Back to Journals » Journal of Pain Research » Volume 19
LC-IL but Not LC-PL Circuit Regulates Comorbid Mechanical Hyperalgesia and Anxiety in Male Rats with Chronic Post-Surgical Pain
Authors Lin S ![]() , Liu C, Jiang X, Lin M, Wang F, Zhuo Y, Lin K, Lin C, Liu R
, Liu C, Jiang X, Lin M, Wang F, Zhuo Y, Lin K, Lin C, Liu R
Received 11 January 2026
Accepted for publication 1 July 2026
Published 10 July 2026 Volume 2026:19 585416
DOI https://doi.org/10.2147/JPR.S585416
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Alaa Abd-Elsayed
Shenshen Lin,1,2,* Congxu Liu,3,* Xiaoling Jiang,4,* Minyi Lin,2 Fengchai Wang,2 Yanmei Zhuo,2 Kaixing Lin,2 Chun Lin,5 Rongguo Liu1,6
1Department of Pain Management, the First Affiliated Hospital of Fujian Medical University, Fuzhou, Fujian, People’s Republic of China; 2Department of Pain Management, the First Hospital of Putian City, Putian, Fujian, People’s Republic of China; 3Pathology Department of Quangang District Hospital, Quanzhou, Fujian, People’s Republic of China; 4Department of Anesthesiology, Affiliated Hospital of Putian University, Putian, Fujian, People’s Republic of China; 5Pain Research Institute, School of Basic Medical Sciences, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China; 6Department of Pain Management, National Regional Medical Center, Binhai Campus of the First Affiliated Hospital, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chun Lin, Pain Research Institute, School of Basic Medical Sciences, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China, Email [email protected] Rongguo Liu, Department of Pain Management, The First Affiliated Hospital of Fujian Medical University, Fuzhou, Fujian, People’s Republic of China, Email [email protected]
Objective: This study investigated the differential roles of Locus Coeruleus-Infralimbic cortex (LC-IL) and Locus Coeruleus-Prelimbic cortex (LC-PL) circuits in comorbid mechanical hyperalgesia and anxiety-like emotions in male rats with chronic post-surgical pain (CPSP).
Methods: A total of 100 adult male Sprague-Dawley rats were randomly divided into 10 groups (n=10/group). Group included LC activation (sham+ChR2, sham+mch), LC–IL inhibition/activation (CPSP+NpHR, CPSP+mch; sham+ChR2, sham+mch), and LC–PL inhibition/activation (CPSP+NpHR, CPSP+mch; sham+ChR2, sham+mch). Using the skin/muscle incision and retraction procedure to establish CPSP or sham surgery. Optogenetic viruses (ChR2 for activation, NpHR for inhibition) or control virus (mCherry) were injected into the LC, with optical fibers implanted in the LC, IL, or PL. After 4 weeks of viral expression, blue or yellow light stimulation was applied. Mechanical paw withdrawal threshold was measured by Von-Frey filaments. Anxiety-like emotions was assessed by open field test and elevated plus maze.
Results: Blue light irradiation of the LC region and LC-IL neural circuit both induced mechanical hyperalgesia and anxiety-like emotions in the rats. Yellow light irradiation of the LC-IL neural circuit alleviated mechanical hyperalgesia and anxiety-like emotions in CPSP model rats. Blue or yellow light irradiation of the LC-PL neural circuit had no significant effect on mechanical hyperalgesia or anxiety-like emotions in the rats.
Conclusion: The LC-IL circuit specifically regulates comorbid mechanical hyperalgesia and anxiety-like emotions in male CPSP rats, while the LC-PL circuit does not. These findings identify a sex-specific circuit mechanism and a potential therapeutic target for CPSP.
Keywords: locus coeruleus, medial prefrontal cortex, infralimbic cortex, prelimbic cortex, chronic post-surgical pain
Introduction
With the aging population and the increasing number of surgical procedures performed, chronic post-surgical pain (CPSP) has become one of the increasingly severe health issues.1,2 According to the international classification of diseases 11th revision (ICD-11), CPSP is defined as pain at the surgical site that persists for more than 3 months after surgical wound healing and can not be explained by other causes such as infection, malignancy, or pre-existing pain.3–5 The incidence of CPSP is as high as 20% after surgeries such as thoracic surgery, inguinal hernia repair, lumbar spine surgery, and hip or knee replacement;6 persistent pain after trauma and burn surgeries is also common.7 CPSP is often accompanied by psychological distress such as anxiety and depression,8,9 which severely impairs patients’ post-surgical quality of life.
Most drugs used clinically for acute pain can not effectively prevent the development of CPSP and may lead to excessive use of opioids;10 currently, there are no effective methods for the treatment or prevention of CPSP. Therefore, clarifying the pathogenesis of CPSP comorbid with anxiety-like emotions and exploring effective therapeutic strategies are of great importance.
The locus coeruleus (LC) is the main source of norepinephrine in the brain. Norepinephrine, a neurotransmitter released by the LC, primarily acts on cells and circuits in the forebrain, brainstem, cerebellum, and spinal cord.11–13 Studies have shown that the LC can be activated when the body is exposed to stress stimuli, leading to increased synthesis and secretion of norepinephrine.14,15 During acute pain, stimulating the LC exerts an analgesic effect; however, during chronic pain, activation of the LC can induce anxiety-like emotions.16,17 Nevertheless, whether the LC regulates chronic pain and anxiety-like emotions in CPSP requires further investigation.
The LC has a unique neural projection pattern and coding characteristics, projecting to a wide range of brain regions including the forebrain, brainstem, cerebellum, and medulla oblongata.18,19 The projection circuits of the LC, by releasing norepinephrine, are involved in regulating numerous bodily functions such as arousal, stress, emotion, learning, pain, decision-making, and perception.20–22 For example, projections from the LC to the amygdala and anterior cingulate cortex (ACC) circuits are involved in regulating anxiety or aversion;23–25 projections from the LC to the spinal cord produce analgesic effects;26–28 and projection circuits from the LC to the medial prefrontal cortex (mPFC) are involved in the formation and extinction of fear memory.29 These findings suggest that different LC projection circuits act on different brain regions: although the main neurotransmitter released is norepinephrine, the regulated functions vary depending on the circuit and brain region.Both the LC and mPFC are involved in the regulation of pain and emotion,30,31 but whether the LC-mPFC projection circuit participates in the regulation of comorbid mechanical hyperalgesia and anxiety-like emotions in CPSP remains unclear, which warrants investigation.
The mPFC consists of three subregions: the ACC, prelimbic cortex (PL), and infralimbic cortex (IL). The role of the ACC subregion in regulating pain and emotion has been clearly established in previous studies; therefore, this study focused on exploring the roles of the IL and PL subregions in regulating mechanical hyperalgesia and anxiety-like emotions in CPSP. The IL exerts antinociceptive/inhibitory effects on pain hypersensitivity, primarily contributing to pain relief and the extinction of fear/anxiety-related behaviors.32 It modulates the descending pain inhibitory pathway via projections to the ventromedial periaqueductal gray, attenuating hyperalgesia and allodynia, additionally, the IL is critical for regulating emotional valence.33,34 The PL is predominantly involved in encoding the affective-motivational dimension of pain, and drives central sensitization and hyperalgesia in chronic pain states.35,36 In terms of emotional regulation, the PL is specialized in mediating anxiety-like behavioral expression, fear acquisition, and the amplification of negative affective states.37,38 Anatomically, the PL receives dense noradrenergic innervations from the LC, and LC-derived norepinephrine release activates PL pyramidal neurons to exacerbate pain hypersensitivity and comorbid anxiety.39,40 This discrete functional profile distinguishes the PL from the IL, which primarily mediates pain inhibition and fear extinction.Notably, the LC noradrenergic system projects differentially to the PL and IL, exerting distinct regulatory effects on pain hypersensitivity and comorbid anxiety. Whether the LC-IL and LC-PL neural circuits exhibit specificity in regulating mechanical hyperalgesia and comorbid anxiety-like emotions in CPSP remains unclear.
In summary, we hypothesize that the specific LC–IL and LC–PL neural circuits within the LC-mPFC network are differentially modulated under CPSP conditions, thereby regulating mechanical hyperalgesia and anxiety-like emotions, and that this regulation may exhibit specificity for the LC-IL and LC-PL neural circuits. Therefore, in this study, the CPSP rat model was established using the skin/muscle incision and retraction (SMIR) procedure described by Flatters. Taking the LC as the entry point, we used optogenetics combined with behavioral and molecular biological methods to explore the regulatory roles of the LC-IL and LC-PL neural circuits (within the LC brain region) in mechanical pain sensitivity and anxiety-like emotions in CPSP rats, aiming to provide new research evidence for the clinical treatment of CPSP. Anatomical validations including viral injection sites, optical fiber placement, and axon terminal expression in the IL and PL were performed to ensure pathway specificity. Notably, viral vectors were injected into the LC to target noradrenergic neurons, and optical stimulation was applied at the axon terminals of LC→IL and LC→PL projections rather than directly activating or inhibiting local IL or PL neurons.
To our knowledge, this is the first study to investigate the effects of the LC-IL and LC-PL neural circuits on mechanical hyperalgesia and comorbid anxiety-like emotions in CPSP, which has certain innovativeness. Considering the well-established sex differences in chronic postsurgical pain, locus coeruleus-noradrenergic signaling, prefrontal anxiety circuits, and pain-anxiety comorbidity, this study was performed only in male rats as an exploratory mechanistic study.41–43 Subsequent studies in female animals are required to verify the universality of the present results.
Materials and Methods
Materials
Experimental Animals
Specific-pathogen-free (SPF) male Sprague-Dawley (SD) rats, weighing 200–220 g, were provided by the Animal Center of Fujian Medical University. All animal experiments were approved by the Laboratory Animal Ethics Committee of Fujian Medical University (FJMU-IACUC-2023-Y-1044). The animal use license number is SYXK (Min) 2022–0003. All procedures were performed in accordance with the Guidelines for Ethical Review of Welfare of Laboratory Animals.
Animals undergoing surgical modeling and oral administration were housed individually to prevent mutual biting of wounds (which could cause infection) and to control drug dosage. The five basic welfare principles for experimental animals were followed: the animal housing room was equipped with a good ventilation and air filtration system; animals were housed under a natural light cycle, with room temperature maintained at approximately26°Cand humidity at approximately 55%; food and water were provided ad libitum. The experimental protocol was optimized to minimize the number of animals in accordance with the “3R” principles (Replacement, Reduction, Refinement) to avoid unnecessary sacrifice and waste. During the experiment, efforts were made to minimize pain caused to the animals. All procedures strictly complied with national policies and regulations on experimental animals, and all animal operations were performed by trained personnel in accordance with standard operating procedures.
Main Drugs, Reagents, and Materials
10μL microsyringes (Hamilton, Switzerland), Von-Frey filaments (Aesthesio, USA), Animal behavior testing platform (RWD Life Science Co., Ltd., Shenzhen, China), Pipettes of various specifications (Eppendorf, Germany),Viral vectors: rAAV-hSyn-ChR2(H134R)- mCherry- WPRE -hGH polyA (serotype9, ChR2), rAAV-hSyn-mCherry-WPRE-hGH polyA (serotype9, mch), and rAAV-hSyn-eNpHR3.0-mCherry -WPRE-hGH polyA (serotype 9, NpHR), scAAV-hSyn-EGFP-WPREs(retroserotype,PT-3975), (BrainVTA Co., Ltd., Wuhan, China).
Methods
Experimental Design and Animal Grouping
A total of 100 adult male Sprague-Dawley rats were randomly divided into 10 groups (n=10/group). Group included LC activation (sham+ChR2, sham+mch), LC–IL inhibition/activation (CPSP+NpHR, CPSP+mch; sham+ChR2, sham+mch), and LC–PL inhibition/activation (CPSP+NpHR, CPSP+mch; sham+ChR2, sham+mch) (Table 1).
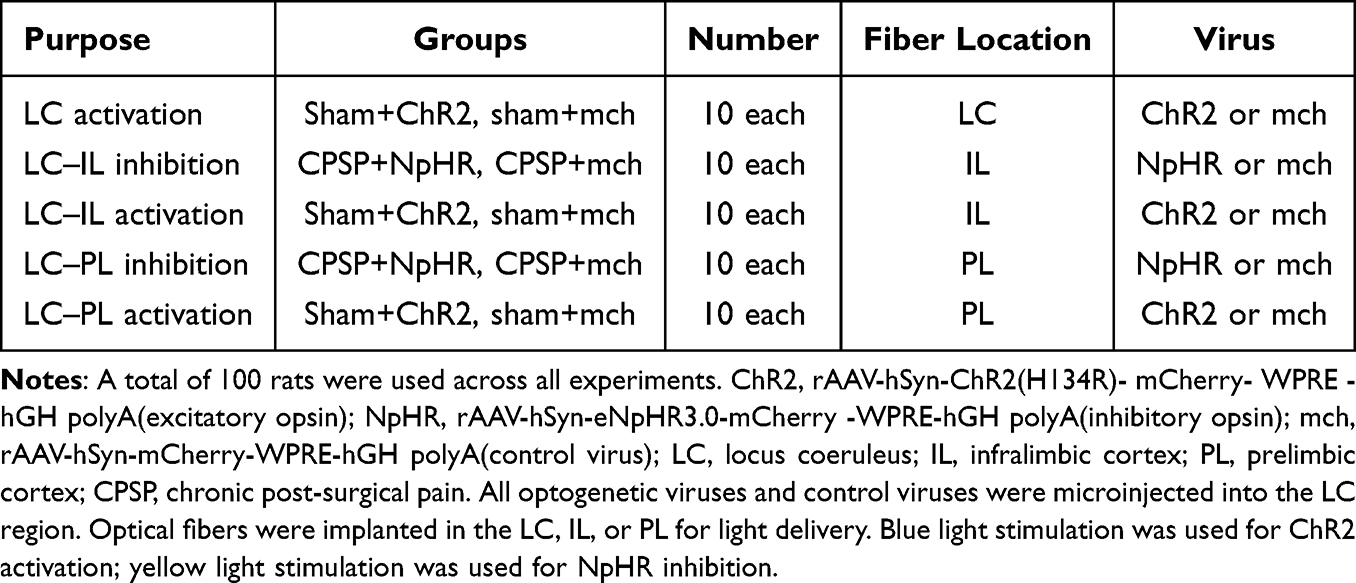 |
Table 1 Experimental Design and Group Structure for Optogenetic Manipulations |
Rats were randomly allocated to experiments by a technician not involved in behavioral assessment, using a random number generator. Baseline mechanical thresholds and body weight were balanced across groups (stratified randomization).
Single-blinding was strictly performed during behavioral testing and data analysis. All animals were coded with unique numbers, and group information was kept confidential by an independent researcher. The investigators who conducted behavioral tests and data collection were completely blind to the experimental grouping. Data were pooled and decoded only after all statistical analyses were completed.
Establishment of the Chronic Post-Surgical Pain (CPSP) Rat Model
The CPSP model was established using the skin/muscle incision and retraction (SMIR) procedure described by Flatters. For the sham-operated group (sham), all procedures were identical to those of the CPSP group except that no retraction was performed. The specific procedure was as follows: Male SD rats (200–220 g) were fasted for 12 hours before surgery with free access to water, all surgical instruments were autoclaved. The operating room temperature was maintained at 25°C. Rats were placed in the supine position and fixed on a disposable surgical drape, then connected to a small animal-specific gas anesthetic machine. The oxygen flow rate and isoflurane concentration were adjusted to maintain an appropriate depth of anesthesia.
After anesthesia took effect, the hair in the right surgical field was removed using an animal hair clipper. The hind limbs were abducted and fixed to the operating table with medical tape; the rat’s head was turned to one side, and the tongue was pulled out to prevent asphyxiation. The surgical field was disinfected to clearly visualize the saphenous nerve, and a sterile surgical drape with a hole was applied.
The skin on the medial side of the right hind paw was stretched taut. A sterile scalpel was used to make a 1.5–2 cm longitudinal skin incision 4 mm medial to the saphenous vein in the middle of the thigh to expose the leg muscles. A 7–10 mm incision was then made in the superficial muscle (gracilis muscle). Blunt-tipped scissors were used to further bluntly dissect the superficial muscle at the incision site; after the white fascia of the adductor muscle group was visible, a micro-retractor (with four prongs spaced 8 mm apart, each 4 mm in length) was inserted into the incision. The skin and superficial muscles were retracted to 2 cm to expose the underlying adductor muscle fascia, and retraction was maintained for 1 hour. During retraction, care was taken to avoid damaging the saphenous nerve (which could cause neuropathic pain).
During retraction, the incision site was covered with sterile normal saline gauze to maintain moisture and temperature, preventing incision dryness caused by prolonged evaporation (which might affect the role of skin incision and retraction in inducing chronic post-surgical hyperalgesia). After 1 hour of retraction, the muscle and skin were sutured layer by layer using sterilized 4–0 absorbable sutures and 3–0 silk sutures, respectively. Penicillin (80,000 U) was intraperitoneally injected for 3 consecutive days after surgery to prevent post-surgical infection. After the rats regained consciousness naturally, they were transferred to the animal room and housed individually. During surgery, the rat’s breathing and physiological status were closely monitored to maintain an appropriate depth of anesthesia.
Stereotaxic Viral Injection and Fiber Implantation in the Brain
Stereotaxic Localization of the Locus Coeruleus (LC)
All surgeries were performed under sterile conditions. Rats were fasted for 12 hours before the experiment with free access to water. After weighing, they were connected to a small animal-specific gas anesthetic machine. Deep anesthesia was induced with isoflurane before surgery (criteria for adequate anesthesia: slow and deep breathing, reduced corneal reflex, loss of limb tone, and no body twitching when the tail was pinched with forceps, indicating that the rat was in a state of deep anesthesia).
After the rat was anesthetized, the hair in the surgical area was removed with a hair clipper. The rat was placed in the prone position and fixed on a rat stereotaxic instrument: the root of the upper incisors was fixed to the incisor hook, and ear bars were inserted into the external auditory canals on both sides. The knobs were tightened, and the stability of the rat’s head fixation was checked.
A scalpel was used to make a longitudinal skin incision along the midline of the rat’s cranial roof. The skin, subcutaneous tissue, and deep fascia were incised sequentially to expose the skull, and the periosteum was peeled off to expose the bregma and lambda for localization. For viral transfection of LC nucleus neurons, a hole with a diameter of approximately 2 mm was drilled in the skull at the LC location (AP: −9.8 mm, RL:±1.4 mm, DV: −7.2 mm) according to the Paxinos & Watson Rat Brain Atlas.
A 10μL microsyringe(RWD) was used to inject 0.5μL virus at a rate of 0.030μL/min, the viral titer was 5.25×1012 vg/mL. After injection, the needle was raised by 0.1 mm and held in place for the same duration as the injection to allow sufficient diffusion of the virus at the injection site. After the microsyringe was slowly withdrawn, an optical fiber was implanted at the same location as the viral injection. Rats in the optogenetic virus group received unilateral injections.
Viral Transfection of LC-IL and LC-PL Neural Circuits
According to the Paxinos & Watson Rat Brain Atlas, optogenetic viruses were injected into the LC, and optical fibers were implanted into the ipsilateral IL (AP: 3.0 mm, RL:±0.6 mm; DV: −4.7 mm) or PL (AP: 3.0 mm, RL:±0.6 mm; DV: −3.2 mm). The viral injection rate and post-injection waiting time were the same as described above.
After the above steps were completed, screws were used to fix the head of rats that underwent stereotaxic brain surgery. Dental cement was used to fill the gaps between the optical fibers, screws, and the skull surface to fix the optical fibers to the rat’s head. Rats were housed individually after stereotaxic surgery. Penicillin (200,000 units per rat) was injected for 3 consecutive days post-surgery, and rats were allowed a 2-week recovery period before proceeding to the next experiment.
Pain Behavioral Testing in Rats
The 50% paw withdrawal threshold (PWT) was used to reflect changes in the rat’s pain threshold. Before behavioral testing, rats were acclimated to the testing environment to enhance their adaptability. Rats were placed in the testing environment for 2–3 acclimation sessions one week before testing, with 30 minutes of acclimation per day, to reduce the impact of emotional responses caused by the unfamiliar environment on test results.
The mechanical withdrawal threshold was measured in all rats before modeling (as the baseline value) and repeated on days 1, 3, 7, 12, 14, 21, and 28 after modeling.
Determination of Mechanical Paw Withdrawal Threshold:Von-Frey filaments of different intensities were used to measure the mechanical withdrawal threshold using the“up-downmethod”. Rats were placed in Plexiglas compartments on a metal mesh platform. After 30 minutes of acclimation, testing was initiated with a 2 g filament, and stimulation was applied for no more than 4 seconds. A positive response (recorded as“1”) was defined as a rapid paw withdrawal or paw licking; a negative response (recorded as “0”) was defined as the absence of such behavior.
During testing: if a negative response was observed, the intensity of the filament was increased; if a positive response was observed, the next lower intensity filament was used. The maximum intensity was 15 g, and any response exceeding this value was recorded as 15 g. The interval between two tests was at least 1 minute to avoid frequent testing affecting the results. During each measurement, the filament was applied vertically to the plantar surface of the rat’s surgical-side hind paw with a force sufficient to bend the filament by 90 degrees, ensuring consistent stimulation intensity each time.
Testing was terminated under the following conditions: A negative response was observed even with the maximum intensity of the Von-Frey filament; After the first positive response, the up-down method was continued for 4 additional measurements;The total number of measurements using the up-down method reached 9. After testing, the withdrawal response pattern (in grams) was converted to the corresponding 50% mechanical withdrawal threshold using the appropriate formula.
Negative Emotion Testing in Rats
Open Field Test (OFT)
The experimental apparatus consisted of two parts: an open field reaction box and an automatic data collection and processing system. The open field apparatus for rats had a height of 60 cm and a base length of 100 cm, with black inner walls. A digital camera was mounted 1.2 m directly above the apparatus, with a field of view covering the entire interior of the open field.
Experimenters, computers, and other equipment were placed in a separate room to minimize interference with the animals. Background noise in the laboratory was controlled below 65 dB. Animals were acclimated to the experimental environment for 1 hour before testing.
During each test, the rat was gently placed at the center of the base of the square box, holding the tail at 2/3 of its length from the root. Video recording and timing were initiated simultaneously, and the rat’s activity was observed for 5 minutes via the monitor. The total distance traveled (cm), central distance traveled (cm), and central time spent (s) were recorded for each rat. Recording was stopped after 5 minutes.
Between tests, the experimental apparatus was cleaned with a 10% ethanol solution to eliminate residual odor from previous animals (which might affect subsequent test results). The next rat was tested only after the ethanol had evaporated and the open field was dry. After all rats were tested, the videos were analyzed using an autonomous activity video analysis system.
Elevated Plus-Maze Test (EPM)
The EPM is a classic animal anxiety behavior test that uses the conflict between an animal’s exploratory behavior in a novel environment and its fear of elevated open arms to assess the animal’s anxiety state.
The testing environment was kept quiet, and rats were acclimated to the environment for 1 hour before testing. At the start of the experiment, the rat was placed in the central area of the maze facing a closed arm. A video tracking software was used to record the rat’s activity for 5 minutes.
In the EPM, the percentage of open arm entries and the percentage of time spent in open arms were used as indicators of anxiety-like behavior in rats. These two indicators were negatively correlated with anxiety levels: fewer entries into open arms and shorter time spent in open arms indicated more severe anxiety.
Between tests, the experimental apparatus was cleaned with a 10% ethanol solution to eliminate residual odor from previous animals (which might affect subsequent test results). The next rat was tested only after the apparatus was dry. Video analysis was performed using the same system as in the OFT.
Optogenetic Experiments
Optogenetic Activation Experiment
Before the experiment, rats were randomly divided into two groups (n=10,each): one group received injection of ChR2-transfected adenovirus, and the other received mch-transfected adenovirus. After sufficient viral expression, the cannula insert in the rat’s brain was connected to an optical fiber patch cord with a rotary joint, which was linked to a 473 nm wired optogenetic system. This setup allowed the rat to move freely while receiving blue light stimulation, thereby activating LC nucleus neurons or the LC-IL/LC-PL neural circuits.
The light stimulation parameters were 15 Hz, 20 ms, and 20 mW. For activation of LC nucleus neurons, blue light stimulation was applied continuously for 5 minutes, with the laser alternating between 4 seconds on and 6 seconds off. Blue light pulses were delivered to the LC via the implanted cannula, and changes in mechanical pain sensitivity and anxiety-like negative emotions in the rats were observed.
For activation of the LC-IL and LC-PL neural circuits, blue light stimulation was applied continuously for 30 minutes per day for 3 consecutive days, with the laser alternating between 4 seconds on and 6 seconds off. Blue light pulses were delivered to the PL and IL subregions via the implanted cannula, and changes in mechanical pain sensitivity and anxiety-like negative emotions in the rats were observed.
After the experiment, brain tissues were collected for anatomical verification of the light activation site, and data from activation outside the target region were excluded.
Optogenetic Inhibition Experiment
Before the experiment, rats were randomly divided into two groups (n=10,each): one group received injection of NpHR-transfected adenovirus, and the other received mch-transfected adenovirus. After viral injection, rats were allowed a 2-week rest period before CPSP model establishment.
After sufficient viral expression, the cannula insert in the rat’s brain was connected to an optical fiber patch cord with a rotary joint, which was linked to a 473 nm wired optogenetic system. This setup allowed the rat to move freely while receiving yellow light stimulation, thereby inhibiting the LC-IL/LC-PL neural circuits.
The light stimulation parameters were 15 Hz, 20 ms, and 20 mW. Yellow light stimulation was applied continuously for 30 minutes per day for 3 consecutive days, with the laser alternating between 4 seconds on and 6 seconds off. Yellow light pulses were delivered to the IL and PL subregions of the prefrontal cortex via the implanted cannula, and changes in mechanical pain sensitivity and anxiety-like negative emotions in the rats were observed.
After the experiment, brain tissues were collected for anatomical verification of the light inhibition site, and data from inhibition outside the target region were excluded.
Frozen Sectioning
Perfusion and Tissue Collection
Perfusion: Rats were anesthetized by intraperitoneal injection of 0.3% pentobarbital sodium at a dose of 50 mg/kg body weight (0.1 mL/10 g). Anesthesia was confirmed by loss of righting reflex and absence of withdrawal response to hind toe pinch. Then placed in the supine position, A“V”shaped incision was made in the abdominal wall skin of the upper abdomen, and the thoracic cavity was opened below the xiphoid process to expose the heart. A perfusion needle was inserted from the left ventricle to the root of the ascending aorta, and a hemostat was used to fix the perfusion needle to the heart. The perfusion system was connected, and a small incision was made in the right atrial appendage as the outflow port for the perfusion solution.
First, 4°Cnormal saline was perfused rapidly until the liver and lungs turned white and the fluid flowing out of the right atrial appendage became clear (indicating that blood in the blood vessels had been completely flushed out). Then, 500 mL of 4% paraformaldehyde was perfused: the first 100 mL was perfused rapidly, and the remaining 400 mL was perfused slowly. During paraformaldehyde perfusion, the rat exhibited generalized tremors; perfusion was completed when muscle tremors ceased and the limbs became rigid. The entire perfusion process lasted approximately 3 hours.
Tissue collection: Rats were euthanized by decapitation following pentobarbital sodium anesthesia and subsequent perfusion to minimize suffering,in accordance with applicable veterinary guidelines of the American veterinary medical association. Scissors were inserted into the foramen magnum to make a crack in the parietal bone, and hemostats were used to split the parietal bone to both sides. Care was taken to tear the membrane structure on the surface of the brain to avoid damaging the integrity of the brain tissue during collection. Successfully perfused brain tissue appeared pale and avascular.
The brain tissue was fixed in 4% paraformaldehyde at 4°Cfor 2 hours, then blotted gently with filter paper to remove excess fluid around the tissue. It was then placed in 20% sucrose and stored in a 4°Crefrigerator. After the tissue sank to the bottom, it was transferred to 30% sucrose for gradient dehydration. Dehydration was completed when the tissue sank to the bottom again, and the tissue was then ready for embedding.
Frozen Sectioning of Brain Tissue
Embedding: A uniform layer of OCT embedding medium was spread on the bottom of a mold. When the bottom of the medium turned slightly white, the tissue was placed horizontally in the mold (with the head-tail direction marked). OCT was then slowly poured into the mold to completely cover the tissue, avoiding air bubble formation. The mold was placed at −80°Cfor rapid freezing.
Sectioning: The tail end of the tissue block was placed on a cryostat stage and clamped to the specimen holder of the microtome. The position and angle of the blade were adjusted to obtain complete and uniform tissue sections. The angle and height of the anti-roll plate were adjusted to ensure flat sections. The thickness of the brain sections was set to 20μm.
Mounting: Adhesive slides were numbered sequentially. A brush was used to pick up the sections and mount them flat on the slides. If air bubbles were present, a small amount of PBS was dipped with the brush to expel the bubbles.
Section Selection: After mounting, sections with intact tissue were selected using an optical microscope and stored in a −20°Crefrigerator for long-term use.
Immunofluorescence
Confocal Immunofluorescence (IF)
Slide Washing: Tissue sections were rewarmed at room temperature for 30 minutes, then placed in PBS buffer and shaken gently (5 minutes per wash×2 washes), followed by washing in PBST (5 minutes×1 wash).
Blocking: Filter paper was used to gently blot excess fluid around the tissue. A hydrophobic pen was used to draw a circle around the tissue, and blocking solution was added dropwise to the tissue within the circle. Blocking was performed at room temperature in a humid chamber for 1 hour.
Primary Antibody Incubation: The blocking solution was gently blotted with filter paper, and diluted primary antibody against c-fos (1:500, Abcam) was added dropwise directly to the tissue. Incubation was performed in a humid chamber at 4°C for 48 hours.
Secondary Antibody Incubation: The primary antibody was gently blotted with filter paper. The sections were placed in PBS buffer and shaken gently (5 minutes per wash×2 washes), followed by washing in PBST (5 minutes×1 wash). Under dark conditions, diluted fluorescent secondary antibody (donkey anti-mouse, 1:500; donkey anti-rabbit, 1:500) was added dropwise, and incubation was performed in a humid chamber at room temperature for 2 hours.
DAPI Nuclear Staining: The secondary antibody was gently blotted with filter paper. The sections were placed in PBS buffer and shaken gently (5 minutes per wash×2 washes), followed by washing in PBST (5 minutes×1 wash). Under dark conditions, ready-to-use DAPI staining solution was added dropwise, and incubation was performed in a dark box for 5–10 minutes.
Mounting: The DAPI solution was gently blotted with filter paper. No additional washing was performed; anti-fluorescence quenching mounting medium was added dropwise directly to the tissue sections, and a coverslip was placed on top to completely cover the tissue without air bubbles. The sections were stored in a dark box at 4°Cuntil observation.
Imaging: A Leica SP5 laser confocal microscope was used for observation and imaging.
Statistical Analysis
All quantitative data are expressed as mean ± SEM. The primary analysis for behavioral outcomes (PWT, OFT, EPM measures) employed two-way repeated-measures ANOVA with group × time interaction (SPSS 26.0), followed by Bonferroni post-hoc tests for multiple comparisons. The between-subjects factor was group (eg., CPSP+NpHR vs. CPSP+mch), and the within-subjects factor was time (pre-stimulation vs. post-stimulation). For LC soma activation (single session), time points were pre, during, and post-stimulation. For terminal manipulations (3-day protocol), time points were baseline (pre-surgery), pre-stimulation (day 0), and post-stimulation (day 3).
Where sphericity was violated (Mauchly’s test p<0.05), Greenhouse-Geisser correction was applied. Effect sizes are reported as partial η2. For two-group comparisons at single time points, independent samples t-test (normal distribution, homogeneous variance) or Welch’s t-test (heterogeneous variance) was used. Normality was assessed by Shapiro–Wilk test. A p-value <0.05 was considered statistically significant. Exact p-values and 95% confidence intervals are reported for all key comparisons.
Results
Successful Induction of the Chronic Post-Surgical Pain (CPSP) Rat Model by SMIR Surgery
In this study, the CPSP rat model was induced using the SMIR procedure. The mechanical withdrawal threshold of the rat’s surgical-side hind paw was measured to evaluate the success of model establishment (Figure 1A and B).
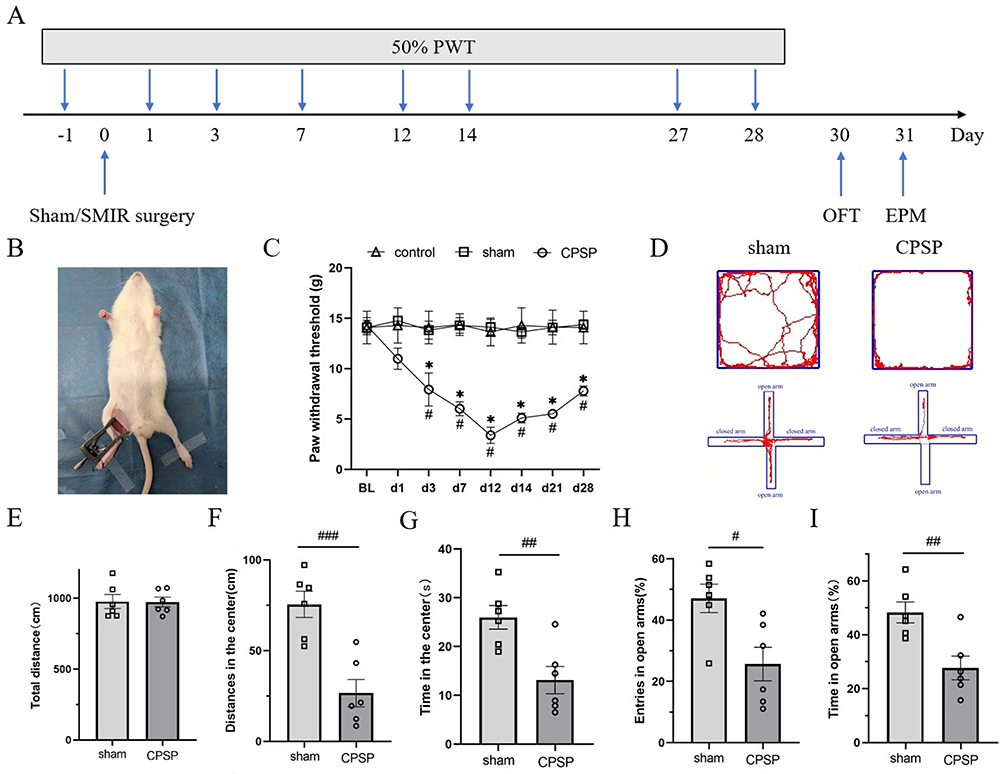 |
Figure 1 Mechanical hyperalgesia and anxiety-like negative emotion in CPSP rats.(A) Schematic of the experimental design; (B) The schematic diagram of the model; (C) Compared with the preoperative basic value, the PWT of the hindfoot of the CPSP group began to decrease from the first day after operation, decreased to the lowest on the 12th day and lasted for 28 days; compared with the sham group and the control group, the PWT of the CPSP group was significantly different on the 3rd, 7th, 12th, 14th, 21st and 28th day after operation; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; *p<0.05, vs baseline; #p<0.05, vs sham/control group, BL=baseline.) (D) On the left, the original trajectories of the open field test and elevated plus maze test of rats in sham group are represented, and on the right, the original trajectories of open field test and elevated maze test of rats in CPSP group are represented; (E–G) The total distance statistical chart of open field experiment, the statistical chart of central activity distance and the statistical chart of central activity time, respectively.(independent Samples t-test Welch’s correction, ##p<0.01, ###p<0.001, vs sham group.); (H and I) The statistical graphs of the percentage of open arm entry times and open arm residence time in elevated cross maze experiment, respectively. (independent Samples t-test Welch’s correction, #p<0.05, ##p<0.01, vs sham group.). |
There was no statistically significant difference in the baseline PWT among the three groups before surgery (p > 0.05). The PWT of rats in the CPSP group began to decrease on post-surgery day 1, reached the lowest level on day 12, and persisted until day 28. Compared with the sham group and control group, the PWT of the CPSP group showed statistically significant differences on post-surgery days 3, 7, 12, 14, 21, and 28 (p<0.05) (Figure 1C). There was no statistically significant difference in PWT between the control group and sham group at any time point; therefore, the sham group was used as the control for the CPSP group in subsequent experiments. These results indicate that the SMIR procedure successfully induced mechanical hyperalgesia in rats and established the CPSP model.
To evaluate whether CPSP model rats exhibited negative emotions, the OFT was used to detect anxiety-like emotions. The results showed that CPSP rats generally developed anxiety-like emotions 12–14 days after modeling. Although there was no statistically significant difference in total distance traveled, compared with the sham group, the CPSP group showed a significant decrease in central distance traveled (t=4.696, p<0.001) and central time spent (t=3.489, p=0.006), with statistically significant differences (Figure 1D–G).
In the EPM, compared with the sham group, the CPSP group showed a significant decrease in the percentage of open arm entries (t=2.985, p=0.014) and the percentage of time spent in open arms (t=3.523, p=0.006), with statistically significant differences (Figure 1H and I). These results indicate that CPSP rats were accompanied by anxiety-like negative emotions, but there was no significant change in motor ability.
Histological Verification of Light Stimulation Sites
Histological examination confirmed that the ChR2 light-sensitive virus were specifically expressed in the LC region, with no significant off-target diffusion to adjacent brain regions. The on-target expression rate of the ChR2 light-sensitive virus in the LC was 93.6 ± 2.1%. Abundant viral-labeled axonal terminals were distributed in the IL and PL, with targeting accuracy quantified as 91.2 ± 2.5% (IL) and 92.5 ± 1.8% (PL), respectively.
Functional Validation by c-Fos Immunoreactivity
To verify that optogenetic manipulations produced intended neural modulation, c-Fos expression was quantified in IL and PL following light stimulation (n=3 rats/condition, 90 min post-stimulation, 3 sections/rat, counter blinded to condition).
LC-IL Circuit Activation
Blue light stimulation of LC-ChR2 terminals in IL significantly increased c-Fos cell density compared to sham+mch+light controls (156.3±18.7 vs. 48.2±7.4 cells/mm2, independent t-test: t=5.89, df=4, p=0.004, Cohen’s d=2.63, 3.2-fold increase). This confirms that terminal activation drives postsynaptic neural activation in IL.
LC-IL Circuit Inhibition
Yellow light stimulation of LC-NpHR terminals in CPSP rats significantly decreased c-Fos cell density compared to CPSP+mch+light controls (31.5±6.2 vs. 82.7±11.3 cells/mm2, t=4.12, df=4, p=0.014, Cohen’s d=1.84, 62% decrease). This confirms effective suppression of terminal neurotransmission.
Light-Alone Controls
No significant difference in c-Fos density between mch+light and mch-dark conditions in either IL (45.3±8.1 vs. 42.8±7.6, t=0.24, p=0.82) or PL (38.7±6.4 vs. 41.2±7.1, t=0.31, p=0.77), confirming that behavioral effects were not attributable to light exposure, heat, or insertion alone.
LC-PL Circuit
C-Fos changes in PL following LC-PL terminal manipulation showed variable trends but did not reach statistical significance (activation: 89.3±15.2 vs.72.6±11.8, t=0.98, p=0.38; inhibition: 68.4±9.7 vs. 75.1±12.3, t=0.52, p=0.63), consistent with the absence of behavioral effects.
Blue Light Activation of the LC Region Induces Mechanical Hyperalgesia and Anxiety-Like Emotions in Sham-Operated Rats
Literature suggests that the LC region is activated during chronic pain. We hypothesized that optogenetic activation of the LC region in sham-operated (sham) rats might induce mechanical hyperalgesia and negative emotions similar to those observed in CPSP model rats. To verify this, the experimental protocol was designed as shown in Figure 2A: the light-sensitive virus ChR2 or the empty virus mch was microinjected into the rat LC region using stereotaxic technology (Figure 2B). Sham surgery was performed 2 weeks later, and 4 weeks after viral expression, confocal microscopy was used to observe the expression of the ChR2 light-sensitive virus in the LC region (Figure 2C).
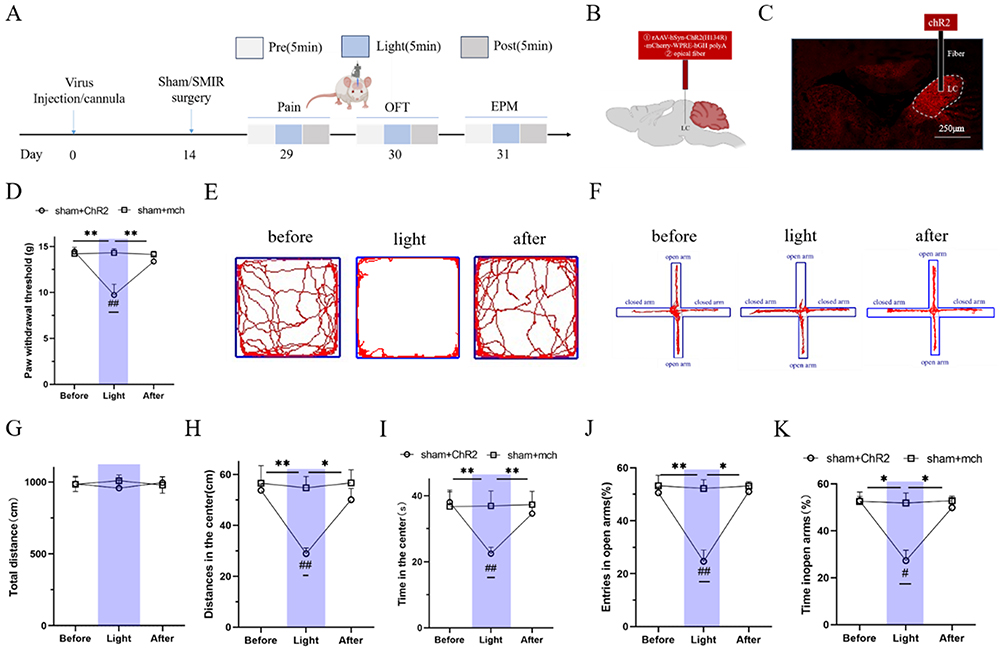 |
Figure 2 Blue light activation of LC causes mechanical hyperalgesia and anxiety-like negative emotion in sham operaterd group rats.(A) Schematic of the experimental design; (B) The original map of photogenetic virus expression at LC; (C) The schematic diagram of LC injection of photogenetic virus ChR2 or mch group; (D) The statistical map of the threshold of mechanical paw contraction of blue light activated normal rat LC neurons; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method, **p<0.01, vs before/after, ##p<0.01, vs sham+mch group. (E) the original representative map of the open field test trajectory of the ChR2 group with blue light activation; (F) The original trajectory representation of the elevated plus maze of the ChR2 group with blue light activation; (G–I) The statistical chart of the total activity distance, the statistical chart of the central activity distance and the statistical chart of the central activity time of the open-field test,*p<0.05, **p<0.01, vs before/after; ##p<0.01, vs sham+mch group; (J and K) The statistical chart of the percentage of open-arm entry times and open-arm retention time of the elevated cross maze experiment, respectively. (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method, *p<0.05, **p<0.01, vs before/after; #p<0.05, ##p<0.01, vs sham+mch group.). |
Blue light was used to stimulate the LC region, and changes in PWT were measured. The results showed that compared with the sham+mch group, the PWT of rats in the sham+ChR2 group was significantly decreased (p=0.002), with a statistically significant difference. Compared with the pre-stimulation period, the PWT of the sham+ChR2 group was significantly decreased during blue light stimulation (p=0.007), with a statistically significant difference. Compared with the post-stimulation period, the PWT of the sham+ChR2 group was also significantly decreased during blue light stimulation (p=0.008), with a statistically significant difference (Figure 2D).
Subsequently, the OFT and EPM were used to observe the effect of blue light activation of the LC region on anxiety-like emotions. In the OFT, compared with the sham+mch group, the sham+ChR2 group showed a significant decrease in central distance traveled (p=0.001) and central time spent (p=0.008) during blue light stimulation, with statistically significant differences, and there was no impact on the rats’ motor ability. Compared with the pre-stimulation period, the sham+ChR2 group showed a significant decrease in central distance traveled (p=0.003) and central time spent (p=0.006) during blue light stimulation, with statistically significant differences. Compared with the post-stimulation period, the sham+ChR2 group also showed a significant decrease in central distance traveled (p=0.043) and central time spent (p=0.021) during blue light stimulation, with statistically significant differences. These results suggest that blue light activation of the LC region in sham rats can induce anxiety-like negative emotions.
In the EPM, compared with the sham+mch group, the sham+ChR2 group showed a significant decrease in the percentage of open arm entries (p=0.009) and the percentage of time spent in open arms (p=0.013) during blue light activation of the LC region, with statistically significant differences (Figure 2E–I). Compared with the pre-stimulation period, the sham+ChR2 group showed a significant decrease in the percentage of open arm entries (p=0.004) and the percentage of time spent in open arms (p=0.013) during blue light stimulation, with statistically significant differences. Compared with the post-stimulation period, the sham+ChR2 group also showed a significant decrease in the percentage of open arm entries (p=0.016) and the percentage of time spent in open arms (p=0.021) during blue light stimulation, with statistically significant differences (Figure 2F–K).
Collectively, these results indicate that blue light activation of the LC region in sham rats can induce mechanical hyperalgesia and anxiety-like negative emotions similar to those in CPSP model rats.
Yellow Light Inhibition of the LC-IL Neural Circuit Alleviates Mechanical Hyperalgesia and Anxiety-Like Negative Emotions in CPSP Rats
Our optogenetic experiments showed that blue light activation of the LC region in sham rats induced mechanical hyperalgesia and anxiety-like emotions similar to those in CPSP model rats, suggesting that the LC-mPFC neural circuit may be involved in regulating mechanical hyperalgesia and anxiety-like emotions in CPSP rats. To confirm this hypothesis, retrograde viral tracing and optogenetic experiments were performed.
The retrograde tracing virus scAAV-hSyn-EGFP-WPREs was microinjected into the mPFC region using stereotaxic technology. One week later, perfusion and sectioning were performed. Under the microscope, high expression of the tracing virus was observed in the injected mPFC region, and viral expression was also observed in the ipsilateral LC region. This verified that the LC region has neural projections to the mPFC region, confirming the existence of the LC-mPFC neural circuit (Figure 3B).
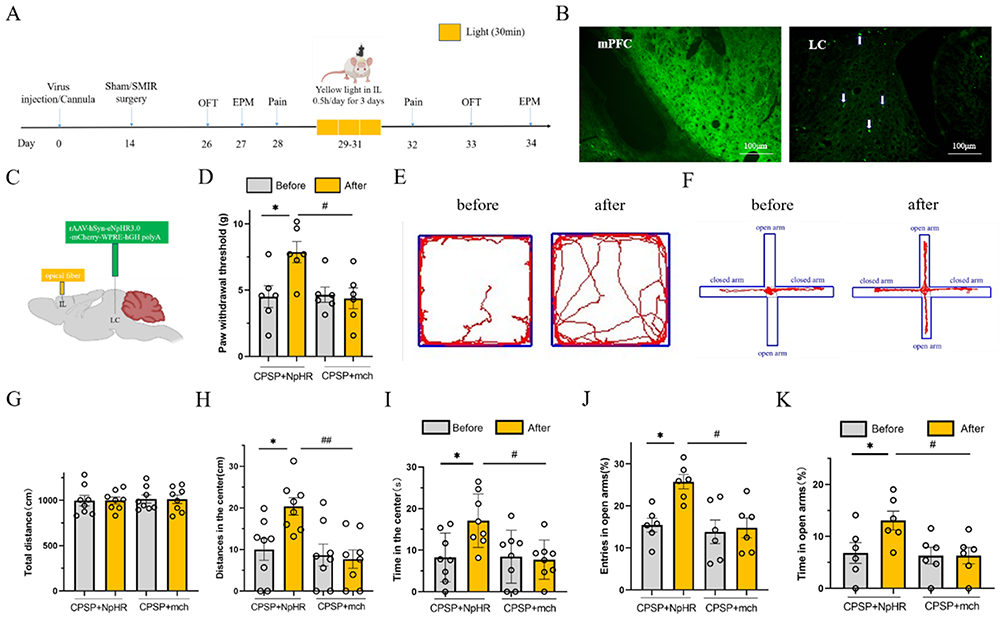 |
Figure 3 Yellow light inhibition of LC-IL neural circuit reduces mechanical hyperalgesia and anxiety negative emotion in CPSP rats.(A) Schematic design of the experiment; (B) The fluorescence expression representation of retrograde tracer virus at mPFC and LC, and the white arrow marks the fluorescence signal at LC; (C) Schematic diagram of NpHR injection and optical fiber embedding location of photogenetic suppression virus; (D) Statistical diagram of the effect of LC-IL neural circuit on mechanical foot reduction threshold in CPSP rats inhibited by yellow light; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; *p<0.05 vs before, #p<0.05 vs CPSP+mch group.) (E) The original track representation of open field experiment before and after photogenetic inhibition of LC-IL neural circuit in CPSP rats; (F) Are the tracks of elevated plus maze experiment in front and back of LC-IL neural circuits in photogenetically inhibited CPSP rats; (G–I) Total distance, central distance and central time of activity in open field test of LC-IL neural circuit in CPSP rats; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; *p<0.05 vs before; #p<0.05, ##p<0.05 vs CPSP+mch group.) (J and K) Statistical diagram of percentage of open arm entry times and percentage of open arm retention time in the elevated plus maze testof LC-IL neural circuit in CPSP rats with photogenetic suppression. (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; *p<0.05 vs before, #p<0.05 vs CPSP+mch group.). |
The IL is a major component of the mPFC. We used optogenetic technology to inhibit the LC-IL neural circuit with yellow light, and measured changes in PWT and anxiety-like emotions to explore the role of the LC-IL neural circuit. The experimental protocol is shown in Figure 3A: the light-sensitive virus NpHR and the empty virus mch were microinjected into the rat LC region, and an optical fiber was implanted in the IL region (Figure 3C). CPSP modeling was performed 2 weeks later. Four weeks after viral expression, the rats received 30 minutes of yellow light stimulation in the IL region once a day for 3 consecutive days, and PWT was measured 1 day later.
The results showed that compared with the CPSP+mch group, the PWT of rats in the CPSP+NpHR group was significantly increased (p=0.018), with a statistically significant difference. Compared with the pre-stimulation period, the PWT of the CPSP+NpHR group was significantly increased after yellow light inhibition (p=0.023), with a statistically significant difference (Figure 3D).
In the OFT, compared with the CPSP+mch group, the CPSP+NpHR group showed a significant increase in central distance traveled (p=0.018) and central time spent (p=0.013), with statistically significant differences. Compared with the pre-stimulation period, the CPSP+NpHR group showed a significant increase in central distance traveled (p=0.028) and central time spent (p=0.027) after yellow light inhibition, with statistically significant differences (Figure 3E–I).
Similar results were observed in the EPM: compared with the CPSP+mch group, the CPSP+NpHR group showed a significant increase in the percentage of open arm entries (p=0.014) and the percentage of time spent in open arms (p=0.016), with statistically significant differences. Compared with the pre-stimulation period, the CPSP+NpHR group showed a significant increase in the percentage of open arm entries (p=0.021) and the percentage of time spent in open arms (p=0.027) after yellow light inhibition, with statistically significant differences (Figure 3F–K).
Comprehensive analysis of the above experimental results indicates that inhibition of the LC-IL neural circuit can alleviate mechanical hyperalgesia and anxiety-like negative emotions in CPSP rats.
Blue light activation of the LC-IL neural circuit induces mechanical hyperalgesia and anxiety-like negative emotions in sham-operated rats since inhibition of the LC-IL neural circuit alleviates mechanical hyperalgesia and anxiety-like negative emotions in CPSP rats, we further explored whether activation of the LC-IL neural circuit in sham rats could induce hyperalgesia and negative emotions similar to those in CPSP model rats using optogenetic technology.
The experimental protocol is shown in Figure 4A: rats were randomly divided into two groups (n=10,each), and the LC region was injected with the light-sensitive virus ChR2 or the empty virus mch, respectively (Figure 4B). Sham surgery was performed after 2 weeks of recovery. Four weeks after viral expression, the rats received 30 minutes of blue light stimulation in the IL region once a day for 3 consecutive days (Figure 4A–C), and PWT was measured 1 day later.
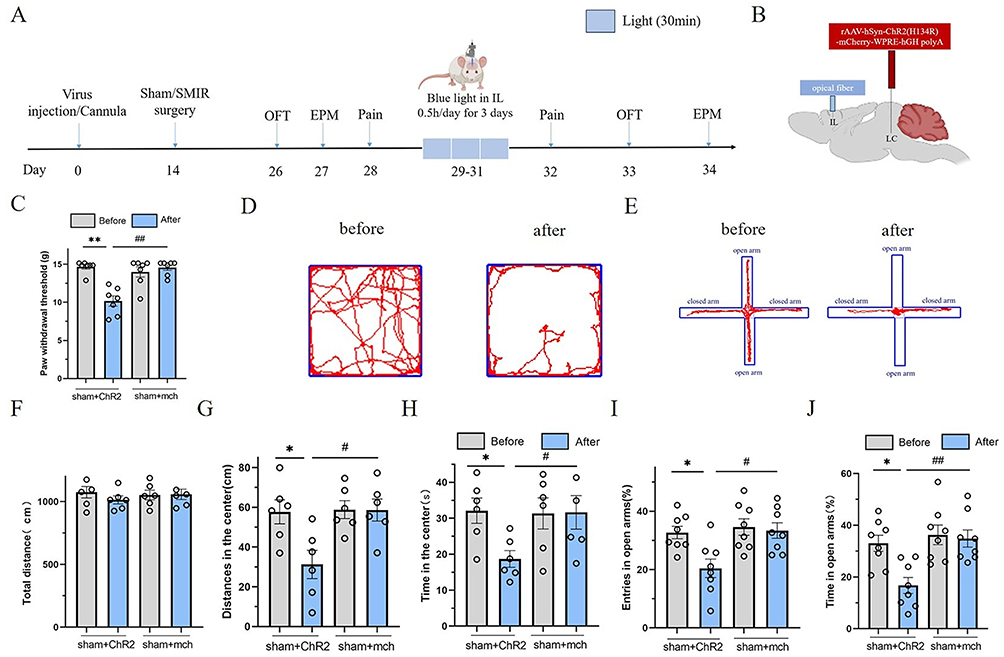 |
Figure 4 Blue light activation of LC-IL neural circuit causes mechanical pain sen-sitization and anxiety-like emotion in sham operaterd group rats.(A) Schematic design of the experiment; (B) Schematic diagram of ChR2 injection and optical fiber embedding location of optogenetic virus; (C) Statistical diagram of the effect of blue light activation of normal rat LC-IL neural circuit on the threshold of mechanical foot contraction; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; **p<0.01, vs before; ##p<0.01, vs sham+mch group.) (D) The original track representation of open field experiment before and after LC-IL neural circuit activated by blue light in ChR2 group rats; (E) the original track representation of elevated plus maze test before and after activation of LC-IL neural circuit in ChR2 group rats by blue light; (F–H) Are the total open field distance, the central active distance and the central time of open field test of LC-IL neural circuit in chR2 group and sham group, respectively; (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method;*p<0.05, vs before; #p<0.05, vs sham+mch group.) (I and J) Are the percentage statistical graphs of open-arm entry times and percentage statistical graphs of open-arm retention time of blue-activated ChR2 group rats and sham group rats in the LC-IL neural circuit elevated plus maze test, respectively. (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method;*p<0.05, vs before; #p<0.05, ##p<0.01, vs sham+mch group.). |
The results showed that compared with the sham+mch group, the PWT of rats in the sham+ChR2 group was significantly decreased (p=0.001), with a statistically significant difference. Compared with the pre-stimulation period, the PWT of the sham+ChR2 group was significantly decreased after blue light activation (p=0.001), with a statistically significant difference (Figure 4C).
In the OFT, compared with the sham+mch group, the sham+ChR2 group showed a significant decrease in central distance traveled (p=0.012) and central time spent (p=0.045), with statistically significant differences. Compared with the pre-stimulation period, the sham+ChR2 group showed a significant decrease in central distance traveled (p=0.015) and central time spent (p=0.037) after blue light activation, with statistically significant differences (Figure 4D–H).
Similar results were observed in the EPM: compared with the sham+mch group, the sham+ChR2 group showed a significant decrease in the percentage of open arm entries (p=0.022) and the percentage of time spent in open arms (p=0.009), with statistically significant differences. Compared with the pre-stimulation period, the sham+ChR2 group showed a significant decrease in the percentage of open arm entries (p=0.031) and the percentage of time spent in open arms (p=0.021) after blue light activation, with statistically significant differences (Figure 4E–J).
These results indicate that activation of the LC-IL neural circuit in sham rats can induce hyperalgesia and anxiety-like negative emotions, confirming that the rat LC-IL neural circuit is involved in regulating hyperalgesia and anxiety-like negative emotions.
The LC-PL Neural Circuit Does Not Affect Mechanical Hyperalgesia or Anxiety-Like Negative Emotions in CPSP Rats
Studies have shown that the PL subregion of the mPFC is also involved in pain and emotion coding. However, further experiments are needed to confirm whether the LC-PL neural circuit regulates mechanical hyperalgesia and anxiety-like negative emotions in CPSP rats. We adopted a strategy similar to that used to explore the LC-IL circuit, with the optical fiber placed in the PL subregion.
The experimental protocol is shown in Figure 5A: the light-sensitive virus NpHR and the empty virus mch were microinjected into the rat LC region, and an optical fiber was implanted in the PL subregion (Figure 5B). CPSP modeling was performed 2 weeks later. Four weeks after viral expression, the rats received 30 minutes of yellow light stimulation in the PL subregion once a day for 3 consecutive days, and PWT was measured 1 day later.
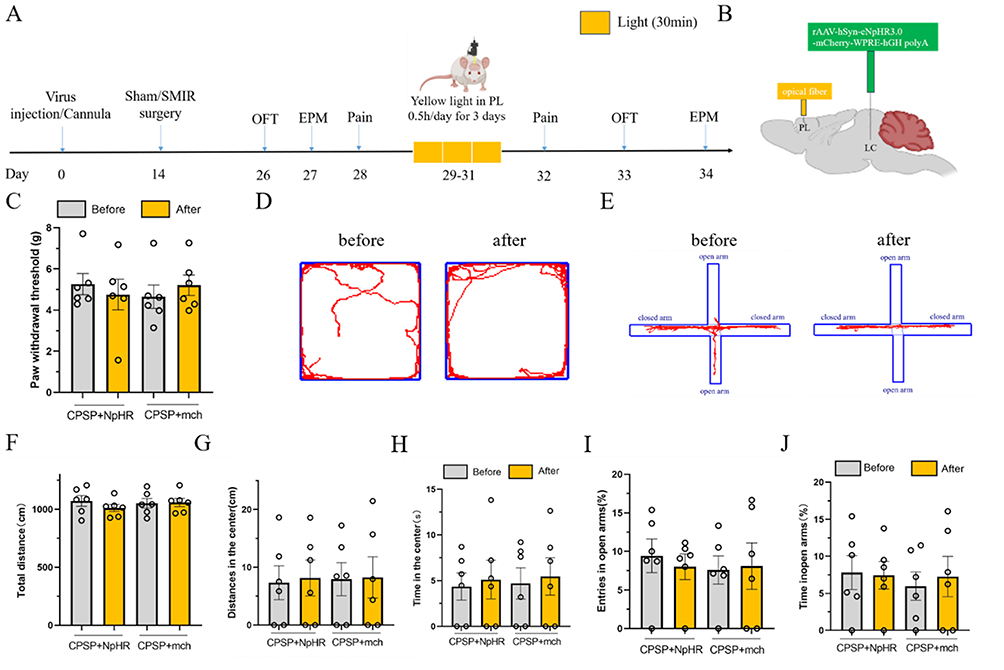 |
Figure 5 Yellow light inhibition of LC-PL neural circuit does not affect mechanical pain sensitivity and anxiety-like emotion in CPSP rats.(A) Schematic of the experimental design; (B) Schematic diagram of NpHR injection of photogenetic virus and embedding position of optical fiber; (C) Statistical diagram of mechanical pain threshold changes in rats with yellow light inhibition of LC-PL nerve circuit; (D) The representative map of the original trajectory of the open field test before and after the LC-PL nerve circuit of yellow light inhibition in CPSP rats; (E) The trajectory representation of elevated plus maze test before and after yellow light inhibition of LC-PL nerve circuit in CPSP rats; (F–H) Total distance statistics, central activity distance statistics and central time statistics of open field test in LC-PL neural circuit of CPSP rats were inhibited by yellow light inhibition; (I and J) Yellow light inhibition CPSP rats LC-PL nerve circuit elevated plus maze test open arm entry percentage statistical chart and open arm retention time percentage statistical chart. (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; p> 0.05, vs before; p> 0.05, vs CPSP+mch group.). |
The results showed that compared with the CPSP+mch group, there was no statistically significant difference in PWT in the CPSP+NpHR group (p>0.05). Compared with the pre-stimulation period, there was also no statistically significant difference in PWT in the CPSP+NpHR group after yellow light inhibition (p>0.05, Figure 5C).
In the OFT, compared with the CPSP+mch group, there were no statistically significant differences in central distance traveled (p>0.05) or central time spent (p>0.05) in the CPSP+NpHR group. Compared with the pre-stimulation period, there were also no statistically significant differences in central distance traveled (p>0.05) or central time spent (p>0.05) in the CPSP+NpHR group after yellow light inhibition (p>0.05, Figure 5D–H).
Similar results were observed in the EPM: compared with the CPSP+mch group, there were no statistically significant differences in the percentage of open arm entries (p>0.05) or the percentage of time spent in open arms (p>0.05) in the CPSP+NpHR group. Compared with the pre-stimulation period, there were also no statistically significant differences in the percentage of open arm entries (p>0.05) or the percentage of time spent in open arms (p>0.05) in the CPSP+NpHR group after yellow light inhibition (Figure 5E–J).
These experimental results indicate that the LC-PL neural circuit is not involved in regulating mechanical hyperalgesia or anxiety-like negative emotions in CPSP rats.
Based on this, we hypothesized that blue light activation of the LC-PL neural circuit in sham rats would not induce mechanical hyperalgesia or negative emotions, and experiments were conducted to verify this. The experimental protocol is shown in Figure 6A: rats were randomly divided into two groups (n=10,each), and the LC region was injected with the light-sensitive virus ChR2 or the empty virus mch, respectively. Sham surgery was performed after 2 weeks of recovery. Four weeks after viral expression, the rats received 30 minutes of blue light stimulation in the PL subregion once a day for 3 consecutive days, and PWT was measured 1 day later.
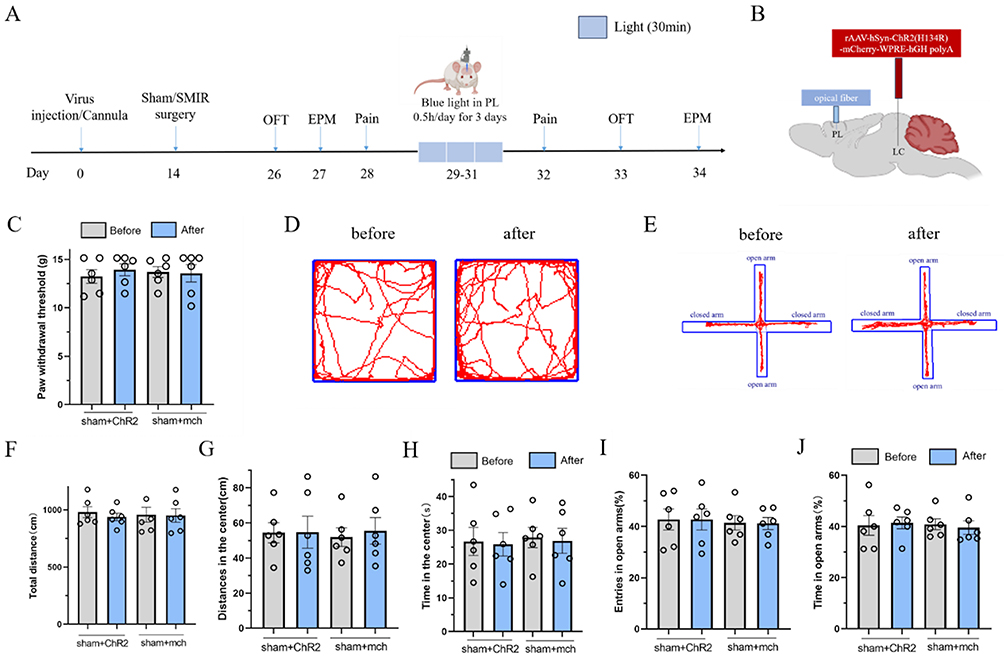 |
Figure 6 Blue light activation of LC-PL neural circuit does not affect mechanical pain sensitivity and anxiety-like emotion in rats.(A) Schematic of the experimental design; (B) Schematic diagram of ChR2 injection of photogenetic virus and embedding position of optical fiber; (C) The effect of blue light activated LC-PL nerve circuit on the mechanical PWT in normal rats; (D) The open field experimental trajectory before and after photogenetic activation of LC-PL nerve circuit in rats. (E) The left side is the representative map of the elevated plus maze test before and after blue light activated LC-PL nerve circuit in normal rats; (F–H) Blue light activated LC-PL nerve circuit rats open field test total distance statistical chart, central activity distance statistical chart and central activity time statistical chart; (I and J) The percentage statistics of open arm entry times and open arm retention time in LC-PL nerve circuit elevated plus maze test of rats activated by blue light heredity. (values represent the Mean±SEM; two-way ANOVA of repeated measures, multiple comparison of Bonferroni method; p> 0.05, vs before; p> 0.05, vs sham+mch group.). |
The results showed that compared with the sham+mch group, there was no statistically significant difference in PWT in the sham+ChR2 group (p>0.05, Figure 6C). In the OFT, compared with the sham+mch group, there were no statistically significant differences in central distance traveled (p>0.05) or central time spent (p>0.05) in the sham+ChR2 group (Figure 6D–H). Similar results were observed in the EPM: compared with the sham+mch group, there were no statistically significant differences in the percentage of open arm entries (p>0.05) or the percentage of time spent in open arms (p>0.05) in the sham+ChR2 group (Figure 6E–J).
These results indicate that activation of the LC-PL neural circuit in sham rats fails to induce mechanical hyperalgesia and anxiety-like emotions similar to those in CPSP model rats, suggesting that the LC-PL neural circuit is not involved in mechanical hyperalgesia or anxiety-like emotions in CPSP model rats.
Discussion
The high incidence of chronic post-surgical hyperalgesia poses a severe challenge in clinical practice. CPSP patients not only suffer from chronic pain but also exhibit anxiety-like negative emotions.44 Consistent with this, our study found that CPSP model rats showed mechanical hyperalgesia and anxiety-like emotions. Functional magnetic resonance imaging (fMRI) studies have revealed significantly increased activity in brain regions associated with emotion during pain,44 suggesting that long-term pain in CPSP patients can activate brain regions regulating emotion, leading to the generation and maintenance of negative emotions. In turn, negative emotions can exacerbate the duration and intensity of pain, forming a vicious cycle that greatly complicates clinical treatment.
Previous studies have shown that numerous brain nuclei are involved in pain regulation, such as the LC, central nucleus of the amygdala (CEA), mPFC, ACC, lateral habenula (LHb), and dorsal raphe nucleus (DRN).45–49 LC neurons are the main source of norepinephrine in the forebrain, and the norepinephrine system plays a crucial role in regulating chronic pain and anxiety-like emotions. Studies have found that when the LC responds to aversive stimuli, widespread norepinephrine release signals can be observed in the forebrain.16,29
The LC-norepinephrine system exhibits time-dependent neuroplastic changes: during acute pain, activation of LC noradrenergic neurons enhances endogenous anti-nociceptive effects, thereby exerting an analgesic effect. In contrast, during long-term chronic pain, activation of the LC may enhance pain perception in patients and further lead to comorbid negative emotions such as anxiety.16,29 Our study found that optogenetic activation of the LC region in sham rats with blue light induced mechanical hyperalgesia and anxiety-like emotions similar to those in CPSP model rats. Conversely, inhibition of LC neurons projecting to the IL subregion in CPSP model rats with yellow light alleviated mechanical hyperalgesia and anxiety-like emotions. These findings confirm that the LC is involved in comorbid mechanical hyperalgesia and anxiety in CPSP. Blocking time-dependent neuroplastic changes in the LC-norepinephrine system may prevent or reduce the chronicization of pain and the development of comorbid anxiety-like emotions.
Studies in both humans and rodents have shown that the mPFC plays a key role in pain signal processing and emotional regulation during acute and chronic pain.50,51 Notably, clinical studies have reported that stimulation of the human dorsolateral mPFC can effectively alleviate hyperalgesia. Noradrenergic neurons in the LC project to the mPFC region, and our retrograde viral tracing experiments confirmed this projection circuit. The mPFC consists of three subregions: the ACC, IL, and PL. Since the role of the ACC in pain and emotion regulation has been extensively studied, we focused on exploring the roles of the LC-IL and LC-PL neural circuits in regulating mechanical hyperalgesia and anxiety-like emotions in CPSP model rats.
The IL subregion of the mPFC is involved in both negative and positive emotional behaviors.52,53 Lesions of the IL subregion in rats result in an anxiolytic phenotype, while activation of this subregion induces anxiety-like behaviors.54 Electrical stimulation of the IL significantly inhibits conditioned fear responses and promotes fear extinction, whereas inactivation of the IL consolidates and reinstates fear.55,56 Non-selective lesions of the IL prevent reward-seeking behaviors, indicating that the IL serves as an integration center for reward-seeking behaviors.54,57 These studies collectively suggest that the IL subregion is closely associated with emotional processing but exerts different regulatory effects in different emotional disorders.
Our study found that optogenetic inhibition of the LC-IL neural circuit with yellow light in CPSP model rats alleviated mechanical hyperalgesia and anxiety-like emotions. Conversely, activation of the LC region and LC-IL neural circuit in sham rats with blue light induced mechanical hyperalgesia and anxiety-like emotions. These findings not only confirm that the IL subregion is involved in regulating comorbid chronic pain and anxiety in CPSP rats but also indicate that activation of the IL subregion is directly associated with activation of LC norepinephrine.
The PL subregion of the mPFC plays important roles in decision-making, cognition, emotional regulation, and pain.35,36,58 Electrical stimulation of the rat PL region significantly prolonged the latency of hot plate and tail-flick responses, suggesting that activation of the PL subregion exerts an analgesic effect, possibly through the periaqueductal gray-rostral ventromedial medulla (PAG-RVM) descending pain regulatory pathway.59 In a neuropathic pain model, optogenetic activation of parvalbumin (PV)-positive inhibitory neurons in the PL subregion also exerted an analgesic effect, while activation of excitatory neurons in this region promoted pain.60 In a chronic inflammatory pain model, specific activation of PL neurons contralateral to the lesion reduced mechanical hyperalgesia and alleviated anxiety-like behaviors in rats, whereas inhibition of contralateral PL neurons induced significant pro-nociceptive and anxiety-like behaviors.61,62 These studies suggest that the function of the PL subregion varies significantly across different pain models and emotional disorders.
In our study, optogenetic inhibition of the LC-PL circuit in CPSP model rats failed to alleviate mechanical hyperalgesia or anxiety-like emotions. Furthermore, activation of the LC-PL neural circuit in sham rats did not induce mechanical hyperalgesia or anxiety-like negative emotions similar to those in CPSP rats. The present study revealed a pathway-specific role of the LC-IL circuit, but not the LC-PL circuit, in regulating mechanical hyperalgesia and comorbid anxiety-like behaviors in CPSP. To ensure the validity of this differential effect, rigorous anatomical verifications were performed, including confirmation of accurate viral targeting in the LC, optical fiber placement in IL and PL, and comparable axon terminal expression of LC-derived noradrenergic projections in both subregions. Symmetrical experimental designs and control treatments were applied across all groups to exclude potential bias from viral expression efficiency or light stimulation parameters. Statistical analyses including repeated-measures ANOVA and group × time interaction analysis further supported the reliability of the null effect observed in the PL pathway. The lack of effect in PL is unlikely to be attributable to insufficient statistical power, inadequate viral expression, or misplaced optical fibers. Instead, it likely reflects the divergent functional roles of IL and PL in pain and emotional regulation, whereby the IL mediates descending pain inhibition and fear extinction, whereas the PL contributes to pain-related affective processing but is not critically involved in the CPSP phenotype examined in this study.
Several limitations should be noted in the current study. First and foremost, this is an exploratory study exclusively conducted in male rats. Substantial clinical and preclinical evidence has verified prominent sex differences in the development of chronic postoperative pain, the structural and functional characteristics of the LC-noradrenergic system, infralimbic cortical circuits mediating anxiety-like behaviors, and the manifestation patterns of pain-anxiety comorbidity.41–43 To avoid the confounding effects of sex hormones, we only selected male animals for this preliminary mechanistic research. Accordingly, our findings cannot be directly extrapolated to females, and further studies using both male and female rodents are urgently needed to validate the LC-IL circuit mechanism and assess its sex-specific translational potential. Second, while we report quantitative c-Fos counts confirming bidirectional neural modulation, publication-quality immunofluorescence images were not available for all conditions due to suboptimal initial imaging resolution. Future studies will employ standardized high-resolution confocal protocols to provide complete pictorial documentation alongside quantification. Third, we did not explore the role of the ACC, the other subregion of the mPFC, in hyperalgesia and comorbid emotions in CPSP rats. Fourth, we did not further identify the specific neurons in the LC and IL that regulate mechanical hyperalgesia and comorbid emotions in CPSP rats. The molecular mechanisms underlying the function of the LC-IL neural circuit were not investigated.
In conclusion, The LC-IL neural circuit is critically involved in regulating comorbid mechanical hyperalgesia and anxiety in male CPSP rats, whereas the LC-PL neural circuit is not. To our knowledge, this is the first study to investigate the differential roles of LC-IL and LC-PL pathways in mechanical hyperalgesia and comorbid anxiety-like negative emotions in a CPSP model. The present findings should be interpreted as male-specific circuit mechanisms due to the exclusive use of male subjects, and may not generalize to females. The lack of significant effect in the LC-PL pathway was further supported by adequate statistical analysis and controlled comparisons, indicating a genuine absence of effect rather than insufficient statistical power. These findings provide novel and pathway-specific evidence for understanding CPSP-related pain and emotion comorbidity, and hold implications for exploring potential intervention targets and developing therapeutic strategies for CPSP. Targeting the LC–IL circuit, which we identified as a key regulator of CPSP-related mechanical hyperalgesia and anxiety, could open up new therapeutic avenues. For instance, deep brain stimulation (DBS) of the LC or its projections to the IL may modulate noradrenergic signaling to restore the descending pain inhibitory pathway, thereby alleviating persistent pain and comorbid anxiety. Transcranial magnetic stimulation (TMS) targeting the mPFC (including the IL) could also indirectly regulate LC–IL circuit activity, offering a non-invasive therapeutic option. Additionally, pharmacological interventions that selectively enhance noradrenergic transmission in the LC–IL pathway, while avoiding off-target effects on other LC projections, may provide a more precise and tolerable treatment strategy for CPSP. Although these potential applications require further preclinical and clinical validation, our study lays a foundation for translating basic circuit research into novel therapeutic approaches for CPSP.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Project supported by Natural Science Foundation of Fujian Province, China (Grant No. 2023J011723); National Natural Science Foundation of China (NSFC) (Grant No. 82471229).
Disclosure
The authors have no commercial association (ie, consultancies, stock ownership, equity interest, patent/licensing arrangements, etc.) that might pose a conflict of interest in connection with the submitted manuscript.
References
1. Kehlet H, S JT, Woolf CJ. Persistent postsurgical pain: risk factors and prevention. Lancet. 2006;367(9522):1618–21. doi:10.1016/S0140-6736(06)68700-X
2. Wylde V, Dennis J, Beswick AD, et al. Systematic review of management of chronic pain after surgery. Br J Surg. 2017;104(10):1293–1306. doi:10.1002/bjs.10601
3. Kubulus C, Mahlstedt S, Wagenpfeil G, et al. Chronic pain patients and time to sustained acceptable pain scores after major surgery - A retrospective registry analysis. J Clin Anesth. 2023;89:111152. doi:10.1016/j.jclinane.2023.111152
4. C BM, Ceko M, Low LA. Cognitive and emotional control of pain and its disruption in chronic pain. Nat Rev Neurosci. 2013;14(7):502–511. doi:10.1038/nrn3516
5. H KC, Cohen RC. Chronic anal pain: a review of causes, diagnosis, and treatment. Cleve Clin J Med. 2022;89(6):336–343. doi:10.3949/ccjm.89a.21102
6. Aziz Q, Giamberardino MA, Barke A, et al. The IASP classification of chronic pain for ICD-11: chronic secondary visceral pain. Pain. 2019;160(1):69–76. doi:10.1097/j.pain.0000000000001362
7. Mauck MC, Smith J, Liu AY, et al. Chronic pain and itch are common, morbid sequelae among individuals who receive tissue autograft after major thermal burn injury. Clin J Pain. 2017;33(7):627–634. doi:10.1097/AJP.0000000000000446
8. Vandenkerkhof EG, Hopman WM, Reitsma ML, et al. Chronic pain, healthcare utilization, and quality of life following gastrointestinal surgery. Can J Anaesth. 2012;59(7):670–680. doi:10.1007/s12630-012-9712-x
9. Llorca-Torralba M, Camarena-Delgado C, Suárez-Pereira I, et al. Pain and depression comorbidity causes asymmetric plasticity in the locus coeruleus neurons. Brain. 2022;145(1):154–167. doi:10.1093/brain/awab239
10. Baker DW. History of the joint commission’s pain standards: lessons for today’s prescription opioid epidemic. JAMA. 2017;317(11):1117–1118. doi:10.1001/jama.2017.0935
11. Galgani A, Lombardo F, Della Latta D, et al. Locus Coeruleus magnetic resonance imaging in neurological diseases. Curr Neurol Neurosci Rep. 2020;21(1):2. doi:10.1007/s11910-020-01087-7
12. Li J, Wei Y, Zhou J, et al. Activation of locus coeruleus-spinal cord noradrenergic neurons alleviates neuropathic pain in mice via reducing neuroinflammation from astrocytes and microglia in spinal dorsal horn. J Neuroinflammation. 2022;19(1):123. doi:10.1186/s12974-022-02489-9
13. Suárez-Pereira I, Llorca-Torralba M, Bravo L, et al. The role of the locus coeruleus in pain and associated stress-related disorders. Biol Psychiatry. 2022;91(9):786–797. doi:10.1016/j.biopsych.2021.11.023
14. Sara S, Bouret S. Orienting and reorienting: the locus coeruleus mediates cognition through arousal. Neuron. 2012;76(1):130–141. doi:10.1016/j.neuron.2012.09.011
15. Mccall JG, Al-Hasani R, Siuda ER, et al. CRH Engagement of the Locus Coeruleus Noradrenergic System Mediates Stress-Induced Anxiety. Neuron. 2015;87(3):605–620. doi:10.1016/j.neuron.2015.07.002
16. Valentino RJ, Van Bockstaele E. Convergent regulation of locus coeruleus activity as an adaptive response to stress. Eur J Pharmacol. 2008;583(2–3):194–203. doi:10.1016/j.ejphar.2007.11.062
17. Maren S. Holmes A Stress and Fear Extinction. Neuropsychopharmacol. 2016;41(1):58–79.
18. Poe GR, Foote S, Eschenko O, et al. Locus coeruleus: a new look at the blue spot. Nat Rev Neurosci. 2020;21(11):644–659. doi:10.1038/s41583-020-0360-9
19. Benarroch EE. Benarroch E E Locus coeruleus. Cell Tissue Res. 2018;373(1):221–232. doi:10.1007/s00441-017-2649-1
20. Arnsten AFT. Arnsten A F Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci. 2009;10(6):410–422. doi:10.1038/nrn2648
21. Berridge CW, Waterhouse BD. Waterhouse B D The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42(1):33–84. doi:10.1016/s0165-0173(03)00143-7
22. Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. doi:10.1146/annurev.neuro.28.061604.135709
23. Kebschull JM, Garcia Da Silva P, Reid AP, et al. High-throughput mapping of single-neuron projections by sequencing of barcoded RNA. Neuron. 2016;91(5):975–987. doi:10.1016/j.neuron.2016.07.036
24. Schwarz LA, Miyamichi K, Gao XJ, et al. Viral-genetic tracing of the input-output organization of a central noradrenaline circuit. Nature. 2015;524(7563):88–92. doi:10.1038/nature14600
25. Uematsu A, Tan BZ, Ycu EA, et al. Modular organization of the brainstem noradrenaline system coordinates opposing learning states. Nat Neurosci. 2017;20(11):1602–1611. doi:10.1038/nn.4642
26. Hirschberg S, Li Y, Randall A, et al. Functional dichotomy in spinal- vs prefrontal-projecting locus coeruleus modules splits descending noradrenergic analgesia from ascending aversion and anxiety in rats. Elife. 2017;6.
27. Chandler DJ, Gao WJ, Waterhouse BD. Heterogeneous organization of the locus coeruleus projections to prefrontal and motor cortices. Proc Natl Acad Sci U S A. 2014;111(18):6816–6821. doi:10.1073/pnas.1320827111
28. Chandler DJ, Waterhouse BD, Gao W-J. New perspectives on catecholaminergic regulation of executive circuits: evidence for independent modulation of prefrontal functions by midbrain dopaminergic and noradrenergic neurons. Front Neural Circuits. 2014;8:53. doi:10.3389/fncir.2014.00053
29. Fan X, Song J, Ma C, et al. Noradrenergic signaling mediates cortical early tagging and storage of remote memory. Nat Commun. 2022;13(1):7623. doi:10.1038/s41467-022-35342-x
30. Shiers S, Price TJ. circuit, and anatomical changes in the prefrontal cortex in chronic pain. Pain. 2020;161(8):1726–1729. doi:10.1097/j.pain.0000000000001897
31. Chudasama Y. Chudasama Y Animal models of prefrontal-executive function. Behav Neurosci. 2011;125(3):327–343. doi:10.1037/a0023766
32. Lacroix L, Spinelli S, A HC, et al. Differential role of the medial and lateral prefrontal cortices in fear and anxiety. Behav Neurosci. 2000;114(6):1119–1130. doi:10.1037/0735-7044.114.6.1119
33. Sullivan RM, Gratton A. Behavioral effects of excitotoxic lesions of ventral medial prefrontal cortex in the rat are hemisphere-dependent. Brain Res. 2002;927(1):69–79. doi:10.1016/S0006-8993(01)03328-5
34. Bi LL, Wang J, Luo ZY, et al. Enhanced excitability in the infralimbic cortex produces anxiety-like behaviors. Neuropharmacology. 2013;72:148–156. doi:10.1016/j.neuropharm.2013.04.048
35. Peng WW, Tang ZY, Zhang FR, et al. Neurobiological mechanisms of TENS-induced analgesia. Neuroimage. 2019;195:396–408. doi:10.1016/j.neuroimage.2019.03.077
36. Patrick Hardy SG, Haigler HJ. Prefrontal influences upon the midbrain: a possible route for pain modulation. Brain Res. 1985;339(2):285–293. doi:10.1016/0006-8993(85)90094-0
37. Longyu M, Yue L, Liu S, et al. Dynamic changes of the infralimbic cortex and its regulation of the prelimbic cortex in rats with chronic inflammatory pain. Neurosci Bull. 2024;40:872–886. doi:10.1007/s12264-023-01159-x
38. Chen L, Cao S, Liu Y-Z, et al. CDK5-Induced HCN2 channel dysfunction in the prelimbic cortex drives allodynia and anxiety-like behaviors in neuropathic pain. Neurosci Bull. 2025;41:2254–2271. doi:10.1007/s12264-025-01546-6
39. Bouras NN, Mack NR, Gao W-J. Prefrontal modulation of anxiety through a lens of noradrenergic signaling. Front Syst Neurosci. 2023;17:1173326. doi:10.3389/fnsys.2023.1173326
40. Pan Q, Guo S-S, Chen M, et al. Representation and control of pain and itch by distinct prefrontal neural ensembles. Neuron. 2023;111:2414–2431.e7. doi:10.1016/j.neuron.2023.04.032
41. Vanneste S, Castejón-España J, Arulchelvan E, et al. Disentangling sex-specific mechanisms in neuropathic and nociplastic chronic pain: a review. Front Neurol. 2026;16:1730291. doi:10.3389/fneur.2025.1730291
42. Shen Z, Wei L, Chang W, et al. Sex differences in chronic pain-induced mental disorders: mechanisms of cerebral circuitry. Front Mol Neurosci. 2023;16:1102808. doi:10.3389/fnmol.2023.1102808
43. Silva JTD, Hernandez-Rojas LG, Mekonen HK, et al. Sex differences in visceral sensitivity and brain activity in a rat model of comorbid pain: a longitudinal study. Pain. 2023;165:698–706. doi:10.1097/j.pain.0000000000003074
44. Messina R, Gollion C, H CR, et al. Functional MRI in migraine. Curr Opin Neurol. 2022;35(3):328–335. doi:10.1097/WCO.0000000000001060
45. Halili A. Halili A Temporal model for central sensitization: a hypothesis for mechanism and treatment using systemic manual therapy, a focused review. MethodsX. 2023;10:101942. doi:10.1016/j.mex.2022.101942
46. Neugebauer V, Mazzitelli M, Cragg B, et al. Amygdala, neuropeptides, and chronic pain-related affective behaviors. Neuropharmacology. 2020;170:108052. doi:10.1016/j.neuropharm.2020.108052
47. D WT, Valdivia S, Khan A, et al. Dual and opposing functions of the central amygdala in the modulation of pain. Cell Rep. 2019;29(2):332–346.e5. doi:10.1016/j.celrep.2019.09.011
48. Y WX, B JW, Xu X, et al. A glutamatergic DRN-VTA pathway modulates neuropathic pain and comorbid anhedonia-like behavior in mice. Nat Commun. 2023;14(1):5124. doi:10.1038/s41467-023-40860-3
49. Zhou W, Jin Y, Meng Q, et al. A neural circuit for comorbid depressive symptoms in chronic pain. Nat Neurosci. 2019;22(10):1649–1658. doi:10.1038/s41593-019-0468-2
50. Dehdar K, Mooziri M, Samii Moghaddam A, et al. Corticosteroid treatment attenuates anxiety and mPFC-amygdala circuit dysfunction in allergic asthma. Life Sci. 2023;315:121373. doi:10.1016/j.lfs.2023.121373
51. Etkin A, Egner T, Kalisch R. Kalisch R Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cognit Sci. 2011;15(2):85–93. doi:10.1016/j.tics.2010.11.004
52. Pfarr S, Schaaf L, K RJ, et al. Choice for drug or natural reward engages largely overlapping neuronal ensembles in the infralimbic prefrontal cortex. J Neurosci. 2018;38(14):3507–3519. doi:10.1523/JNEUROSCI.0026-18.2018
53. Huang WC, Zucca A, Levy J, et al. Social behavior is modulated by valence-encoding mpfc-amygdala sub-circuitry. Cell Rep. 2020;32(2):107899. doi:10.1016/j.celrep.2020.107899
54. Chen YH, Wu JL, Hu NY, et al. Distinct projections from the infralimbic cortex exert opposing effects in modulating anxiety and fear. J Clin Invest. 2021;131(14). doi:10.1172/JCI145692.
55. Milad MR, Vidal-Gonzalez I, Quirk GJ. Electrical stimulation of medial prefrontal cortex reduces conditioned fear in a temporally specific manner. Behav Neurosci. 2004;118(2):389–394. doi:10.1037/0735-7044.118.2.389
56. Do-Monte FH, Manzano-Nieves G, Quiñones-Laracuente K, et al. Revisiting the role of infralimbic cortex in fear extinction with optogenetics. J Neurosci. 2015;35(8):3607–3615. doi:10.1523/JNEUROSCI.3137-14.2015
57. Izquierdo I, Furini CRG, Myskiw JC. Fear Memory. Physiol Rev. 2016;96(2):695–750. doi:10.1152/physrev.00018.2015
58. Yu H, Chen L, Lei H, et al. Infralimbic medial prefrontal cortex signalling to calbindin 1 positive neurons in posterior basolateral amygdala suppresses anxiety- and depression-like behaviours. Nat Commun. 2022;13(1):5462. doi:10.1038/s41467-022-33139-6
59. Yin JB, Liang SH, Li F, et al. dmPFC-vlPAG projection neurons contribute to pain threshold maintenance and antianxiety behaviors. J Clin Invest. 2020;130(12):6555–6570. doi:10.1172/JCI127607
60. Zhang Z, M GV, Chen L, et al. Role of prelimbic gabaergic circuits in sensory and emotional aspects of neuropathic pain. Cell Rep. 2015;12(5):752–759. doi:10.1016/j.celrep.2015.07.001
61. Gao F, Huang J, Huang G, et al. Elevated prelimbic cortex-to-basolateral amygdala circuit activity mediates comorbid anxiety-like behaviors associated with chronic pain. J Clin Invest. 2023;133(9). doi:10.1172/JCI166356.
62. Zhao YT, Deng J, Liu HM, et al. Adaptation of prelimbic cortex mediated by IL-6/STAT3/Acp5 pathway contributes to the comorbidity of neuropathic pain and depression in rats. J Neuroinflammation. 2022;19(1):144. doi:10.1186/s12974-022-02503-0
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.