBR® Green Realtime PCR Master Mix (TOYOBO CO., LTD. Life Science Department OSAKA JAPAN). The sequences of PCR primers are listed. Lnc-SNHG5-qPCR-S: 5’ CGCTTGGTTAAAACCTGACACT3’; Lnc-SNHG5-qPCR-AS: 5’CCAAGACAATCTGGCCTCTATC3’; RPS3-qPCR-S: 5’CTGGAGTTGAGGTGCGAGTTA3’; RPS3-qPCR-AS:5’ACAGCAGTCAGTTCCCGAATC3’; GAPDH-qPCR-S: 5’GAGTCAACGGATTTGGTCGT3’; GAPDH-qPCR-AS: 5’GACAAGCTTCCCGTTCTCAG3’. The reaction conditions were as follows: 94°C for 4 min, 94°C for 20s, 58°C for 20s, and 72°C for 20s for 40 cycles.
CCK8 Assay
Briefly, 24 hours before transfection, cells were plated into 96-well plates with a density of 2×103 cells in 100 μL medium per well. Between 0–48 h of culture, each well was added with 10 μL CCK8 reagent (Solarbio, Beijing, China) and incubated at 37°C for 1 h. The relative cell proliferation rate was detected using a microplate reader (Thermo Fisher, Ltd., CA, USA) at absorbance of 450 nm.
Colony-Formation Assay
The cells were plated in 12-well petri dishes at a density of 500 cells per dish and cultured in a 5% CO2 incubator at 37°C. Change the medium every 3 days. After 2 weeks, the cells were immobilized with methanol (Beyotime, Shanghai, China) for 20 min and stained with 0.5% crystal violet for 10 min (Beyotime, Shanghai, China) and the colonies formed were counted manually.
Flow Cytometry Assay for Cell Apoptosis
Cell apoptosis was detected by Annexin V-FITC kit (Solarbio, Beijing, China) according to the manufacturer’s protocol and was detected by flow cytometry assay.
Western Blot
Cells were washed in cold 1×PBS and lysed using RIPA (a protease phosphatase inhibitor included) lysis buffer, boiled at 95 °C for 5 min, and centrifuged at 12,000 rpm/min for 10 min. The concentration of protein was determined using the BCA Kit (#CW0014S, CWBIO, China). Supernatants were separated on 10% SDS-PAGE and transferred onto PVDF membranes (Millipore, USA) and blocked with 5% non-fat milk (Solarbio, Beijing, China). Then, protein samples were incubated overnight at 4°C with RPS3 (1:2000, ab140688, Abcam), ACTIN (1:3000, ab ab8227, Abcam), total p65 (1:1000, #8242, Cell Signaling Technology), nuclear p65 (1:2000, ab131100, Abcam). After 1xTBST washing, protein samples were incubated with secondary antibodies (Abcam, MA, USA) for 1 h. Finally, ECL kit (Beyotime, Shanghai, China) was used to assess protein bands.
Tumor Xenograft Model
1×106 SMMC-7721 cells were injected into the right subcutaneous tissue of nude mice for tumor transplantation. The subcutaneous length and width of the tumor were measured every 2 days in vivo transplantation trial. After 25 days, the mice were killed and their weights were measured. The mice were housed under standard laboratory conditions (light cycle, 12 h dark/12 h light; temperature, 22±2°C; humidity, 55±2.5%), and were provided with ad libitum access to food and water. All animal experiments were approved by Tianjin Medical University Cancer Institute and Hospital (LLSP2019-017) and performed in compliance with the Ethical Guidelines for The Study of Experimental Pain in Conscious Animals. Great efforts were made to minimize the number and pain of animals.
Statistical Analysis
GraphPad Prism V (GraphPad Software, Inc.) was used for image editing and SPSS 19.0 (IBM Corp.) Statistical software was used for data analysis. The relative expression of SNHG5 in the HCC tissues and the normal tissues was analyzed by the Wilcoxon signed-rank test. Measurement data were expressed as the mean ± SD and compared using the unpaired t-test. P<0.05 was considered to indicate a statistically significant difference.
Results
Lnc-SNHG5 is Up-Regulated in HCC
In order to explore the potential role of lncRNA SNHG5 in HCC, we used some databases and qPCR to show the level of lnc-SNHG5. TIMER showed that lnc-SNHG5 was up-regulated in many cancers, including cholangiocarcinoma (CHOL), glioblastoma multiforme (GBM), kidney renal papillary cell carcinoma (KIRP), kidney renal clear cell carcinoma (KIRC), colon adenocarcinoma (COAD), liver hepatocellular carcinoma (LIHC), lung squamous cell carcinoma (LUSC) (Figure 1A). In addition, UALCAN data base also show that lnc-SNHG5 was up-regulated in the indicated cancers (Figure 1B). Finally, we individually analyzed the level of lnc-SNHG5 in TCGA and found it is observably enhanced in HCC (p<0.05) (Figure 1C). Results of qPCR demonstrated that lnc-SNHG5 expression was enhanced in tumor tissues with HCC (Figure 1D). To find appropriate cell lines for the research, we evaluated the expression of lnc-SNHG5 by qPCR in THLE-2, HepG2, Huh-7, SMMC-7721 and QGY-7703 cell lines. Lnc-SNHG5 was up-regulated in cancer cell lines and was the highest expression in QGY-7703 among the cancer cell lines (Figure 1E). These data suggest that SNHG5 is obviously up-regulated in HCC. We analyzed the TCGA data of 371 HCC tissues and 50 normal liver tissues. The results showed that lnc-SNHG5 is positively related to tumor grade of HCC patients (Figure 2F), but not the age (Figure 2A), gender (Figure 2B), weight (Figure 2C), histological subtypes (Figure 2D), tumor stage (Figure 2E). More importantly, Kaplan-Meier Plots analysis demonstrated that HCC patients with higher lnc-SNHG5 levels revealed poor overall survival compared with the HCC patients with lower lnc-SNHG5 expression (Figure 2G). GO enrichment analyses showed that the top 10 regulation pathways were mainly enriched in terms of cellular process, biological process and so on (Figure 3A). KEGG enrichment analyses showed that the top 10 pathways were mainly enriched in terms of p53 pathway, cancer pathway and so on (Figure 3B). And the dysregulation of lnc-SNHG5 expression was involved in different cancer hallmark which was analyzed by LnCeVar database (Figure 3C).
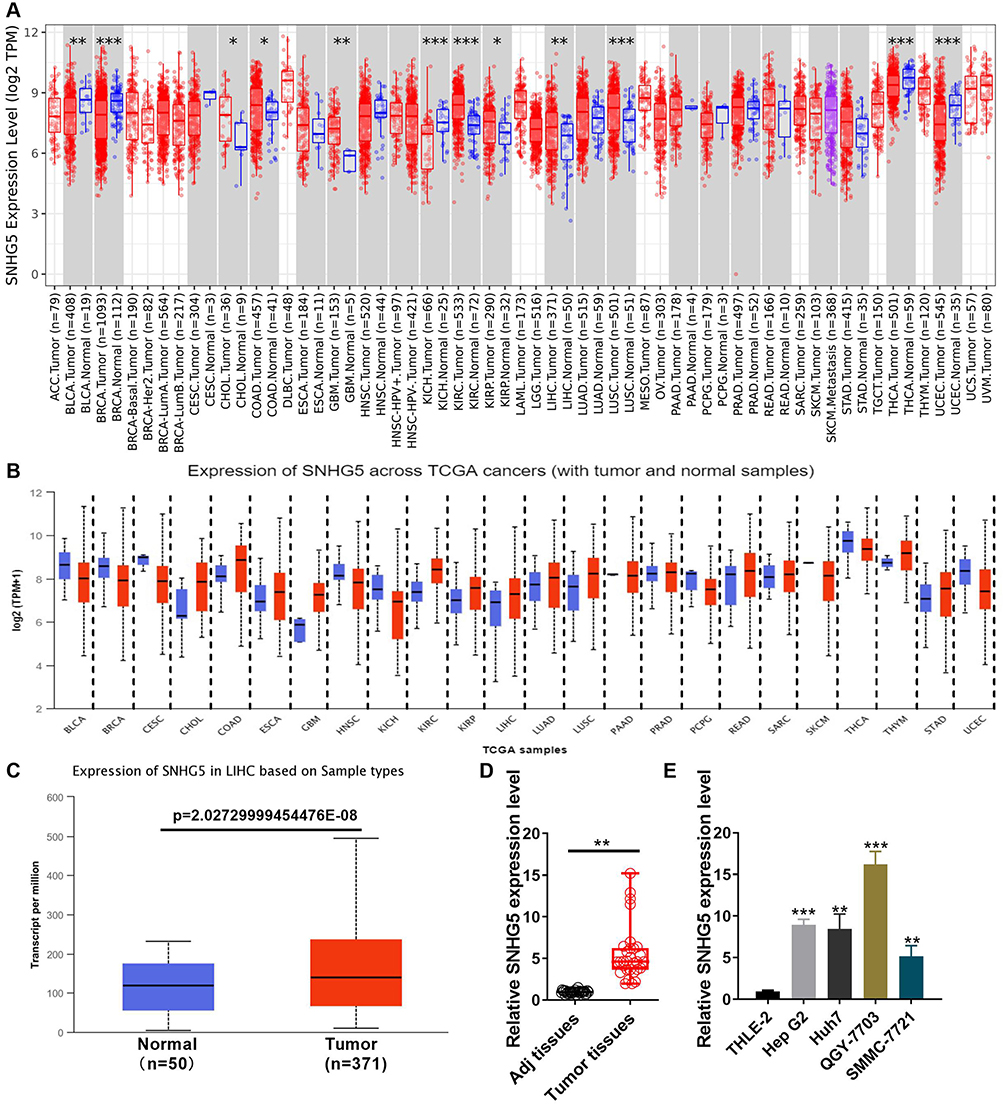 |
Figure 1 Lnc-SNHG5 is up-regulated in HCC. (A) TIMER database showed the expression of lnc-SNHG5 in the indicated cancers. (B) UALCAN showed the expression of lnc-SNHG5 in the indicated cancers. (C) TCGA showed the expression of lnc-SNHG5 in liver cancer. (D) qPCR showed the expression of lnc-SNHG5 in HCC tissues and the normal tissues. (E) qPCR showed the expression of lnc-SNHG5 in the indicated cell lines. Data are presented as the mean±standard deviation. Statistical differences were analyzed using the independent samples t-test. *P<0.05; **P<0.01; ***P<0.001. |
 |
Figure 2 Lnc-SNHG5 is positively related to tumor grade of HCC patients. (A) TCGA showed the expression of lnc-SNHG5 in different ages in HCC patients. (B) TCGA showed the expression of lnc-SNHG5 in Male and Female in HCC patients. (C) TCGA showed the expression of lnc-SNHG5 in different weights in HCC patients. (D) TCGA showed the expression of lnc-SNHG5 in different subtypes in HCC patients. (E) TCGA showed the expression of lnc-SNHG5 in individual cancer stages in HCC patients. (F) TCGA showed the expression of lnc-SNHG5 in different tumor grades in HCC patients. (G) Kaplan-Meier plots showed high lnc-SNHG5 levels predicted the poor prognosis in HCC. *P<0.05. Abbreviation: ns, not significant. |
 |
Figure 3 LnCeVar database showed the different cancer hallmark of lnc-SNHG5 involved. (A) GO analysis showed the different cell process of lnc-SNHG5 involved. (B) KEGG analysis showed the different pathways of lnc-SNHG5 involved. (C) The related cancer hallmarks of lnc-SNHG5. |
Lnc-SNHG5 Promoted Malignant Phenotypes in HCC Cells
Next, the abnormal expression and function of lnc-SNHG5 was investigated in HCC development. Results of qPCR exhibited lnc-SNHG5 group promoted the level of SNHG5 in SMMC-7721 cells, shR-SNHG5 group inhibited the expression level of SNHG5 in QGY-7703 cells (Figure 4A). CCK-8 results exhibited that transfection of lnc-SNHG5 promoted the proliferation ability of SMMC-7721 cells and transfection of shR-SNHG5 inhibited the proliferation ability of QGY-7703 cells (Figure 4B). A colony formation assay showed that lnc-SNHG5 overexpression led to an increase of the colony number in SMMC-7721 cells, whereas lnc-SNHG5 knockdown showed the opposite effect in QGY-7703 cells (Figure 4C). Furthermore, while lnc-SNHG5 inhibited cell apoptosis in SMMC-7721 cells, shR-SNHG5 antagonized such effect in QGY-7703 cells (Figure 4D and E). In conclusion, lnc-SNHG5 acts as a tumor oncogene in HCC.
 |
Figure 4 Lnc-SNHG5 promoted cell proliferation. (A) The efficiency of lnc-SNHG5 or shR-SNHG5 was detected by qPCR assay in SMMC-7721 and QGY-7703 cells. (B) The role of altered lnc-SNHG5 expression on cellular viabilities of SMMC-7721 and QGY-7703 cells were detected by CCK8 assay. (C) Colony formation assay showed the proliferation ability in SMMC-7721 and QGY-7703 cells. (D and E) Flow cytometric assay showed the cell apoptosis ability in SMMC-7721 and QGY-7703 cells. Data are presented as the mean±standard deviation. Statistical differences were analyzed using the independent samples t-test. *P<0.05; ***P<0.001. |
The Interaction Between Lnc-SNHG5 and RPS3
To confirm the role of the lncRNA SNHG5-RPS3 interaction in HCC progression, LNCipedia database was used to predict the interactional genes for SNHG5 and the results showed that RPS3 is a gene of the highest score (Figure 5A). lnc-SNHG5 was positively related to RPS3 in many cancers (Figure 5B). We analyzed the positive expressive correlation of SNHG5 and RPS3 in HCC (Figure 5C). IntaRNA was used to analyze the target sites of lnc-SNHG5 on RPS3 and found that 685–696 site on RPS3 was the indicated sites (Figure 6A). The minimal energy spectrum and the sequence position were also shown (Figure 6B). The results of qPCR corroborated that lnc-SNHG5 promoted the mRNA level of RPS3 after the overexpression with lnc-SNHG5 in SMMC-7721 cells and shR-SNHG5 reduced the RPS3 mRNA level in QGY-7703 cells (Figure 6C). Western blot assay showed overexpression of lnc-SNHG5 further elevated RPS3 protein levels in SMMC-7721 cells, whereas lnc-SNHG5 knockdown reduced RPS3 protein levels in QGY-7703 cells (Figure 6D).
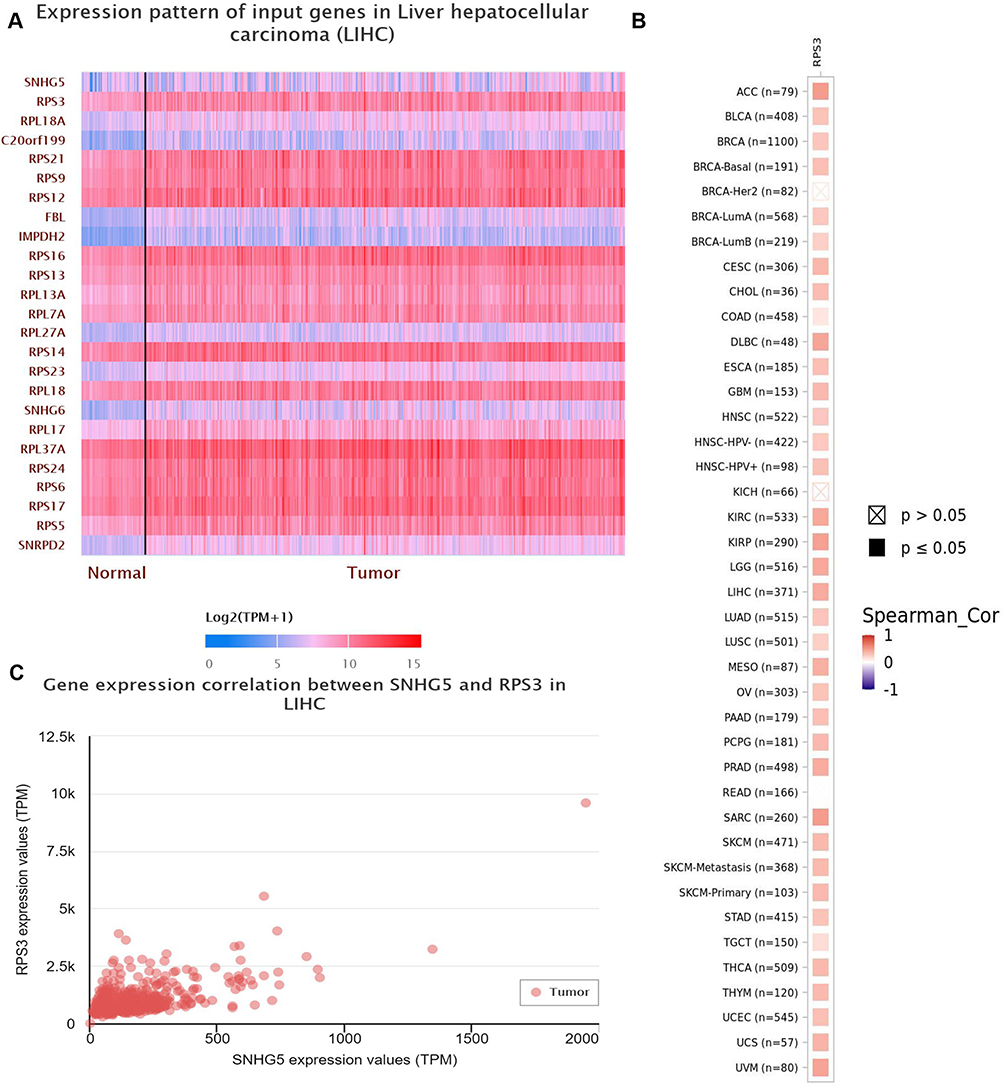 |
Figure 5 The interaction between Lnc-SNHG5 and RPS3. (A and B) LNCipedia showed the interaction of lnc-SNHG5 and the indicated proteins. (C) The positive expressive correlation of SNHG5 and RPS3 in LIHC. |
 |
Figure 6 The interactive sites of lnc-SNHg5 and RPS3. (A) IntaRNA database showed the interactive sites of lnc-SNHG5 and RPS3. (B) The interactive landscape of lnc-SNHG5 and RPS3. (C) qPCR showed the mRNA level of RPS3 in SMMC-7721 and QGY-7703 cells. (D) Western blot showed the protein level of RPS3 in SMMC-7721 and QGY-7703 cells. Data are presented as the mean±standard deviation. Statistical differences were analyzed using the independent samples t-test. *P<0.05; **P<0.01. |
Up-Regulated RPS3 in HCC Was Positively Related to Tumor Grade
TCGA data showed the level of RPS3 in HCC and found it is observably enhanced (p=1.62447832963153E-12) (Figure 7A). The results showed that RPS3 is positively related to tumor grade of HCC patients (Figure 7E), but not the age (Figure 7B), gender (Figure 7C) and weight (Figure 7D). More importantly, Kaplan-Meier survival analysis results showed that HCC patients with higher RPS3 levels revealed poor overall survival compared with the HCC patients with lower RPS3 levels (Figure 7F). In addition, we detected the expression level of RPS3 in HCC tissues and adjacent tissues and found that the RPS3 level was up-regulated in HCC (Figure 7G). In cell lines, we also found that the RPS3 level was increased in the cancer cell lines compared to the THLE-2 cells (Figure 7H).
 |
Figure 7 Up-regulated RPS3 in HCC was positively related to tumor grade. (A) TCGA showed the expression of lnc-SNHG5 in liver cancer tissues and the adjacent normal tissues. (B) TCGA showed the expression of RPS3 in different ages in HCC patients. (C) TCGA showed the expression of RPS3 in Male and Female in HCC patients. (D) TCGA showed the expression of lnc-SNHG5 in different weights in HCC patients. (E) TCGA showed the expression of RPS3 in different tumor grades in HCC patients. (F) Kaplan-Meier survival analysis showed high levels of RPS3 predicted poor prognosis in HCC. (G) IHC showed the RPS3 expression level in HCC tissues. (H) Western blot showed the RPS3 expression level in cell lines. *P<0.05; **P<0.01; ***P<0.001. Abbreviation: ns, not significant. |
SNHG5 Regulated the NFκB Pathway Through RPS3 in vitro and in vivo
In order to determine whether SNHG5 is involved in NFκB pathway, the levels of p65 in NFκB signaling were evaluated using Western blot analysis. lnc-SNHG5 overexpression significantly elevated the phosphorylated p65 (p-p65) expression, but not the total p65 expression (t-p65) in SMMC-7721 cells (Figure 8A). Furthermore, mice injected with lnc-SNHG5 showed increased tumor size, growth rates and tumor weight at day 25 (Figure 8B–D). Finally, we performed IF to detect the expression level of phosphorylated p65 in lnc-SNHG5-mediated tumor in vivo. The results showed that lnc-SNHG5 promoted the protein levels of phosphorylated p65 and knockdown of RPS3 rescued the stimulative effect mediated by lnc-SNHG5 (Figure 8E). Together, these results demonstrate that SNHG5 regulates HCC progression via the RPS3-NF-κB pathway.
 |
Figure 8 SNHG5 regulated the NF-κB pathway through RPS3 in vitro and in vivo. (A) Western blot showed the total and phosphorylated p65 protein level in SMMC-7721 cells. (B–D) lnc-SNHG5 promoted and shR-RPS3 inhibited tumor size, tumor growth rate and tumor weight. (E) IF showed the protein expression of phosphorylated p65 in tumor tissues. Data are presented as the mean±standard deviation. Statistical differences were analyzed using the independent samples t-test. *P<0.05; **P<0.01. Abbreviation: ns, not significant. |
Discussion
New evidence suggests that lncRNAs are involved in the occurrence of tumors and may be used as biomarkers for diagnosis and prognosis in many cancers.21,22 SNHG5 is the small nucleolar RNA host gene 5, located in Chromosome 6 (85677007-85678733), which represents a snoRNA host gene and produces a long non-coding RNA. Abnormal expression of lnc-SNHG5 has been found in various cancers.23 In addition, Yu et al reported lncRNA SNHG5 promoted cell proliferation, migration and invasion by sponging miR-655-3p/FZD4 axis in oral squamous cell carcinoma.24 Recently, Li et al reported the SNHG5 expression was increased in cancer cell lines. Mechanically, SNHG5, as a competitive endogenous RNA (ceRNA) to microRNA-23c (miR-23c), promoted the expression of high mobility substrate box 2 (HMGB2).25 Li et al reported that LncSNHG5 promoted cancer stem cell-like properties and cell proliferation of HCC through the regulation of UPF1 and the Wnt pathway.26 In addition, lnc-SNHG5 promoted the progression of human hepatocellular carcinoma by regulating miR-26a-5p/GSK3β signal pathway.27 Our results are consistent with the above data.25–27 In this study, we found lnc-SNHG5 was up-regulated through the large database including TIMER and TCGA, which is positively related to tumor grade. In addition, lnc-SNHG5 promoted cell proliferation by regulating cell cycle and apoptosis in vitro. Mice injected with lnc-SNHG5 showed increased tumor size, growth rates and tumor weight in vivo.
It has been reported that plenty of ribosomal proteins (RPs) play a vital role in many cancers.28,29 RPS3 is mainly involved in ribosome maturation and initiation of translation and is a part of the small subunit of the ribosome in the 40S.30 RPS3 has various extra-ribosomal functions, including DNA repair, cell metabolism, transcriptional regulation, and cell signaling.31–33 Zhao et al reported that RPS3 contributed to the development of hepatocellular carcinoma through the up-regulation of SIRT1.34 In this study, we found lnc-SNHG5 promoted RPS3 expression at mRNA and protein levels. Mechanically, SNHG5 binds directly to RPS3 through the bioinformatic analysis. Mice injected with shR-RPS3 showed decreased tumor size, growth rates and tumor weight in lnc-SNHG5 treated group in vivo.
The NF-KB families contain five members: cRel, RelA (p65), NF-kB1 (p50), NF-kB2 (p52) and RelB.35,36 Inhibition of NFκB using an IκBα-super-repressor variant boosts the death of premalignant hepatocytes and arrests the development of HCC.37 Papers have shown that RPS3 interacted with the p65 subunit of NFκB in the cytoplasm and the nucleus.38 Blocking the nuclear translocation of RPS3 weakens the RPS3/NF-κB target gene transcription, not affect NF-κB stimuli-triggered p65 nuclear accumulation.39 In this study, we found lnc-SNHG5 could activate the NF-κB pathway including the upregulation of p-p65 through RPS3.
To conclude, our findings constitute the first report to delineate the lnc-SNHG5-RPS3-NFκB axis in HCC. This axis may help elucidate the molecular mechanisms of HCC progression and provide potential therapeutic targets for this deadly disease.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was partially supported by Tianjin Key Medical Discipline (Specialty) Construction Project [TJYXZDXK-058B]; Tianjin Health Research Project [NO. TJWJ2022XK024].
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Bao L, Li P, Zhao H, et al. Pseudogene PLGLA exerts anti-tumor effects on hepatocellular carcinoma through modulating miR-324-3p/GLYATL1 axis. Dig Liver Dis. 2022;54(7):918–926. doi:10.1016/j.dld.2021.10.003
2. Torre LA, Bray F, Siegel RL, et al. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
3. Zhang S, Zhou Y, Wang Y, et al. The mechanistic, diagnostic and therapeutic novel nucleic acids for hepatocellular carcinoma emerging in past score years. Briefings Bioinf. 2021;22(2):1860–1883. doi:10.1093/bib/bbaa023
4. Sun JH, Luo Q, Liu LL, et al. Liver cancer stem cell markers: progression and therapeutic implications. World J Gastroenterol. 2016;22(13):3547–3557. doi:10.3748/wjg.v22.i13.3547
5. Dagogo-Jack I, Shaw AT. Tumour heterogeneity and resistance to cancer therapies. Nat Rev Clin Oncol. 2018;15(2):81–94. doi:10.1038/nrclinonc.2017.166
6. Huang Z, Zhou JK, Peng Y, et al. The role of long noncoding RNAs in hepatocellular carcinoma. Mol Cancer. 2020;19(1):77. doi:10.1186/s12943-020-01188-4
7. Lai Y, Huang H, Abudoureyimu M, et al. Non-coding RNAs: emerging regulators of glucose metabolism in hepatocellular carcinoma. Am J Cancer Res. 2020;10(12):4066–4084.
8. Wang M, Yu F, Chen X, et al. The underlying mechanisms of noncoding RNAs in the chemoresistance of hepatocellular carcinoma. Mol Ther Nucleic Acids. 2020;21:13–27. doi:10.1016/j.omtn.2020.05.011
9. Wong LS, Wong CM. Decoding the roles of long noncoding RNAs in hepatocellular carcinoma. Int J Mol Sci. 2021;22(6):3137. doi:10.3390/ijms22063137
10. Zhang F, Li Y, Gan L, et al. HBx-upregulated MAFG-AS1 promotes cell proliferation and migration of hepatoma cells by enhancing MAFG expression and stabilizing nonmuscle myosin IIA. FASEB J. 2021;35(5):e21529.
11. Yukimoto A, Watanabe T, Sunago K, et al. The long noncoding RNA of RMRP is downregulated by PERK, which induces apoptosis in hepatocellular carcinoma cells. Sci Rep. 2021;11(1):7926. doi:10.1038/s41598-021-86592-6
12. Chen K, Hou Y, Liao R, et al. LncRNA SNHG6 promotes G1/S-phase transition in hepatocellular carcinoma by impairing miR-204-5p-mediated inhibition of E2F1. Oncogene. 2021;40(18):3217–3230. doi:10.1038/s41388-021-01671-2
13. Luo M, Liu Z, Hu Z, et al. Quercetin improves contrast-induced acute kidney injury through the HIF-1α/lncRNA NEAT1/HMGB1 pathway. Pharm Biol. 2022;60(1):889–898. doi:10.1080/13880209.2022.2058558
14. Han JB, Wang Y, Yang R, et al. LncRNA FAM225A activates the cGAS-STING signaling pathway by combining FUS to promote CENP-N expression and regulates the progression of nasopharyngeal carcinoma. Pathol Res Pract. 2022;236:154005. doi:10.1016/j.prp.2022.154005
15. Qin Y, Sun W, Wang Z, et al. Long Non-Coding small nucleolar RNA host genes (SNHGs) in endocrine-related cancers. Onco Targets Ther. 2020;13:7699–7717. doi:10.2147/OTT.S267140
16. Xiao X, Xu J, Sheng X, Wang C, Dong J, Shi X. Tobacco nicotine promotes TRAIL resistance in lung cancer through SNHG5. Exp Ther Med. 2023;25(3):131. doi:10.3892/etm.2023.11830
17. Sen R, Baltimore D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell. 1986;46(5):705–716. doi:10.1016/0092-8674(86)90346-6
18. Macià A, Vaquero M, Gou-Fàbregas M, et al. Sprouty1 induces a senescence-associated secretory phenotype by regulating NFκB activity: implications for tumorigenesis. Cell Death Differ. 2014;21(2):333–343. doi:10.1038/cdd.2013.161
19. Yang G, Wang Y, Feng J, et al. Aspirin suppresses the abnormal lipid metabolism in liver cancer cells via disrupting an NFκB-ACSL1 signaling. Biochem Biophys Res Commun. 2017;486(3):827–832. doi:10.1016/j.bbrc.2017.03.139
20. Leslie J, Hunter JE, Collins A, et al. c-Rel-dependent Chk2 signaling regulates the DNA damage response limiting hepatocarcinogenesis. Hepatology. 2023;78(4):1050–1063. doi:10.1002/hep.32781
21. Cao J, Wu L, Lei X, et al. Long non-coding RNA-based signature for predicting prognosis of hepatocellular carcinoma. Bioengineered. 2021;12(1):673–681. doi:10.1080/21655979.2021.1878763
22. Ding X, Xu X, He XF, et al. Muscleblind-like 1 antisense RNA 1 inhibits cell proliferation, invasion, and migration of prostate cancer by sponging miR-181a-5p and regulating PTEN/PI3K/AKT/mTOR signaling. Bioengineered. 2021;12(1):803–814. doi:10.1080/21655979.2021.1890383
23. Li YH, Hu YQ, Wang SC, et al. LncRNA SNHG5: a new budding star in human cancers. Gene. 2020;749:144724. doi:10.1016/j.gene.2020.144724
24. Yu L, Huo L, Shao X, et al. lncRNA SNHG5 promotes cell proliferation, migration and invasion in oral squamous cell carcinoma by sponging miR-655-3p/FZD4 axis. Oncol Lett. 2020;20(6):310. doi:10.3892/ol.2020.12173
25. Li W, Lu Y, Wu Y, et al. SNHG5 functions as competitive RNA with miR-23c to regulate HMGB2 expression in hepatocellular carcinoma. Am J Transl Res. 2020;12(5):2192–2200.
26. Li Y, Hu J, Guo D, et al. LncRNA SNHG5 promotes the proliferation and cancer stem cell-like properties of HCC by regulating UPF1 and Wnt-signaling pathway. Cancer Gene Therapy. 2022;29(10):1373–1383. doi:10.1038/s41417-022-00456-3
27. Li Y, Guo D, Zhao Y, et al. Long non-coding RNA SNHG5 promotes human hepatocellular carcinoma progression by regulating miR-26a-5p/GSK3β signal pathway. Cell Death Dis. 2018;9(9):888. doi:10.1038/s41419-018-0882-5
28. Ceci M, Fazi F, Romano N. The role of RNA-binding and ribosomal proteins as specific RNA translation regulators in cellular differentiation and carcinogenesis. Biochim Biophys Acta Mol Basis Dis. 2021;1867(4):166046. doi:10.1016/j.bbadis.2020.166046
29. Harold CM, Buhagiar AF, Cheng Y, et al. Ribosomal RNA transcription regulation in breast cancer. Genes. 2021;12(4):502. doi:10.3390/genes12040502
30. Schäfer T, Maco B, Petfalski E, et al. Hrr25-dependent phosphorylation state regulates organization of the pre-40S subunit. Nature. 2006;441(7093):651–655. doi:10.1038/nature04840
31. Kim HD, Kim TS, Kim J. Aberrant ribosome biogenesis activates c-Myc and ASK1 pathways resulting in p53-dependent G1 arrest. Oncogene. 2011;30(30):3317–3327. doi:10.1038/onc.2011.47
32. Kim Y, Kim HD, Kim J. Cytoplasmic ribosomal protein S3 (rpS3) plays a pivotal role in mitochondrial DNA damage surveillance. Biochim Biophys Acta. 2013;1833(12):2943–2952. doi:10.1016/j.bbamcr.2013.07.015
33. Alam E, Maaliki L, Nasr Z. Ribosomal protein S3 selectively affects colon cancer growth by modulating the levels of p53 and lactate dehydrogenase. Mol Biol Rep. 2020;47(8):6083–6090. doi:10.1007/s11033-020-05683-1
34. Zhao L, Cao J, Hu K, et al. RNA-binding protein RPS3 contributes to hepatocarcinogenesis by post-transcriptionally up-regulating SIRT1. Nucleic Acids Res. 2019;47(4):2011–2028. doi:10.1093/nar/gky1209
35. Ghosh S, May MJ, Kopp EB. NF-kappa B and Rel proteins: evolutionarily conserved mediators of immune responses. Annu Rev Immunol. 1998;16(1):225–260. doi:10.1146/annurev.immunol.16.1.225
36. Taniguchi K, Karin M. NF-κB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol. 2018;18(5):309–324. doi:10.1038/nri.2017.142
37. Pikarsky E, Porat RM, Stein I, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004;431(7007):461–466. doi:10.1038/nature02924
38. Wan F, Anderson DE, Barnitz RA, et al. Ribosomal protein S3: a KH domain subunit in NF-kappaB complexes that mediates selective gene regulation. Cell. 2007;131(5):927–939. doi:10.1016/j.cell.2007.10.009
39. Hodgson A, Wier EM, Fu K, et al. Metalloprotease NleC suppresses host NF-κB/inflammatory responses by cleaving p65 and interfering with the p65/RPS3 interaction. PLoS Pathog. 2015;11(3):e1004705. doi:10.1371/journal.ppat.1004705
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.