Back to Journals » OncoTargets and Therapy » Volume 13
LINC00461 Overexpression Can Induce Docetaxel Resistance in Breast Cancer by Interacting with miR-411-5p
Authors Zhang C, Wang J, Zhang J, Qu H, Tang X
Received 30 January 2020
Accepted for publication 14 May 2020
Published 12 June 2020 Volume 2020:13 Pages 5551—5562
DOI https://doi.org/10.2147/OTT.S247776
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Carlos E Vigil
Cheng Zhang,1 Jizhao Wang,1 Jiawei Zhang,1 Hangying Qu,1 Xiaojiang Tang2
1Department of Thoracic Surgery, First Affiliated Hospital, Xi’an Jiaotong University, Xi’an 710061, Shaanxi Province, People’s Republic of China; 2Department of Breast Surgery, First Affiliated Hospital, Xi’an Jiaotong University, Xi’an 710061, Shaanxi Province, People’s Republic of China
Correspondence: Xiaojiang Tang Email [email protected]
Purpose: Breast cancer (BC) is the most common malignant cancer in women worldwide. Recently, long non-coding RNAs (LncRNAs) have been reported to have essential roles in BC tumorigenesis.
Patients and Methods: Tumor and adjacent non-tumor tissue samples were collected from patients with BC (n = 168) for comparison of LncRNA and miRNA expression levels. Patient clinical, demographic, and molecular data were analyzed by univariate and multivariate methods to identify factors associated with patient survival, and a nomogram was generated using significant risk/protective factors. Further, analyses of LINC00461 and miR-411-5p expression and function were conducted in BC and normal breast epithelial cell lines, by quantitative RT-PCR, cell proliferation, wound-healing, RNA pull-down, RNA immunoprecipitation, and luciferase assays. Docetaxel (DTX)-resistant BC cell lines were also generated and used to assess the roles of LINC00461 and miR-411-5p in drug resistance.
Results: LINC00461 was up-regulated in BC tissues relative to adjacent non-tumor samples, and expression of LINC00461 was correlated with poor patient prognosis. LINC00461 knockdown could inhibit proliferation of BC cells in vitro. Further, LINC00461 expression was higher in DTX-resistant than in non-resistant BC cell lines. Our data support a role for LINC00461 as a competitive endogenous RNA sponge involved in regulation of miR-411-5p expression in BC. miR-411-5p was down-regulated in both BC tissues and cell lines, with levels negatively correlated with those of LINC00461. Moreover, miR-411-5p was down-regulated in DTX-resistant BC cell lines compared with non-resistant cell lines.
Conclusion: In conclusion, LINC00461 promotes proliferation, migration, and DTX-resistance in BC by acting as a sponge for miR-411-5p. This process represents a potential therapeutic target for patients with BC.
Keywords: lncRNA, miRNA, proliferation, migration, ceRNA
Introduction
Breast cancer (BC) is the most common malignant cancer in women and a leading cause of cancer-related death in affected individuals, which accounts for 11.6% of all cancers and 6.6% of all cancer death in the world.1–3 There were an estimated 232,670 newly diagnosed cases and 40,000 deaths caused by BC in the United States in 2014,4 with an increase in diagnosed cases to approximately 1.7 million in 2016.5 In recent decades, early diagnosis and treatment regimens for BC have advanced greatly, assisted by the advent of targeted pharmacological treatments, sophisticated surgical resection techniques, and advanced chemotherapy.6 Nevertheless, patient survival and prognosis remain poor, due to high rates of recurrence and drug resistance,7 providing impetus for further efforts to discover new diagnostic methods and therapeutic approaches, as well as to investigate the molecular mechanisms underlying BC.
Long non-coding RNAs (lncRNAs) are a class of RNA transcripts of >200 nucleotides that do not encode proteins.8,9 Research has increasingly focused on the various functions of lncRNAs in development, including in RNA processing, and transcriptional regulation of gene expression.10,11 Similar to proteins, the subcellular localization of lncRNAs is crucial to their function.12,13 Many lncRNAs are regarded as modulators of nuclear functions. In particular, lncRNAs regulate the cellular behavior of the central dogma (ie, DNA → RNA → protein) through diverse mechanisms, such as epigenetic alterations, protein–lncRNA interactions, lncRNA-miRNA/DNA interactions, and genetic variation.14
Some studies have also focused on potential functions of lncRNAs in disease etiology.15–17 Analyses of normal and tumor cells indicate that expression of some lncRNAs is altered in various forms of cancer,18 with lncRNAs, such as HOTAIR, identified as potential biomarkers in cancer diagnosis and prognosis.19
The lncRNA, LINC00461, promotes cell proliferation in glioma, hepatocellular carcinoma, and multiple myeloma.20–22 LINC00461 may act as an oncogene in various types of tumors. However, the function of LINC00461 in BC has yet to be determined. In our study, we found that LINC00461 was significantly up-regulated in BC compared with normal tissues and that its expression was correlated with poor patient prognosis. Further, LINC00461 knockdown could inhibit BC proliferation in vitro. Moreover, expression of LINC00461 in docetaxel (DTX)-resistant cell lines was higher than that in non-resistant BC cell lines. Importantly, knockdown of LINC00461 in DTX-resistant cell lines could reverse DTX resistance. Mechanistically, our data show that LINC00461 acts as a sponge for miR-411-5p, thereby promoting BC cell proliferation.
Patients and Methods
Patients and Samples
BC and adjacent non-cancerous tissue samples were collected from 168 patients with BC at The First Affiliated Hospital of Xi’an Jiao Tong University by surgical resection. All tissue specimens were examined by expert pathologists and stored in liquid nitrogen. No patients had received chemotherapy, radiotherapy, or other treatments at the time of sample collection. This research was approved by the Ethics Committee of The First Affiliated Hospital of Xi’an Jiao Tong University. All patients provided the written informed consent according to the Declaration of Helsinki.
Cell Lines
The normal human mammary epithelial cell line, MCF-10A, and the human BC cell lines, MCF-7 and MDA-MB-231, were purchased from the American Type Culture Collection. Human DTX-resistant BC cell lines, MCF-7-DTX and MDA-MB-231-DTX, were obtained by culturing MCF-7 and MDA-MB-231 cells with elevating doses of DTX for 10 months. All cells were cultured in Dulbecco’s modified Eagle’s medium (Hyclone) supplemented with 10% fetal serum (Logan, UT, USA). The human DTX-resistant BC cell lines, MCF-7-DTX and MDA-MB-231-DTX were obtained by culturing MCF-7 and MDA-MB-231 cells with elevating doses of DTX for 10 months. The wild-type cells were firstly cultured in the complete medium with 200 nM DTX for 1 month, followed by escalation to 400nM, 800nM, 1.5µM, 3µM, 5µM, 7µM, 9µM, 10µM doses until the resistance stabilized. The DTX-resistant cells were characterized by Cell Counting Kit-8 assays for the inhibitory concentration 50.
Vectors and Cell Transfection
Recombinant lentivirus vectors for LINC00461 overexpression and knockdown, as well as negative controls, were obtained from Genechem Biotech (Shanghai, China). Cells (5 × 105) were seeded in 6-well plates and infected with lentivirus using 10 μg/mL Polybrene. Then, cells were grown in media supplemented with 2 μg/mL puromycin for 1 week to select for infected cells. Stable overexpression and knockdown cell lines were identified by qRT-PCR. The miR-411-5p mimics, inhabitor, and negative controls were designed and synthesized by GenePharm Biotech (Shanghai, China) and transfected into cells seeded in 12-well plates using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, lnc.).
RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from BC cells using RNAiso Plus (TaKaRa Biotechnology, Dalian, China), following the manufacturer’s instructions. Then, a Reverse Transcription Kit (TaKaRa Biotechnology, Dalian, China) was used to synthesize cDNA. qRT-PCR was performed using a SYBR Green PCR Kit (TaKaRa Biotechnology, Dalian, China). GAPDH and U6 were used as internal controls for expression quantitation by the 2−ΔΔCt method. Primer sequences were as follows: LINC00461, 5′‐GCGTGGACTACTCTGATG‐3′ (forward) and 5′‐CCAAGTGCTTACTGTCT‐3′ (reverse); miR-411-5p, 5′-GCGACTGACGTCTTTTTGCGGTCTGG-3ʹ (forward) and 5′-CAGA ACAGTGTCGTGACAGTGACGAT-3′ (reverse); U6, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and 5ʹ-CGCTTCACGAATTTGCGTGTCAT-3ʹ (reverse); and GAPDH, 5ʹ-TATGATGATATCAAGAGGGTAGT-3ʹ (forward) and 5ʹ-TGTATCCAAACTCATTGTCATAC-3′ (reverse).
Cell Proliferation Assay
The proliferation capacity of cells was assessed using a Cell Counting Kit-8 (CCK-8, Beyotime Institute of Biotechnology, Shanghai, China) assay, following the manufacturer’s instructions. Briefly, 1 × 103 cells were seeded in each well of 96-well plates and cultured in complete medium at 37°C. Proliferation capacity was plotted by measuring absorbance at 450 nm after incubation with 100 µL complete medium containing 10% CCK-8 in each well for 2 h. Experiments were repeated three times.
Wound-Healing Assay
Cells were infected with LINC00461 overexpression, miR-411-5p overexpression, or corresponding knockdown, lentiviral particles. After 2–3 days, levels of green fluorescent protein in the cells were observed under a fluorescence microscope (a fluorescence rate of 50–80% was expected). Cells were collected when cell density reached 80%. After digestion with trypsin, cells were resuspended in complete culture medium, and the cell density was fixed at 50,000/well. Cells were then cultured in an incubator at 37°C and 5% CO2. When the cells were confluent, using a pipette tip to creating an area free from adherent cells by scraping them off. At 0 and 24 h after inoculation, cells were analyzed using a Celigo scanner (Nexcelom, USA) to determine the migration area.
RNA Immunoprecipitation (RIP) Assay
RIP assays were performed using an EZMagna RIP RNA-binding protein immunoprecipitation kit (Millipore, USA), according to the manufacturer’s instructions. After cells were lysed, lysates were incubated in RIP buffer containing magnetic beads conjugated with Ago2 antibodies (Millipore) or negative control IgG. Then, the samples were incubated at 4°C for 2 h. Subsequently, co-precipitated RNA was isolated using proteinase K and detected by PCR.
RNA Pull-Down Assay
Biotinylated LINC00461 and a control probe were obtained from Genechem Biotech (Shanghai, China). Cell lysates were incubated with LINC00461 or the control probe and streptavidin-coupled Dynabeads (Invitrogen) to generate probe-bound Dynabeads. After washing using wash buffer (Invitrogen), RNA complexes were isolated and purified using lysis buffer and proteinase K. Purified RNA was analyzed by qRT-PCR.
Luciferase Reporter Assay
The online software, starBase 3.0, was used to predict miRNA binding sites in LINC00461. Then, sequences including wild-type (WT) or mutant (MUT) LINC00461 binding sites were cloned into a luciferase reporter vector (Promega, Madison, WI, USA). Cells were plated in 24-well plates and transfected with WT-LINC00461 or MUT-LINC00461 and miR-411-5p mimics using Lipofectamine 2000. After 48 h of transfection, the dual-luciferase reporter assay system (Promega, Madison, WI) was used to evaluate luciferase activity, based on the manufacturer’s instructions. Experiments were performed independently in triplicate.
Statistical Analysis
All statistical analyses were performed in GraphPad (GraphPad Prism version 5.0, San Diego, USA) and SPSS (version 17.0; SPSS, Inc., Chicago, IL, USA) software. A paired Student’s t-test was conducted to analyze LINC00461 in BC and paired adjacent non-cancerous tissue samples. Correlation analysis was conducted to analyze the expression of LINC00461 and miR-411-5p. Survival analysis was conducted to analyze the association between overall survival and LINC00461 and miR-411-5p expression, tumor stage, and estrogen receptor (ER) and progesterone receptor (PR) positive-status. P < 0.05 was considered statistically significant.
Results
LINC00461 Is Up-Regulated in BC
We found that LINC00461 was expressed at high levels in 168 BC tissues (Figure 1A), while miR-411-5p was down-regulated (Figure 1B), relative to adjacent non-tumor tissue samples. Correlation analysis showed that LINC00461 expression was negatively associated with levels of miR-411-5p and miR-491 (Figure 1C). In vitro experiments demonstrated high expression of LINC00461 in the MDA-MB-231 and MCF-7 BC cell lines (Figure 1D), while levels of miR-411-5p were low (Figure 1E); however, no significant miR-491 expression was detected (Figure 1F), relative to levels in the normal human mammary epithelial cell line, MCF-10A. Further, evaluation of LINC00461 subcellular localization demonstrated that it is primarily localized to the cytoplasm (Figure 1G and H).
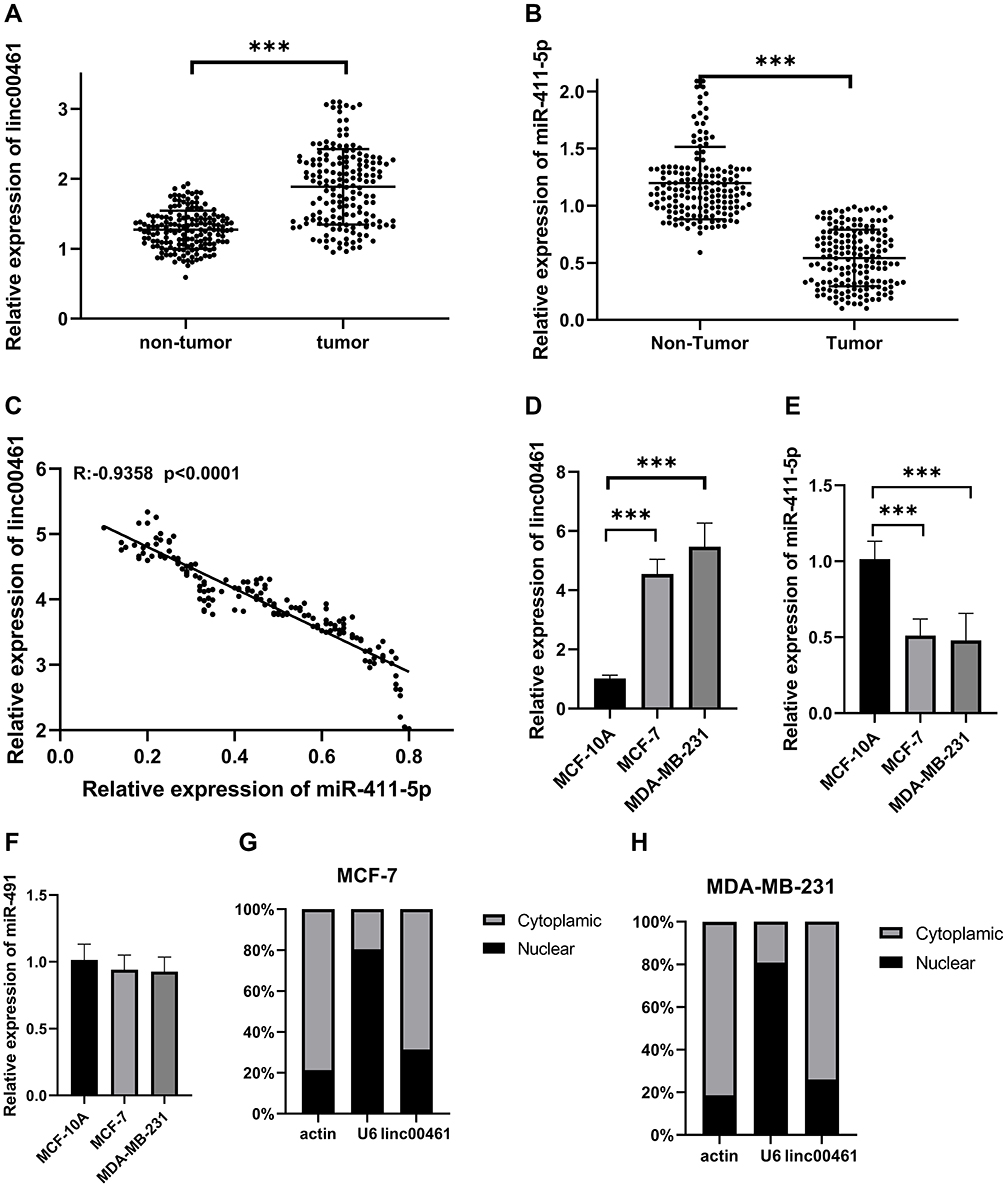 |
Figure 1 (A) LINC00461 expression in 168 paired BC and adjacent non-tumor tissues by RT-qPCR. (B) miR-411-5p expression in 168 paired BC and adjacent non-tumor tissues by qRT-PCR. (C) Correlation between LINC0041 and miR-411-5p. (D) Expression of LINC00461 in MCF-7 and MDA-MB-231 cell lines. (E) Expression of miR-411-5p in MCF-7 and MDA-MB-231 cell lines. (F) Expression of miR-491 in MCF-7 and MDA-MB-231 cell lines. (G) Subcellular localization of LINC00461 in MCF-7 cell lines. (H) Subcellular localization of LINC00461 in MDA-MB-231 cell lines. Data represent the mean ± SD. ***P <0.001. The experiments were independently repeated at least three times. |
Association of LINC00461 and microRNA Expression with Survival of Patients with BC
In this research, we collected 168 BC tissues and the clinical characteristics are shown in Table 1. As shown in Figure 2A–C and Supplemental Fig A and B, LINC00461 and miR-411-5p expression, tumor stage, and estrogen receptor (ER) and progesterone receptor (PR) positive-status, were associated with overall survival in patients with BC. Therefore, we selected M stage, LINC00461, ER+, PR+, and miR-411-5p for inclusion in a nomogram for prediction of 3- and 5-year survival rates for patients with BC. The results showed that M stage was the most important factor influencing survival rate, followed by LINC00461 expression (Figure 2D). Calibration curves for 3-year (Figure 2E) and 5-year (Figure 2F) overall survival prediction and the C-index (0.796 ± 0.061) suggested that the nomogram could precisely and accurately predict survival rates.
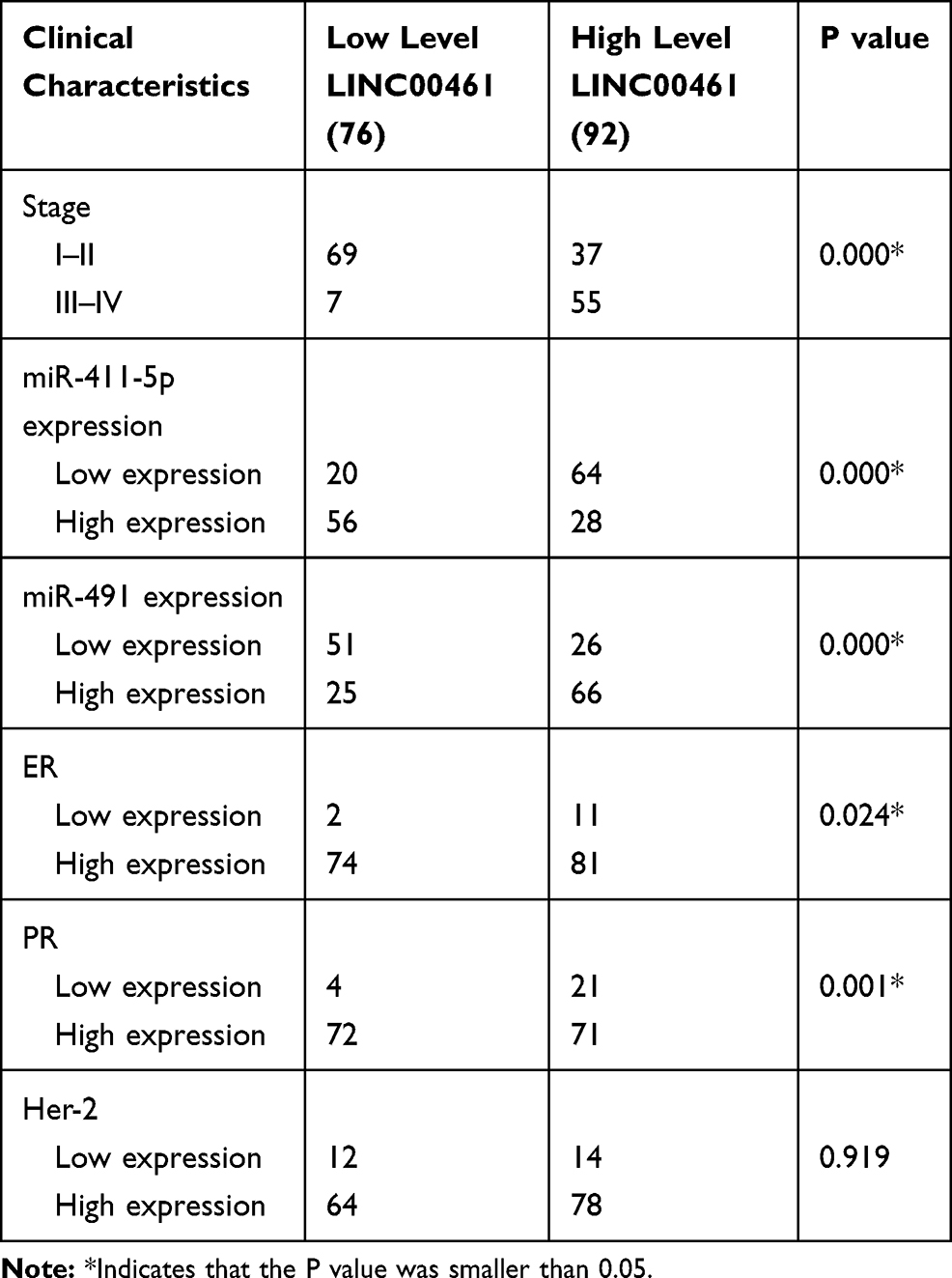 |
Table 1 Clinical Characteristics of Low and High Expression LINC00461 Breast Cancer Patients |
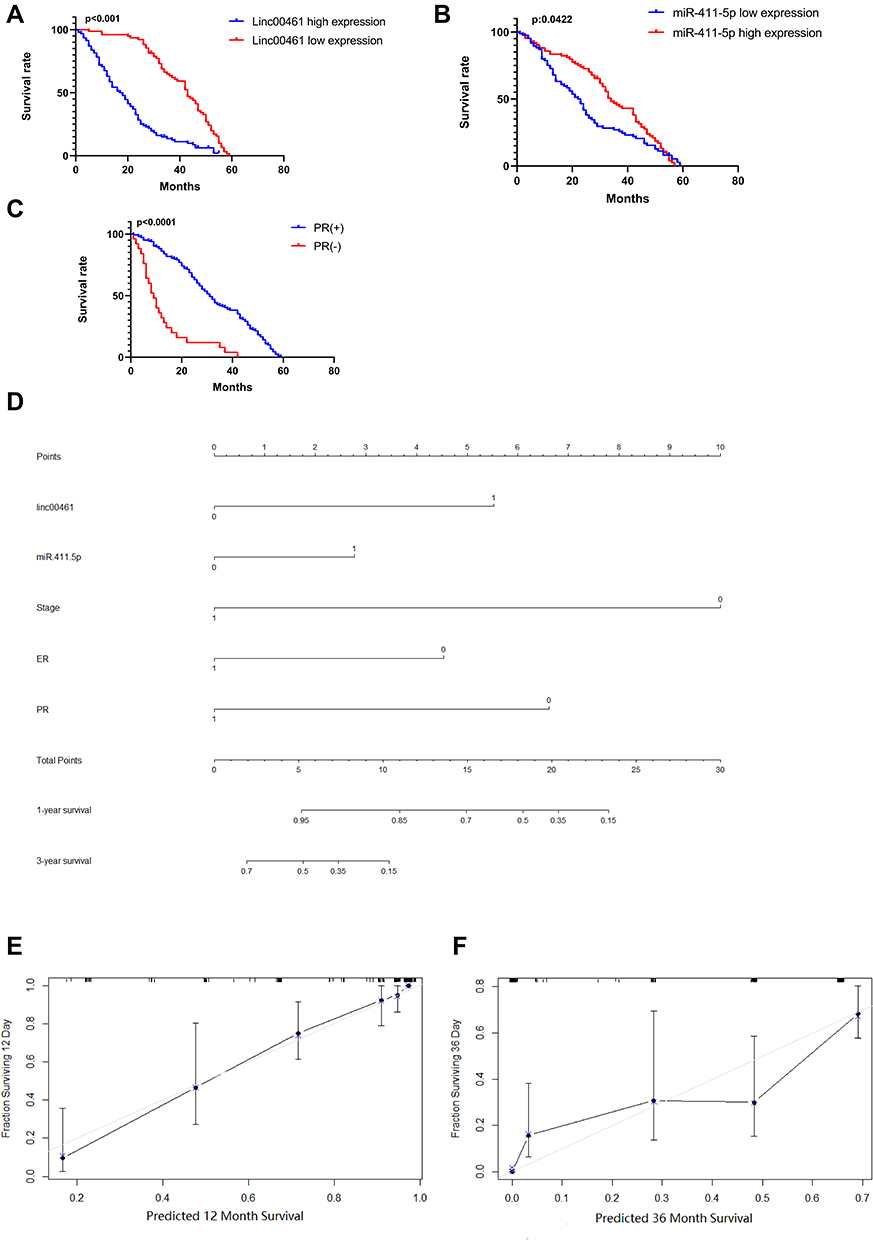 |
Figure 2 (A) Kaplan–Meier survival analysis was used to assess LINC00461 expression and BC patients’ overall survival. (B) Kaplan–Meier survival analysis was used to assess miR-411-5p expression and BC patients’ overall survival. (C) Kaplan–Meier survival analysis was used to assess PR status and BC patients’ overall survival. (D) Nomogram for prediction of 3- and 5-year survival rates for patients with BC. (E) Calibration curves for 3-year overall survival prediction. (F) Calibration curves for 5-year overall survival prediction. |
LINC00461 Can Negatively Regulate miR-411-5p Expression
We found that miR-411-5p was expressed at high levels in MDA-MB-231 and MCF-7 cell lines when LINC00461 expression was knocked down ( Supplemental Fig C and D and Figure 3A); however, while miR-491 levels were also influenced by LINC00461 knockdown, the difference was not statistically significant (Figure 3B). Further, we found that up- or down-regulation of miR-411-5p did not influence LINC00461 expression, indicating that LINC00461 may directly target miR-411-5p (Figure 3C and D).
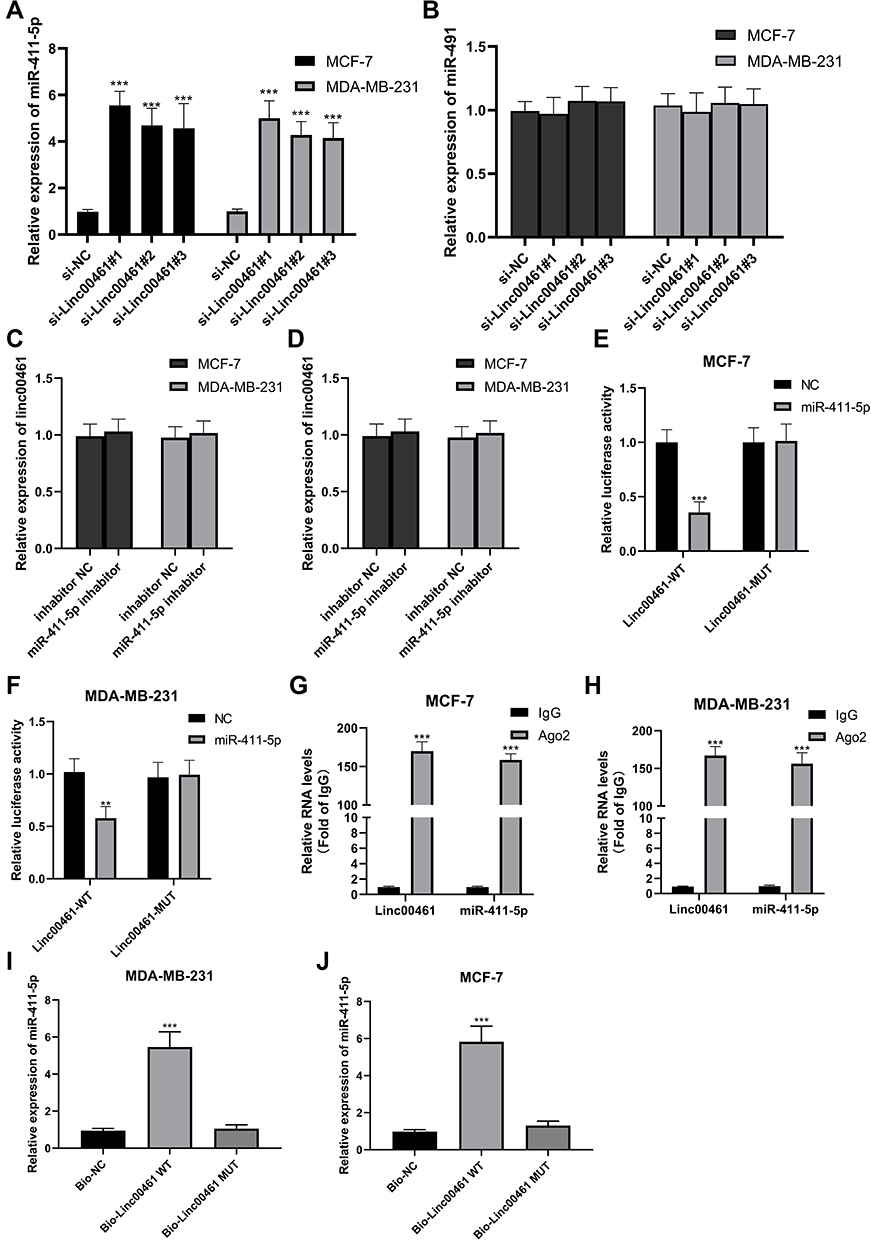 |
Figure 3 (A) miR-411-5p expression in LINC00461 down-regulated MCF-7 and MDA-MB-231 cell lines. (B) miR-491 expression in LINC00461 down-regulated MCF-7 and MDA-MB-231 cell lines. (C) LINC00461 expression in miR-411-5p up-regulated MCF-7 and MDA-MB-231 cell lines. (D) LINC00461 expression in miR-411-5p down-regulated MCF-7 and MDA-MB-231 cell lines. (E) The relative luciferase activity in MCF-7 cells cotransfected with luciferase reporter vectors containing Linc00461-WT or Linc00461-MUT and miR-control or miR-411-5p. (F) The relative luciferase activity in MDA-MB-231 cells transfected with luciferase reporter vectors containing Linc00461-WT or Linc00461-MUT and miR-control or miR-411-5p. (G) RIP assays were performed in MCF-7 cells. (H) RIP assays were performed in MDA-MB-231 cells. (I) RNA pull-down assays were performed in MCF-7 cells. (J) RNA pull-down assays were performed in MDA-MB-231 cells. Data represent the mean ± SD. **P <0.01. ***P <0.001. The experiments were independently repeated at least three times. |
To further understand the mechanism underlying LINC00461 regulation of miR-411-5p, we performed dual-luciferase reported gene assays. The result showed that miR-411-5p could significantly decrease the luciferase activity driven by LINC00461-WT in the MDA-MB-231 and MCF-7 cell lines, while activity from LINC00461-MUT was not significantly influenced (Figure 3E and F). Further, RIP experiments revealed that miR-411-5p was expressed at high levels in the LINC00461-WT group in both BC cell lines (Figure 3G and H). Moreover, RNA pull-down assays showed that larger amounts of LINC00461 were pulled down from BC cells expressing miR-411-5p-WT than from those expressing miR-411-5p-MUT, demonstrating that LINC00461 can bind directly to the WT miR-411-5p sequence (Figure 3I and J). Together, these data suggest that LINC00461 can directly interact with miR-411-5p to exert its biological functions.
LINC00461 Is a Potential Oncogene and miR-411-5p a Possible Tumor Suppressor
Knockdown of LINC00461 led to decreased proliferation of MDA-MB-231 and MCF-7 cells (Figure 4A-B), while up- and down-regulation of miR-411-5p inhibited and promoted proliferation of these cells, respectively (Figure 4C–F). In migration tests, LINC00461 knockdown enhanced migration of MDA-MB-231 and MCF-7 cells (Figure 4G and H). In contrast, over- and under-expression of miR-411-5p led to decreased and increased cell migration rates, respectively, in MDA-MB-231 and MCF-7 cells (Figure 4I and J).
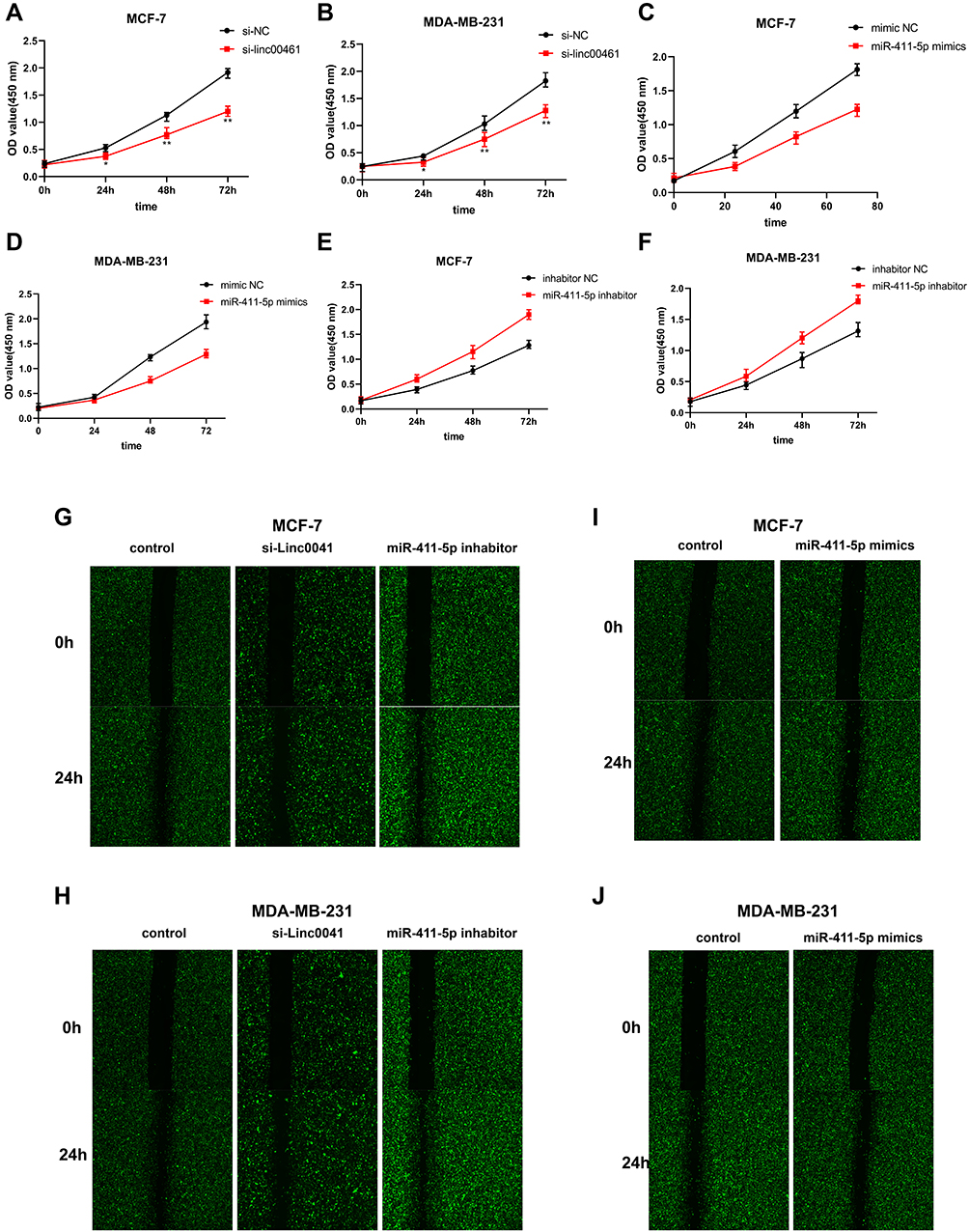 |
Figure 4 (A and B) CCK8 assays of MCF-7 and MDA-MB-231 cell lines following NC or LINC00461 knockdown. (C and D) CCK8 assays of MCF-7 and MDA-MB-231 cell lines following NC or miR-411-5p overexpression. (E and F) CCK8 assays of MCF-7 and MDA-MB-231 cell lines following NC or miR-411-5p knockdown. (G–J) Wound-healing assays of MCF-7 and MDA-MB-231 cell lines following NC or LINC00461 knockdown and NC or miR-411-5p overexpression and NC or miR-411-5p knockdown. Data represent the mean ± SD. *P <0.05. **P <0.01. The experiments were independently repeated at least three times. |
LINC00461 and miR-411-5p May Regulate DTX Resistance
Our results indicated that LINC00461 expression was higher in MCF-7-DTX and MDA-MB-231-DTX cells than that in their corresponding non-resistant parent cell lines (MCF-7 and MDA-MB-231) (Figure 5A and B). Moreover, LINC00461 knockdown significantly decreased the survival rates of MCF-7-DTX and MDA-MB-231-DTX cells treated with DTX (Figure 5C and D).
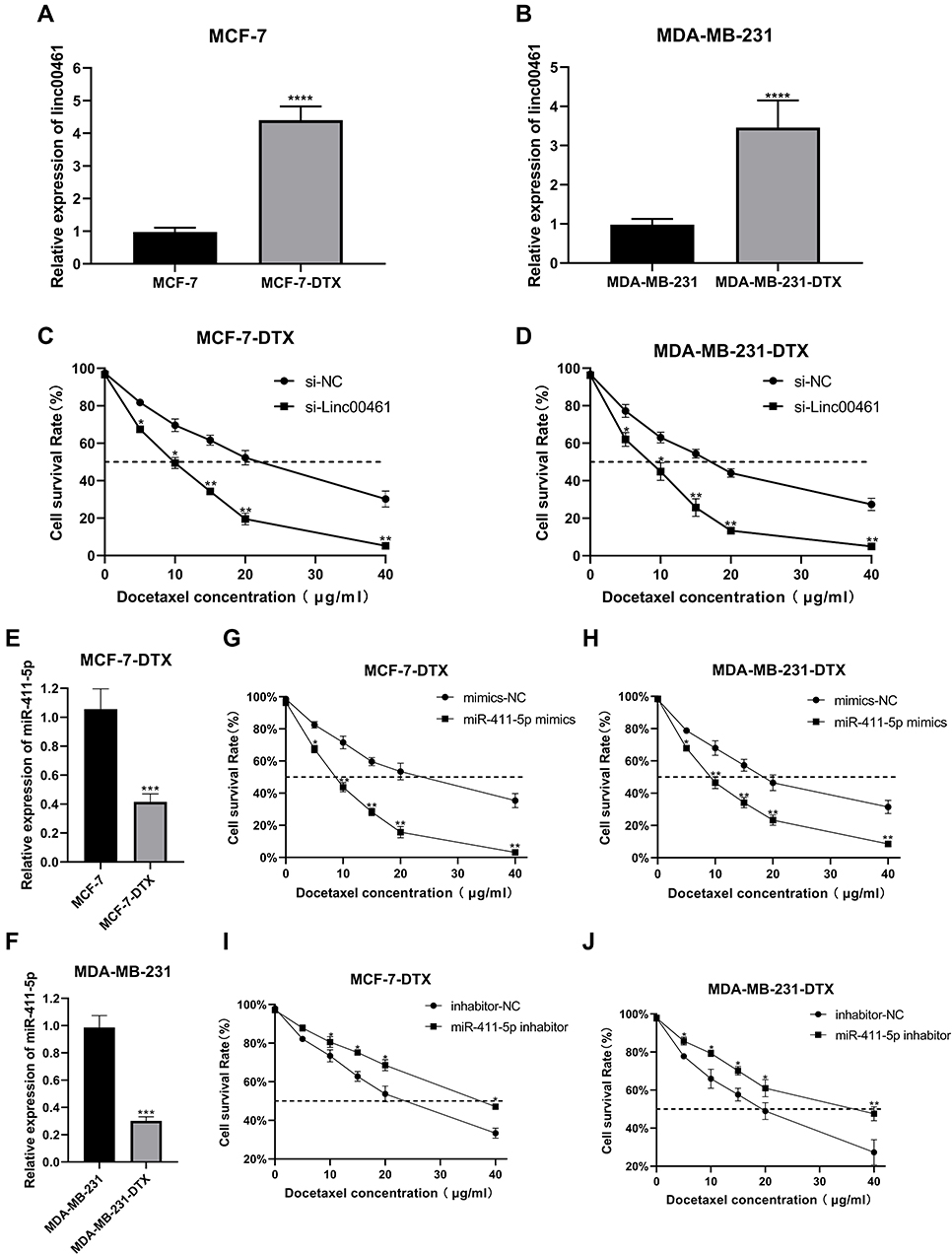 |
Figure 5 (A and B) Expression of LINC00461 in MCF-7-DTX and MDA-MB-231-DTX cell lines. (C and D) CCK8 assays of MCF-7-DTX and MDA-MB-231-DTX cell lines following NC or LINC00461 knockdown. (E and F) Expression of miR-411-5p in MCF-7-DTX and MDA-MB-231-DTX cell lines. (G and H) CCK8 assays of MCF-7-DTX and MDA-MB-231-DTX cell lines following NC or miR-411-5p overexpression. (I and J) CCK8 assays of MCF-7-DTX and MDA-MB-231-DTX cell lines following NC or miR-411-5p knockdown. Data represent the mean ± SD. *P <0.05. **P <0.01. ***P <0.001. ****P <0.0001. The experiments were independently repeated at least three times. |
miR-411-5p was also down-regulated in DTX-resistant BC cell lines (Figure 5E and F), while overexpression of miR-411-5p led to decreased survival rates (Figure 5G and H) and miR-411-5p knockdown resulted in increased survival rates in response to DTX treatment (Figure 5I and J).
Discussion
BC is the most prevalent gynecological malignancy worldwide.2,23 Although there has been great progress in the diagnosis and treatment of BC, overall survival rates remain far from satisfactory.24 Numerous investigations have found that lncRNAs are involved in tumorigenesis and cancer development.25–30 Yang et al identified lncRNA LINC00461 as significantly up-regulated in glioma tissues.22 Further, Dong et al found that LINC00461 was up-regulated in BC and can promote BC cell migration and invasion through miR-30a-5p/ITGB3 axis.31 In our early research, we found that the expression of LINC00461 was up-regulated in chemotherapy-resistant cells. Additional LINC00461 can promote glioma cell proliferation and migration; however, the biological functions of LINC00461 remain unknown. Deeper research into the physiological roles of LINC00461 has the potential to contribute to more effective therapeutic modalities.
In this study, we found that LINC00461 is significantly up-regulated in BC tumors relative to adjacent non-tumor tissue samples. Further, survival analysis indicated that high levels of LINC00461 are closely associated with poor prognosis in patients with BC, and our nomogram predicts LINC00461 as an important risk factor for BC prognosis. Consistent with these findings, LINC00461 expression was significantly increased in BC cell lines. Moreover, levels of LINC00461 in DTX-resistant cell lines (MCF-7-DTX and MDA-MB-231-DTX) were higher than those in corresponding non-resistant lines (MCF-7 and MDA-MB-231).
To further understand the biological role of LINC00461 in BC, we also conducted functional assays, which showed that down-regulation of this lncRNA can inhibit BC cell proliferation and enhance cell migration in vitro. Moreover, knockdown of LINC00461 in DTX-resistant cell lines significantly decreased cell viability in response to DTX treatment. Together, these findings indicate that LINC00461 may have an important role in BC development and is a potential oncogene contributing to DTX resistance in this disease.
Previous research showed that lncRNAs can participate in multiple diseases by acting as competitive endogenous RNAs (ceRNAs) through binding to specific miRNAs.32 As LINC00461 was mainly localized to the cytoplasm, we hypothesized it may act as a ceRNA in BC. Hence, we used bioinformatic analysis to identify possible targets, resulting in the identification of miR-411-5p as a potential downstream target of LINC00461. Subsequent experiments showed that miR-411-5p is down-regulated in both BC tissues and cell lines. Further, miR-411-5p expression increased on LINC00461 knockdown in BC cells, while neither up- nor down-regulation of miR-411-5p in BC cells influenced LINC00461 expression. The results of dual-luciferase reporter gene, RIP, and RNA pull-down assays further demonstrated that LINC00461 can directly interact with miR-411-5p to act as a ceRNA. Moreover, overexpression of miR-411-5p could inhibit proliferation and decrease migration rates of BC cells, and miR-411-5p was down-regulated in DTX-resistant BC cell lines compared with their corresponding parent BC cell lines. In CCK-8 experiments, up-regulation of miR-411-5p decreased BC cell proliferation and cell viability, while down-regulation of this miR-411-5p had the opposite effects.
Conclusion
Overall, this study illustrated that LINC00461 was increased in BC tissues and cells. The clinical significance of LINC00461 was also detected. Moreover, our results indicate that LINC00461 promotes DTX-resistance in BC by acting as a sponge for miR-411-5p molecules. These findings highlight the potential role of LINC00461/miR-411-5p as a new prognostic evaluation and a therapeutic target in patients with BC.
Disclosure
The authors report no conflicts of interest in this work.
References
1. DeSantis CE, Fedewa SA, Goding SA, et al. Breast cancer statistics, 2015: convergence of incidence rates between black and white women. CA Cancer J Clin. 2016;66(1):31–42.
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
3. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
4. Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. doi:10.3322/caac.21208
5. Shao X, Sun B, Shen Y. Axillary reverse mapping (ARM): where to go. Breast Cancer. 2019;26(1):1–10. doi:10.1007/s12282-018-0886-0
6. Gradishar WJ, Anderson BO, Balassanian R, et al. NCCN guidelines insights: breast cancer, version 1.2017. J Natl Compr Canc Netw. 2017;15(4):433–451. doi:10.6004/jnccn.2017.0044
7. Gradishar WJ, Anderson BO, Balassanian R, et al. Invasive breast cancer version 1.2016, NCCN clinical practice guidelines in oncology. J Natl Compr Canc Netw. 2016;14(3):324–354. doi:10.6004/jnccn.2016.0037
8. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
9. Guttman M, Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature. 2012;482(7385):339–346. doi:10.1038/nature10887
10. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. doi:10.1038/nrg2521
11. Xie C, Yuan J, Li H, et al. NONCODEv4: exploring the world of long non-coding RNA genes. Nucleic Acids Res. 2014;42(Databaseissue):D98–D103. doi:10.1093/nar/gkt1222
12. Batista PJ, Chang HY. Long noncoding RNAs: cellular address codes in development and disease. Cell. 2013;152(6):1298–1307. doi:10.1016/j.cell.2013.02.012
13. Chen LL, Carmichael GG. Decoding the function of nuclear long non-coding RNAs. Curr Opin Cell Biol. 2010;22(3):357–364. doi:10.1016/j.ceb.2010.03.003
14. Jalali S, Bhartiya D, Lalwani MK, Sivasubbu S, Scaria V. Systematic transcriptome wide analysis of lncRNA-miRNA interactions. PLoS One. 2013;8(2):e53823. doi:10.1371/journal.pone.0053823
15. Tang Q, Ni Z, Cheng Z, et al. Three circulating long non-coding RNAs act as biomarkers for predicting NSCLC. Cell Physiol Biochem. 2015;37(3):1002–1009. doi:10.1159/000430226
16. Zhou X, Ye F, Yin C, et al. The interaction between MiR-141 and lncRNA-H19 in regulating cell proliferation and migration in gastric cancer. Cell Physiol Biochem. 2015;36(4):1440–1452. doi:10.1159/000430309
17. Sun M, Liu XH, Lu KH, et al. EZH2-mediated epigenetic suppression of long noncoding RNA SPRY4-IT1 promotes NSCLC cell proliferation and metastasis by affecting the epithelial-mesenchymal transition. Cell Death Dis. 2014;5:e1298. doi:10.1038/cddis.2014.256
18. Wang X, Goodrich KJ, Gooding AR, et al. Targeting of polycomb repressive complex 2 to RNA by short repeats of consecutive guanines. Mol Cell. 2017;65(6):1056–1067.e5. doi:10.1016/j.molcel.2017.02.003
19. Wang W, He X, Zheng Z, et al. Serum HOTAIR as a novel diagnostic biomarker for esophageal squamous cell carcinoma. Mol Cancer. 2017;16(1):75. doi:10.1186/s12943-017-0643-6
20. Ji D, Wang Y, Li H, Sun B, Luo X. Long non-coding RNA LINC00461/miR-149-5p/LRIG2 axis regulates hepatocellular carcinoma progression. Biochem Biophys Res Commun. 2019;512(2):176–181. doi:10.1016/j.bbrc.2019.03.049
21. Deng M, Yuan H, Liu S, Hu Z, Xiao H. Exosome-transmitted LINC00461 promotes multiple myeloma cell proliferation and suppresses apoptosis by modulating microRNA/BCL-2 expression. Cytotherapy. 2019;21(1):96–106. doi:10.1016/j.jcyt.2018.10.006
22. Yang Y, Ren M, Song C, et al. LINC00461, a long non-coding RNA, is important for the proliferation and migration of glioma cells. Oncotarget. 2017;8(48):84123–84139. doi:10.18632/oncotarget.20340
23. Tao Z, Shi A, Lu C, et al. Breast cancer: epidemiology and etiology. Cell Biochem Biophys. 2015;72(2):333–338. doi:10.1007/s12013-014-0459-6
24. Chou J, Wang B, Zheng T, et al. MALAT1 induced migration and invasion of human breast cancer cells by competitively binding miR-1 with cdc42. Biochem Biophys Res Commun. 2016;472(1):262–269. doi:10.1016/j.bbrc.2016.02.102
25. Shi Y, Li J, Liu Y, et al. The long noncoding RNA SPRY4-IT1 increases the proliferation of human breast cancer cells by upregulating ZNF703 expression. Mol Cancer. 2015;14(1):51. doi:10.1186/s12943-015-0318-0
26. Liang Y, Song X, Li Y, et al. A novel long non-coding RNA-PRLB acts as a tumor promoter through regulating miR-4766-5p/SIRT1 axis in breast cancer. Cell Death Dis. 2018;9(5):563. doi:10.1038/s41419-018-0582-1
27. Liang Y, Li Y, Song X, et al. Long noncoding RNA LINP1 acts as an oncogene and promotes chemoresistance in breast cancer. Cancer Biol Ther. 2018;19(2):120–131. doi:10.1080/15384047.2017.1394543
28. Huarte M. The emerging role of lncRNAs in cancer. Nat Med. 2015;21(11):1253. doi:10.1038/nm.3981
29. Gao Y, Zhang Z, Li K, et al. Linc-DYNC2H1-4 promotes EMT and CSC phenotypes by acting as a sponge of miR-145 in pancreatic cancer cells. Cell Death Dis. 2017;8(7):e2924. doi:10.1038/cddis.2017.311
30. Xu Y, Qin L, Sun T, et al. Twist1 promotes breast cancer invasion and metastasis by silencing Foxa1 expression. Oncogene. 2017;36(8):1157–1166. doi:10.1038/onc.2016.286
31. Dong L, Qian J, Chen F, Fan Y, Long J. LINC00461 promotes cell migration and invasion in breast cancer through miR-30a-5p/integrin beta3 axis. J Cell Biochem. 2019;120(4):4851–4862. doi:10.1002/jcb.27435
32. Thomson DW, Dinger ME. Endogenous microRNA sponges: evidence and controversy. Nat Rev Genet. 2016;17(5):272. doi:10.1038/nrg.2016.20
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.