")
Back to Journals » International Journal of Nanomedicine » Volume 15
Isolation and Functions of Extracellular Vesicles Derived from Parasites: The Promise of a New Era in Immunotherapy, Vaccination, and Diagnosis
Authors Khosravi M, Mirsamadi ES, Mirjalali H , Zali MR
Received 23 February 2020
Accepted for publication 10 April 2020
Published 28 April 2020 Volume 2020:15 Pages 2957—2969
DOI https://doi.org/10.2147/IJN.S250993
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Thomas Webster
Mojdeh Khosravi,1 Elnaz Sadat Mirsamadi,2 Hamed Mirjalali,3 Mohammad Reza Zali4
1Department of Pharmacy and Pharmaceutical Technology and Parasitology, University of Valencia, Valencia, Spain; 2Department of Microbiology, Faculty of Medicine, Tehran Medical Sciences, Islamic Azad University, Tehran, Iran; 3Foodborne and Waterborne Diseases Research Center, Research Institute for Gastroenterology and Liver Diseases, Shahid Beheshti University of Medical Sciences, Tehran, Iran; 4Gastroenterology and Liver Diseases Research Center, Research Institute for Gastroenterology and Liver Diseases, Shahid Beheshti University of Medical Sciences, Tehran, Iran
Correspondence: Hamed Mirjalali
Foodborne and Waterborne Diseases Research Center, Research Institute for Gastroenterology and Liver Diseases, Shahid Beheshti University of Medical Sciences, Tehran, Iran
Email [email protected]
Abstract: Experimental and epidemiological evidence shows that parasites, particularly helminths, play a central role in balancing the host immunity. It was demonstrated that parasites can modulate immune responses via their excretory/secretory (ES) and some specific proteins. Extracellular vesicles (EVs) are nano-scale particles that are released from eukaryotic and prokaryotic cells. EVs in parasitological studies have been mostly employed for immunotherapy of autoimmune diseases, vaccination, and diagnosis. EVs can carry virulence factors and play a central role in the development of parasites in host cells. These molecules can manipulate the immune responses through transcriptional changes. Moreover, EVs derived from helminths modulate the immune system via provoking anti-inflammatory cytokines. On the other hand, EVs from parasite protozoa can induce efficient immunity, that makes them useful for probable next-generation vaccines. In addition, it seems that EVs from parasites may provide new diagnostic approaches for parasitic infections. In the current study, we reviewed isolation methods, functions, and applications of parasite’s EVs in immunotherapy, vaccination, and diagnosis.
Keywords: parasites, extracellular vesicles, immunotherapy, vaccination, diagnosis
Introduction
Parasitic infections have a history as long as human living.1,2 Despite lethal infections due to some parasites such as human-infecting species of Leishmania3 and Plasmodium,4,5 Naegleria,6 Entamoeba histolytica,7 most of them, particularly helminths, live within their hosts without considerable symptoms.8,9 Parasitic infections are the cause of economic loss in developing and developed countries. Based on the latest reports, more than one billion people around the world suffer from parasitic diseases.10,11 For example, it was estimated that 219 million cases in 2017 suffered from malaria with 435,000 deaths that makes this parasitic infection a global public health problem and one of the most important debilitating infectious diseases in developing countries.12 From the economic point of view, only single malaria reduces economic growth in Africa by 1.3% per annum.12 Leishmaniasis is the second lethal parasitic disease after malaria that the lack of effective vaccine has led to serious problems in tropical regions.13,14 Therefore, the most important challenges facing with these infections are detection, and immunization.
However, studies on different applications of EVs for in parasitic diseases and the interesting roles of these micro/nano-particles in cellular and molecular biology are being dramatically increased. In the current study, we comprehensively gathered and categorized different applications of EVs in the field of parasitology that helps researchers for their future studies. For this purpose, available English electronic major libraries including: PubMed, Scopus and Web of Science were searched with keywords: “Parasite” AND “Extracellular Vesicles” AND “Immunotherapy” OR “Vaccination” OR “Diagnosis” OR “Detection”.
Parasites and the Immune System
Apart from the traditional view emphasizing on the pathogenic role of parasites, a body of evidence has demonstrated the positive consequences of facing with some parasites in the immune system homeostasis,15–18 which is famously discussed as “hygiene hypothesis”. This hypothesis was supported by studies showing less exposure to the parasite infections during early childhood may increase the susceptibility to immune disorders during adulthood.18,19 Actually, parasites, particularly helminths, can modulate the immune system through effective soluble mediators which interact with the host immune cells to escape from the immune responses.20,21 This subject has been supported by recent studies showing the manipulation of the immune system by secreted proteins and carbohydrate molecules expressed by helminths.22
During the contact between the normal immune system and foreign antigens, a plethora of proteins are produced and released from immune and non-immune cells.23,24 Accordingly, the role of some helminths and their components were practiced to ameliorate the pathological conditions and clinical manifestations of some autoimmune diseases such as asthma, inflammatory bowel diseases (IBD),25–27 multiple sclerosis (MS)28,29 and etc., using in vivo/in vitro studies. However, the most important limitations of these studies were either alive helminthic materials such as eggs or low efficacy of the derived proteins.30,31
On the other hand, during the invasion of some parasite protozoa such as Leishmania, Plasmodium, and Toxoplasma gondii, the cellular immune responses are the most important defense mechanisms. Studies on this subject have been led to engage of either total antigen or derived specific proteins of protozoan parasites as a vaccination strategy against themselves or other intracellular microorganisms.32–36
During the last decade, focuses on EVs released from parasites or immune cells sensed by parasites have been explosively increased. Up to now, several distinct types of EVs were known of which exosomes (size between 30 to150 nm, spherical shape with membrane bound) are an important subtype.37,38 Based on the growing evidence, the clinical applications of EVs in the field of parasitology are mainly categorized into three major classes.39 A) Immunotherapy by EVs to modulate the immune responses. In this application, EVs derived from helminths are mostly employed to attenuate the immunological responses during autoimmune diseases such as IBD and allergy. B) Increasing the immunity responses against some parasitic infections, also known as vaccine. This application has majorly been practiced for parasitic protozoa in both medical and veterinary researches. C) Detection of specific EVs (particularly their contents) as a promising tool for diagnosis of parasitic diseases, particularly blood protozoa (Figure 1).
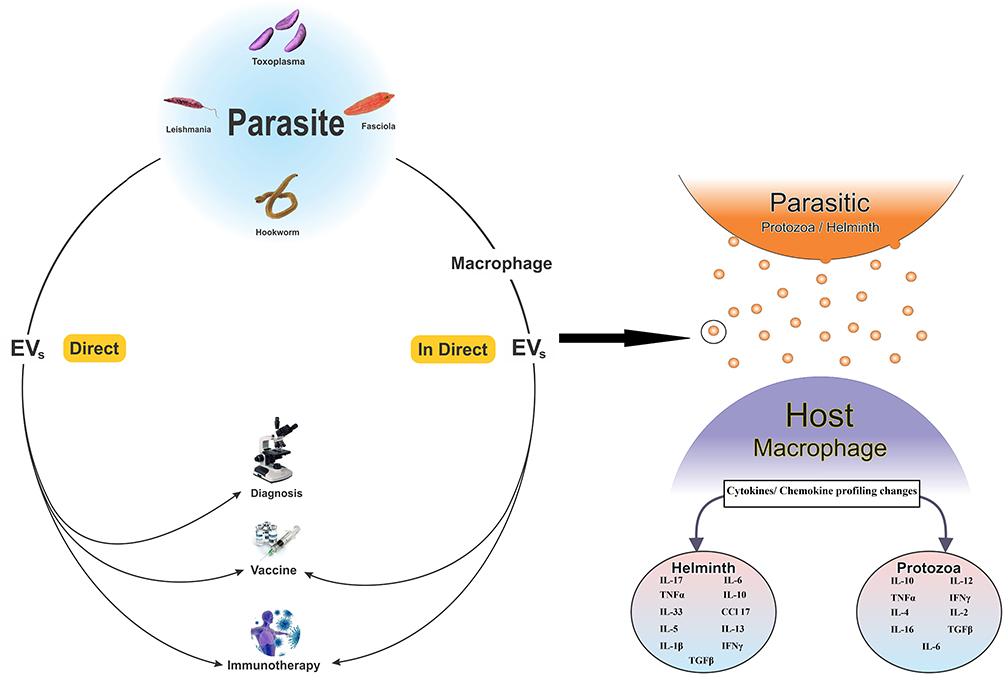 |
Figure 1 Extracellular vesicles (EVs) extracted directly from parasites or indirectly from host cells sensed by parasites. Indirectly released EVs from host cells sensed by parasites are mostly used for vaccination and immunotherapy while for diagnosis, EVs originated from parasites are a regular target. EVs can also change the cytokine/chemokine pattern in recipient cells. Protozoan parasites mostly increase the level of cytokine/chemokine pattern involved in polarization of cellular immunity while EVs from helminths usually increase the level of immunomodulatory cytokines such as IL-4 and TGFβ. However, there are studies showing elevated levels of immunomodulatory cytokines in host cells affected by EVs from parasite protozoa or vice versa. Abbreviations: IL, interleukin; TNF, tumor necrosis factor; IFN- γ, interferon-gamma; TGF-β, tumor growth factor-beta; CCL, chemokine (C-C motif) ligand. |
EVs in the parasitology studies have mostly been employed for immunization against a parasitic infection (8/17; 47.05%) while only four studies (23.53%), particularly in recent years, were carried out to use EVs for detection of parasitic infections (Table 1). In addition, ultracentrifugation more than 100,000 × g for at least 1 hr at 4ºC has been the most common technique for isolation and purification of EVs.
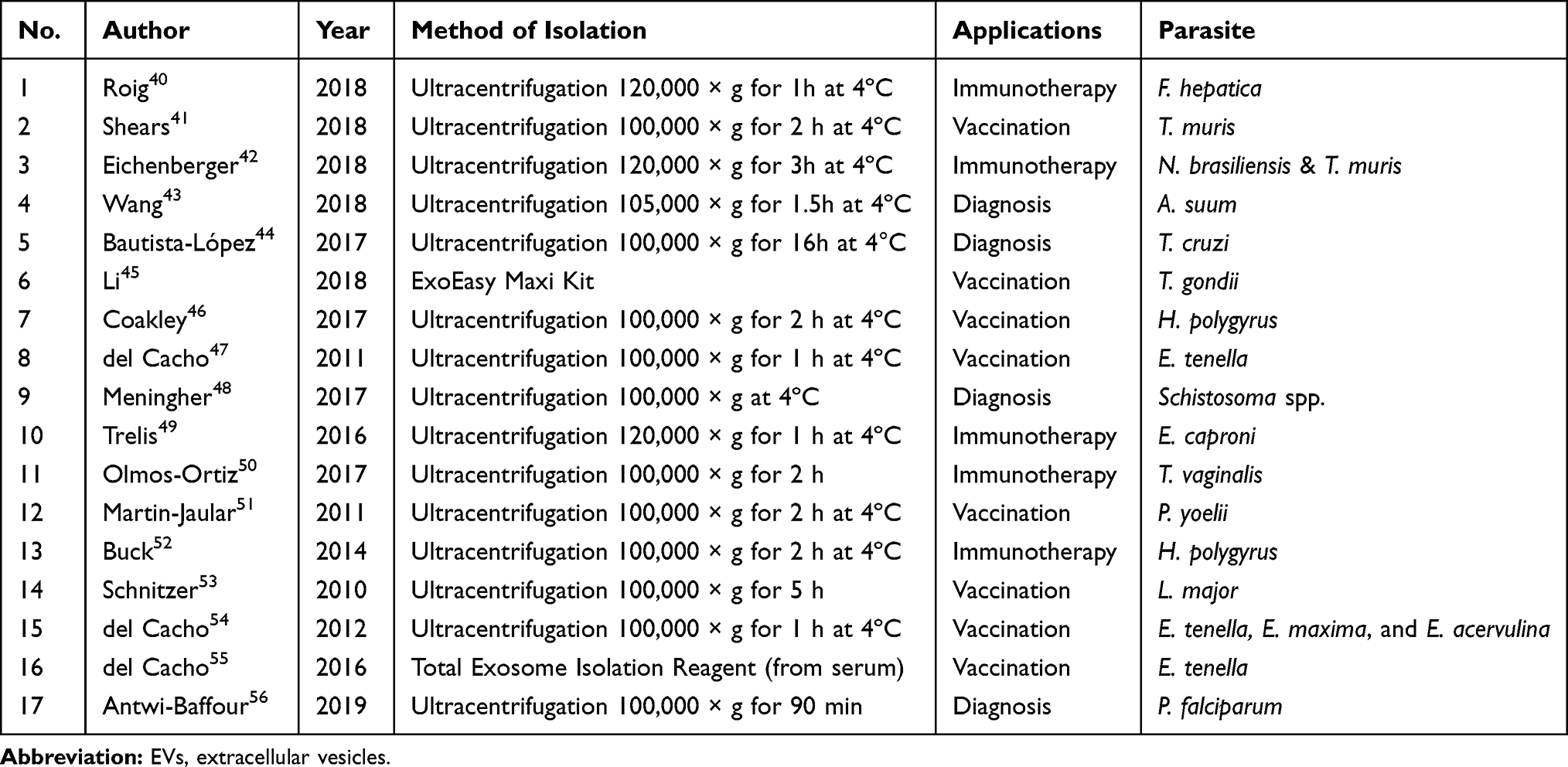 |
Table 1 All Eligible Studies Were Categorized Based on Methods of Isolation of EVs, Clinical Applications and Target Parasites |
Furthermore, IBD was the autoimmune diseases that were frequently investigated for evaluation of the effects of EVs. In most of the studies, mice were the choice of host models and laboratory chickens were mostly the choice animal model for study on Eimeria (Table 2).40–56
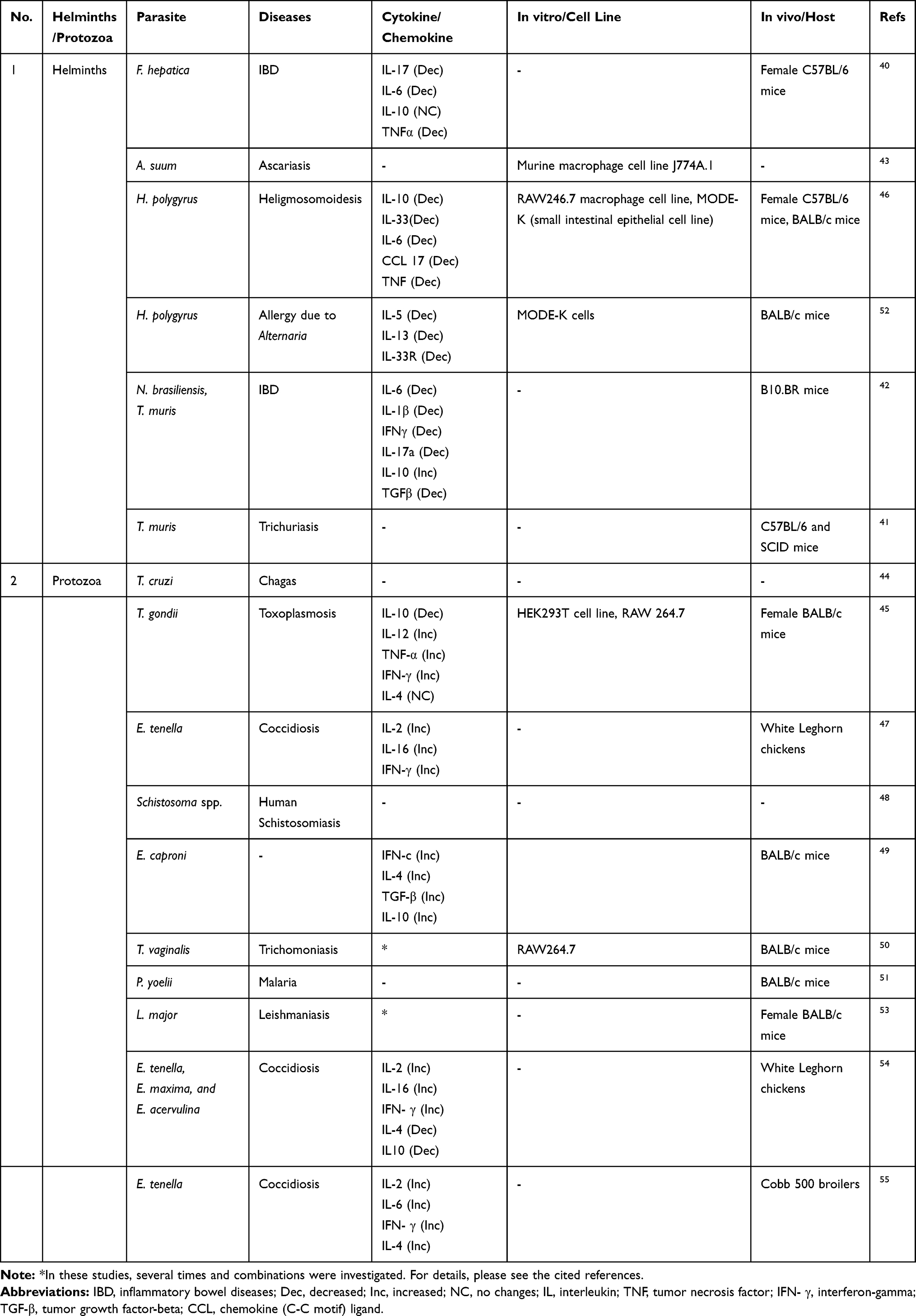 |
Table 2 Targeted Diseases, Cytokine Profile, Cell Line and Animal Models Employed in Each Study |
Parasite’s EVs
Parasites are eukaryotic microorganisms that during the time have learnt to communicate with their hosts to provide favor conditions in their niches.1,2 Although some of the parasites such as human-infecting species of Plasmodium and Leishmania, E. histolytica, and some other parasites usually hurt their hosts, the hygiene hypothesis claims that some parasites, as an old friend of humans, have played an important role in evolution, improvement and homeostasis of the immune system.31
However, it seems that parasites are mostly companied with their host via ES products.57,58 Numerous studies have explored the role of EVs released from either parasites or different types of host cells sensed by parasites in communication between parasites and host cells.59 Actually, derived EVs from parasites can manipulate the target cells via delivering the pathogenic, immunomodulatory, and genetic materials to provide a pleasant condition for surviving and multiplication of parasites.39,60 Furthermore, it was suggested that during the unpleasant environments, host cells may release EVs recruiting immune cells to defense against parasites.39 Experimental studies have demonstrated that EVs mostly contain lipid, proteins, DNA, RNA, and metabolites61–63 (Figure 2). Apparently, EVs have also played a crucial role during transferring of drug-resistant,64 virulence,65,66 and regulatory genes67–69 into the host cells.
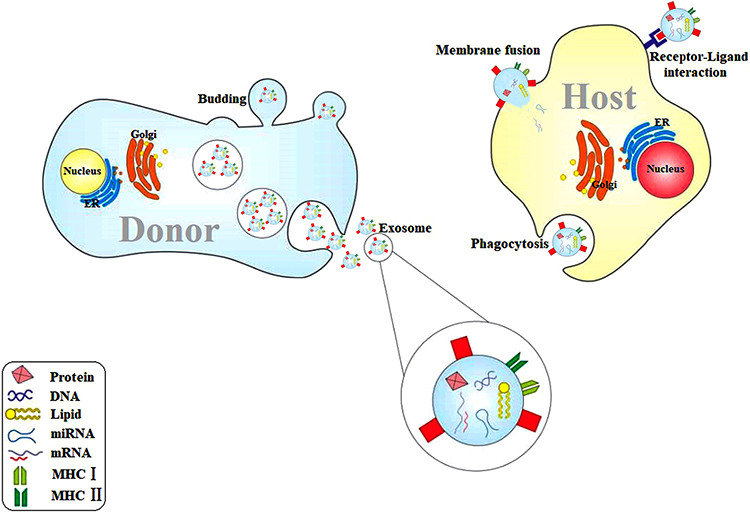 |
Figure 2 Host cells usually release EVs in response to the environmental changes such as contact with parasites or their products. EVs contain DNA, messenger RNA, MicroRNA, proteins and lipids, and express MHC I and II. EVs are released via budding and vesicle cargo. EVs may affect the recipient cell through direct fusion, receptor–ligand interaction, as well as phagocytosis. Abbreviations: EVs, extracellular vesicles; MHC, major histocompatibility complex; ER, endoplasmic reticulum; mRNA, messenger RNA; miRNA, micro RNA. |
Extracellular Vesicles: Classification and Isolation Techniques
Up to now, there is no consensus terminology on the EVs. Based on the size and origin of the mother cell, a spectrum of terms has been given to the extracted cells.70 As a traditional nomenclature that categorized EVs based on their size and origin, microvesicles, apoptotic bodies, and exosomes are the major classes with size range from 5 µM to 30 nm.39,70 During recent years, a class of EVs originally released from tumor cells, known as oncosomes, was described which has the size of 10 µm.71,72 Nevertheless, two main types of EVs, endosomes, and ectosomes, based on their biogenesis have been defined. Accordingly, endosomes are mostly smaller which as an organelle, formed within the mammalian cells. Exosomes are considered as endosomes and fused to the plasma membrane of the recipient cells. Contrary, ectosomes are formed as budding from the plasma membrane of the donor cells and included microvesicles, apoptotic cells, and oncosomes.73,74 Over the recent years, a couple of terms have been used for the EVs based on the study context and the mother cells of the released EVs. However, EVs reflect the donor cell conditions.70 Over the past decade, many isolation and purification techniques have been practiced based on the purpose of studies, available facilities, and the type of EVs.
Overviews of the isolation techniques represented that there are two major isolation methods: (1) size-dependent and (2) size-independent techniques.75–77 Size-dependent techniques comprise a broad spectrum of methods which isolate EVs based on their size, buoyant density, and physical features. In this approach, differential centrifugation ranged from 300 × g to 200,000 × g is the most frequently used method.75,78 In this technique, cell debris and dead cells sediment in 300 × g, apoptotic bodies sediment in 2000 × g, microvesicles sediment in 10,000 × g, and exosomes sediment in 100,000–200,000 × g.79 Although searching among literature reveals that most of the studies employed differential centrifugation to recover exosomes, it seems that the recovery rate and purity of this technique are insufficient.80 However, utilizing differential centrifugation followed by floatation using sucrose or iodixanol gradient improves the purity and recovery rate of EVs.77,81,82
Precipitation-based techniques using polyethylene glycol (PEG), protamine, acetate, and precipitation of proteins with organic solvent (PROSPR) are used to cover the limitations of differential centrifugation-based techniques.75,83 Although precipitation-based methods improve the recovery rate of EVs, the purity of EVs remains as a challenge.
Ultrafiltration is used to separate EVs based on their molecular weight and size, and increases recovery of EVs compared to the conventional differential centrifugation. However, this technique is not able to differentiate EVs type in a sample based on their size; therefore, the purity of EVs is still insufficient.77 To overcome the limitations of conventional ultrafiltration, size-exclusion chromatography (SEC), and asymmetrical flow field-flow fractionation (AF4) were developed.76,77 Recent studies performed by Mol et al84 and Gamez-Valero et al85 represented the higher functionality and lower alteration of EVs isolated by SEC compared to ultracentrifugation and precipitating agents in clinical practices, respectively. EVs separation based on the microfluidic technology was recently developed and practiced. This technique isolates EVs according to their size. However, this method has not been standardized and needs to be compared to the available common isolation techniques.43,86-88
Although many techniques based on the size of EVs have been developed, improved, and practiced, two major size-independent EVs isolation methods were described during the recent years. In this regard, immunoaffinity isolation and flow cytometry are two techniques which isolate EVs regarding the expressed proteins on their surface, not their physical criteria. These techniques are usually performed to characterize EVs based on the expressed proteins on their surface in a small volume of samples. However, immunoaffinity isolation followed by a magnetic isolation was described to increase the volume of isolated EVs for further analyses.89,90 There are available commercial kits which employ specific antibodies to detect CD markers and EVs proteins such as heat shock proteins (HSP).90–92 Some studies designed and developed flow cytometry-based method for detection of EVs in which the most important limitations for this technique are the particles smaller than 600 nm and low refraction index.90,93
The Major Content of EVs
It is widely accepted that EVs contain divergent molecules such as lipids, proteins, genetic materials, and even metabolites which affect the recipient cell’s functions.70 EVs released from parasites contain a complex of molecules ranged from genetic materials to functional proteins. Firstly, Couper et al94 demonstrated that microparticles with size <1 µm derived from the plasma of mice infected with P. berghei were able to activate macrophages. Although it seems that the isolated microparticles were EVs, one of the first studies which showed the presence of EVs in parasites was performed by Marcilla et al95 who demonstrated the presence of EVs in ESP of Echinostoma caproni and Fasciola hepatica. Moreover, the proteome analysis of the isolated EVs indicated the similarity of 54% and 52% of protein content of isolated EVs from E. caproni and F. hepatica with their secretome, respectively. Furthermore, the protein content of F. hepatica comprised a higher number of proteases (such as cathepsins and leucine aminopeptidase) and detoxifying enzymes compared to E. caproni. Later, Bayer-Santos et al96 described two types of EVs which were derived from the plasma membrane and within the flagellar pocket of both epimastigotes and metacyclic trypomastigotes stages of Trypanosoma cruzi, respectively. The proteome analysis showed that there was EVs content exclusive for either epimastigote or metacyclic trypomastigote, while major identified proteins were known to play crucial roles in metabolism, signaling, nucleic acid binding, parasite survival, and virulence of the parasite.
Later, a couple of small RNAs were characterized within the EVs derived from T. cruzi and it was suggested that the parasite can modulate its communication with the host cells via the small RNAs.97–99 During recent years, focusing on the non-coding small RNA content of the EVs derived from both helminths and protozoa demonstrated a couple of unique small RNAs together with common small RNAs which previously identified in hosts.100–102 Juntao Liu et al,103 determined the RNAs content of EVs released by Schistosoma japonicum using high-throughput sequencing and then evaluated up taking the EVs by peripheral blood immune cells. Accordingly, they demonstrated that approximately 32% of the RNA content was micro RNA consisted of mir-125b, mir-61, mir-3505, and a helminth-specific micro RNA, bantam. However, they concluded that S. japonicum derived EVs increased macrophage proliferation and TNF-α production in host cells via mir-125b and bantam.
The Role of EVs in Pathogenicity of Parasites
Parasite–host interaction studies have elucidated that invasive parasites usually release EVs in response to the environmental changes to provide suitable conditions for their pathogenicity. It was shown that EVs are able to modulate the immune system and carry virulence factors to the target- or the related cells in order to manipulate their life cycle using switching on/off the signaling pathways involved in cell death.104 Recently, it was reported that promastigotes of L. infantum release major surface proteases (MSP), an important virulence factor, via EVs.66
Furthermore, it was shown that the released EVs can induce transcriptional changes in the target cells, modulate the immune responses, and affect the severity of diseases. Sampaio et al105 elucidated that monocytes sensed with EVs derived from PfEMP1 transport knock-out P. falciparum-infected RBC, induced more transcriptional changes. Furthermore, it was demonstrated that the released EVs carried drug-resistant and virulence genes, and affect the expression level of some host genes. Studies in animal models showed that the released EVs from parasites may affect the pathogenicity of them. Lovo-Martins et al106 showed that EVs released from T. cruzi led to parasitemia and reduced the level of inflammatory biomarkers such as nitric oxide (NO), IL6, and TNFα. It was reported that EVs released from L. amazonensis could provide favor conditions in host cells which led to the recruitment of Th2 responses and higher parasite burden.107 However, it seems that EVs play a central role in the development of a parasite in the host cells.108
EVs as a Promising Target for Immunotherapy of Autoimmune Diseases
History of the role of parasites, particularly helminths, in the modulation of the immune system in autoimmune diseases backs to the theory firstly described by Strachan, so-called “Hygiene hypothesis”.109,110 Regarding the chronic infections due to helminth parasites, distracting the immune system seems to be the most important defense tool of these parasites. Actually, helminths modulate the host’s immunity via changing the surface antigen or releasing the immunomodulatory components. The ES components are also the well-known mechanism used by helminths to escape from the immune system.22,58 Until know, ESP of helminths were described to be able to manipulate the immune responses in humans.111–113 In fact, helminth parasites provoke the polarization of the Th2 responses that lead to the secretion of anti-inflammatory cytokines such as IL4, IL5, IL10, IL13, as well as regulatory cytokine, transforming growth factor (TGFβ).31 These unique features of helminth parasites were established based on the use of non-human- or human-infecting worms and their eggs,114,115 released ESP from helminths111,112 and specific synthesized proteins,116,117 which are originally secreted from helminths.
The most important autoimmune diseases which have been included in worm therapy plans are IBD, rheumatoid arthritis (RA), MS, diabetes type 1 and asthma.118 Despite the promising results of worm therapy in almost all of these autoimmune diseases, the major challenge was working with alive worm/egg in humans. Therefore, recent studies employed either natural proteins from the worms or recombinant proteins modeled based on their specific secreted proteins. Tissue inhibitory metalloproteinase-2 (TIMP-2), which is currently known as anti-inflammatory protein-2 (AIP-2), is a major protein secreted in ESP of hookworms that showed favorable results in the attenuation of the clinical manifestations of asthma and IBD in animal models.119
During the recent decade, it has been focused on the applications of EVs extracted from either helminth ESP or macrophages stimulated by helminths and their products to ameliorate the clinical manifestations of Th1 autoimmune diseases. EVs as a cargo carry a heterogenic complex of genetic and metabolic materials which are deployed by parasites to communicate with hosts.39,120 Recently, a growing number of studies experienced both in vitro and in vivo applications of EVs derived from helminths or immune cells stimulated with parasites to cure the symptoms of autoimmune diseases, particularly IBD, that the results of most of them have been satisficing.40,42,52 However, there is still no clinical trial to demonstrate the pleasant consequents of parasite-released EVs to control the clinical manifestations of autoimmune diseases.
EVs as a New Hope for Designing Next-Generation Vaccines
EVs-based vaccines were widely introduced, practiced, and even proved for bacterial infections and some cancer types, until now. Among bacterial infections, EVs extracted from Helicobacter pylori,121 Salmonella enterica (typhimurium),122 Neisseria meningitis,123 Staphylococcus aureus,124,125 Escherichia coli126 and Acinetobacter127 have been studied. Notably, a vaccination strategy based on EVs with trade name “Bexcero” for N. meningitis was performed in some countries.128,129 There is evidence showing that EVs released from tumor cells may control the growth of tumor cells via presenting MHC I and MHC II, and immune system stimulatory molecules.130,131 Furthermore, during recent years, bioengineered EVs using interference RNA (iRNA) have motivated scientists around the world to design a cost-benefit and effective vaccination strategy for cancers.132,133
In case of parasitic diseases, immunization with attenuated parasites or specific synthetic proteins has a long history while there are few studies focusing on the ability of EVs to induce protection against parasites. Although protective effects of EVs were reported in Trichuris muris,41 these experiences were mostly successful in case of parasitic protozoa due to the immunological pathways involved in defending against protozoa.45,47,55
Macrophages and dendritic cells, as the progeny of monocytes, stimulated with EVs are able to manage the innate and adaptive immunity against a pathogen. Macrophages and dendritic cells act as antigen-presenting cells (APCs) and are the first barrier against microbial pathogens. These cells after up taking EVs derived from parasites and presenting antigen can initiate cellular or humoral immune responses. However, it seems that shifting towards Th1 responses together with releasing some cytokines, such as IL2, IL12, IL16, and IFNγ, is the main immune system’s arms against parasitic protozoa. However, successful induction of protective immunity against Leishmania, Toxoplasma, and Eimeria suggest EVs and their contents as the next generation vaccines45,47,53,55(Figure 1).
EVs are Specific Targets for Detection of Parasitic Diseases
Rapid and reliable diagnosis of the infectious agents provides all choices on the table for clinicians for on-time interventions. Since EVs represent the features of their mother’s cell, these particles have been considered as new biomarkers for diagnosis of different types of cancers, and also detection of infectious diseases. Although it is controversial,134 based on the infrastructure of the envelope of EVs that is similar to liposomes, it seems that these particles remain stable enough to employ as a diagnostic biomarker. This feature of EVs makes them more applicable to be a biomarker than miRNAs and specific proteins.133
A meta-analysis conducted by Zhou showed that the prediction of exosomal miRNA such as miR21, miR-451a, miR-1290, and miR-638 were significantly correlated with the prognosis of solid tumors.135 EVs (in this study exosome) due to carrying oncogenic materials, were suggested as a prediction tool for human malignant mesothelioma.136 Studies on EVs for diagnosis of infectious diseases have suggested these interesting tiny particles as a biomarker for the detection of infectious diseases. Anyanwu et al137 observed human immunodeficiency virus (HIV) protein in EVs released in urine of infected patients and concluded that EVs might be a diagnostic tool for the detection of HIV. EVs as biomarkers were recently employed for parasitic infections as well. In this regard, released EVs carrying ESP of T. cruzi, from mammalian cells were suggested as reliable diagnostic tools.44 EVs for diagnosis of parasitic infections were later investigated and proposed for Ascaris43 and Schistosoma.48
However, regarding the high specificity and reliability of EVs, particularly their contents, for the detection of infectious diseases, it seems that a diagnostic panel based on EVs would be a useful, applicable and rapid method for the detection of infectious diseases.
Conclusion and Future Perspectives
During the recent decade, the number of studies that investigated the potential applications of EVs has explosively grown. The most important reason of this growth backs to the interesting results of studies on EVs. Many studies characterized the EV’s content in human- and animal-parasites. However, characterization of the specific contents such as micro RNAs and proteins can provide a reliable panel for diagnosis of parasitic diseases, particularly lethal infections. Although there are well-successful attempts for diagnosis of malaria, leishmaniasis, and schistosomiasis using EVs-based approaches, we think that there is a gap for rapid and reliable diagnosis of primary amoebic meningoencephalitis (PAM) caused by free-living amoeba (FLA), Naegleria fowleri. The disease caused by this FLA is almost always fatal and because of the rapid progress of the disease, diagnosis mostly happens post-mortem. Therefore, a rapid and reliable diagnosis panel using the parasite-specific EVs probably improves on-time intervention and prognosis.
There are only a few studies evaluating the effects of parasite-derived EVs on the host cells in vivo or in vitro. Since infections caused by parasites are mostly chronic, parasites communicate with their host for a long time. Therefore, the role of parasites-derived EVs in the modulation of the immune system seems to be an important portion of host-parasite interaction. This fact makes parasite’s EVs a terra incognita that needs to explore, particularly in the correlation with autoimmune diseases.
Furthermore, it seems that together with increasing our knowledge on the content of released EVs from parasites, new opportunities will be opened in the field of EVs bioengineering. In the other words, parasites can communicate with the hosts using their EVs; therefore, characterization of the EVs content may describe new useful parasite-specific molecules which makes packaging of EVs using some parasite-specific proteins, metabolites, and iRNA (such as micro RNA, siRNA and Inc RNA) achievable interesting target during the next years, particularly for immunotherapy and vaccination.
Acknowledgments
The authors would like to thank all the staff of the Foodborne and Waterborne Diseases Research Center for their collaboration.
Disclosure
The authors report no conflicts of interest for this work. This study was financially supported by the Research Institute for Gastroenterology and Liver Diseases, Shahid Beheshti University of Medical Sciences with grant number: 1029, and received ethical approval from the ethics committee of the university (no. IR.SBMU.RCGLD.REC.1398.015).
References
1. Cox FE. History of human parasitology. Clin Microbiol Rev. 2002;15:595–612. doi:10.1128/cmr.15.4.595-612.2002
2. Kochin BF, Bull JJ, Antia R. Parasite evolution and life history theory. PLoS Biol. 2010;8(10):e1000524. doi:10.1371/journal.pbio.1000524
3. Akhoundi M, Kuhls K, Cannet A, et al. A historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl Trop Dis. 2016;10(3):e0004349. doi:10.1371/journal.pntd.0004349
4. Moore KA, Fowkes FJI, Wiladphaingern J, et al. Mediation of the effect of malaria in pregnancy on stillbirth and neonatal death in an area of low transmission: observational data analysis. BMC Med. 2017;15(1):98. doi:10.1186/s12916-017-0863-z
5. Sypniewska P, Duda JF, Locatelli I, et al. Clinical and laboratory predictors of death in African children with features of severe malaria: a systematic review and meta-analysis. BMC Med. 2017;15(1):147. doi:10.1186/s12916-017-0906-5
6. Matanock A, Mehal JM, Liu L, Blau DM, Cope JR. Estimation of undiagnosed Naegleria fowleri primary Amebic Meningoencephalitis, United States1. Emerg Infect Dis. 2018;24(1):162–164. doi:10.3201/eid2401.170545
7. Maldonado-Barrera CA, Campos-Esparza MDR, Muñoz-Fernández L, et al. Clinical case of cerebral amebiasis caused by E. histolytica. Parasitol Res. 2012;110(3):1291–1296. doi:10.1007/s00436-011-2617-8
8. Stensvold CR, Clark CG. Current status of Blastocystis: a personal view. Parasitol Int. 2016;65(6):763–771. doi:10.1016/j.parint.2016.05.015
9. Einarsson E, Ma’ayeh S, Svard SG. An up-date on Giardia and giardiasis. Curr Opin Microbiol. 2016;34:47–52. doi:10.1016/j.mib.2016.07.019
10. Jensen K The Global Burdon of Parasitic Diseases; 2019.
11. Kuris AM. The global burden of human parasites: who and where are they? How are they transmitted? J Parasitol. 2012;98(6):1056–1064. doi:10.1645/12-90.1
12. WHO. World malaria report 2018. Geneva; 2018.
13. Desjeux P. The increase in risk factors for leishmaniasis worldwide. Trans Royal Soc Trop Med Hyg. 2001;95(3):239–243. doi:10.1016/s0035-9203(01)90223-8
14. Oryan A, Akbari M. Worldwide risk factors in leishmaniasis. Asian Pacific J Trop Med. 2016;9(10):925–932. doi:10.1016/j.apjtm.2016.06.021
15. Leonardi I, Frey I, Rogler G. Helminth therapy for organic diseases? Transl Res. 2015;166(6):586–601. doi:10.1016/j.trsl.2015.06.016
16. Zarowiecki M, Berriman M. What helminth genomes have taught us about parasite evolution. Parasitology. 2015;142(Suppl 1):S85–97. doi:10.1017/s0031182014001449
17. Maizels RM. Infections and allergy - helminths, hygiene and host immune regulation. Curr Opin Immunol. 2005;17(6):656–661. doi:10.1016/j.coi.2005.09.001
18. Santiago HC, Nutman TB. Human Helminths and allergic disease: the hygiene hypothesis and beyond. Am J Trop Med Hyg. 2016;95(4):746–753. doi:10.4269/ajtmh.16-0348
19. Ludvigsson J, Jones MP, Faresjo A. Worm infestations and development of autoimmunity in children - The ABIS study. PLoS One. 2017;12(3):e0173988. doi:10.1371/journal.pone.0173988
20. McKay DM. The therapeutic helminth? Trend Parasitol. 2009;25(3):109–114. doi:10.1016/j.pt.2008.11.008
21. Wang M, Wu L, Weng R, et al. Therapeutic potential of helminths in autoimmune diseases: helminth-derived immune-regulators and immune balance. Parasitol Res. 2017;116(8):2065–2074. doi:10.1007/s00436-017-5544-5
22. Johnston MJ, MacDonald JA, McKay DM. Parasitic helminths: a pharmacopeia of anti-inflammatory molecules. Parasitology. 2009;136(2):125–147. doi:10.1017/s0031182008005210
23. He Z, Zhu X, Shi Z, Wu T, Wu L. Metabolic regulation of dendritic cell differentiation. Front Immunol. 2019;10:410. doi:10.3389/fimmu.2019.00410
24. Okeke EB, Uzonna JE. The pivotal role of regulatory T cells in the regulation of innate immune cells. Front Immunol. 2019;10:680. doi:10.3389/fimmu.2019.00680
25. Huang X, Zeng L-R, Chen F-S, Zhu J-P, Zhu M-H. Trichuris suis ova therapy in inflammatory bowel disease: a meta-analysis. Medicine. 2018;97(34):e12087. doi:10.1097/md.0000000000012087
26. Wangchuk P, Shepherd C, Constantinoiu C, et al. Hookworm-derived metabolites suppress pathology in a mouse model of colitis and inhibit secretion of key inflammatory cytokines in primary human leukocytes. Infect Immun. 2019;87(4). doi:10.1128/iai.00851-18
27. Xu J, Liu M, Yu P, Wu L, Lu Y. Effect of recombinant Trichinella spiralis cysteine proteinase inhibitor on TNBS-induced experimental inflammatory bowel disease in mice. Int Immunopharmacol. 2019;66:28–40. doi:10.1016/j.intimp.2018.10.043
28. Correale J, Farez MF. Does helminth activation of toll-like receptors modulate immune response in multiple sclerosis patients? Front Cell Infect Microbiol. 2012;2:112. doi:10.3389/fcimb.2012.00112
29. Dixit A, Tanaka A, Greer JM, Donnelly S. Novel therapeutics for multiple sclerosis designed by parasitic worms. Int J Mol Sci. 2017;18. doi::10.3390/ijms18102141.
30. Van Kruiningen HJ, West AB. Potential danger in the medical use of Trichuris suis for the treatment of inflammatory bowel disease. Inflam Bowl Dis. 2005;11(5):515. doi:10.1097/01.mib.0000160369.47671.a2
31. Smallwood TB, Giacomin PR, Loukas A, et al. Helminth immunomodulation in autoimmune disease. Front Immunol. 2017;8:453. doi:10.3389/fimmu.2017.00453
32. Lu G, Zhou J, Zhou A, et al. SAG5B and SAG5C combined vaccine protects mice against Toxoplasma gondii infection. Parasitol Int. 2017;66(5):596–602. doi:10.1016/j.parint.2017.06.002
33. Wang JL, Zhang N-Z, Li T-T, et al. Advances in the development of anti-Toxoplasma gondii vaccines: challenges, opportunities, and perspectives. Trend Parasitol. 2019;35(3):239–253. doi:10.1016/j.pt.2019.01.005
34. Draper SJ, Sack BK, King CR, et al. Malaria vaccines: recent advances and new horizons. Cell Host Microbe. 2018;24(1):43–56. doi:10.1016/j.chom.2018.06.008
35. Iborra S, Solana JC, Requena JM, Soto M. Vaccine candidates against Leishmania under current research. Expert Rev Vaccines. 2018;17:323–334. doi:10.1080/14760584.2018.1459191
36. Seyed N, Peters NC, Rafati S. Translating observations from leishmanization into non-living vaccines: the potential of dendritic cell-based vaccination strategies against Leishmania. Front Immunol. 2018;9:1227. doi:10.3389/fimmu.2018.01227
37. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
38. Thery C. Exosomes: secreted vesicles and intercellular communications. F1000 Biol Rep. 2011;3:15. doi:10.3410/b3-15
39. Wu Z, Wang L, Li J, et al. Extracellular vesicle-mediated communication within host-parasite interactions. Front Immunol. 2018;9:3066. doi:10.3389/fimmu.2018.03066
40. Roig J, Saiz ML, Galiano A, et al. Extracellular vesicles from the helminth Fasciola hepatica prevent DSS-induced acute ulcerative colitis in a T-lymphocyte independent mode. Front Microbiol. 2018;9:1036. doi:10.3389/fmicb.2018.01036
41. Shears RK, Bancroft AJ, Hughes GW, Grencis RK, Thornton DJ. Extracellular vesicles induce protective immunity against Trichuris muris. Parasit Immunol. 2018;40(7):e12536. doi:10.1111/pim.12536
42. Eichenberger RM, Ryan S, Jones L, et al. Hookworm secreted extracellular vesicles interact with host cells and prevent inducible colitis in mice. Front Immunol. 2018;9:850. doi:10.3389/fimmu.2018.00850
43. Wang Y, Yuan W, Kimber M, Lu M, Dong L. Rapid differentiation of host and parasitic exosome vesicles using microfluidic photonic crystal biosensor. ACS Sensor. 2018;3(9):1616–1621. doi:10.1021/acssensors.8b00360
44. Bautista-Lopez NL, Ndao M, Camargo FV, et al. Characterization and diagnostic application of Trypanosoma cruzi trypomastigote excreted-secreted antigens shed in extracellular vesicles released from infected mammalian cells. J Clin Microbiol. 2017;55(3):744–758. doi:10.1128/jcm.01649-16
45. Li Y, Liu Y, Xiu F, et al. Characterization of exosomes derived from Toxoplasma gondii and their functions in modulating immune responses. Int J Nanomedicine. 2018;13:467–477. doi:10.2147/ijn.s151110
46. Coakley G, McCaskill JL, Borger JG, et al. Extracellular vesicles from a helminth parasite suppress macrophage activation and constitute an effective vaccine for protective immunity. Cell Rep. 2017;19(8):1545–1557. doi:10.1016/j.celrep.2017.05.001
47. Del Cacho E, Gallego M, Lee SH, et al. Induction of protective immunity against Eimeria tenella infection using antigen-loaded dendritic cells (DC) and DC-derived exosomes. Vaccine. 2011;29(21):
48. Meningher T, Lerman G, Regev-Rudzki N, et al. Schistosomal microRNAs isolated from extracellular vesicles in sera of infected patients: a new tool for diagnosis and follow-up of human schistosomiasis. J Infect Dis. 2017;215(3):378–386. doi:10.1093/infdis/jiw539
49. Trelis M, Galiano A, Bolado A, et al. Subcutaneous injection of exosomes reduces symptom severity and mortality induced by Echinostoma caproni infection in BALB/c mice. Int J Parasitol. 2016;46(12):799–808. doi:10.1016/j.ijpara.2016.07.003
50. Olmos-Ortiz LM, Barajas-Mendiola MA, Barrios-Rodiles M, et al. Trichomonas vaginalis exosome-like vesicles modify the cytokine profile and reduce inflammation in parasite-infected mice. Parasite Immunol. 2017;39(6):e12426. doi:10.1111/pim.12426
51. Martin-Jaular L, Nakayasu ES, Ferrer M, Almeida IC, Del Portillo HA. Exosomes from Plasmodium yoelii-infected reticulocytes protect mice from lethal infections. PLoS One. 2011;6(10):e26588. doi:10.1371/journal.pone.0026588
52. Buck AH, Coakley G, Simbari F, et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat Commun. 2014;5(1):5488. doi:10.1038/ncomms6488
53. Schnitzer JK, Berzel S, Fajardo-Moser M, Remer KA, Moll H. Fragments of antigen-loaded dendritic cells (DC) and DC-derived exosomes induce protective immunity against Leishmania major. Vaccine. 2010;28(36):
54. Del Cacho E, Gallego M, Lee SH, et al. Induction of protective immunity against Eimeria tenella, Eimeria maxima, and Eimeria acervulina infections using dendritic cell-derived exosomes. Infect Immun. 2012;80(5):1909–1916. doi:10.1128/iai.06413-11
55. Del Cacho E, Gallego M, Lillehoj HS, et al. Induction of protective immunity against experimental Eimeria tenella infection using serum exosomes. Vet Parasitol. 2016;224:1–6. doi:10.1016/j.vetpar.2016.04.043
56. Antwi-Baffour S, Malibha-Pinchbeck M, Stratton D, et al. Plasma mEV levels in Ghanain malaria patients with low parasitaemia are higher than those of healthy controls, raising the potential for parasite markers in mEVs as diagnostic targets. J Extracell Vesicles. 2020;9(1):
57. Crowe J, Lumb FE, Harnett MM, Harnett W. Parasite excretory-secretory products and their effects on metabolic syndrome. Parasite Immunol. 2017;39(5):e12410. doi:10.1111/pim.12410
58. Kahl J, Brattig N, Liebau E. The untapped pharmacopeic potential of helminths. Trend Parasitol. 2018;34(10):828–842. doi:10.1016/j.pt.2018.05.011
59. Coakley G, Maizels RM, Buck AH. Exosomes and other extracellular vesicles: the new communicators in parasite infections. Trend Parasitol. 2015;31(10):477–489. doi:10.1016/j.pt.2015.06.009
60. Zakeri A, Hansen EP, Andersen SD, Williams AR, Nejsum P. Immunomodulation by helminths: intracellular pathways and extracellular vesicles. Front Immunol. 2018;9:2349. doi:10.3389/fimmu.2018.02349
61. Abels ER, Breakefield XO. Introduction to extracellular vesicles: biogenesis, RNA cargo selection, content, release, and uptake. Cell Mol Neurobiol. 2016;36(3):301–312. doi:10.1007/s10571-016-0366-z
62. Nicolao MC, Rodriguez Rodrigues C, Cumino AC. Extracellular vesicles from Echinococcus granulosus larval stage: isolation, characterization and uptake by dendritic cells. PLoS Negl Trop Dis. 2019;13:e0007032. doi:10.1371/journal.pntd.0007032
63. Siles-Lucas M, Sánchez-Ovejero C, González-Sánchez M, et al. Isolation and characterization of exosomes derived from fertile sheep hydatid cysts. Vet Parasitol. 2017;236:22–33. doi:10.1016/j.vetpar.2017.01.022
64. Samuel P, Fabbri M, Carter DRF. Mechanisms of drug resistance in cancer: the role of extracellular vesicles. Proteomics. 2017;17(23–24):
65. Szempruch AJ, Dennison L, Kieft R, Harrington JM, Hajduk SL. Sending a message: extracellular vesicles of pathogenic protozoan parasites. Nat Rev Microbiol. 2016;14(11):669–675. doi:10.1038/nrmicro.2016.110
66. Marshall S, Kelly PH, Singh BK, et al. Extracellular release of virulence factor major surface protease via exosomes in Leishmania infantum promastigotes. Parasite Vectors. 2018;11(1):355. doi:10.1186/s13071-018-2937-y
67. Jiang L, Vader P, Schiffelers RM. Extracellular vesicles for nucleic acid delivery: progress and prospects for safe RNA-based gene therapy. Gene Ther. 2017;24(3):157–166. doi:10.1038/gt.2017.8
68. Shahabipour F, Barati N, Johnston TP, et al. Exosomes: nanoparticulate tools for RNA interference and drug delivery. J Cell Physiol. 2017;232(7):1660–1668. doi:10.1002/jcp.25766
69. Saari H, Lázaro-Ibáñez E, Viitala T, et al. Microvesicle- and exosome-mediated drug delivery enhances the cytotoxicity of Paclitaxel in autologous prostate cancer cells. J Controlled Release. 2015;220:727–737. doi:10.1016/j.jconrel.2015.09.031
70. Jaiswal R, Sedger LM. Intercellular vesicular transfer by exosomes, microparticles and oncosomes - implications for cancer biology and treatments. Front Oncol. 2019;9:125. doi:10.3389/fonc.2019.00125
71. Wendler F, Stamp GW, Giamas G. Tumor-stromal cell communication: small vesicles signal big changes. Trend Cancer. 2016;2:326–329. doi:10.1016/j.trecan.2016.05.007
72. Devhare PB, Ray RB. Extracellular vesicles: novel mediator for cell to cell communications in liver pathogenesis. Mol Aspects Med. 2018;60:115–122. doi:10.1016/j.mam.2017.11.001
73. Cocucci E, Meldolesi J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trend Cell Biol. 2015;25(6):364–372. doi:10.1016/j.tcb.2015.01.004
74. Meldolesi J. Exosomes and ectosomes in intercellular communication. Curr Biol. 2018;28(8):R435–r444. doi:10.1016/j.cub.2018.01.059
75. Konoshenko MY, Lekchnov EA, Vlassov AV, Laktionov PP. Isolation of extracellular vesicles: general methodologies and latest trends. Biomed Res Int. 2018;2018:8545347. doi:10.1155/2018/8545347
76. Carnino JM, Lee H, Jin Y. Isolation and characterization of extracellular vesicles from Broncho-alveolar lavage fluid: a review and comparison of different methods. Respiratory Res. 2019;20(1):240. doi:10.1186/s12931-019-1210-z
77. Monguio-Tortajada M, Galvez-Monton C, Bayes-Genis A, Roura S, Borras FE. Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cell Mol Life Sci. 2019;76(12):
78. Gardiner C, Vizio DD, Sahoo S, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles. 2016;5:32945. doi:10.3402/jev.v5.32945
79. Momen-Heravi F. Isolation of extracellular vesicles by ultracentrifugation. Methods Mol Biol. 2017;1660:25–32. doi:10.1007/978-1-4939-7253-1_3
80. Baranyai T, Herczeg K, Onódi Z, et al. Isolation of exosomes from blood plasma: qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS One. 2015;10(12):e0145686. doi:10.1371/journal.pone.0145686
81. Webber J, Clayton A. How pure are your vesicles? J Extracell Vesicles. 2013;2(1):19861. doi:10.3402/jev.v2i0.19861
82. Pan B-T, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–978. doi:10.1016/0092-8674(83)90040-5
83. Lobb RJ, Becker M, Wen Wen S, et al. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J Extracell Vesicles. 2015;4(1):27031. doi:10.3402/jev.v4.27031
84. Mol EA, Goumans MJ, Doevendans PA, Sluijter JPG, Vader P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomedicine. 2017;13:2061–2065. doi:10.1016/j.nano.2017.03.011
85. Gamez-Valero A, Monguió-Tortajada M, Carreras-Planella L, et al. Size-exclusion chromatography-based isolation minimally alters extracellular vesicles’ characteristics compared to precipitating agents. Sci Rep. 2016;6(1):33641. doi:10.1038/srep33641
86. Kanwar SS, Dunlay CJ, Simeone DM, Nagrath S. Microfluidic device (ExoChip) for on-chip isolation, quantification and characterization of circulating exosomes. Lab Chip. 2014;14(11):1891–1900. doi:10.1039/c4lc00136b
87. Gholizadeh S, Shehata Draz M, Zarghooni M, et al. Microfluidic approaches for isolation, detection, and characterization of extracellular vesicles: current status and future directions. Biosens Bioelectron. 2017;91:588–605. doi:10.1016/j.bios.2016.12.062
88. Guo SC, Tao SC, Dawn H. Microfluidics-based on-a-chip systems for isolating and analysing extracellular vesicles. J Extracell Vesicles. 2018;7:
89. Koliha N, Wiencek Y, Heider U, et al. A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J Extracell Vesicles. 2016;5:29975. doi:10.3402/jev.v5.29975
90. Wiklander OPB, Bostancioglu RB, Welsh JA, et al. Systematic methodological evaluation of a multiplex bead-based flow cytometry assay for detection of extracellular vesicle surface signatures. Front Immunol. 2018;9:1326. doi:10.3389/fimmu.2018.01326
91. Ghosh A, Davey M, Chute IC, et al. Rapid isolation of extracellular vesicles from cell culture and biological fluids using a synthetic peptide with specific affinity for heat shock proteins. PLoS One. 2014;9(10):e110443. doi:10.1371/journal.pone.0110443
92. Nakai W, Yoshida T, Diez D, et al. A novel affinity-based method for the isolation of highly purified extracellular vesicles. Sci Rep. 2016;6(1):33935. doi:10.1038/srep33935
93. Welsh JA, Holloway JA, Wilkinson JS, Englyst NA. Extracellular vesicle flow cytometry analysis and standardization. Front Cell Develop Biol. 2017;5:78. doi:10.3389/fcell.2017.00078
94. Couper KN, Barnes T, Hafalla JCR, et al. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010;6(1):e1000744. doi:10.1371/journal.ppat.1000744
95. Marcilla A, Trelis M, Cortés A, et al. Extracellular vesicles from parasitic helminths contain specific excretory/secretory proteins and are internalized in intestinal host cells. PLoS One. 2012;7(9):e45974. doi:10.1371/journal.pone.0045974
96. Bayer-Santos E, Aguilar-Bonavides C, Rodrigues SP, et al. Proteomic analysis of trypanosoma cruzi secretome: characterization of two populations of extracellular vesicles and soluble proteins. J Proteome Res. 2013;12(2):883–897. doi:10.1021/pr300947g
97. Bayer-Santos E, Lima FM, Ruiz JC, Almeida IC, da Silveira JF. Characterization of the small RNA content of Trypanosoma cruzi extracellular vesicles. Mol Biochem Parasitol. 2014;193(2):71–74. doi:10.1016/j.molbiopara.2014.02.004
98. Garcia-Silva MR, das Neves RF, Cabrera-Cabrera F, et al. Extracellular vesicles shed by Trypanosoma cruzi are linked to small RNA pathways, life cycle regulation, and susceptibility to infection of mammalian cells. Parasitol Res. 2014;113:285–304. doi:10.1007/s00436-013-3655-1
99. Fernandez-Calero T, Garcia-Silva R, Pena A, et al. Profiling of small RNA cargo of extracellular vesicles shed by Trypanosoma cruzi reveals a specific extracellular signature. Mol Biochem Parasitol. 2015;199(1–2):19–28. doi:10.1016/j.molbiopara.2015.03.003
100. Nowacki FC, Swain MT, Klychnikov OI, et al. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J Extracell Vesicles. 2015;4(1):28665. doi:10.3402/jev.v4.28665
101. Zhu L, Liu J, Dao J, et al. Molecular characterization of S. japonicum exosome-like vesicles reveals their regulatory roles in parasite-host interactions. Sci Rep. 2016;6(1):25885. doi:10.1038/srep25885
102. Zhu S, Wang S, Lin Y, et al. Release of extracellular vesicles containing small RNAs from the eggs of Schistosoma japonicum. Parasites & Vectors. 2016;9(1):574. doi:10.1186/s13071-016-1845-2
103. Liu J, Zhu L, Wang J, et al. Schistosoma japonicum extracellular vesicle miRNA cargo regulates host macrophage functions facilitating parasitism. PLoS Pathog. 2019;15(6):e1007817. doi:10.1371/journal.ppat.1007817
104. Castelli G, Bruno F, Saieva L, et al. Exosome secretion by Leishmania infantum modulate the chemotactic behavior and cytokinic expression creating an environment permissive for early infection. Exp Parasitol. 2019;198:39–45. doi:10.1016/j.exppara.2019.01.014
105. Sampaio NG, Emery SJ, Garnham AL, et al. Extracellular vesicles from early stage Plasmodium falciparum -infected red blood cells contain PfEMP1 and induce transcriptional changes in human monocytes. Cell Microbiol. 2018;20(5):e12822. doi:10.1111/cmi.12822
106. Lovo-Martins MI, Malvezi AD, Zanluqui NG, et al. Extracellular vesicles shed by Trypanosoma cruzi potentiate infection and elicit lipid body formation and pge2 production in murine macrophages. Front Immunol. 2018;9:896. doi:10.3389/fimmu.2018.00896
107. Barbosa FMC, Dupin TV, Toledo MDS, et al. Extracellular vesicles released by Leishmania (Leishmania) amazonensis promote disease progression and induce the production of different cytokines in macrophages and B-1 cells. Front Microbiol. 2018;9:3056. doi:10.3389/fmicb.2018.03056
108. Kotrbová A, Štěpka K, Maška M, et al. TEM ExosomeAnalyzer: a computer-assisted software tool for quantitative evaluation of extracellular vesicles in transmission electron microscopy images. J Extracell Vesicles. 2019;8(1):
109. Strachan DP. Hay fever, hygiene, and household size. BMJ. 1989;299:1259–1260. doi:10.1136/bmj.299.6710.1259
110. Rook GA, Brunet LR. Old friends for breakfast. Clin Exp Allergy. 2005;35(7):841–842. doi:10.1111/j.1365-2222.2005.02112.x
111. Finlay CM, Stefanska AM, Walsh KP, et al. Helminth products protect against autoimmunity via innate type 2 cytokines IL-5 and IL-33, which promote eosinophilia. J Immunol. 2016;196:703–714. doi:10.4049/jimmunol.1501820
112. Amdare N, Khatri V, Yadav RSP, et al. Brugia malayi soluble and excretory-secretory proteins attenuate development of streptozotocin-induced type 1 diabetes in mice. Parasit Immunol. 2015;37(12):624–634. doi:10.1111/pim.12283
113. Radovic I, Gruden-Movsesijan A, Ilic N, et al. Immunomodulatory effects of Trichinella spiralis-derived excretory–secretory antigens. Immunol Res. 2015;61:312–325. doi:10.1007/s12026-015-8626-4
114. Bruun JM, Carstensen JM, Vejzagić N, et al. OvaSpec – A vision-based instrument for assessing concentration and developmental stage of Trichuris suis parasite egg suspensions. Comput Biol Med. 2014;53:94–104. doi:10.1016/j.compbiomed.2014.07.009
115. Taghipour N, Mosaffa N, Rostami-Nejad M, et al. Syphacia obvelata: a new hope to induction of intestinal immunological tolerance in C57BL/6 Mice. Korean J Parasitol. 2017;55(4):439–444. doi:10.3347/kjp.2017.55.4.439
116. Navarro S, Pickering DA, Ferreira IB, et al. Hookworm recombinant protein promotes regulatory T cell responses that suppress experimental asthma. Sci Transl Med. 2016;8:362ra143. doi:10.1126/scitranslmed.aaf8807
117. Cho MK, Park MK, Kang SA, et al. TLR2-dependent amelioration of allergic airway inflammation by parasitic nematode type II MIF in mice. Parasit Immunol. 2015;37(4):180–191. doi:10.1111/pim.12172
118. Maizels RM. Parasitic helminth infections and the control of human allergic and autoimmune disorders. Clin Microbiol Infect. 2016;22:481–486. doi:10.1016/j.cmi.2016.04.024
119. Ferreira IB, Pickering DA, Troy S, et al. Suppression of inflammation and tissue damage by a hookworm recombinant protein in experimental colitis. Clin Transl Immunology. 2017;6(10):e157. doi:10.1038/cti.2017.42
120. de Pablos Torro LM, Retana Moreira L, Osuna A. Extracellular vesicles in chagas disease: a new passenger for an old disease. Front Microbiol. 2018;9:1190. doi:10.3389/fmicb.2018.01190
121. Keenan J, Day T, Neal S, et al. A role for the bacterial outer membrane in the pathogenesis of Helicobacter pylori infection. FEMS Microbiol Letter. 2000;182(2):259–264. doi:10.1111/j.1574-6968.2000.tb08905.x
122. Liu Q, Liu Q, Yi J, et al. Outer membrane vesicles derived from Salmonella typhimurium mutants with truncated LPS induce cross-protective immune responses against infection of Salmonella enterica serovars in the mouse model. Int J Med Microbiol. 2016;306(8):697–706. doi:10.1016/j.ijmm.2016.08.004
123. Bakke H, Lie K, Haugen IL, et al. Meningococcal outer membrane vesicle vaccine given intranasally can induce immunological memory and booster responses without evidence of tolerance. Infect Immun. 2001;69(8):
124. Choi SJ, Kim M-H, Jeon J, et al. Active immunization with extracellular vesicles derived from Staphylococcus aureus effectively protects against staphylococcal lung infections, mainly via Th1 cell-mediated immunity. PLoS One. 2015;10(9):e0136021. doi:10.1371/journal.pone.0136021
125. Wang X, Thompson CD, Weidenmaier C, Lee JC. Release of Staphylococcus aureus extracellular vesicles and their application as a vaccine platform. Nat Commun. 2018;9(1):1379. doi:10.1038/s41467-018-03847-z
126. Kim OY, Hong BS, Park K-S, et al. Immunization with Escherichia coli outer membrane vesicles protects Bacteria induced Lethality via Th1 and Th17 cell responses. J Immunol. 2013;190(8):
127. McConnell MJ, Rumbo C, Bou G, Pachon J. Outer membrane vesicles as an acellular vaccine against Acinetobacter baumannii. Vaccine. 2011;29(34):
128. Carter NJ. Multicomponent meningococcal serogroup B vaccine (4CMenB; Bexsero®): a review of its use in primary and booster vaccination. BioDrugs. 2013;27(3):263–274. doi:10.1007/s40259-013-0029-2
129. Martinon-Torres F, Safadi MAP, Martinez AC, et al. Reduced schedules of 4CMenB vaccine in infants and catch-up series in children: immunogenicity and safety results from a randomised open-label phase 3b trial. Vaccine. 2017;35(28):
130. Escola JM, Kleijmeer MJ, Stoorvogel W, et al. Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human B-lymphocytes. J Biological Chem. 1998;273(32):
131. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi:10.1084/jem.183.3.1161
132. Campos JH, Soares RP, Ribeiro K, et al. Extracellular vesicles: role in inflammatory responses and potential uses in vaccination in cancer and infectious diseases. J Immunol Res. 2015;2015:832057. doi:10.1155/2015/832057
133. Stremersch S, De Smedt SC, Raemdonck K. Therapeutic and diagnostic applications of extracellular vesicles. J Controlled Release. 2016;244:167–183. doi:10.1016/j.jconrel.2016.07.054
134. Simbari F, McCaskill J, Coakley G, et al. Plasmalogen enrichment in exosomes secreted by a nematode parasite versus those derived from its mouse host: implications for exosome stability and biology. J Extracell Vesicles. 2016;5(1):30741. doi:10.3402/jev.v5.30741
135. Zhou J, Guo H, Yang Y, Zhang Y, Liu H. A meta-analysis on the prognosis of exosomal miRNAs in all solid tumor patients. Medicine. 2019;98(16):e15335. doi:10.1097/md.0000000000015335
136. Greening DW, Ji H, Chen M, et al. Secreted primary human malignant mesothelioma exosome signature reflects oncogenic cargo. Sci Rep. 2016;6(1):32643. doi:10.1038/srep32643
137. Anyanwu SI, Doherty A, Powell MD, et al. Detection of HIV-1 and human proteins in urinary extracellular vesicles from HIV+ patients. Adv Virol. 2018;2018:7863412. doi:10.1155/2018/7863412
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.