")
Back to Journals » Drug Design, Development and Therapy » Volume 8
Involvement of NF-ΚB and HSP70 signaling pathways in the apoptosis of MDA-MB-231 cells induced by a prenylated xanthone compound, α-mangostin, from Cratoxylum arborescens
Authors Ibrahim MY, Mohd Hashim N, Mohan S, Abdulla M, Abdelwahab S, Kamalidehghan B, Ghaderian M, Dehghan F, Ali LZ, Karimian H, Yahayu M, Ee GC, Farjam AS, Mohd Ali H
Received 21 April 2014
Accepted for publication 25 June 2014
Published 6 November 2014 Volume 2014:8 Pages 2193—2211
DOI https://doi.org/10.2147/DDDT.S66574
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
The paper has been retracted.
Mohamed Yousif Ibrahim,1 Najihah Mohd Hashim,1 Syam Mohan,2 Mahmood Ameen Abdulla,3 Siddig Ibrahim Abdelwahab,2 Behnam Kamalidehghan,1 Mostafa Ghaderian,1,4 Firouzeh Dehghan,1,5 Landa Zeenelabdin Ali,1 Hamed Karimian,1 Maizatulakmal Yahayu,6 Gwendoline Cheng Lian Ee,7 Abdoreza Soleimani Farjam,8 Hapipah Mohd Ali9
1Department of Pharmacy, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia; 2Medical Research Centre, Jazan University, Jazan, Saudi Arabia; 3Department of Molecular Medicine, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia; 4Epigenetics Lab, HIR Building, Institute of Biological Sciences, Faculty of Science, University of Malaya, Kuala Lumpur, Malaysia; 5Department of Physiology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia; 6Department of Bioproduct Research and Innovation, Institute of Bioproduct Development (IBD), Universiti Teknologi Malaysia, UTM Johor Bahru, Johor, Malaysia; 7Department of Chemistry, Faculty of Science, Universiti Putra Malaysia (UPM), Serdang, Selangor, Malaysia; 8Institute of Tropical Agriculture, Universiti Putra Malaysia (UPM), Serdang, Selangor, Malaysia; 9Department of Chemistry, University of Malaya, Kuala Lumpur, Malaysia
Background: Cratoxylum arborescens has been used traditionally in Malaysia for the treatment of various ailments.
Methods: α-Mangostin (AM) was isolated from C. arborescens and its cell death mechanism was investigated. AM-induced cytotoxicity was observed with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Acridine orange/propidium iodide staining and annexin V were used to detect cells in early phases of apoptosis. High-content screening was used to observe the nuclear condensation, cell permeability, mitochondrial membrane potential, and cytochrome c release. The role of caspases-3/7, -8, and -9, reactive oxygen species, Bcl-2 and Bax expression, and cell cycle arrest were also investigated. To determine the role of the central apoptosis-related proteins, a protein array followed by immunoblot analysis was conducted. Moreover, the involvement of nuclear factor-kappa B (NF-ΚB) was also analyzed.
Results: Apoptosis was confirmed by the apoptotic cells stained with annexin V and increase in chromatin condensation in nucleus. Treatment of cells with AM promoted cell death-transducing signals that reduced MMP by downregulation of Bcl-2 and upregulation of Bax, triggering cytochrome c release from the mitochondria to the cytosol. The released cytochrome c triggered the activation of caspase-9 followed by the executioner caspase-3/7 and then cleaved the PARP protein. Increase of caspase-8 showed the involvement of extrinsic pathway. AM treatment significantly arrested the cells at the S phase (P<0.05) concomitant with an increase in reactive oxygen species. The protein array and Western blotting demonstrated the expression of HSP70. Moreover, AM significantly blocked the induced translocation of NF-ΚB from cytoplasm to nucleus.
Conclusion: Together, the results demonstrate that the AM isolated from C. arborescens inhibited the proliferation of MDA-MB-231 cells, leading to cell cycle arrest and programmed cell death, which was suggested to occur through both the extrinsic and intrinsic apoptosis pathways with involvement of the NF-ΚB and HSP70 signaling pathways.
Keywords: mitochondria, protein array, caspase-3/7
Corrigendum for this paper has been published
Introduction
Breast cancer is one of the most deadly cancers affecting women worldwide because of its high rates of incidence and mortality. According to the American Cancer Society (ACS), breast cancer is reported to account for 26% of all new cancer cases, which is the highest percentage of all the cancers in American women.1 In Malaysia, the National Cancer Registry (NCR) reported that one in 20 Malaysian women are at risk of acquiring breast cancer in their lifetime.2 This incidence rate is still considered low compared to one in eight women in Europe and the United States.3 The causes due to which up to 70% of breast cancer development occur in women is reported to be of environmental factors and lifestyle. The remaining 30% may be attributed to genetic factors.4,5
Among the cancer treatment strategies, radiation therapy became a valuable tool for the control of local and regional diseases after 1960 with the invention of the linear accelerator, but, like surgery, radiation therapy alone cannot enucleate metastatic cancer. Nevertheless, the success of the treatment depends on the capacity to reach every organ in the body of patients. Drugs, biological molecules, and immune-mediated remedies have therefore become the focus of current efforts to cure cancer. Chemotherapy is defined as a mode of treatment in which drugs are used to retard or destroy the growth of malignant cells. These drugs are either used alone or in combination with one or more drugs. For many years, tamoxifen has been considered the drug of choice for the cure of estrogen-dependent cases. The antitumor effect of tamoxifen is due to the arrest of growth and prompting of apoptosis,6 which is mediated by linkage to the intracellular estrogen receptor on the breast cancer cells; blocking of steroid hormone action;7 inhibition of protein kinase C;8 and its binding to calmodulin.9
However, there are various side effects, including development of liver cancer, increased blood clotting, retinopathy, and corneal opacities upon performing chemotherapy with tamoxifen.10 Due to these side effects, the use of herbs as an alternative therapy has attracted a great deal of interest by virtue of the lower toxicity and cost benefits.11 Many clinical attempts using herbs or natural products are being conducted around the world.12 Furthermore, some herbs and their constituents have been reported to suppress the proliferation of cancer cells directly. Thus, the use of herbs could be a more effective strategy in curing cancer patients.13
Cratoxylum arborescens (Vahl) Blume (Figure 1A and B) is a traditional herb belonging to the Guttiferae family, and its natural range of distribution includes Malaysia, South Myanmar (Burma), Sumatra, and Borneo.14 This plant is used traditionally as a cure for fever, cough, diarrhea, and other ailments.15 The phytochemicals found in C. arborescens include a pharmacologically superior class of phytochemical, xanthones.14,16 α-Mangostin (AM) (Figure 1C) is one of the major xanthones extracted from the stem bark of this plant.17 AM possesses a wide spectrum of biological activities, which includes anti-inflammatory,18,19 cardioprotective,20 antitumor,21,22 antidiabetic,23 antibacterial,24 antifungal,25 antioxidant,18,26 antiparasitic,27 and anti-obesity28 properties.
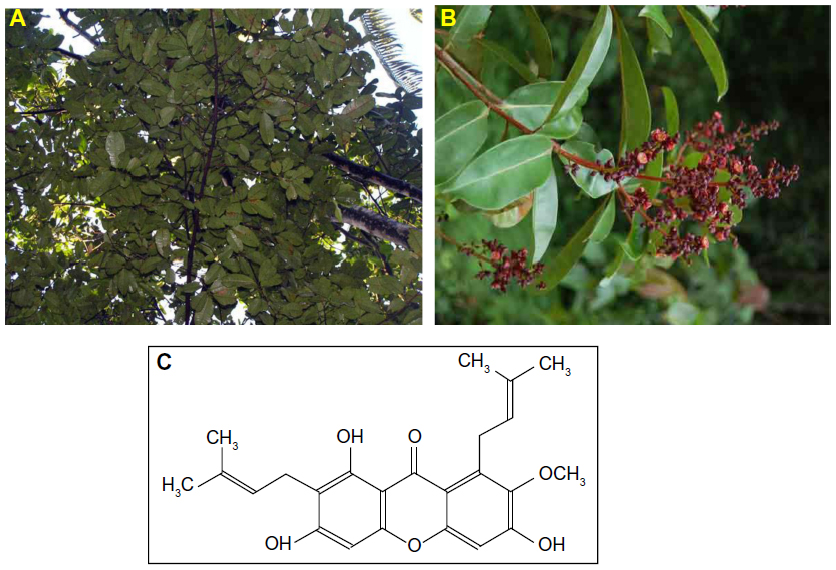 | Figure 1 Cratoxylum arborescens (Guttiferae). |
The breast cancer cell line MDA-MB-231 was isolated in 1974 from a pleural effusion of a patient with disseminated disease relapsing several years after removal of her primary tumor.29 It is used as a model of estrogen receptor-negative and HER-2/neu-negative breast cancers. The cell line is highly aggressive both in vitro and in vivo.30
In this study, we evaluated the apoptotic cell death mechanism prompted by AM on breast cancer using MDA-MB-231 cells as an in vitro model.
Materials and methods
Plant materials
The stem bark of C. arborescens (Guttiferae) was collected from wild trees growing in Malaysia, in June 2009. A voucher specimen was deposited at the Herbarium, Department of Biology, University Putra Malaysia, Serdang, Malaysia.
Extraction and isolation of AM from C. arborescens
The finely ground, air-dried stem bark of C. arborescens (1.0 kg) was extracted consecutively with hexane, chloroform, and methanol to obtain 6.12, 28.18, and 40.27 g of dark, viscous semisolid material on solvent removal, respectively. The hexane extract was chromatographed over a vacuum column and eluted with solvent of gradually increasing polarity to get 26 fractions of 200 mL each. After extensive fractionation and purification, fractions 14–20 yielded AM (Figure 1C).
Identification of AM
The melting point of AM was between 181°C−182°C. Ultraviolet MeOH λmax nm (log ε): 390 (2.41), 358 (3.99), 316 (3.99), and 238 (2.65). Infrared vmax cm−1 (KBr): 3,369 (OH), 2,934 (CH), 1,608 (C=C), 1,462, and 1,286. Electron ionization mass spectrometry m/z (% intensity): 410 (43.06), 395 (6.14), 379 (1.61), 354 (25.77), 339 (100.00), 311 (32.57), 296 (12.89), 285 (18.90), 257 (6.46), and 162 (14.16). Proton nuclear magnetic resonance (500 MHz, acetone-d6): δ 13.79 (OH-1), 9.62 (OH-6), 9.52 (OH-3), 6.81 (s, 1H, H-5), 6.38 (s, 1H, H-4), 5.26 (t, J=6.85 Hz, 2H, H-12, and H-17), 4.12 (d, J=6.85 Hz, 2H, H-11), 3.78 (OMe-7), 3.35 (d, J=8.00 Hz, 2H, H-16), 1.82 (s, 3H, Me-14), 1.71 (s, 3H, Me-19), and 1.64 (s, 6H, Me-15, and Me 20). Carbon-13 nuclear magnetic resonance (125 MHz, acetone-d6): δ 182.0 (C-9), 162.1 (C-4a), 160.9 (C-1), 156.6 (C-10a), 155.4 (C-6), 154.9 (C-3), 143.6 (C-7), 137.3 (C-8), 130.6 (C-18 and C-13), 123.9 (C-12), 122.6 (C-17), 111.2 (C-8a), 110.2 (C-2), 102.8 (C-9a), 101.9 (C-5), 92.3 (C-4), 62.5 (OMe-7), 26.1 (C-11), 25.1 (C-15 and C-20), 21.1 (C-16), 17.5 (C-14), and 17.1 (C-19).
Cell culture
Normal breast cells, MCF-10A, and human mammary cancer cells, MDA-MB-231 (estrogen-negative cells which are isolated from pleural effusions of a breast cancer patient), were acquired from the American Type Culture Collection ([ATCC] Manassas, VA, USA) and then kept at 37°C in an incubator with 5% CO2 saturation. They were grown in Roswell Park Memorial Institute (RPMI)-1640 medium (PAA Laboratories, Cölbe, Germany) together with 10% fetal bovine serum (FBS).
Antiproliferative effect of AM on MDA-MB-231 cells
The inhibitory effect of AM was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, in which 1×105 of MDA-MB-231 cells/mL were seeded in triplicate in 96-well plates and kept for 24 hours at 37°C with 5% CO2 saturation. After 24 hours’ incubation, a serial dilution for different concentrations of AM was prepared and transferred to the MDA-MB-231 cells and incubated for 24 hours in 37°C and 5% CO2; 20 μL of MTT solution (5 mg/mL) was added to the treated cells in a dark place, covered with foil, and incubated for 4 hours. All media were discharged and a total of 100 μL volume of dimethyl sulfoxide (DMSO) was poured into each well until the purple formazan crystals dissolved. The plate was measured using a microplate reader at absorbance 570 nm. The experiment was conducted in triplicate to evaluate the IC50 (half-maximal inhibitory concentration) for AM against the MDA-MB-231 cell line.
Acridine orange (AO)/propidium iodide (PI) double staining using fluorescent microscopy
MDA-MB-231 cells were quantified using AO and PI double staining in conformity with the standard procedures, and inspected under a fluorescence microscope with Q Fluoro software (Leica Microsystems, Wetzlar, Germany). Concisely, the treatment was conducted in a 25 mL culture flask, in which 1×105 cells/mL MDA-MB-231 were seeded and treated with different concentrations of AM (5, 10, and 20 μg/mL) for 24 hours. After 24 hours, the cells were centrifuged at 1,800 rpm for 5 minutes to discharge the old medium; then, cool phosphate-buffered saline (PBS) was used to wash the cell two times before staining with AO/PI. Equal amounts (10 μL [10 μg/mL]) of fresh AO and PI were prepared and covered from light to stain the cell onto a glass slide, which was covered with a cover slip. The slides were viewed under a fluorescence microscope 30 minutes prior to the fluorescence fading.
Assay of the apoptotic rate by annexin V–fluorescein isothiocyanate (FITC) staining
MDA-MB-231 (concentration of 1×105 cells/mL) was exposed to 5, 10, and 20 μg/mL concentrations of AM, and the annexin V assay was executed with the use of the BD Pharmingen™ Annexin V-FITC Apoptosis Detection Kit (APOAlert® Annexin V; Clon Tech, Mountain View, CA, USA). The treated cells were centrifuged for 5 minutes at 1,800 rpm to eliminate the media. Subsequently, cells were washed with 1× binding buffer provided by the manufacturer. Then, the cells were resuspended in 200 μL of binding buffer; 5 μL of annexin V and 10 μL of PI (Sigma-Aldrich Co, St Louis, MO, USA) were added before incubating at 37°C in the dark for 15 minutes. The binding buffer raised the reaction volume to 500 μL for the flow cytometric analysis. Flow cytometric analysis was conducted utilizing FACSCanto™ II Flow cytometry b (BD Biosciences, San Jose, CA, USA). The DMSO-treated (0.1%, v/v) cells were employed as the control.
Nuclear morphology, membrane permeability, mitochondrial membrane potential (ΔÈm) (MMP), and cytochrome c release analysis
A Multiparameter Cytotoxicity 3 Kit (Thermo Fisher Scientific, Waltham, MA, USA) was employed as formerly defined at length.31 The kit facilitates concurrent measurements of six parameters at the same time for the same cell population. Chromatin condensation, morphological change, detection of the MMP, release of cytochrome c, and cell permeability were used to test the loss of the MDA-MB-231 cells. MDA-MB-231 (1×105 cells/mL) was seeded into 96 wells plate (Molecular Devices LLC, Sunnyvale, CA, USA) and treated with AM (5, 10, and 20 μg/mL). After 24 hours, the plates were processed according to the kit procedure and analyzed using the ArrayScan® high-content screening (HCS) system (Cellomics). The images and the intensity data were reading the texture of the fluorescence inside each of the cells, together with the typical fluorescence of the cell population inside the well. The experiment was done in triplicate.
Immunofluorescence analysis of Bax/Bcl-2
A total of 5×103 MDA-MB-231 cells were seeded in a 96-well plate (genetix), followed by 5, 10, and 20 μg/mL of AM treatment. Then, the cells were fixed for 15 minutes at 37°C by 4% paraformaldehyde after rinsing two times with PBS. The cells were treated using a blocking buffer for 60 minutes’ incubation in 0.03% Triton X-100/PBS and normal serum before the cells were rinsed once more with PBS. The diluted primary antibody solution containing 1× PBS/1% BSA/0.3% Triton X-100 was added after the aspiration of the blocking buffer. The cells underwent incubation overnight at 4°C. Bcl-2 and Bax fluorochrome-conjugated secondary antibody diluted (Santa Cruz Biotechnology Inc., Dallas, TX, USA) in antibody dilution or in PBS only was added to the cells and incubated for 1 hour. After rinsing three times in PBS, the cells were treated with 2-(4-amidinophenyl)-1H -indole-6-carboxamidine (DAPI) and examined using the CellReporter™ cytofluorimeter system (Gentix/Molecular Devices LLC, Sunnyvale, CA, USA).
Bioluminescent assay of caspases-3/7, -8, and -9
A concentration-dependent study of the activity of caspases -3/7, -8, and -9 was determined using luminescence-based assay (Caspase-Glo™ 3/7, Caspase-Glo™ 8, and Caspase-Glo™ 9 Assay; Promega Corporation, Fitchburg, WI, USA). The cells were seeded in 96-well plates in 50 μL of RPMI-1640 together with 10% FBS and incubated for 24 hours. Then, the cells were treated with AM 5, 10, and 20 μg/mL and incubated for 24 hours at 37°C with 5% CO2. After 24 hours, 100 μL of the assay reagent was added and the plate was incubated for 1 hour at 37°C. Luminescence was calculated using a Tecan Infinite®200 Pro (Tecan, Männedorf, Switzerland) microplate reader.
Western blot analysis
MDA-MB-231 cells were cultured in a 25 mL flask (TPP Brand, Trasadingen, Switzerland) and treated with AM 5, 10, and 20 μg/mL. The cells were collected, washed with cool PBS, and mixed with a lysis buffer (50 mM Tris hydrochloride pH 8.0, 120 mM NaCl, 0.5% NP-40, 1 mM phenylmethanesulfonyl fluoride) to isolate the total proteins from the cells. A 40 μg measure of the extracted protein was separated using 12% SDS-PAGE, then moved to a polyvinylidenedifluoride membrane (Bio-Rad Laboratories Inc., Hercules, CA, USA), and blocked with 5% nonfat milk in a tris-buffered saline–Tween buffer 7 (0.12 M Tris-base, 1.5 M NaCl, 0.1% Tween 20) for 30 minutes at room temperature, and incubated with the appropriate primary antibody overnight at 4°C, then incubated with alkaline phosphatase-conjugated secondary antibody for 30 minutes at room temperature, and then washed in tris-buffered saline and Tween buffer 20. The primary antibodies, β-actin (sc-130300), Bax (sc-20067), HSP70 (sc-69705), proliferating cell nuclear antigen (PCNA) (sc-25280), nuclear factor-kappa B (NF-κB)/P65 (sc-398442), cytochrome c (sc-13560), caspase-7 (sc-81660), caspase-8 (sc-81657), and caspase-9 (sc-56073) were purchased from Santa Cruz Biotechnology Inc., but Bcl2 (ab38629) and Poly (ADP-ribose) polymerase (PARP) (ab4830) were purchased from Abcam (Cambridge, UK). The membranes were then incubated for 1 hour at room temperature with alkaline phosphatase (AP)-conjugated goat anti-mouse or goat anti-rabbit secondary antibodies in a ratio of 1:5,000 and then washed twice with TBST for 10 minutes three times on an orbital shaker. Then, the blots were developed using the BCIP/NBT (Santa Cruz Biotechnology Inc.) for sensitive colorimetric detection to quantify the target protein band.
Determination of the intracellular reactive oxygen species (ROS) level
ROS was determined with 2′,7′-dichlorofluorescin diacetate (DCFH-DA).31 Concisely, 10 mM DCFH-DA stock solution (in methanol) was diluted 500-fold in Hank’s Balanced Salt Solution with no serum or further additives to yield a 20 μM working solution. After exposure to AM for 24 hours, the cells in the 96-well black plate were rinsed two times with Hank’s Balanced Salt Solution before incubation in a 100 μL working solution of DCFH-DA at 37°C for 30 minutes. Using a fluorescence microplate reader (Tecan Infinite®200 Pro), the result was determined at 485 nm excitation and 520 nm emission.
DNA content and cell cycle phase distribution
MDA-MB-231 cells at a concentration of 1×105 cells/mL were seeded into a 25 mL flask (TPP Brand) containing RPMI-1640 medium augmented with 10% FBS and dosed with AM 5, 10, and 20 μg/mL. After incubation for 24 hours, the cells were collected, centrifuged to remove the old medium, and washed two times using warm PBS, then the pellet was fixed with 90% cool ethanol (kept at −20°C for 1 day before use) and kept at 4°C overnight. After incubation, the cells were spun down for 5 minutes at 1,800 rpm, the ethanol was removed, and the pellets were washed with PBS to clear ethanol from the fixed cells. An amount of 600 μL warmed PBS was added and mixed gently. Then, 25 μL RNase A in a concentration of 10 μg/mL and 50 μL of PI at a concentration of 1 μg/mL were decanted into the pellet and preserved in a dark place for 30 minutes. The DNA content of the cells was then examined using a FACSCanto II Flow cytometry by studying at least 10,000 cells per sample. The percentage of cells in the G1, S, and G2 phases were investigated using Diva software (BD Biosciences, San Jose, CA, USA).
Immunofluorescence of NF-κB activity
HCS was utilized to assess the inhibitory effects of AM on TNF-α-induced NF-κB activation, ie, nuclear translocation of NF-κB. The experiments were conducted according to the company’s instructions for the NF-κB activation kit (Cellomics). An ArrayScan reader was utilized to measure the variance between the strength of the nuclear and cytoplasmic NF-κB-associated fluorescence, and reported as a translocation parameter.
Human apoptosis proteome profiler array
To examine the pathways through which AM prompts apoptosis, we determined the apoptosis-related proteins utilizing the Proteome Profiler Array (RayBio® Human Apoptosis Antibody Array Kit; Raybiotech, Norcross, GA, USA), in accordance with the manufacturer’s instructions. In short, the cells were dosed with 20 μg/mL AM. Three hundred micrograms of proteins from each sample were incubated overnight with the human apoptosis array. The data from the apoptosis array were measured by scanning the membrane using a Biospectrum AC ChemiHR 40 (UVP, Upland, CA, USA) and examination of the file of the array image was conducted using Image J analysis software in accordance with the instructions of the company.
Statistical analysis
The results were reported as the mean ± standard deviation for at least three analyses for each sample. The normality and homogeneity of variance assumptions were checked, and statistical analysis was performed according to the SPSS 16.0 package and GraphPad Prism 5.0 software. Analysis of variance (ANOVA) was performed using the one-way ANOVA test followed by Tukey’s post hoc HSD multiple comparison tests. The type 1 error level was set at P<0.05 for all tests.
Results
AM inhibits the growth of MDA-MB-231 cells
The cytotoxic effects of AM on the MDA-MB-231 cells were ascertained by means of MTT assay. As shown in Table 1, AM inhibited the growth of MDA-MB-231 cells and significant inhibition was exhibited at IC50 9.2±0.21 after 24 hours. Tamoxifen, which was used as standard control, showed an IC50 of 2.92 μg/mL. Meanwhile, AM did not exhibit any cytotoxicity toward the normal breast cell line MCF-10A with IC50 >30 μg/mL.
AO/PI double staining cell morphological analysis
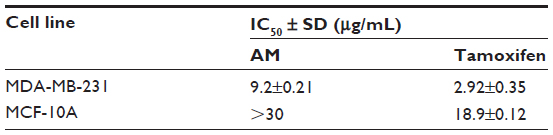 | Table 1 concentrations of AM and tamoxifen |
To determine the viable cells, as well as those in early apoptosis, late apoptosis, and secondary necrosis, the cells were counted using a fluorescence microscope. A total of 200 cells were counted arbitrarily and differentially, along with the negative control, which was untreated. Early apoptosis was detected using AO within the fragmented DNA with a bright green fluorescence. In addition, the green nuclear structure of the control cells was shown to be intact (Figure 2A). Following treatment with 5 μg/mL AM, moderate apoptosis was observed by blebbing and nuclear chromatin condensation (Figure 2B). In addition, in the later phases of apoptosis, alterations, including a reddish-orange color due to the binding of AO to the denatured DNA, were noted after 10 and 20 μg/mL of AM treatment (Figure 2C and D). The results indicated that the AM produced morphological features that are linked to apoptosis in a concentration-dependent manner. A statistically significant (P<0.05) difference was noted in the cell population through the differential recording of treated cells (200-cell population) (Figure 2E).
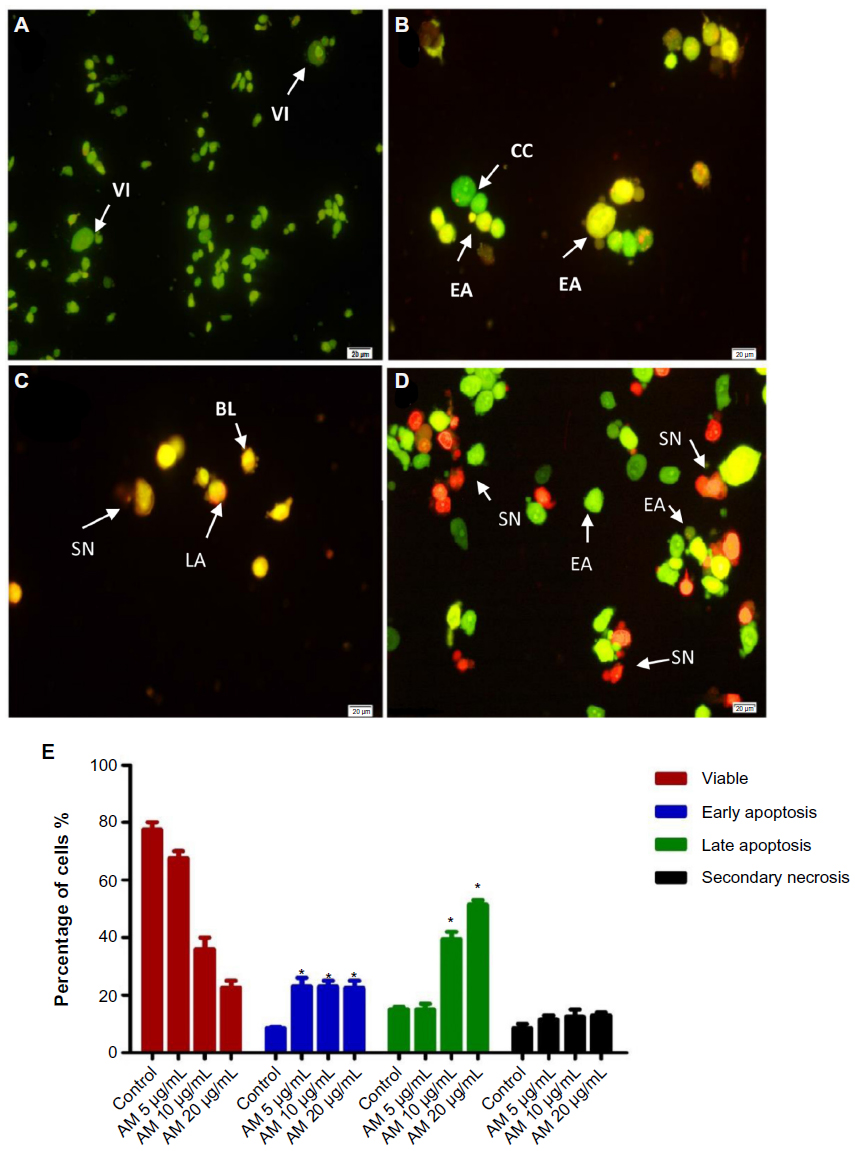 | Figure 2 Fluorescent micrographs of acridine orange and propidium iodide double-stained MDA-MB-231 cells. |
Effect of AM on the ratio of apoptotic cells by annexin V–FITC staining
The flow cytometric analysis was used to confirm the induced apoptotic effect of AM. As shown in Figure 3, the annexin V assay revealed that the apoptotic induction in MDA-MB-231 cells began after being exposed to AM at 5 μg/mL. For the untreated control, 98.2% of cells were viable, 1.8% were in the early stages of apoptotic induction, and no cells were in the late stages of apoptosis/dead (Figure 3A). At 5 μg/mL treatment, the early apoptotic population (annexin V- positive, PI-negative) significantly (P<0.05) increased from 1.8% to 3.2% (Figure 3B). At 10 μg/mL incubation, the early apoptotic population (annexin V-positive, PI-negative) significantly (P<0.05) raised to 6.5% (Figure 3C). After 20 μg/mL exposure, the number of viable cells decreased to 49.6% with a concomitant increase in both early apoptotic and late stages of apoptotic/dead cell (positive for both annexin V and PI) populations to 9.9% and 20.3%, respectively (Figure 3D). These results indicated that the AM allowed the translocation of phosphatidylserine to occur and hence induced early apoptotic induction in MDA-MB-231 cells.
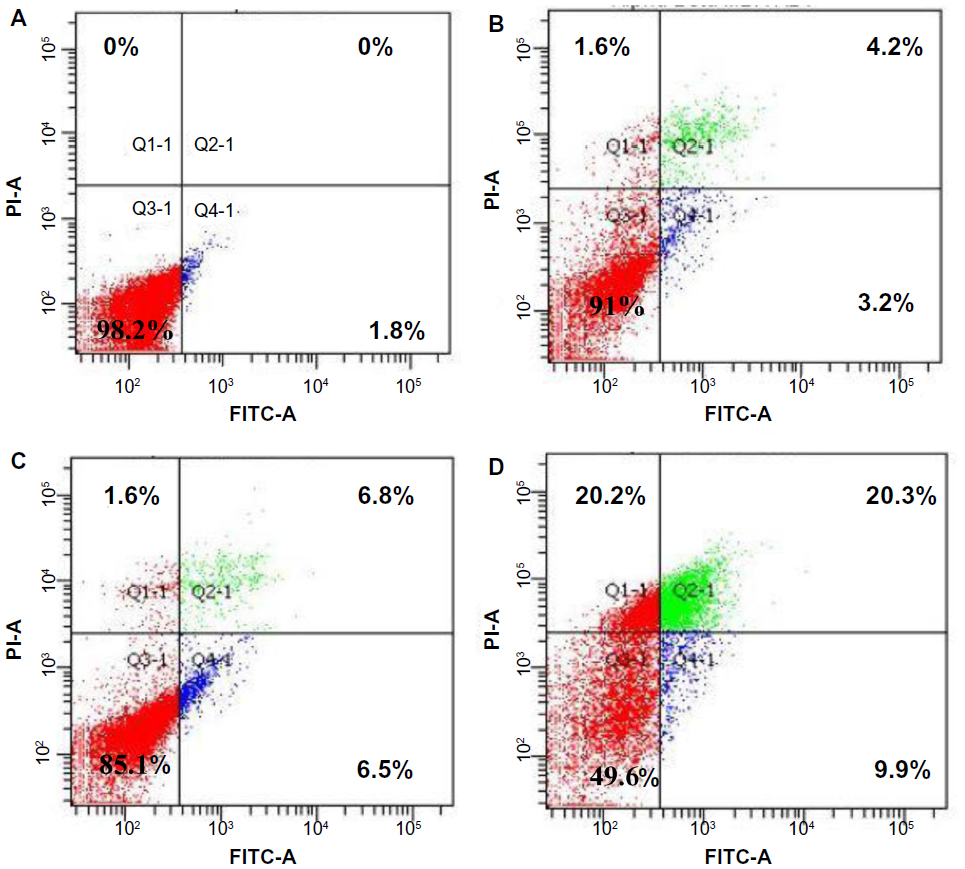 | Figure 3 The effect of AM on early apoptosis of MDA-MB-231 cells. |
AM-induced apoptosis in MDA-MB-231 cells
To verify the incidence of apoptosis, we inspected the alterations in the nuclear morphology of the MDA-MB-231 cells by determining the nuclear condensation and fragmentation hallmark for apoptosis (Figure 4A). The Hoechst 33342 staining indicated that cells exhibited nuclear condensation with 20 μg/mL AM treatment. The intensity of the nucleus that directly relates to the apoptotic chromatin changes, such as blebbing, fragmentation, and condensation, are quantified in Figure 4B. A simultaneous increase in the cells’ permeability was also observed (Figure 4C).
 | Figure 4 Effect of AM on nuclear intensity, MMP, permeability, and cytochrome c release. |
AM-induced MMP disruption and release of cytochrome c
In the phenomenon of apoptosis, MMP is often disturbed by the forming of permeability transition pores or the insertion of proapoptotic proteins such as Bax or Bid in the membrane of mitochondria. Hence, we studied the effect of AM on the changes of MMP in MDA-MB-231 cells with the use of a mitochondria-specific voltage-dependent dye. As displayed in Figure 4D, the MMP in the cells treated with AM (P<0.05) showed a significant reduction. In addition, a considerable decrease in the intensity of fluorescence (Figure 4A), which reflected the breakdown of MMP, AM also activated a major translocation of cytochrome c from the mitochondria into the cytosol. At 20 μg/mL, AM activated three times the amount of cytochrome c released in MDA-MB-231 cells compared to the control (P<0.05) (Figure 4E). Moreover, since the release of cytochrome c from the mitochondria triggers the activation of procaspase-9, we measured the immunoblotting of cytosol cytochrome c. AM significantly increased the level of cytosol cytochrome c in MDA-MB-231 cells compared to the control (P<0.05) (Figure 5D).
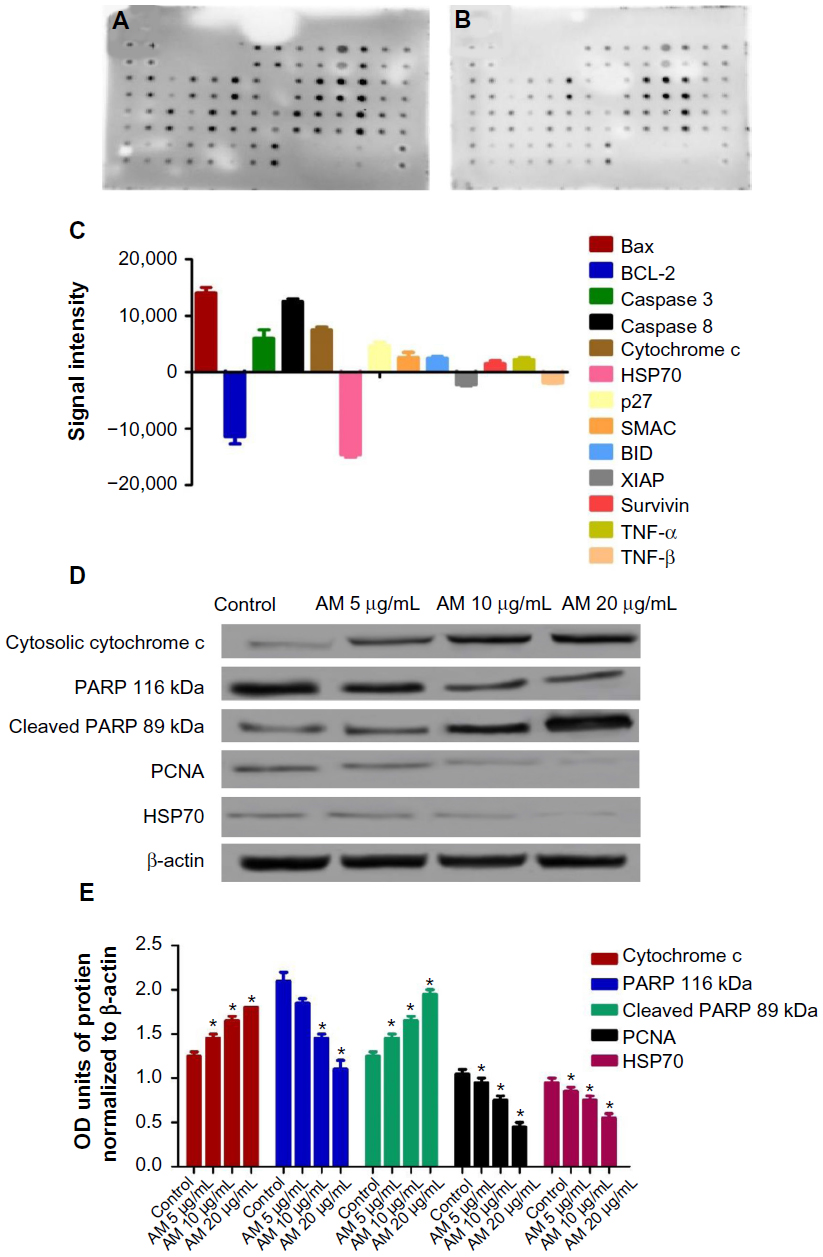 | Figure 5 Screening of the apoptotic markers and proteins expression induced by AM. |
AM regulates the expression of Bcl-2 and Bax protein
The expression of proapoptotic proteins, such as Bax, is an early incident that sensitizes cells to undergo apoptosis. Compared with the controls, MDA-MB-231 cells that were treated with AM displayed a considerable escalation in fluorescence intensity (Figure 6A) when stained with a specific Bax antibody. This outcome indicated that the expression of Bax is upregulated in MDA-MB-231 cells after dosing with 20 μg/mL AM. Nevertheless, the antiapoptotic, Bcl-2 levels were minimal during the period of treatment (Figure 6A). Figure 6B shows that both up- and downregulation of Bax and Bcl-2 were statistically significant (P<0.05) in a concentration-dependent manner. To confirm the immunofluorescence result, the expression of Bax and Bcl-2 in the MDA-MB-231 cells treated with AM were determined using Western blot analysis. AM treatment resulted in an upregulation of Bax and a downregulation of Bcl-2 in a concentration-dependent manner (Figure 6C), which resulted in an increase in the ratio of Bax to Bcl-2, therefore prompted apoptosis process (Figure 6D).
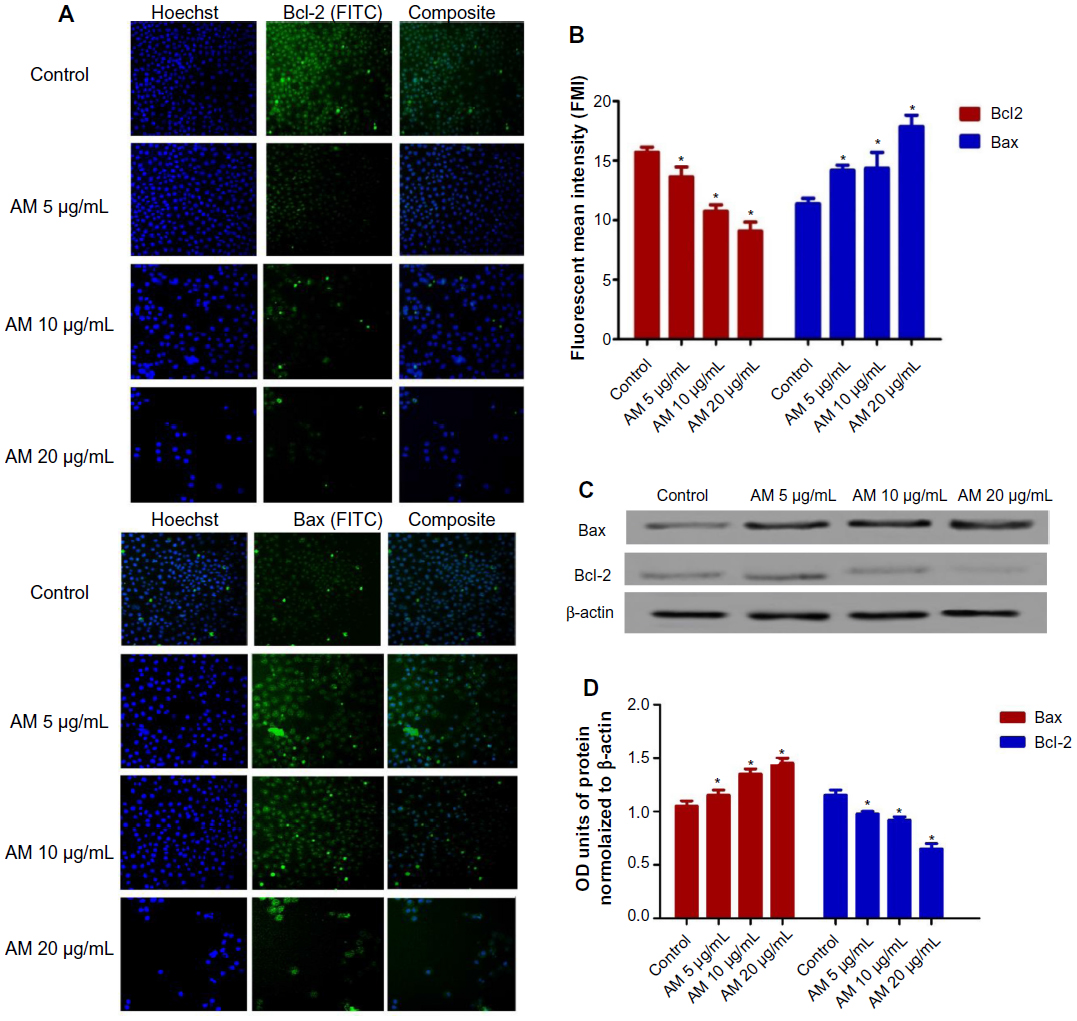 | Figure 6 AM changed the regulation of Bax/Bcl-2 expression. |
Effect of AM treatment on caspases-3/7, -8, and -9
All the caspase enzymes examined calorimetrically were shown to be induced by the treatment in a concentration-dependent manner. High levels of caspase-3/7, caspase-8, and caspase-9 were found in the greatest treatment concentration (20 μg/mL) with a significant variance from the control (P<0.05). Our results provide validation of the activation of all three caspases by AM in MDA-MB-231 cells (Figure 7A). To confirm the immunofluorescence result, the cell extracts were obtained after treatments and processed for Western blot analysis. The changes in protein levels in the MDA-MB-231 cells treated with different concentrations of AM were determined using Western blot analysis. In a concentration-dependent manner, AM treatment resulted in partial cleavage of procaspase-7, -9, and -8 (Figure 7B), which indicated that AM could increase the cleavage maturation of caspase-7, -9, and -8 (Figure 7C).
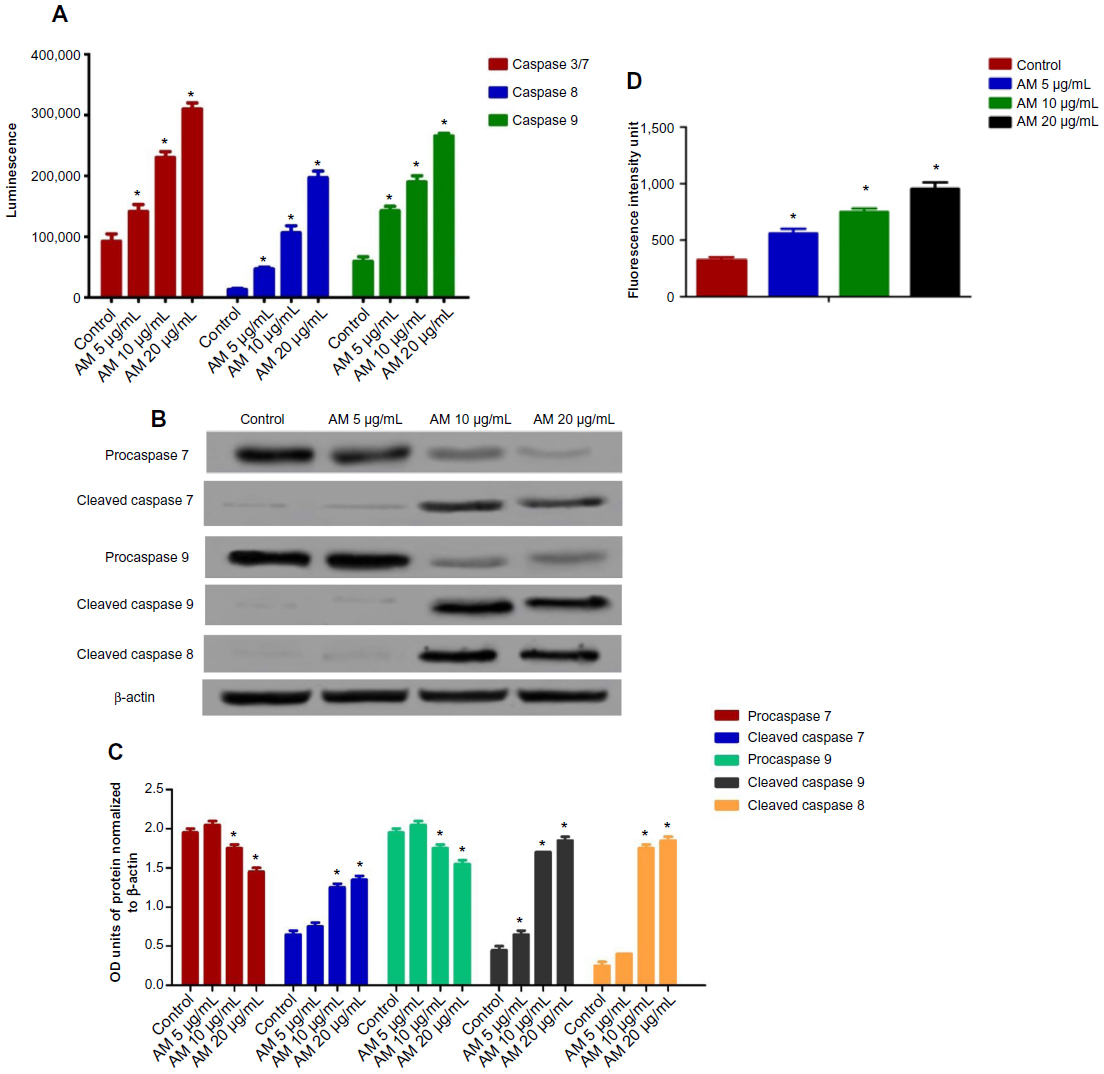 | Figure 7 Effects of AM on MDA-MB-231 cells’ procaspases, caspases, cleaved caspases, and ROS generation. |
Suppression of the PCNA and PARP protein expressions by AM
The expression of PCNA and PARP cleavage on MDA-MB-231 cells with or without AM treatment was tested by Western blot analysis. As shown in Figure 5D and E, after treatment with 5, 10, and 20 μg/mL of AM, the level of PCNA protein decreased significantly.
Since PARP cleavage results in cleaved product of 89 kDa, we observed a reduction in the PARP (116 kDa) and concomitant presence of cleaved product in a dose-dependent manner (Figure 5D and E).
AM-induced cell death includes increased ROS formation
The production of ROS was generally linked to the MMP disturbance and cell apoptosis. To determine this relation, we inspected the levels of ROS in MDA-MB-231 cells treated with AM. ROS was observed by the oxidation-sensitive fluorescent dye DCFH-DA. The escalation of dose-dependence in dichlorofluorescein diacetate fluorescence was observed in the treated cells. In addition, a considerable and immediate formation of ROS, up to twice as high as observed in the control, was identified following the dosing of cells with AM at 10 and 20 μg/mL (Figure 7D). This result indicated that this compound triggered the intracellular ROS development of MDA-MB-231 cells.
Cell cycle analysis
We conducted this trial to determine the influence of AM on the DNA content of MDA-MB-231 cells by the cell cycle phase distribution (G0, G1, S, G2, and M) after treatment (Figure 8). The results indicated that AM halted the cell cycle progression in the S phase (P<0.05). The results displayed in Figure 8E show that there is a significant S phase arrest in a concentration-dependent manner in the MDA-MB-231 cells, which accounts for 32.77%, 35.84%, and 49.33% of cells following treatment with 5, 10, and 20 μg/mL, respectively, for 24 hours (P<0.05). Meanwhile, the cells in both the G0/G1 and G2/M phases diminished with an increase in the treatment concentration.
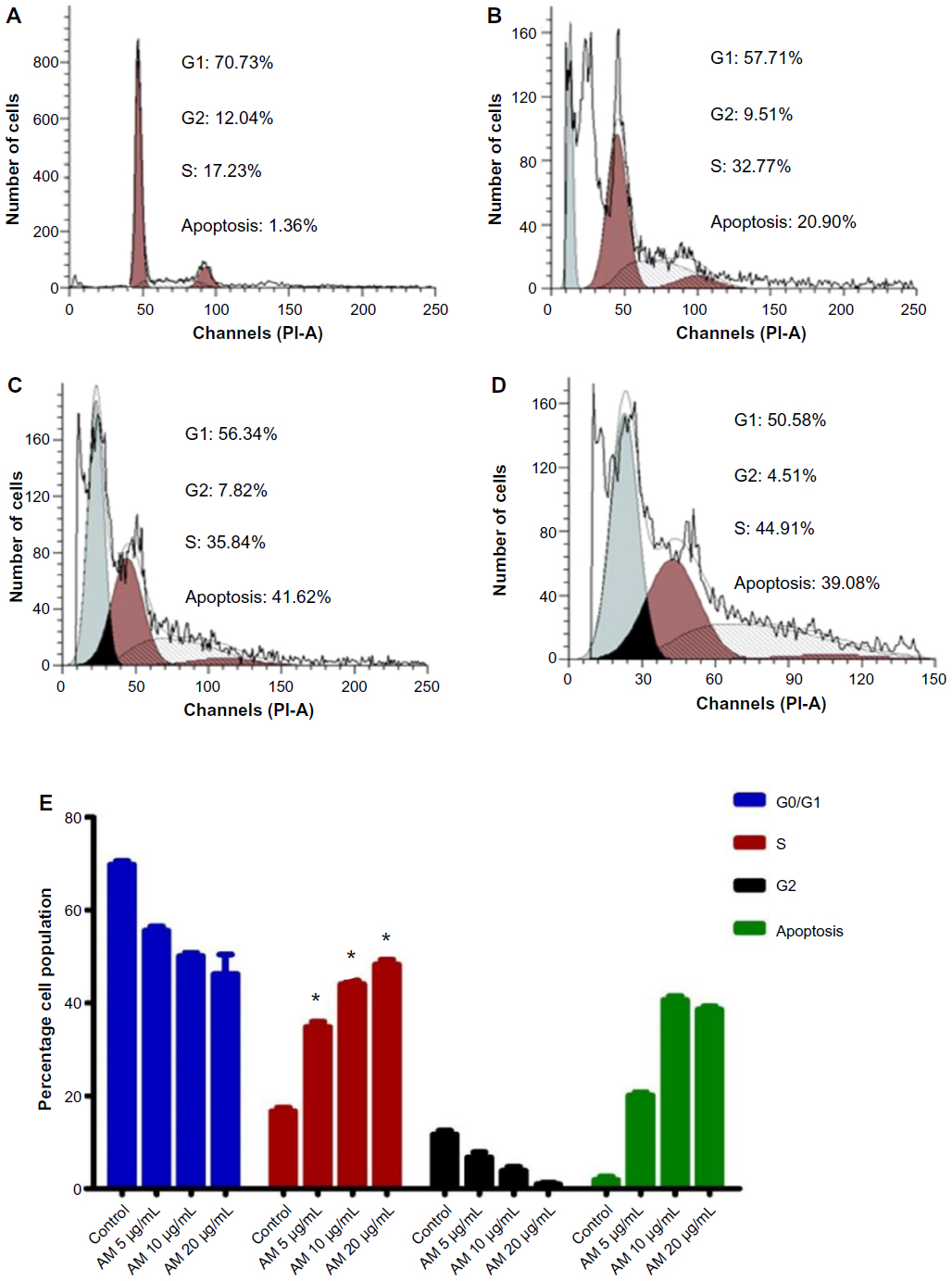 | Figure 8 AM induced S phase cell cycle arrest in MDA-MB-231 cells. |
Inhibition of TNF-α-induced NF-κB nuclear translocation by AM
The obstruction to apoptosis and cell proliferation was considered to be closely related to the activation of NF-κB. Hence, the role of AM in the suppression of activated NF-κB induced by the inflammatory cytokine, TNF-α, was examined using Alexa Fluor 488-conjugated anti-NF-κB antibody. Although a high NF-κB fluorescent intensity was observed in the cytoplasm in the control cells (medium alone) (Figure 9A), it was only faint in the nuclei, which suggests an absence of NF-κB activation in the control cells (medium alone). In addition, TNF-α alone considerably intensified the NF-κB fluorescent intensity in the nuclei. AM demonstrated a significant inhibitory effect on the triggering of NF-κB (Figure 9B). In the cells treated with curcumin, a known inhibitor of NF-κB activation, a significant suppression of TNF-α-induced NF-κB nuclear translocation was observed, as evidenced by the low nuclear NF-κB-related fluorescence intensity (Figure 9B). In parallel, the morphological alterations of NF-κB translocation, as shown by the immunofluorescence staining (Figure 9A), indicated the inhibitory effect of AM on TNF-α-induced NF-κB translocation in a concentration-dependent manner with significant inhibition for the 20 μg/mL concentration of AM. To affirm the immunofluorescence result, we measured the immunoblotting of the nuclear content of NF-κB p65. As shown in Figure 9C and D, treatment with AM significantly decreased the escalation of this nuclear protein induced by positive control (TNF-α) in a dose-dependent manner, particularly at 20 μg/mL of AM.
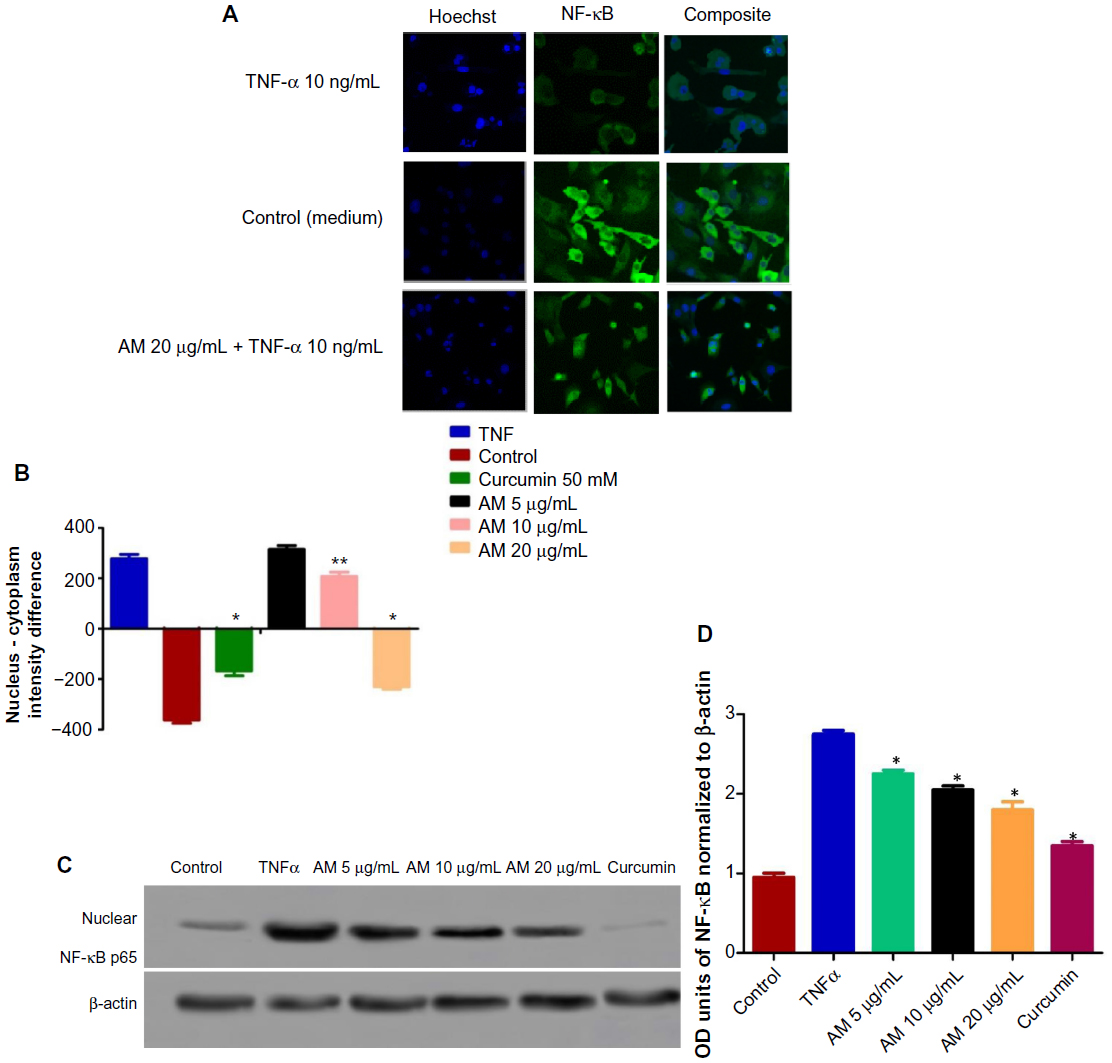 | Figure 9 Inhibition of TNF-α-induced NF-κB nuclear translocation by AM. |
Human apoptosis protein array
Following AM exposure for 24 hours, the MDA-MB-231 cells were lysed, and the apoptotic markers were screened using a protein array. In Figure 5A–C, the images show representative changes detected. All the major markers responsible for the apoptosis signaling pathway, including Bax, Bcl-2, Bim, cytochrome c, and caspase-3/7 and -8, were expressed in the two models. A significant chaperone Hsp70, which played a part in the apoptosis, was also downregulated, while the cell proliferation repressor protein p27, as well as X-linked IAP (XIAP), were also induced in this in vitro model. To confirm the protein array result, the expression of Hsp70 in the MDA-MB-231 cells treated with AM were determined using Western blot analysis. AM treatment resulted in a downregulation of this protein in a concentration-dependent manner (Figure 5D and E).
Discussion
The role of apoptosis is crucial in many functions, ranging from fetal development to adult tissue homeostasis.32 Tumors are attributed to uncontrolled proliferation as well as reduced apoptosis. One critical method by which cytotoxic drugs destroy cancer cells is the activation of apoptotic pathways.33 Herbal medicines have been a major source from which numerous apoptosis-inducing agents are derived,34 and, according to several reports, many of these naturally occurring compounds may contribute partially in human cancer prevention or therapy.35,36 Such studies showed that bioactive compounds elicit apoptosis in cancer cells.37,38 Renewed interest in the application of oriental medicine for cancer treatment, along with auspicious clinical results, has led to much emphasis being given to medicinal plants. Nevertheless, the chemical components as well as definite mechanisms of many herbal medicines remain obscure.
In this regard, C. arborescens is one well-known plant used in Asian countries for the prevention and treatment of different kinds of ailments.14 AM, as a natural compound, is a major prenylated xanthone isolated from this plant. Therefore, the present study elucidated the mechanism of apoptosis provoked by AM towards MDA-MB-231 cells.
MTT assay was the antiproliferative assay employed in this study. Based on the data gathered from this experiment, the IC50 of AM on MDA-MB-231 cell lines was shown to be 9 μg/mL. The IC50 value of AM falls within the significant range of cytotoxic effects stipulated by the National Cancer Institute Standard, United States.39 However, AM treatment did not exhibit any cytotoxicity toward normal MCF-10A breast cells, exhibiting no IC50 value until 30 μg/mL. According to Shier,40 compounds that demonstrate an IC50 value of more than 30 μg/mL are not considered potentially cytotoxic, while compounds with an IC50 value of less than 5.0 μg/mL are considered very active. These findings showed that AM has different levels of cytotoxicity on normal and cancer cells that are cytotoxic toward mammary gland cancer cells, unlike its effect toward normal cells. Therefore, AM could act as a potential antitumor compound that is worth studying, especially pertaining to its cell death mechanism.
The use of AO and PI fluorescent dyes in the analysis of cell culture viability and morphological characteristics is common. Hence, we used these fluorescent dyes to observe the different stages of apoptosis, beginning from the condensation of the chromatin up to the formation of apoptotic bodies, with AM treatment. Although the morphological features were clearly noticed, the assay of annexin V was then conducted in an attempt to quantify the cells of the apoptotic population. Annexin V was found to link specifically to phosphatidylserine, which is located at the outer membrane leaflet of cells in the presence of calcium.41,42 The present study established that AM treatment can induce cell death in MDA-MB-231 cells through apoptosis. In addition, the results showed a significant dose-dependent increase occurring in the early stage of apoptosis.
Although both extrinsic and intrinsic pathways are involved in apoptosis, the sensitivity of the intrinsic pathways causes tumors to occur more frequently through this route.39 Mitochondria are the main cellular component for the intrinsic means of apoptosis due to their ability to directly initiate the apoptotic cellular program. The primary involvement of the mitochondria is in the cell’s redox status, although they can also execute multiple cellular functions, including energy production, cell proliferation, and death.43,44 The permeabilization of the outer membrane and alteration of the mitochondrial transmembrane potential (Δψm) by the mitochondria initiating the apoptotic cascade, releases soluble proapoptotic proteins including cytochrome c, which, ultimately, leads to the activation of caspases-9 and -3.39 It is essential that the MMP is decreased for this process to begin. The ability of AM to reduce the MMP has been revealed by the fluorescence-based HCS analysis. Concurrently, the release of cytochrome c also increased. Our results regarding AM correlate well with previous studies on AM against human chondrosarcoma cells (SW1353) and human hepatoma cells (SK-Hep-1), showing the disruption of MMP and the release of cytochrome c from the mitochondria to the cytosol.45,46
Subsequently, with MMP disruption and the release of cytochrome c, AM treatment of MDA-MB-231 cells triggered the activation of caspases-3/7, -8, and -9. In fact, the stimulation of caspases-3/7, -8, and -9 were shown to be induced by AM in a concentration-dependent manner. Caspase-9 is located in the intermembrane space of the mitochondria. It is discharged in a Bcl-2-inhibitable manner following the induction of permeability transition, whereas, in cells, the release is upon the induction of apoptosis.47 The caspase-9 that is released then activates the post-mitochondrial caspases, which includes caspases-3 and -7; the disassembly of the cell takes place in what is termed the execution phase of apoptosis.48 Caspase-8 is closely implicated in apoptosis signaling via the extrinsic pathway, even though it is found in the upstream and downstream of the mitochondria.47,49 Moreover, the engagement of caspase-8 in mitochondrial pathways through the cleavage of the Bcl-2 family member Bid to tBid has been shown on numerous occasions.50,51 Our results regarding AM are well correlated with previous studies on AM against BJMC3879 metastatic murine mammary adenocarcinoma cells and MDA-MB-231 an estrogen receptor-negative and HER-2/neu-negative human breast cancer cells that showed the activation of caspases-3, -9, and -8.52,53
Oxidative stress is considered to be a vital state in promoting programmed cell death in reaction to a variety of signals and pathophysiological situations. Moreover, studies have verified that ROS is an inevitable factor in cell death in which mitochondria are significantly involved.54,55 To determine this relationship, we measured the ROS level upon AM treatment of MDA-MB-231 cells. The results clearly underline the significant relation (P<0.05). There was elevated (twofold) intracellular ROS with AM treatment (20 μg/mL) of MDA-MB-231 cells and this could be due to the free radical generation during the cytotoxicity.
Concurrently, a number of Bcl-2 family members, including Bcl-2 and Bax, are known to have the ability to control apoptosis.56 In ascertaining the vulnerability of cells toward death, it is crucial for a balance to exist between antiapoptotic and proapoptotic Bcl-2 family members.57 It has been shown in previous reports that susceptibility to apoptosis can be caused by the upregulation of Bax and downregulation of Bcl-2.58,59 Thus, we examined the effect of AM on the expression of proteins that belong to this family. Bcl-2 is a cytoplasmic protein that plays a significant part in inhibiting apoptosis that is caused by factors, such as chemotherapeutic agents and irradiation, as well as the withdrawal of growth factors.60,61 The immunofluorescence and immunoblotting results in this study demonstrated the Bcl-2 downregulation in MDA-MB-231 cells after treatment with the different concentrations of AM. This occurrence likely explains the apoptotic effects of AM on MDA-MB-231 cells. The exertion of these effects on the expression of Bcl-2 may also be connected to the production of mitochondrial apoptotic factors, which, eventually, lead to apoptosis.62 Our findings regarding AM support previous findings, which showed that the induction of apoptosis seems to be modulated by the levels of Bcl-2 and Bax in the head and neck squamous carcinoma cell line (HNSCC), human Caucasian colon adenocarcinoma cell line (COLO 205), undifferentiated human colon carcinoma cell line (MIP-101), and human colon epithelial adenocarcinoma cell line (SW 620).21,63
PARP can detect DNA damage due to its sensitivity. This enzyme is strongly triggered by DNA strand breaks and has primary functions in processes, such as DNA repair and the maintenance of genome stability. In addition, it also performs a crucial role in the induction of cell death initiated by a variety of stimuli.64 The proteolytic degradation of PARP that takes place at the beginning of apoptosis is caused by the activated caspase-3.65 As demonstrated by the immunoblot analysis, AM treatment significantly increased thee cleaved PARP protein. Based on the results, it was shown that the stimulation of caspases-9 and -3 is induced by the released cytochrome c, which in turn cleaves 116 kDa PARP into 85 kDa fragments in MDA-MB-231 treated cells.
The growth of cells is moderated by cell proliferation (division) and apoptosis (programmed cell death),66 and homeostasis is sustained by a balance between the two. One key characteristic of cancer cells is the absence of the homeostasis process, therefore dietary management of this process may result in a deeper understanding regarding cancer prevention.67 Resveratrol, curcumin, and epigallocatechin are among the many natural compounds that have displayed antiproliferative actions against cancer cells through the initiation of apoptosis. Therefore, a highly conserved 36 kDa nuclear protein of DNA polymerase-delta known as the PCNA has been identified as a useful marker that can be used to assess tumor cell proliferation and progression.68 An overexpression of PCNA has been reported in different malignancies, including breast cancer.69 The antiproliferative efficacy of AM has been established in that it was found to decrease the immunoblotting expression of this proliferative marker in MDA-MB-231 cells. Our results corroborate previous reports on the antiproliferative activities of AM toward canine osteosarcoma cells and human lung adenocarcinoma A549 cells.70,71
The mode of apoptosis caused by numerous natural compounds is closely related to cell cycle arrest.72–74 It has been established that cell cycle control is a significant event in safeguarding precise cellular division. Many carcinogenic processes are reported to cause the abnormalities of cell cycle regulators. For this reason, it is reasonable to target and alter the cell cycle regulators in cancer cells for the purpose of chemoprevention and treatment.75–77 Following dosage with various concentrations of AM, the examination of the cell cycle of MDA-MB-231 cells revealed a higher number of cells in the S phase. On the other hand, the amount of cells in the G0/G1 and G2/M phase reduced in comparison with the untreated cells (Figure 8E). These results indicate the ability of AM in inhibiting cellular proliferation via S phase arrest.
The NF-κB, is a protein complex that plays a role in regulating DNA transcription. In addition, it has also been considered to be an apoptosis inhibitor.78,79 Thus, repression of the NF-κB activity can induce apoptosis. In this study, we have demonstrated that AM can prevent the TNF-α-induced NF-κB translocation of the cytoplasm to the nucleus of the MDA-MB-231 cells using immunofluorescence and immunoblotting techniques, suggesting the participation of an NF-κB inhibition mechanism in apoptosis, which indicated that AM could become a useful antitumor agent.
Apoptosis involves a large number of proteins.80,81 To determine the role of the central apoptosis-related proteins, a protein array analysis was conducted. In this study, various proteins in the extrinsic and intrinsic pathways were examined. These proteins include those that are known to cause apoptosis, including Bax, caspase-3, cytochrome c, caspase-8, and SMAC, as well as those that have been recognized as antiapoptotic, including Bcl-2 and XIAP, of which XIAP is a member of the inhibitors of apoptosis (IAP) family of proteins,82 and SMAC is a proapoptotic protein that acts together with IAP to reduce their inhibitory effects.83,84 Together with the Bcl-2 family members, heat shock proteins have also been deemed to be apoptosis inhibitors, due to their significant role in cell survival, through stopping the release of cytochrome c from mitochondria or by preventing the formation of apoptosome.85 The protein array analysis and Western blot results showed a significant reduction in Hsp70 following treatment with AM. This finding is consistent with the findings from earlier studies, which indicated that apoptosis could be suppressed by an overexpression of Hsp70.86,87 The results from the protein array displayed a characteristic profile of protein levels related to both the extrinsic and intrinsic apoptosis pathways in MDA-MB-231 cells treated with AM.
According to the observations mentioned in this report, it can be concluded that AM has the ability to induce apoptosis in MDA-MB-231 cells with cell death-transducing signals that regulate the MMP through a downregulation of Bcl-2 and an upregulation of Bax, which, in turn, triggers the release of the cytochrome c from mitochondria to cytosol. The activation of caspase-9 is triggered once cytochrome c enters the cytosol, followed by the activation of the downstream executioner caspase-3/7. Subsequently, it cleaves specific substrates, making way for the occurrence of apoptotic changes. Increase of caspase-8 revealed the involvement of the extrinsic pathway. This form of apoptosis was suggested to occur through both the extrinsic and intrinsic apoptosis pathways with involvement of the NF-κB and HSP70 signaling pathways.
Acknowledgments
This study was financially supported by the University of Malaya through the Postgraduate Research Fund (PPP) grant PG 141-2012B, University Malaya Research Grant RP001C-13BIO, and High Impact Research Grant UM-MOHE M.C/625/1/HIR/MOHE//SC/09 from the Ministry of Higher Education Malaysia.
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63(1):11–30. | |
Hortobagyi GN, de la Garza Salazar J, Pritchard K, et al; ABREAST Investigators. The global breast cancer burden: variations in epidemiology and survival. Clin Breast Cancer. 2005;6(5):391–401. | |
Bray F, Ren JS, Masuyer E, Ferlay J. Global estimates of cancer prevalence for 27 sites in the adult population in 2008. Int J Cancer. 2013;132(5):1133–1145. | |
Schottenfeld D, Beebe-Dimmer JL, Buffler PA, Omenn GS. Current perspective on the global and United States cancer burden attributable to lifestyle and environmental risk factors. Annu Rev Public Health. 2013;34:97–117. | |
Varghese JS, Smith PL, Folkerd E, et al. The heritability of mammographic breast density and circulating sex-hormone levels: two independent breast cancer risk factors. Cancer Epidemiol Biomarkers Prev. 2012;21(12):2167–2175. | |
Charalambous C, Pitta CA, Constantinou AI. Equol enhances tamoxifen’s anti-tumor activity by induction of caspase-mediated apoptosis in MCF-7 breast cancer cells. BMC Cancer. 2013;13(1):238. | |
Yu X, Filardo EJ, Shaikh ZA. The membrane estrogen receptor GPR30 mediates cadmium-induced proliferation of breast cancer cells. Toxicol Appl Pharmacol. 2010;245(1):83–90. | |
Kuo JR, Wang CC, Huang SK, Wang SJ. Tamoxifen depresses glutamate release through inhibition of voltage-dependent Ca2+ entry and protein kinase Cα in rat cerebral cortex nerve terminals. Neurochem Int. 2012;60(2):105–114. | |
Pawar P, Ma L, Byon CH, et al. Molecular mechanisms of tamoxifen therapy for cholangiocarcinoma: role of calmodulin. Clin Cancer Res. 2009;15(4):1288–1296. | |
Abbasalipourkabir R, Salehzadeh A, Abdullah R. Antitumor activity of tamoxifen loaded solid lipid nanoparticles on induced mammary tumor gland in Sprague-Dawley rats. African Journal of Biotechnology. 2010;9(43):7337–7345. | |
Shukla Y, George J. Combinatorial strategies employing nutraceuticals for cancer development. Ann N Y Acad Sci. 2011;1229(1):162–175. | |
Cragg GM, Newman DJ. Natural products: a continuing source of novel drug leads. Biochim Biophys Acta. 2013;1830(6):3670–3695. | |
Tang J, Feng Y, Tsao S, Wang N, Curtain R, Wang Y. Berberine and Coptidis rhizoma as novel antineoplastic agents: a review of traditional use and biomedical investigations. J Ethnopharmacol. 2009;126(1):5–17. | |
José M, Kijjoa A, Gonzalez TG, et al. Xanthones from Cratoxylum maingayi. Phytochemistry. 1998;49(7):2159–2162. | |
Srithi K, Balslev H, Wangpakapattanawong P, Srisanga P, Trisonthi C. Medicinal plant knowledge and its erosion among the Mien (Yao) in northern Thailand. J Ethnopharmacol. 2009;123(2):335–342. | |
Obolskiy D, Pischel I, Siriwatanametanon N, Heinrich M. Garcinia mangostana L.: a phytochemical and pharmacological review. Phytother Res. 2009;23(8):1047–1065. | |
Pedraza-Chaverri J, Cárdenas-Rodríguez N, Orozco-Ibarra M, Pérez-Rojas JM. Medicinal properties of mangosteen (Garcinia mangostana). Food Chem Toxicol. 2008;46(10):3227–3239. | |
Pérez-Rojas JM, Cruz C, García-López P, et al. Renoprotection by alpha-Mangostin is related to the attenuation in renal oxidative/nitrosative stress induced by cisplatin nephrotoxicity. Free Radic Res. 2009;43(11):1122–1132. | |
Saleem M. Lupeol, a novel anti-inflammatory and anti-cancer dietary triterpene. Cancer Lett. 2009;285(2):109–115. | |
Sampath PD, Kannan V. Mitigation of mitochondrial dysfunction and regulation of eNOS expression during experimental myocardial necrosis by alpha-mangostin, a xanthonic derivative from Garcinia mangostana. Drug Chem Toxicol. 2009;32(4):344–352. | |
Kaomongkolgit R, Chaisomboon N, Pavasant P. Apoptotic effect of alpha-mangostin on head and neck squamous carcinoma cells. Arch Oral Biol. 2011;56(5):483–490. | |
Wang JJ, Sanderson BJ, Zhang W. Significant anti-invasive activities of α-mangostin from the mangosteen pericarp on two human skin cancer cell lines. Anticancer Res. 2012;32(9):3805–3816. | |
Nelli GB, K AS, Kilari EK. Antidiabetic effect of α-mangostin and its protective role in sexual dysfunction of streptozotocin induced diabetic male rats. Syst Biol Reprod Med. 2013;59(6):319–328. | |
Koh JJ, Qiu S, Zou H, et al. Rapid bactericidal action of alpha-mangostin against MRSA as an outcome of membrane targeting. Biochim Biophys Acta. 2013;1828(2):834–844. | |
Kaomongkolgit R, Jamdee K, Chaisomboon N. Antifungal activity of alpha-mangostin against Candida albicans. J Oral Sci. 2009;51(3):401–406. | |
Ngawhirunpat T, Opanasopi P, Sukma M, Sittisombut C, Kat A, Adachi I. Antioxidant, free radical-scavenging activity and cytotoxicity of different solvent extracts and their phenolic constituents from the fruit hull of mangosteen (Garcinia mangostana). Pharm Biol. 2010;48(1):55–62. | |
Kumar RB, Shanmugapriya B, Thiyagesan K, Kumar SR, Xavier SM. A search for mosquito larvicidal compounds by blocking the sterol carrying protein, AeSCP-2, through computational screening and docking strategies. Pharmacognosy Res. 2010;2(4):247–253. | |
Devalaraja S, Jain S, Yadav H. Exotic fruits as therapeutic complements for diabetes, obesity and metabolic syndrome. Food Res Int. 2011;44(7):1856–1865. | |
Acharyya S, Oskarsson T, Vanharanta S, et al. A CXCL1 paracrine network links cancer chemoresistance and metastasis. Cell. 2012; 150(1):165–178. | |
Otterbach F, Callies R, Adamzik M, et al. Aquaporin 1 (AQP1) expression is a novel characteristic feature of a particularly aggressive subgroup of basal-like breast carcinomas. Breast Cancer Res Treat. 2010;120(1):67–76. | |
Looi CY, Arya A, Cheah FK, et al. Induction of apoptosis in human breast cancer cells via caspase pathway by vernodalin isolated from Centratherum anthelminticum (L.) seeds. PLoS One. 2013;8(2):e56643. | |
Yang HS, Kim JY, Lee JH, et al. Celastrol isolated from Tripterygium regelii induces apoptosis through both caspase-dependent and -independent pathways in human breast cancer cells. Food Chem Toxicol. 2011;49(2):527–532. | |
Zhang X, Wei H, Liu Z, et al. A novel protoapigenone analog RY10-4 induces breast cancer MCF-7 cell death through autophagy via the Akt/mTOR pathway. Toxicol Appl Pharmacol. 2013;270(2):122–128. | |
Pacifico S, Gallicchio M, Lorenz P, et al. Apolar Laurus nobilis leaf extracts induce cytotoxicity and apoptosis towards three nervous system cell lines. Food Chem Toxicol. 2013;62(1):628–637. | |
Tanaka T. Role of apoptosis in the chemoprevention of cancer. J Exp Clin Med. 2013;5(3):89–91. | |
Kumazaki M, Noguchi S, Yasui Y, et al. Anti-cancer effects of naturally occurring compounds through modulation of signal transduction and miRNA expression in human colon cancer cells. J Nutr Biochem. 2013;24(11):1849–1858. | |
Pan MH, Lai CS, Wang H, Lo CY, Ho CT, Li S. Black tea in chemo-prevention of cancer and other human diseases. Food Science and Human Wellness. 2013;2(1):12–21. | |
Plastina P, Bonofiglio D, Vizza D, et al. Identification of bioactive constituents of Ziziphus jujube fruit extracts exerting antiproliferative and apoptotic effects in human breast cancer cells. J Ethnopharmacol. 2012;140(2):325–332. | |
Constant Anatole P, Guru SK, Bathelemy N, et al. Ethyl acetate fraction of Garcina epunctata induces apoptosis in human promyelocytic cells (HL-60) through the ROS generation and G0/G1 cell cycle arrest: a bioassay-guided approach. Environ Toxicol Pharmacol. 2013;36(3):865–874. | |
Shier WT. Mammalian cell culture on $5 a day: a laboratory manual of low cost methods. Los Banos, University of the Philippines. 1991;64. | |
Cheng X, Xiao Y, Wang X, et al. Anti-tumor and pro-apoptotic activity of ethanolic extract and its various fractions from Polytrichum commune L.ex Hedw in L1210 cells. J Ethnopharmacol. 2012;143(1):49–56. | |
van Engeland M, Nieland LJ, Ramaekers FC, Schutte B, Reutelingsperger CP. Annexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry. 1998;31(1):1–9. | |
Olguín-Martínez M, Hernández-Espinosa DR, Hernández-Muñoz R. α-Tocopherol administration blocks adaptive changes in cell NADH/NAD+ redox state and mitochondrial function leading to inhibition of gastric mucosa cell proliferation in rats. Free Radic Biol Med. 2013;65:1090–1100. | |
Giorgi C, Romagnoli A, Pinton P, Rizzuto R. Ca2+ signaling, mitochondria and cell death. Curr Mol Med. 2008;8(2):119–130. | |
Hsieh SC, Huang MH, Cheng CW, Hung JH, Yang SF, Hsieh YH. α-Mangostin induces mitochondrial dependent apoptosis in human hepatoma SK-Hep-1 cells through inhibition of p38 MAPK pathway. Apoptosis. 2013;18(12):1548–1560. | |
Krajarng A, Nakamura Y, Suksamrarn S, Watanapokasin R. α-Mangostin induces apoptosis in human chondrosarcoma cells through downregulation of ERK/JNK and Akt signaling pathway. J Agric Food Chem. 2011;59(10):5746–5754. | |
Mohan S, Abdelwahab SI, Kamalidehghan B, et al. Involvement of NF-κB and Bcl2/Bax signaling pathways in the apoptosis of MCF7 cells induced by a xanthone compound Pyranocycloartobiloxanthone A. Phytomedicine. 2012;19(11):1007–1015. | |
Li Z, Jo J, Jia JM, et al. Caspase-3 activation via mitochondria is required for long-term depression and AMPA receptor internalization. Cell. 2010;141(5):859–871. | |
Jin Z, Li Y, Pitti R, et al. Cullin3-based polyubiquitination and p62-dependent aggregation of caspase-8 mediate extrinsic apoptosis signaling. Cell. 2009;137(4):721–735. | |
Gu Q, Wang JD, Xia HH, et al. Activation of the caspase-8/Bid and Bax pathways in aspirin-induced apoptosis in gastric cancer. Carcinogenesis. 2005;26(3):541–546. | |
Turk B, Stoka V. Protease signalling in cell death: caspases versus cysteine cathepsins. FEBS Lett. 2007;581(15):2761–2767. | |
Shibata MA, Iinuma M, Morimoto J, et al. α-Mangostin extracted from the pericarp of the mangosteen (Garcinia mangostana Linn) reduces tumor growth and lymph node metastasis in an immunocompetent xenograft model of metastatic mammary cancer carrying a p53 mutation. BMC Med. 2011;9:69. | |
Kurose H, Shibata MA, Iinuma M, Otsuki Y. Alterations in cell cycle and induction of apoptotic cell death in breast cancer cells treated with α-mangostin extracted from mangosteen pericarp. J Biomed Biotechnol. 2012;2012:672428. | |
Ham YM, Yoon WJ, Park SY, et al. Quercitrin protects against oxidative stress-induced injury in lung fibroblast cells via up-regulation of Bcl-xL. J Funct Foods. 2012;4(1):253–262. | |
Cui Y, Lu Z, Bai L, Shi Z, Zhao W-e, Zhao B. β-Carotene induces apoptosis and up-regulates peroxisome proliferator-activated receptor γ expression and reactive oxygen species production in MCF-7 cancer cells. Eur J Cancer. 2007;43(17):2590–2601. | |
Sánchez-Pérez Y, Morales-Bárcenas R, García-Cuellar CM, et al. The α-mangostin prevention on cisplatin-induced apoptotic death in LLC-PK1 cells is associated to an inhibition of ROS production and p53 induction. Chem Biol Interact. 2010;188(1):144–150. | |
Adams JM, Cory S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene. 2007;26(9):1324–1337. | |
Lee JS, Jung WK, Jeong MH, Yoon TR, Kim HK. Sanguinarine induces apoptosis of HT-29 human colon cancer cells via the regulation of Bax/Bcl-2 ratio and caspase-9-dependent pathway. Int J Toxicol. 2012;31(1):70–77. | |
Zhu AK, Zhou H, Xia JZ, et al. Ziyuglycoside II-induced apoptosis in human gastric carcinoma BGC-823 cells by regulating Bax/Bcl-2 expression and activating caspase-3 pathway. Braz J Med Biol Res. 2013;46(3):670–675. | |
Ferenc P, Solár P, Kleban J, Mikes J, Fedorocko P. Down-regulation of Bcl-2 and Akt induced by combination of photoactivated hypericin and genistein in human breast cancer cells. J Photochem Photobiol B. 2010;98(1):25–34. | |
Zhang GJ, Kimijima I, Onda M, et al. Tamoxifen-induced apoptosis in breast cancer cells relates to down-regulation of bcl-2, but not bax and bcl-X(L), without alteration of p53 protein levels. Clin Cancer Res. 1999;5(10):2971–2977. | |
Jin S, Zhang QY, Kang XM, Wang JX, Zhao WH. Daidzein induces MCF-7 breast cancer cell apoptosis via the mitochondrial pathway. Ann Oncol. 2010;21(2):263–268. | |
Watanapokasin R, Jarinthanan F, Nakamura Y, Sawasjirakij N, Jaratrungtawee A, Suksamrarn S. Effects of α-mangostin on apoptosis induction of human colon cancer. World J Gastroenterol. 2011;17(16):2086–2095. | |
Bouchard VJ, Rouleau M, Poirier GG. PARP-1, a determinant of cell survival in response to DNA damage. Exp Hematol. 2003;31(6):446–454. | |
Yoon JS, Kasin Yadunandam A, Kim SJ, Woo HC, Kim HR, Kim GD. Dieckol, isolated from Ecklonia stolonifera, induces apoptosis in human hepatocellular carcinoma Hep3B cells. J Nat Med. 2013;67(3):519–527. | |
Wen S, Niu Y, Lee SO, Chang C. Androgen receptor (AR) positive vs negative roles in prostate cancer cell deaths including apoptosis, anoikis, entosis, necrosis and autophagic cell death. Cancer Treat Rev. 2014;40(1):31–40. | |
Lançon A, Michaille JJ, Latruffe N. Effects of dietary phytophenols on the expression of microRNAs involved in mammalian cell homeostasis. J Sci Food Agric. 2013;93(13):3155–3164. | |
Campisi G, Di Fede O, Giovannelli L, et al. Use of fuzzy neural networks in modeling relationships of HPV infection with apoptotic and proliferation markers in potentially malignant oral lesions. Oral Oncol. 2005;41(10):994–1004. | |
Jia Z, Zhao W, Fan L, Sheng W. The expression of PCNA, c-erbB-2, p53, ER and PR as well as atypical hyperplasia in tissues nearby the breast cancer. J Mol Histol. 2012;43(1):115–120. | |
Krajarng A, Nilwarankoon S, Suksamrarn S, Watanapokasin R. Antiproliferative effect of α-mangostin on canine osteosarcoma cells. Res Vet Sci. 2012;93(2):788–794. | |
Shih YW, Chien ST, Chen PS, Lee JH, Wu SH, Yin LT. Alpha-mangostin suppresses phorbol 12-myristate 13-acetate-induced MMP-2/MMP-9 expressions via alphavbeta3 integrin/FAK/ERK and NF-kappaB signaling pathway in human lung adenocarcinoma A549 cells. Cell Biochem Biophys. 2010;58(1):31–44. | |
Mohan S, Abdul AB, Abdelwahab SI, et al. Typhonium flagelliforme induces apoptosis in CEMss cells via activation of caspase-9, PARP cleavage and cytochrome c release: its activation coupled with G0/G1 phase cell cycle arrest. J Ethnopharmacol. 2010;131(3):592–600. | |
Park HS, Park KI, Lee DH, et al. Polyphenolic extract isolated from Korean Lonicera japonica Thunb induce G2/M cell cycle arrest and apoptosis in HepG2 cells: involvements of PI3K/Akt and MAPKs. Food Chem Toxicol. 2012;50(7):2407–2416. | |
Yang F, Sun X, Shen J, et al. A recombined protein (rSj16) derived from Schistosoma japonicum induces cell cycle arrest and apoptosis of murine myeloid leukemia cells. Parasitol Res. 2013;112(3):1261–1272. | |
Khan MA, Chen HC, Wan XX, et al. Regulatory effects of resveratrol on antioxidant enzymes: a mechanism of growth inhibition and apoptosis induction in cancer cells. Mol Cells. 2013;35(3):219–225. | |
Pozo-Guisado E, Alvarez-Barrientos A, Mulero-Navarro S, Santiago-Josefat B, Fernandez-Salguero PM. The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast cancer cells: cell-specific alteration of the cell cycle. Biochem Pharmacol. 2002;64(9):1375–1386. | |
Xiao D, Herman-Antosiewicz A, Antosiewicz J, et al. Diallyl trisulfide-induced G(2)-M phase cell cycle arrest in human prostate cancer cells is caused by reactive oxygen species-dependent destruction and hyperphosphorylation of Cdc 25 C. Oncogene. 2005;24(41):6256–6268. | |
Naugler WE, Karin M. NF-κB and cancer-identifying targets and mechanisms. Curr Opin Genet Dev. 2008;18(1):19–26. | |
Verfaillie T, Garg AD, Agostinis P. Targeting ER stress induced apoptosis and inflammation in cancer. Cancer Lett. 2013;332(2):249–264. | |
Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495–516. | |
Fan TJ, Han LH, Cong RS, Liang J. Caspase family proteases and apoptosis. Acta Biochim Biophys Sin (Shanghai). 2005;37(11):719–727. | |
Deveraux QL, Reed JC. IAP family proteins – suppressors of apoptosis. Genes Dev. 1999;13(3):239–252. | |
Chai J, Du C, Wu JW, Kyin S, Wang X, Shi Y. Structural and biochemical basis of apoptotic activation by Smac/DIABLO. Nature. 2000;406(6798):855–862. | |
Srinivasula SM, Hegde R, Saleh A, et al. A conserved XIAP-interaction motif in caspase-9 and Smac/DIABLO regulates caspase activity and apoptosis. Nature. 2001;410(6824):112–116. | |
Mosser DD, Caron AW, Bourget L, et al. The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol Cell Biol. 2000;20(19):7146–7159. | |
Isa NM, Abdul AB, Abdelwahab SI, et al. Boesenbergin A, a chalcone from Boesenbergia rotunda induces apoptosis via mitochondrial dysregulation and cytochrome c release in A549 cells in vitro: involvement of HSP70 and Bcl2/Bax signalling pathways. J Funct Foods. 2013;5(1):87–97. | |
Yang W, Ahmed M, Tasawwar B, et al. Combination radiofrequency (RF) ablation and IV liposomal heat shock protein suppression: reduced tumor growth and increased animal endpoint survival in a small animal tumor model. J Control Release. 2012;160(2):239–244. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.