Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 16
Leptin Mediates Prostate Stromal Cell Proliferation, Smooth Muscle Contraction, and Mitochondrial Function in Benign Prostate Hyperplasia
Authors Wang X ![]() , Guo L, Liu G, Liu T
, Guo L, Liu G, Liu T
Received 24 May 2023
Accepted for publication 13 August 2023
Published 19 October 2023 Volume 2023:16 Pages 3261—3273
DOI https://doi.org/10.2147/DMSO.S420258
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Gian Paolo Fadini
Xiaolong Wang,1 Linfa Guo,1 Guiyong Liu,2 Tongzu Liu1,3,4
1Department of Urology, Zhongnan Hospital of Wuhan University, Wuhan, People’s Republic of China; 2Qianjiang Central Hospital of Hubei Province, Qianjiang, People’s Republic of China; 3Cancer Precision Diagnosis and Treatment and Translational Medicine Hubei Engineering Research Center, Wuhan, People’s Republic of China; 4Hubei Province Key Laboratory of Urinary System Diseases, Wuhan, People’s Republic of China
Correspondence: Tongzu Liu; Guiyong Liu, Department of Urology, Cancer Precision Diagnosis and Treatment and Translational Medicine Hubei Engineering Research Center, Hubei Province Key Laboratory of Urinary System Diseases, Zhongnan Hospital of Wuhan University, Wuhan, 430071, People’s Republic of China, Email [email protected]; [email protected]
Introduction: Leptin is a metabolic peptide hormone produced by adipocytes, with proven roles in proliferation of prostate cancer cells and of prostate cells in animal models of benign prostatic hyperplasia (BPH). Thus, the role of leptin as a molecular link connecting BPH and lower urinary tract symptoms (LUTS) suggestive of BPH with metabolic symptoms appears feasible but is still unknown. In fact, a connection between metabolic syndrome and BPH is becoming increasingly evident from epidemiologic studies. Key factors of Lower urinary tract symptoms associated with benign prostatic hyperplasia (BPH/LUTS) are increased prostate smooth muscle tone, and prostate enlargement. Here, we examined the effects of leptin on contraction of human prostate smooth muscle and on growth of stromal cells.
Methods: We performed microarray analysis to identify genes (fold change ≥ 1.5) associated with BPH/LUTS progression, such as those involved in proliferation, apoptosis, and mitochondrial metabolism, in rat prostate tissue (data from GSE129561). We then used electric field stimulation (EFS) to induce frequency-dependent, neurogenic contractions of human prostate strips, which were enhanced by leptin. We also examined the effect of leptin on human prostate stromal cells (WPMY-1) and found increased cell proliferation and viability upon exposure. To explore the underlying mechanism, we conducted mitochondrial stress assay using near-infrared (NIR) fluorescent dye and flow cytometry (FACS) analysis and observed reduced cellular apoptosis and preserved mitochondrial membrane potential (∆ψM) after leptin treatment.
Results: Microarray analysis reveals that leptin regulates prostate smooth muscle contraction and stromal cell proliferation, shedding new light on its involvement in BPH/LUTS pathogenesis and mitochondrial function. We found that leptin enhanced the proliferation rate of prostate stromal cells relative to the control group (0.67 ± 0.05 vs 0.54 ± 0.08, p-value= 0.024). Moreover, leptin (100 ng/mL) potentiated the frequency-dependent, neurogenic contractions of prostate strips elicited by EFS (p= 0.047 between leptin and control group). We also show that leptin treatment increased the mitochondrial membrane potential of prostate stromal cells and inhibited mitochondrial apoptosis.
Discussion: Our results indicate that leptin stimulates the contractility and proliferation of smooth muscle and stromal cells in the human prostate, implying a potential role for leptin in exacerbating BPH/LUTS in obese men. Leptin modulation may be a beneficial therapeutic strategy for patients with metabolic syndrome and BPH/LUTS. Further studies are warranted to elucidate the mechanisms and implications of the leptin system in BPH/LUTS.
Keywords: metabolic syndrome, leptin, prostate, contraction, mitochondrial
Introduction
Metabolic syndrome (MetS) is a cluster of metabolic disorders, including insulin resistance, obesity, hypertension, and dyslipidemia. The prevalence of metabolic syndrome is substantial worldwide, with almost 30% of the global adult population being afflicted. Even more alarming is the number of children and adolescents affected by metabolic syndrome, with 25.8 million children and 35.5 million adolescents reportedly affected by the conditions, respectively.1 According to recent studies, the prevalence of MetS is highest in the United States, with 32% of the population affected, whereas 24% and 23% of the European and Asia-Pacific populations are affected, respectively. The high prevalence rates of MetS in these regions may indicate an increased risk of cardiovascular diseases, underscoring the urgent need for preventive measures to mitigate the potential epidemic.2 Additionally, recent studies have suggested that MetS may be linked to various urological disorders, such as erectile dysfunction, urinary stones, and prostate cancer. Therefore, it is essential to investigate the relationship between MetS and urological disorder, as it may have significant implications for the prevention and management of urological conditions. Recent studies have indicated that MetS may contribute to the development of benign prostatic hyperplasia (BPH) and lower urinary tract symptoms (LUTS) in men.3 LUTS include both voiding symptoms such as hesitancy, weak stream, intermittency, straining, and incomplete emptying, and storage symptoms such as frequency, urgency, nocturia, and urgency incontinence. A common and significant health issue is the presence of LUTS associated with benign prostatic hyperplasia (BPH/LUTS), which can impair the quality of life considerably. The association between MetS and BPH/LUTS is complex and multifactorial.4 The link between MetS and BPH/LUTS has been identified as mediated by a number of essential mechanistic elements, such as chronic inflammation, oxidative stress, hormonal imbalances, insulin-like growth factor signaling, sympathetic overactivity, endothelial dysfunction, and adipokine dysregulation.5 These pathways encompass a diverse array of molecular and cellular mechanisms that underly the metabolic and endocrine perturbations that are commonly observed in individuals with MetS and BPH/LUTS. A more thorough comprehension of the pathophysiology of BPH/LUTS related to MetS can be attained through elucidating these ties, potentially leading to the creation of more precise and efficacious therapeutic treatments.
The current consensus among medical professionals is that the approaches to diagnosing and treating LUTS are rooted in the understanding of the lower urinary tract as a unified construct. Moreover, recent studies on LUTS have revealed a number of essential proteins involved in both adrenergic and non-adrenergic mediated prostate smooth muscle contraction and stromal cell proliferation as well, such as LIM kinase,6 G protein-coupled receptor kinase,7 polo-like kinase8 and others.9 A key aspect of the pathophysiology of MetS associated to BPH/LUTS is the alternation of prostate growth and contraction. MetS may provide a circumstance that enhance the smooth muscle tone and proliferation in the unify process. However, the molecular mechanisms underlying this dysregulation remain unclear.
Leptin is a metabolic protein hormone (16 kD in size) that is secreted by adipocytes and acts on the Ob receptor (obesity receptor, Ob-R or leptin Receptor, LepR) on the cell membrane.10 It has a positive correlation with body fat content and body mass index (BMI). MetS patients, however, have notably greater levels of leptin than normal people, and may even develop leptin resistance in cases of cardiovascular diseases caused by MetS.11 Several recent studies have indicated the potential for leptin to augment proliferation and metastasis in prostate cancer.12–14 Additionally, in non-malignant conditions, it has been demonstrated that leptin hastens the re-programming of smooth muscle in the airway and cardiovascular system.15 These findings suggest a broad range of effects and highlight the need for further investigation into the mechanistic pathways activated by leptin in different biological contexts.16 The effect of leptin as a metabolic hormone and its corresponding signaling pathway might be a potential contributor to deviant expansion and contraction of prostate smooth muscle, but the clear mechanism of this phenomenon still unknown.
The aim of this study is to investigate the role of leptin in MetS and BPH/LUTS using a microarray analysis and prostate stromal cells. We propose that leptin modulates gene expression in the prostate to stimulate cell proliferation and smooth muscle contraction, thereby contributing to BPH pathogenesis and progression. Furthermore, we explore the involvement of leptin receptors and their downstream signaling pathways in mediating these effects.
Materials and Methods
Micro-Array
We obtained the raw data for this study from the Gene Expression Omnibus (GEO) database (accession number: GSE129561). This dataset was derived from one of our previous studies, which provided detailed information on the animal model and sample treatment used in this research.
In this study, we incited the microarray data from rats model that simulated the pathological features of BPH/LUTS. Male Sprague Dawley rats (180–220g), aged six weeks, were kept under controlled environmental conditions. After a period of acclimation, rats were castrated and then received subcutaneous injections of dihydrotestosterone (3mg/kg) for four weeks to induce BPH. The rats also received oral administration of vehicle (distilled water) daily, starting 72 h after castration17 (Supplement Figure 1).
We used the R package limma to perform quality control and normalization of the data, and to identify differentially expressed genes (DEGs) between the control and BPH groups. We applied a cut-off of |log2 fold change|>1 and adjusted p-value <0.05 to select the DEGs. We visualized the DEGs with heatmap and volcano plot, and conducted functional enrichment analysis using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. Our results revealed several potential pathways and genes that could be involved in the development and progression of BPH.
CCK-8 and EdU
WPMY-1 is a human prostatic stromal myofibroblast cell line derived from the normal peripheral zone of an adult prostate. This study aimed to evaluate its proliferation and viability in response to leptin treatment using two assays: Cell Counting Kit 8 (WST-8/CCK8) (Abcam, Cambridge, United Kingdom) and 5-ethynyl-2’-deoxyuridine (EdU) (Abcam, Cambridge, United Kingdom). For the CCK-8 assay, WPMY-1 cells were seeded in 96-well plates and treated with different concentrations of leptin for 24, 48, or 72 h. Then, 10μL of CCK-8 solution was added to each well and incubated for 2 h at 37°C. The absorbance at 450 nm was determined by a microplate reader. For the EdU assay, WPMY-1 cells were seeded in glass-bottom dishes and treated with different concentrations of leptin for 24 or 48 h. Then, 50 μM of EdU solution was added to each dish and incubated for an additional 2 h at 37 °C. After permeabilization and fixation, the cells were stained with Apollo® 488 azide (Cell Signaling Technology, Inc., Danvers, MA, United States) and Hoechst 33342 (Thermo Fisher Scientific Inc., Waltham, MA, United States) according to the manufacturer’s instructions. The fluorescence images were captured by a confocal microscope (Leica Confocal System, Mannheim, Germany).
Human Tissue
This study was conducted in accordance with the ethical principles of the World Medical Association’s Declaration of Helsinki and approved by the ethics committee of Wuhan University. We collected prostate tissue samples from ten patients who underwent prostatectomy for prostate cancer. All patients provided their voluntary and informed consent, and their identities were anonymized. A pathologist performed a macroscopic examination of the tissues and excluded those with signs of neoplasia, cancer, or inflammation. The organ bath experiments were carried out within 2 h of tissue collection.
Tension Measurements
We conducted a study to examine the contractile properties of prostatic tissue strips obtained from patients who underwent laparoscopic radical prostatectomy. A pathologist prepared each sample into strips of 5×3×3 mm dimensions. We mounted the strips in four-chamber (Beijing Jinggong Hongtai Tech, Beijing, China), and tissue baths containing 10 mL of oxygenated Krebs-Henseleit solution at 37°C and pH 7.4. We applied a resting tension of 4.9 mN to each strip and let them equilibrate for 45 minutes, adjusting the tension three times to maintain a steady smooth muscle tone. We then measured the maximal contraction induced by 80 mM KCl in each strip. After 30 minutes of incubation, we washed the strips with Krebs-Henseleit solution three times, with or without inhibitors or DMSO as a control. We then performed cumulative concentration-response curves for various contractile agonists and frequency-response curves with EFS, which simulates action potentials and triggers the release of endogenous neurotransmitters. We assessed the extracellular Ca2+ influx by washing KCl with a Ca2+-free Krebs-Henseleit solution, followed by adding KCl (80 mM) to depolarize the cell membrane and activate voltage-dependent Ca2+ channels.
We investigated the effect of leptin on the contractility of human prostate tissues by applying EFS at a series of frequencies (2 Hz, 4 Hz, 8 Hz, 16 Hz, and 32 Hz) (Beijing Jinggong Hongtai Tech, Beijing, China). We pre-incubated the tissues with leptin for 120 min before constructing the frequency curves. To control for variability among prostate samples and patients, such as stromal ratios, smooth muscle content, BPH intensity, or other factors, we normalized the EFS-induced contractions to those induced by 80 mM KCl. We used four chambers of an organ bath for each experiment and allocated them randomly to the control group (DMSO) or the leptin group. We recorded and analysed the maximum contraction levels at each concentration and frequency in leptin incubation chambers and control chambers by SPSS 19 (IBM SPSS Statistics, Chicago, IL, USA) and PRISM Pad 9 (Graphstats Technologies, San Diego, CA, USA).
Cell Contraction Assay
We performed a cell contraction assay (Cell Biolabs, San Diego, CA, USA) in collagen gels to assess the contractile activity of prostate stromal cells, a widely used method to investigate cell-extracellular matrix interactions. After embedding prostate stromal cells in collagen gels and incubating them for 24 h to allow sufficient time for cell attachment and matrix remodelling, we detached the gels from the culture wells and monitored them at distinct time points (1, 3, 12, and 24 h) to observe the degree of contraction. We calculated the gel area using ImageJ software and normalized it to the initial area to quantitatively evaluate the cellular contraction. To ascertain the role of myosin II in the contraction process, we compared a group of gels treated with 2,3-butanedione monoxime (BDM), a non-specific inhibitor of myosin ATPase activity, with untreated gels. This approach enabled us to provide insights into the underlying mechanisms involved in prostate stromal cell contraction in response to leptin treatment.
Fluorescence-Activated Cell Sorting (FACS)
Measurement of Mitochondrial Membrane Potential
To measure the mitochondrial membrane potential (∆ψM) of WPMY-1 cells, we used a mitochondrial staining reagent Near infrared (NIR) (Abcam, Cambridge, United Kingdom) staining method with a fixable viability dye that can be excited by a red laser and detected by a flow cytometer (BD Biosciences, East Rutherford, NJ, USA).18 We incubated the cells with 100 µL of 1X NIR staining solution for 30 minutes at room temperature in the dark, following the manufacturer’s instructions. Then, we washed the cells with 2 mL of cell staining buffer containing bovine serum albumin (BSA) to remove excess dye. Finally, we fixed and permeabilized the cells with 4% paraformaldehyde and 0.1% Triton X-100 for intracellular antibody staining. After staining with antibodies against ∆ψM-related proteins, we analyzed the cells by flow cytometry using a 633 or 635 nm laser for NIR detection and appropriate filters for other fluorophores.
Measurement of Mitochondrial Apoptosis
In apoptotic cells, JC-10 (Abcam, Cambridge, United Kingdom) diffuses into the cytosol as green-fluorescent monomers (Ex/Em=490/525nm).19 To perform JC-10 staining for WPMY-1 cells, a human prostate stromal cell line according to the following steps: Culture WPMY-1 cells in appropriate medium and conditions until 70–80% confluency. Treat cells with or without leptin for a suitable period (24 h and 48 h). Based on the instructure, we trypsinized the cells and washed them with phosphate-buffered saline (PBS). Then, we suspended them in a solution containing 15 µM of JC-10 and incubated them at 37°C for 10 minutes. After that, we washed the cells again with PBS and resuspended them in assay buffer. Finally, we analyzed the cells by flow cytometry, using filters for green (FL1) and orange (FL2) fluorescence channels. We calculated the ratio of orange to green fluorescence intensity for each cell population. Compare the fluorescence intensity between control and treated cells to evaluate the rate of mitochondrial apoptosis.
Data Analysis
To analyse the gene expression, we utilized microarray analysis on the BPH rat model that we previously established and the GEO database, which employs different analytical tools to identify gene expression variances.17 In cell proliferation and NIR assay, we employed t-tests to measure the level of statistical significance. Furthermore, in the evaluation of data from the organ bath and JC-10 assay, we employed a one-way analysis of variance (ANOVA) test.
Results
Microarray Analysis and GO-Defined Category Show the BPH-Enriched Genes Focus on Proliferation, Apoptosis, and Mitochondrial Metabolism
To identify the molecular mechanisms underlying BPH, we performed microarray analysis on prostate tissues from control and BPH model rats. We found that 1023 genes were differentially expressed between the two groups, with 421 genes upregulated and 602 genes downregulated in BPH (Figure 1A). To further explore the biological functions of these genes, we performed Gene Ontology (GO) enrichment analysis and found that the most enriched GO terms were related to proliferation, apoptosis, and mitochondrial metabolism (Table 1 and Table 2 presented herein demonstrates a selection of genes that exhibit a log fold change greater than 1.5, including up-regulated genes and down-regulated genes). These results suggest that BPH is associated with altered regulation of cell growth, death, and energy metabolism in the prostate.
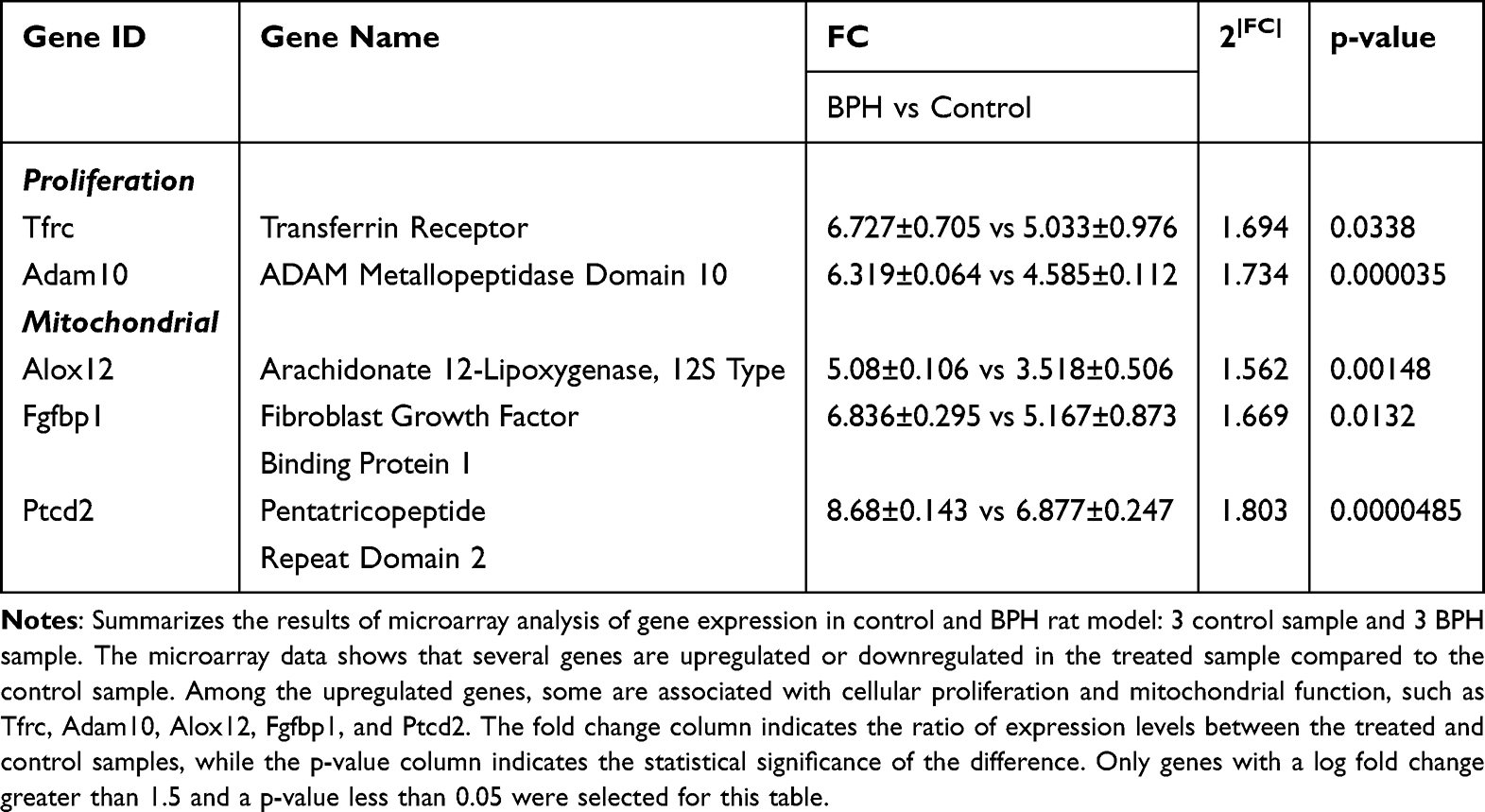 |
Table 1 Expression Levels for Up-Regulated Genes According to Primary Gene Ontology Function |
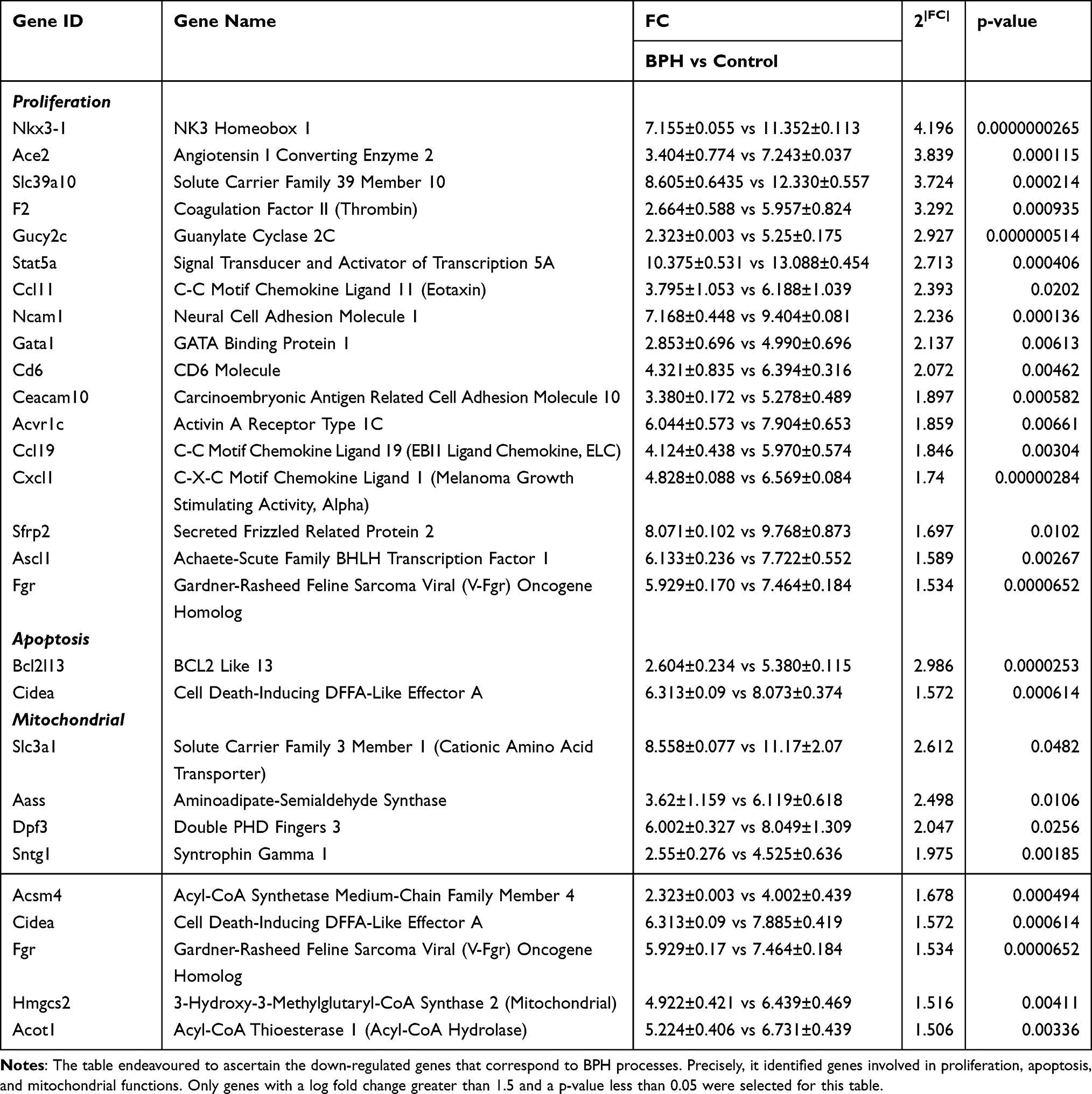 |
Table 2 Expression Levels for Down-Regulated Genes According to Primary Gene Ontology Function |
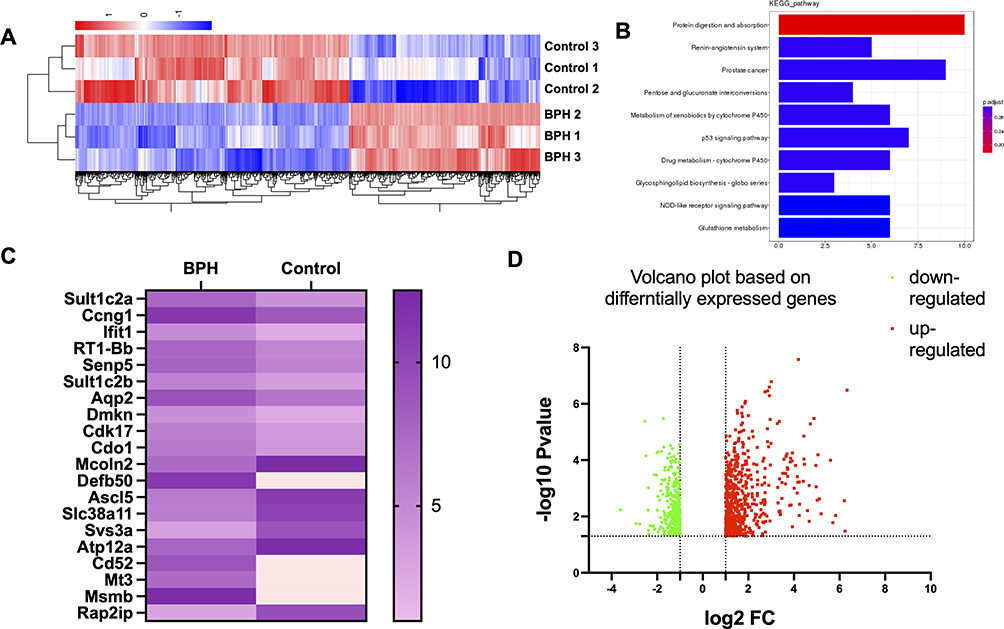 |
Figure 1 The differential gene expression between BPH and control rats (A) displays a heatmap comprising the DEGs enriched in proliferation and mitochondrial-related genes. The color bars on the top of the heatmap indicate these enriched categories. The color scale represents the log2 fold change of gene expression. (B) presents the KEGG analysis of the DEGs. This panel shows the top 10 enriched pathways based on their adjusted p-value. The color of the bars represents the enrichment score, and the number of DEGs in each pathway is indicated. Based on the KEGG analysis, the potential mechanisms and signaling pathways associated with BPH were identified. (C and D) presents a volcano plot and heatmap of the top 10 upregulated and top 10 downregulated target genes of the differentially expressed genes (DEGs) in a rat model of BPH. Among the top target genes, Sult1c2a, Ccng1, Ifit1, RT1-Bb, Senp5, Sult1c2b, Aqp2, Dmkn, Cdk17, Cdo1, and Mcoln2 were found to be upregulated, while Defb50, Ascl5, Slc38a11, Svs3a, Atp12a, Cd52, Mt3, Msmb, and Rap2ip were identified as downregulated in the BPH rats compared to the control group. The volcano plot, with the x-axis representing the log2 fold change and the y-axis the -log10 adjusted p-value, revealed that the downregulated genes were greater in number than the upregulated genes when compared to the control group, based on predefined cut-offs for fold change (>1.5 or <-1.5) and adjusted p-value (<0.05). The upregulated genes are represented by red dots, while the downregulated genes are represented by green dots, as shown by the dashed lines. |
In the present investigation, a microarray analysis was conducted, and gene ontology (GO)-defined categories were assessed utilizing the GEO database, to explore the molecular basis of BPH (Figure 1B and D). Our results revealed that genes enriched in BPH are primarily associated with biological processes such as proliferation, apoptosis, and mitochondrial metabolism (Figure 1C). These findings suggest that BPH is characterized by disrupted regulation of cellular growth, death, and energy production.
Leptin Treatment Enhances the Proliferation of Prostate Stromal Cells
In this study, we aimed to investigate the effect of leptin treatment on the proliferation of WPMY-1. We used two methods to measure cell proliferation. We found that leptin treatment significantly increased the proliferation of WPMY-1 in a dose-dependent manner compared to untreated controls. Our findings suggest that leptin may contribute to prostate enlargement by promoting stromal cell growth (Figure 2B). Exposure of WPMY-1 to Leptin (25 ng/mL) for 24 h resulted in raised proliferation, assessed by CCK-8. Similarly, 25 ng/mL leptin resulted in raised proliferation, compared to control cells by EdU assay (Figure 2A and C). By the expression of newly synthesized WPMY-1 DNA, the proliferation rate was completely increased compared to the control group (0.67 ± 0.05 vs 0.54 ± 0.08, p-value = 0.024).
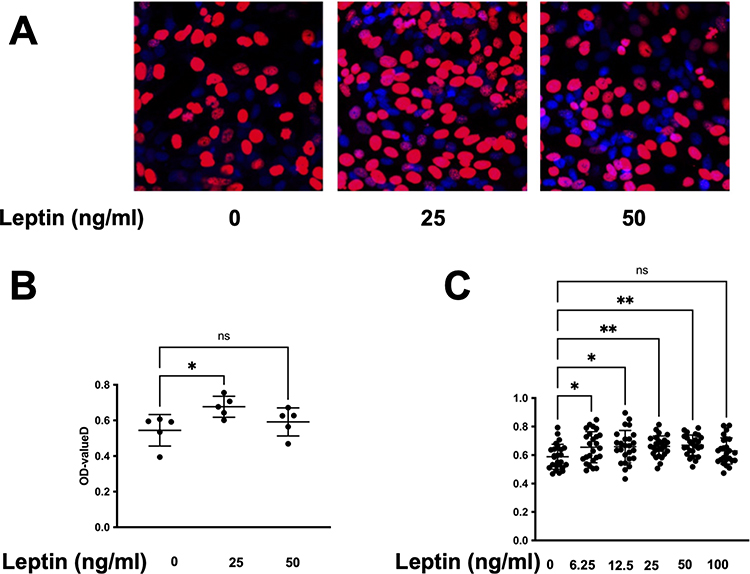 |
Figure 2 Leptin enhances the viability and proliferation of prostatic stromal cells. (A and B) EdU assay was performed to measure the proliferation of prostatic stromal cells treated with different concentrations of leptin (0, 25, 50 ng/mL) for 24 h. The results showed that leptin significantly increased the proliferation ratio of prostatic stromal cells at 25 ng/mL, but not at other concentrations. Data are presented as mean ± SD of three independent experiments. *P < 0.05, **P < 0.01 compared with control group. (C) CCK-8 assay was performed to measure the viability of prostatic stromal cells (WPMY-1) treated with different concentrations of leptin (0, 6.25, 12.5, 25, 50, and 100 ng/mL) for 24 h. The results showed that leptin significantly increased the viability of prostatic stromal cells in a dose-dependent manner. |
Leptin Treatment Facilitates Contraction of Prostate Smooth Muscle and Stromal Cells
We measured the contractile responses of prostate strips to EFS parallel with pharmacological agents before and after exposure to leptin. We found that leptin treatment significantly enhanced the contraction of prostate smooth muscle induced by EFS (Figure 3B). Our findings indicate that leptin may have a physiological role in regulating prostate smooth muscle tone. Electric field stimulation-induced frequency-dependent, neurogenic contractions of prostate strips, which were promoted by leptin (100 ng/mL) (p= 0.047 between leptin and control group). Smooth muscle contraction was increased to 112% of controls by 100 ng/mL, while no effect was observed by 50 ng/mL leptin (Figure 3A).
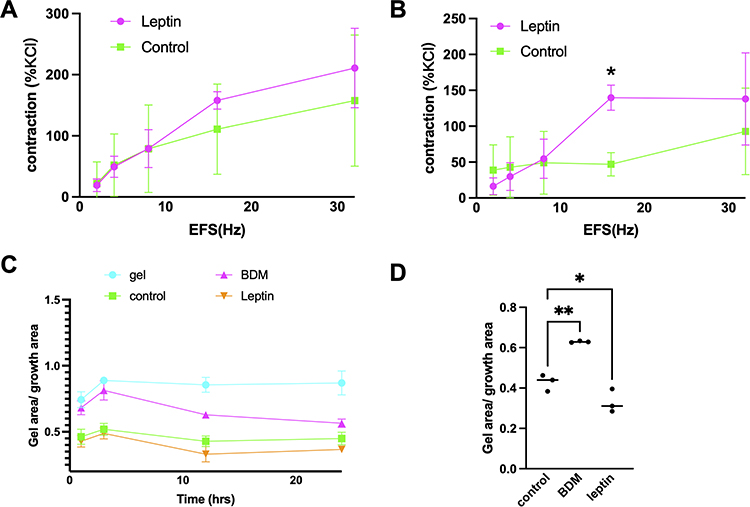 |
Figure 3 Effect of leptin on EFS-induced contraction of prostate strips and stromal cells. (A and B) Prostate strips were incubated with either 50 ng/mL or 100 ng/mL leptin for 30 min before being stimulated with EFS at 16 Hz. The amplitude of the contraction was measured and normalized to the initial response. Leptin significantly enhanced the contraction of prostate strips at 100 ng/mL compared to the control group (*p < 0.05). Data are presented as mean ± SEM of five independent experiments. (C) Leptin induces stromal cell contraction. Stromal cells were cultured in collagen gels and treated with leptin (25 ng/mL) or vehicle for 24 h in standard 24-well-plates. Quantification of gel contraction expressed as percentage of initial area. (D) Data are mean ± SEM of three independent experiments. (*p < 0.05), compared to vehicle control by One-way ANOVA. Control vs BDM: p=0.0017; Control vs Leptin: p=0.0451. (**p < 0.01). |
Leptin treatment significantly increased the contraction of WPMY-1 in 12 h (Figure 3C). The percentage of contraction was 0.428 ± 0.04 for the control group, 0.629 ± 0.04 for BDM, 0.33 ± 0.05 for leptin (p <0.0003) (Figure 3D). These results suggest that leptin stimulates prostate stromal cell contraction.
Leptin Treatment Increased the Mitochondrial Membrane Potential of Prostate Stromal Cells and Suppresses the Mitochondrial Apoptosis
To investigate the effect of leptin treatment on mitochondrial function and apoptosis of prostate stromal cells, we used JC-10 assay (Figure 4A) and near-infrared (NIR) spectroscopy (Figure 4B). We treated WPMY-1 with different concentrations of leptin for 24 h and measured JC-10 fluorescence intensity and NIR signals. We found that leptin treatment increased the NIR signals of WPMY-1, indicating enhanced mitochondrial respiration and oxidative phosphorylation. Leptin treatment also increased the JC-10 fluorescence intensity of WPMY-1, indicating reduced mitochondrial apoptosis, meanwhile higher NIR signals show increased ∆ψM.
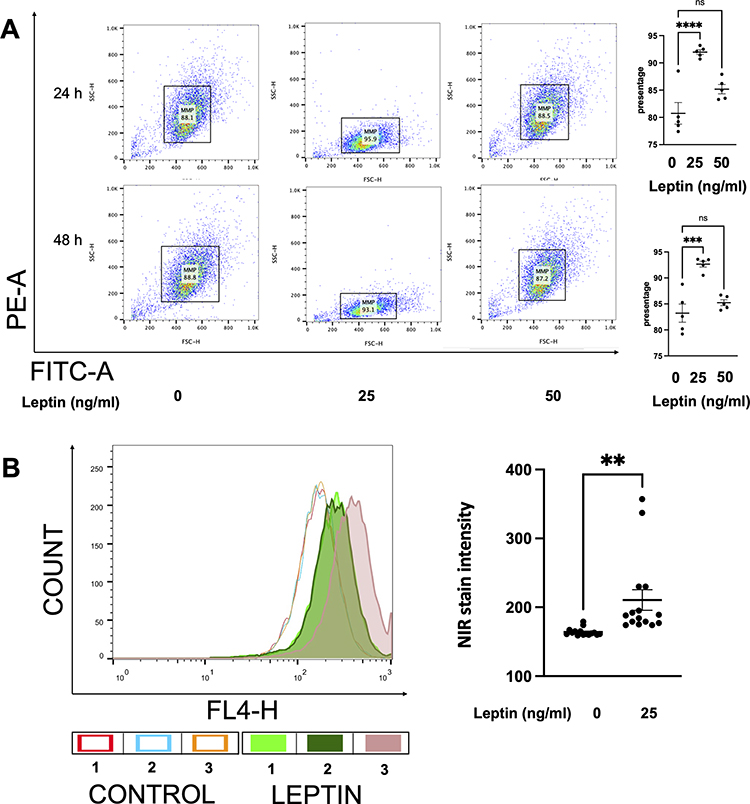 |
Figure 4 The assessment of mitochondrial-related apoptosis of prostatic stromal cells by NIR and JC-10 dye. (A) shows the changes in the mitochondrial associated apoptosis in WPMY-1 cells based on JC-10 fluorescence intensity. Leptin enhanced the green fluorescence intensity of JC-10 aggregates, which means leptin downregulates the mitochondrial associated apoptosis. FACS analysis was carried out, analysing 10,000 cells per group. Data are mean ± SEM of five independent experiments. (***p < 0.001, ****p < 0.0001), compared to vehicle control by ordinary one-way ANOVA test. (B) reflects the activity of the ∆ψM in WPMY-1 cells based on NIR dye staining. After 24 h of incubation, leptin (25 ng/mL) increased the number of NIR-positive cells, indicating a protective effect on ∆ψM. Data are mean ± SEM of five independent experiments. (**p < 0.005), compared to vehicle control by Student’s t-test. |
We evaluated the effect of leptin on the ∆ψM of WPMY-1 using JC-10 and NIR assays. As shown in Figure 4A, leptin at 25 ng/mL significantly enhanced the JC-10 fluorescence intensity of WPMY-1 cells after 24 h and 48 h of exposure (p < 0.0001 for 24h and p = 0.0001 for 48 h). However, leptin at 50 ng/mL did not alter the JC-10 fluorescence intensity (p = 0.054 and p = 0.3623, respectively). Similarly, leptin at 25 ng/mL increased the ∆ψM of WPMY-1 cells in the NIR assay, as indicated by a higher NIR signal compared to the solvent control (210.7 ± 46.67 versus 164.0 ± 15.11, p = 0.0045) (Figure 4B). These results suggest that leptin enhances the mitochondrial function of prostate stromal cells.
Discussion
This study is the first to demonstrate the pivotal role of leptin in the etiology of MetS associated to BPH/LUTS, using both genomic and in vitro approaches. We have previously shown that leptin can enhance the contraction of prostatic smooth muscle tissue, thereby increasing smooth muscle tone and contributing to BPH development. Here, we identify the potential target genes that mediate this effect, leading to BPH pathogenesis. The target genes that were identified in our study through gene expression profiling using micro-array technology, have provided us with a comprehensive understanding of the molecular mechanisms underlying the involvement of leptin in the development of BPH. The differentially expressed or mutated genes in BPH patients were analyzed and potential targets were identified. In our investigation, we employed a rodent model of BPH that was instigated through subcutaneous administration of testosterone propionate consequent to castration. Gene expression analysis of the prostate tissue was performed, and the target genes were found to be mainly associated with pathways related to proliferation, contraction, and mitochondrial function. Specifically, genes such as Tfrc and Adam10 were found to be linked to proliferation, and Alox12, Fgfbp1, and Ptcd2, were linked to mitochondrial function (Table 1). Based on the microarray analysis, the precise mechanism by which MetS contributes to BPH is yet to be fully understood; however, several hypotheses have been proposed. A possible mechanism by which MetS contributes to prostate diseases is through the disruption of mitochondrial function. This disruption may result in increased production of reactive oxygen species (ROS), which can cause oxidative damage to cellular components and trigger inflammatory responses. Furthermore, mitochondrial dysfunction may induce cellular senescence, a state of irreversible growth arrest that is associated with aging and cancer. Therefore, MetS may promote prostate cancer development by impairing mitochondrial homeostasis and inducing oxidative stress, inflammation, and senescence in the prostate tissue.20 The regulation of apoptosis, autophagy and hormone synthesis is among the essential functions performed by mitochondria. Mitochondrial dysfunction can arise due to a range of factors, including genetic mutations, exposure to environmental toxins, aging, and metabolic disturbances. In turn, this may disrupt the balance between ROS and antioxidant defences, leading to oxidative damage of macromolecules such as DNA, proteins, and lipids.21 Notably, oxidative stress can trigger inflammation and prompt prostatic stromal cells to secrete cytokines and growth factors, which, in turn, stimulate the growth of prostatic epithelial cells and enhance extracellular matrix deposition.22
Additionally, oxidative stress can drive cellular senescence, characterized by altered gene expression and definite growth arrest, leading to cellular aging and dysfunction. Senescent cells release pro-inflammatory and pro-fibrotic factors that further impede prostate function by promoting enlargement and fibrosis. As such, mitochondrial function is a crucial regulator of MetS and BPH progression in obese men, and targeting mitochondrial dysfunction holds promise as an intervention strategy to forestall or cure BPH and its complications. However, the evidences that alteration of mitochondrial function in MetS associated to BPH are missing.
We conducted an investigation to identify novel targets that link MetS and BPH. We found that leptin, a wildly reported mediator of MetS, modulates the growth and contraction of prostatic stromal cells in vitro and ex vivo. We also explored the molecular mechanisms underlying these effects and discovered that leptin alters the mitochondrial function and apoptosis of WPMY-1 cells. Our findings suggest that leptin may play a role in the pathogenesis of BPH and provide new insights for potential therapeutic interventions. Our results demonstrate that leptin promotes the proliferation of WPMY-1 cells and inhibits their apoptosis, mimicking the in vivo effects of leptin in the development of BPH. Additionally, leptin enhances the ∆ψM, indicating improved mitochondrial function. Moreover, leptin increases the contractility of WPMY-1 cells in culture with a contraction gel. Overall, our findings suggest that leptin may protect WPMY-1 cells by reducing mitochondrial apoptosis, enhancing proliferation, and inducing contraction ability.
Combining FACS, contraction gel assay, and organ bath experiments, we confirmed from both in the cellular and human tissue levels that leptin has an important impact on the development of BPH. We hypothesized that leptin influences BPH by modulating the mitochondrial function in prostate cells. We found that leptin increased the ∆ψM and reduced the mitochondrial related apoptosis in prostate cells, indicating a shift from cell death to survival.
Our data reveal that BPH up-regulates three genes encoding proteins with distinct roles in mitochondrial function: Alox12, Fgfbp1, and Ptcd2 (Table 1). Alox12 encodes arachidonate 12-lipoxygenase, an enzyme that oxidizes polyunsaturated fatty acids to generate lipid hydroperoxides and lipoxins. Knockout of Alox12 leads to the downregulation of the biomarkers of inflammatory and apoptosis signaling pathway in spinal cord injury.23
Fgfbp1 encodes fibroblast growth factor binding protein 1, a secreted protein that binds to and potentiates the activity of fibroblast growth factors on target cells. A recent study suggested that Fgfbp1 in bladder cancer tissue could be identified as a promising biomarker for the accurate prediction of Bacillus Calmette-Guerin response in bladder cancer patients, which shows a clinical translate value for this target genes.24
The newly study of the pull-down assays indicated that the Ptcd2 protein was partially co-sedimented with its associated mitochondrial ribosome. The protein also displayed an association with various mitochondrial ribosome proteins, further attesting to its involvement in crucial mitochondrial functions. Ptcd2 encodes pentatricopeptide repeat domain-containing protein 2, a protein that modulates the processing and translation of mitochondrial RNA transcripts.25
Several studies have associated these three genes with various aspects of mitochondrial function. Alox12 regulates mitochondrial membrane potential, ROS generation, and apoptosis in cardiomyocytes. Fgfbp1 influences mitochondrial DNA replication, respiratory chain assembly, and energy metabolism in cancer cells. Ptcd2 controls the expression of cytochrome c oxidase subunit III (COIII), a key component of complex IV of the electron transport chain. However, The linkages between leptin and these target genes are not fully understood, which could a promising research topic in the future investigations.
We also show that leptin can influence the mitochondrial-mediated apoptosis pathway. These mechanisms may account for how leptin stimulates the growth of WPMY-1 cells and increases the smooth muscle contraction of the prostate, both of which are involved in BPH progression. Therefore, we suggest that targeting mitochondrial functions could be a valuable research direction to explore the role of leptin in BPH and to develop new treatments for this condition.
In this investigation, we evaluated the impact of leptin on mitochondrial function and mitochondrial associated apoptosis in WPMY-1. Our findings showed that leptin administration increased the ∆ψM, indicating improved mitochondrial energy metabolism and function. Additionally, we observed that leptin induced proliferation and contraction of WPMY-1 cells, corroborated by enhanced cell viability, EdU incorporation, and contractile force. We hypothesize that leptin may facilitate the increase in the ∆ψM via mitochondrial signaling pathways, possibly involved in augmenting smooth muscle tone in BPH. Hence, we propose that leptin can simultaneously trigger proliferation and contraction during the progression and development of BPH.
Leptin demonstrates its capacity to stimulate the proliferation of prostatic stromal cells, which play a crucial role in providing structural and functional support to the prostate gland, as well as the contraction of smooth muscle cells involved in urinary control. These findings suggest that leptin exerts its influence through a shared signaling pathway that governs both cell proliferation and contraction within the prostate gland. Given the common risk factors of obesity and inflammation shared by BPH and MetS, targeting leptin activity may yield beneficial effects in managing both conditions. Moreover, in overweight males, leptin could serve as a potential biomarker for BPH, and the development of pharmacological inhibitors targeting leptin signaling holds promise as a therapeutic approach for BPH treatment.
Furthermore, we postulate that leptin’s activation of different target genes is concentration-dependent, leading to either inhibition or stimulation of cell proliferation, as supported by existing literature.26 In human myometrial cells, low concentrations of leptin appear to induce a signaling pathway that suppresses cell growth, while high concentrations promote cell growth by up-regulating inflammation- and mitochondrial function-related signaling pathways and down-regulating inhibitory genes. Consequently, the overall effect of high concentrations of leptin is an augmentation of cell proliferation.27
Our findings reveal that leptin stimulates the proliferation of prostate stromal cells and the contraction of prostate smooth muscle cells in vitro, which are both involved in MetS associated to BPH. These mechanisms may increase prostatic volume and urethral resistance, leading to BPH/LUTS. We propose that BPH/LUTS should be considered as part of a systemic disorder that affects multiple pathways. Therefore, pharmacological interventions that target multiple pathways simultaneously may offer better clinical outcomes. We suggest that a single agent that can inhibit leptin-induced cell growth and relax smooth muscle tone could be a potential novel therapeutic strategy for BPH/LUTS.
This article is subject to certain limitations that must be considered when interpreting the findings presented. For instance, a potential limitation is the relatively small number of animal models used in the experiments, which may not provide adequate representation of the full range of treatment effects. Additionally, the patients’ sample may have been influenced by medication usage prior to surgeries, potentially introducing bias into the results. Confounding due to medication use is particularly present in clinical studies involving patients with BPH and MetS, as they are often treated with several medications over extended periods. In effect, some medications may either mask or amplify the effects of the treatment by providing anti-inflammatory or analgesic effects. We address these limitations by increasing the number of biological repeats in order to overcome potential issues encountered in organ bath. Through the addition of biological repeats, statistical power and accuracy are increased while random error and data noise are minimized, leading to more robust and consistent results that support the given hypothesis.
Conclusion
The role of leptin in promoting smooth muscle contraction and stromal cell growth in the human prostate represents a potential aggravating factor for the occurrence of LUTS and BPH in obese male patients. The upregulation of leptin in MetS has been implicated in the modulation of mitochondrial function and the activation of genome targets associated with both contraction and proliferation processes in the human prostate. These mechanisms are thought to contribute to the pathogenesis of BPH. Given the widely acknowledged association between metabolic syndrome and BPH/LUTS, the therapeutic management of leptin may prove beneficial for this category of patients. Further investigation into the leptin system in the context of BPH/LUTS is warranted.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This analysis and/or the underlying studies had been funded by the Li Huanying Foundation of Beijing, 2020, NO.PYZ201503 and the China Scholarship Council (grant 201606270233 (WXL)). Both funding sources were not involved in study design, in collection, analysis and interpretation of data, in the writing of the report, or in the decision to submit the article for publication.
Disclosure
All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Noubiap JJ, Nansseu JR, Lontchi-Yimagou E, et al. Global, regional, and country estimates of metabolic syndrome burden in children and adolescents in 2020: a systematic review and modelling analysis. Lancet Child Adolesc Health. 2022;6(3):158–170. doi:10.1016/S2352-4642(21)00374-6
2. Ranasinghe P, Mathangasinghe Y, Jayawardena R, Hills AP, Misra A. Prevalence and trends of metabolic syndrome among adults in the Asia-pacific region: a systematic review. BMC Public Health. 2017;17(1):101. doi:10.1186/s12889-017-4041-1
3. Ngai HY, Yuen KS, Ng CM, Cheng CH, Chu SP. Metabolic syndrome and benign prostatic hyperplasia: an update. Asian J Urol. 2017;4(3):164–173. doi:10.1016/j.ajur.2017.05.001
4. Hammarsten J, Peeker R. Urological aspects of the metabolic syndrome. Nat Rev Urol. 2011;8(9):483–494. doi:10.1038/nrurol.2011.112
5. Cao D, Sun R, Peng L, et al. Immune cell proinflammatory microenvironment and androgen-related metabolic regulation during benign prostatic hyperplasia in aging. Front Immunol. 2022;13:842008. doi:10.3389/fimmu.2022.842008
6. Yu Q, Wu C, Chen Y, et al. Inhibition of LIM kinase reduces contraction and proliferation in bladder smooth muscle. Acta Pharm Sin B. 2021;11(7):1914–1930. doi:10.1016/j.apsb.2021.01.005
7. Yu Q, Gratzke C, Wang Y, et al. Inhibition of prostatic smooth muscle contraction by the inhibitor of G protein-coupled receptor kinase 2/3, CMPD101. Eur J Pharmacol. 2018;831:9–19. doi:10.1016/j.ejphar.2018.04.022
8. Wang X, Li B, Ciotkowska A, et al. Onvansertib, a polo-like kinase 1 inhibitor, inhibits prostate stromal cell growth and prostate smooth muscle contraction, which is additive to inhibition by alpha(1)-blockers. Eur J Pharmacol. 2020;873:172985. doi:10.1016/j.ejphar.2020.172985
9. Steurer S, Hager B, Buscheck F, et al. Up regulation of Rho-associated coiled-coil containing kinase1 (ROCK1) is associated with genetic instability and poor prognosis in prostate cancer. Aging (Albany NY). 2019;11(18):7859–7879. doi:10.18632/aging.102294
10. Pico C, Palou M, Pomar CA, Rodriguez AM, Palou A. Leptin as a key regulator of the adipose organ. Rev Endocr Metab Disord. 2022;23(1):13–30. doi:10.1007/s11154-021-09687-5
11. Poetsch MS, Strano A, Guan K. Role of leptin in cardiovascular diseases. Front Endocrinol (Lausanne). 2020;11:354. doi:10.3389/fendo.2020.00354
12. Ray A, Cleary MP. The potential role of leptin in tumor invasion and metastasis. Cytokine Growth Factor Rev. 2017;38:80–97. doi:10.1016/j.cytogfr.2017.11.002
13. Philp LK, Rockstroh A, Sadowski MC, et al. Leptin antagonism inhibits prostate cancer xenograft growth and progression. Endocr Relat Cancer. 2021;28(5):353–375. doi:10.1530/ERC-20-0405
14. Gorrab A, Pagano A, Ayed K, et al. Leptin promotes prostate cancer proliferation and migration by stimulating STAT3 pathway. Nutr Cancer. 2021;73(7):1217–1227. doi:10.1080/01635581.2020.1792946
15. Noblet JN, Goodwill AG, Sassoon DJ, Kiel AM, Tune JD. Leptin augments coronary vasoconstriction and smooth muscle proliferation via a Rho-kinase-dependent pathway. Basic Res Cardiol. 2016;111(3):25. doi:10.1007/s00395-016-0545-6
16. Genchi VA, D’Oria R, Palma G, et al. Impaired leptin signalling in obesity: is leptin a new thermolipokine? Int J Mol Sci. 2021;22(12):6445. doi:10.3390/ijms22126445
17. Wang X, Wang Y, Gratzke C, et al. Ghrelin aggravates prostate enlargement in rats with testosterone-induced Benign prostatic hyperplasia, stromal cell proliferation, and smooth muscle contraction in human prostate tissues. Oxid Med Cell Longev. 2019;2019:4748312. doi:10.1155/2019/4748312
18. Saha PC, Chatterjee T, Pattanayak R, et al. Targeting and imaging of mitochondria using near-infrared cyanine dye and its application to multicolor imaging. ACS Omega. 2019;4(11):14579–14588. doi:10.1021/acsomega.9b01890
19. Younes N, Alsahan BS, Al-Mesaifri AJ, et al. JC-10 probe as a novel method for analyzing the mitochondrial membrane potential and cell stress in whole zebrafish embryos. Toxicol Res (Camb). 2022;11(1):77–87. doi:10.1093/toxres/tfab114
20. Kowalczyk P, Sulejczak D, Kleczkowska P, et al. Mitochondrial oxidative stress-a causative factor and therapeutic target in many diseases. Int J Mol Sci. 2021;22(24):13384. doi:10.3390/ijms222413384
21. Juan CA, Perez de la Lastra JM, Plou FJ, Perez-Lebena E. The chemistry of Reactive Oxygen Species (ROS) revisited: outlining their role in biological macromolecules (DNA, Lipids and Proteins) and induced pathologies. Int J Mol Sci. 2021;22(9):4642. doi:10.3390/ijms22094642
22. Naiyila X, Li J, Huang Y, et al. A novel insight into the immune-related interaction of inflammatory cytokines in Benign prostatic hyperplasia. J Clin Med. 2023;12(5):1821. doi:10.3390/jcm12051821
23. Li JL, Liang YL, Wang YJ. Knockout of ALOX12 protects against spinal cord injury-mediated nerve injury by inhibition of inflammation and apoptosis. Biochem Biophys Res Commun. 2019;516(3):991–998. doi:10.1016/j.bbrc.2019.06.118
24. Li F, Zhang H, Wang Y, et al. FGFBP1 as a potential biomarker predicting bacillus Calmette-Guerin response in bladder cancer. Front Immunol. 2022;13:954836. doi:10.3389/fimmu.2022.954836
25. Baleva MV, Chicherin I, Piunova U, et al. Pentatricopeptide protein PTCD2 regulates COIII translation in mitochondria of the HeLa cell line. Int J Mol Sci. 2022;23(22):14241. doi:10.3390/ijms232214241
26. Blanquer-Rossello MM, Santandreu FM, Oliver J, Roca P, Valle A. Leptin modulates mitochondrial function, dynamics and biogenesis in MCF-7 cells. J Cell Biochem. 2015;116(9):2039–2048. doi:10.1002/jcb.25158
27. Noda T, Kikugawa T, Tanji N, et al. Long‑term exposure to leptin enhances the growth of prostate cancer cells. Int J Oncol. 2015;46(4):1535–1542. doi:10.3892/ijo.2015.2845
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.