Back to Journals » Journal of Inflammation Research » Volume 15
Investigation on Probable Association Between IL-13, IL-13RA1, and IL-13RA2 Genes Polymorphism and Pulmonary Tuberculosis
Authors Li HM, Tang F, Huang Q, Pan HF ![]() , Zhang TP
, Zhang TP
Received 14 May 2022
Accepted for publication 30 July 2022
Published 8 August 2022 Volume 2022:15 Pages 4527—4536
DOI https://doi.org/10.2147/JIR.S374714
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Ning Quan
Hong-Miao Li,1 Fei Tang,2 Qian Huang,3 Hai-Feng Pan,1 Tian-Ping Zhang4
1Department of Epidemiology and Biostatistics, School of Public Health, Anhui Medical University, Hefei, People’s Republic of China; 2Department of Interventional Pulmonology and Endoscopic Diagnosis and Treatment Center, Anhui Chest Hospital, Hefei, People’s Republic of China; 3Department of Public Health, Medical Department, Qinghai University, Xining, People’s Republic of China; 4The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, People’s Republic of China
Correspondence: Tian-Ping Zhang, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, People’s Republic of China, Email [email protected] Hai-Feng Pan, Department of Epidemiology and Biostatistics, School of Public Health, Anhui Medical University, Hefei, People’s Republic of China, Email [email protected]
Objective: Our study aimed to explore the association of IL-13, IL-13RA1, and IL-13RA2 genes polymorphisms with PTB susceptibility and its clinical features.
Methods: Nine SNPs were genotyped by improved multiple ligase detection reaction (iMLDR) in 476 PTB patients and 473 controls. The association between these SNPs and PTB risk was analyzed using SPSS software and haplotype analysis was assessed using SHEsis software.
Results: The IL-13RA1 rs2495636 GA genotype frequency in PTB patients was significantly decreased, and IL-13RA2 rs5946039 A allele was related to the lower risk of PTB. In IL-13 gene, rs20541 variant was found to be associated with PTB risk under recessive mode. Moreover, IL-13RA1 rs141573089 C allele was significantly lower in PTB presenting with fever, drug resistance, and CC genotype was decreased in PTB presenting with leukopenia. IL-13RA1 rs2495636 polymorphism was associated with drug resistance, pulmonary infection, and IL-13RA2 rs3795175, rs638376 polymorphisms were related to drug resistance in PTB patients.
Conclusion: IL-13 rs20541, IL-13RA1 rs2495636, IL-13RA2 rs5946039 polymorphisms might be contributed to the genetic background of PTB in Chinese population.
Keywords: pulmonary tuberculosis, Mycobacterium tuberculosis, interleukin-13, single nucleotide polymorphisms
Introduction
Tuberculosis (TB) is a common infectious disease caused by Mycobacterium tuberculosis (MTB) infection, which could not only spread to the respiratory system but also to other body systems, and is the leading infectious cause of death worldwide.1 It is worth noting that there were estimated 9.9 million new incidents of TB patients globally in 2021.2 Approximately one third of the world’s population was infected with MTB, with only 10% of them developing active pulmonary TB (PTB),3,4 and this statistic remained largely unexplained. Increasing evidence suggested that genetic and environmental factors, as well as bacterial agents, play key roles in PTB development.5,6 Studying the significant roles of host genetic factors in TB disease and determining the specific host genes associated with PTB susceptibility were helpful to further understand the pathogenesis of PTB and formulate appropriate treatment strategies.
Additionally, immune pathways are important in controlling latent PTB infections, while the mechanisms underlying the progression from latent stage to the onset of active disease are still not fully understood.7 Complex interactions between lymphocytes, antigen-presenting cells, and secreted cytokines are involved in the immune response against PTB.3 Several inflammatory cytokines abnormally expressed in PTB could serve as biomarkers for disease severity and bacterial burden.8 Genetic variations in cytokine genes might alter the expression levels by affecting the recognition sites of transcription factors, and their transcriptional activity.9 Hence, polymorphisms of cytokine genes were closely related to PTB susceptibility.10–12 For example, IFN-γ-874 gene polymorphisms were reported to be related to PTB, and interleukin (IL)-10 gene variations contributed to the progression of PTB.
IL-13 was a cytokine secreted by activated T-helper 2 (Th2) cells, which involved in the regulation of humoral immunity and the type II cytokine-mediated immune response, and had many biological functions, including promoting inflammatory immune response and inhibiting the production of inflammatory cytokines.13,14 Study had shown that IL-13 was involved in the development of PTB, and the plasma level of IL-13 was significantly increased in PTB patients.7 In human TB, the relative increased expression of IL-13, which associated with lung damage, indicated that a subversive Th2 component in the response to MTB might undermine protective immunity and contribute to reactivation and tissue pathology.15 Recently, another study analyzed the association between IL-13 gene polymorphism and PTB susceptibility, while no meaningful result was found.16
IL-13 had two cognate receptors, known as IL-13RA1 and IL-13RA2, which specifically bind to IL-13 in vitro.17,18 The role of genetic variations in IL-13RA1 and IL-13RA2 in human disease had also been reported,19,20 however, similar studies had not been conducted in PTB patients. Therefore, to improve our understanding of the role of IL-13 in the pathogenesis of PTB, we conducted this study to explore the possible association between IL-13, IL-13RA1, and IL-13RA2 gene single nucleotide polymorphisms (SNPs) and susceptibility to PTB in a Chinese population.
Materials and Methods
Study Participants
In total, 476 PTB patients and 473 unrelated healthy individuals were included. All PTB patients were selected from Anhui Chest Hospital and were diagnosed using the following criteria: suspicious clinical symptoms, chest radiography, sputum and/or bronchoalveolar lavage fluid MTB culture, microscopy of acid fast bacilli (AFB), and the effects of anti-TB treatment. The patients with cancer, HIV positivity, hepatitis, and immune-compromised conditions were excluded, as well as those with poor compliance. Healthy individuals with the same ethnic background, no history of TB, cancer, and HIV were enrolled from the health examining center in the same area as controls. Moreover, all controls were examined by a senior physician to ensure that they were asymptomatic with negative sputum smear and culture, and normal chest radiograph.
This study was carried out after being approved by the ethics committee of Anhui Medical University (20200250) in complies with the Declaration of Helsinki. After obtaining informed consent, we collected peripheral blood samples and relevant data from the study subjects, including basic information such as age and sex, as well as some clinical data such as pulmonary infection, leukopenia, fever, drug resistance, drug-induced liver injury (DILI), sputum smear status.
DNA Extraction
Approximately 5 mL of peripheral blood was drawn from the medial cubital vein, placed in EDTA-containing tubes, and stored at −20°C until DNA extraction. The Flexi Gene-DNA Kit (Qiagen, Valencia, CA) was used to extract genomic DNA from the peripheral blood leukocytes with the standard procedures.
SNP Selection and Genotyping
The SNP selection methods mainly included literature searches and tag SNP selection. We systematically searched the previous studies on the relationship between IL-13, IL-13RA1, and IL-13RA2 gene polymorphisms and human diseases to look for the SNPs associated with human disease susceptibility. Meanwhile, we obtained genotype data of IL-13, IL-13RA1, and IL-13RA2 in CHB from the Ensembl Genome Browser 85 and CHBS_1000 g and used the pairwise option of the HaploView 4.0 software (Cambridge, MA, USA) to select the tag SNPs of these genes through linkage disequilibrium (LD) analysis with r2 threshold >0.8. The tag SNPs were selected with a minor allele frequency (MAF) ≥0.05 capturing all the common SNPs located in the chromosome locus transcribed into these genes and their flanking 2000 bp region. Finally, we selected IL-13 rs2066960, rs2069744, rs20541, IL-13RA1 rs147857000, rs141573089, rs2495636, IL-13RA2 rs3795175, rs638376, rs5946039 for genotyping.
Genetic polymorphisms were detected by an improved multiple ligase detection reaction (iMLDR) genotyping assay with the technical support of the Center for Genetic & Genomic Analysis, Genesky Biotechnologies (Inc., Shanghai). IMLDR is a commonly used, well-established genotyping method, which is widely used in our study and other studies, and its specific steps can be found in the previous studies.21,22 The detailed experimental steps were as follows: (1) 1μL DNA sample was extracted, and the quality of the sample was checked and the concentration was estimated by 1% agarose electrophoresis. Then, the DNA sample was diluted to 5–10 ng/μL according to the estimated concentration. (2) Multiplex PCR reaction was carried out with 20μL reaction system included 1x HotStarTaq bufer, 3.0 mMMg2+, 0.3mM dNTP, 1U HotStarTaq polymerase (Qiagen Inc.), 1μL sample DNA and 1μL multiple PCR primers. (3) Purification of multiple PCR products: 5U SAP enzyme and 2U Exonuclease I enzyme were added to 20 μL PCR product, 37°C warm bath for 1 h, then 75°C inactivated for 15 min. (4) Ligating reaction system: 1ul 10x ligating buffer, 0.25 ul high-temperature ligase, 0.4 ul 5’ ligating primer mixture (1μM), 0.4ul primer 3’ ligating primer mixture (2μM), 2ul purifed multiple PCR products, 6ul ddH2O mixing. (5) The 0.5 μL diluted product was mixed with 0.5 μL Liz500 SIZE STANDARD, 9 μL Hi-Di, denatured at 95°C for 5 min, then placed on the ABI3730XL sequencer. (6) The raw data collected on the ABI3730XL sequencer are analyzed by GeneMapper 4.1 (AppliedBiosystems, USA). Only the subjects with all nine SNP successfully genotyped were included in the final analysis.
Statistical Analysis
All statistical analyses were two-sided and conducted using SPSS (version 23.0; Armonk, NY: IBM Corp, USA). Hardy-Weinberg equilibrium (HWE) of all SNPs was evaluated by Chi-square (χ2) in normal controls. The differences in genotype, allele frequencies differences of all SNPs between PTB patients and normal controls were compared using the chi-square test (χ2), and logistic regression analysis was performed to calculate odds ratios (OR), 95% confidence intervals (CI). Haplotype analysis was assessed using SHEsis software,23 and the associations between all SNPs and PTB risk in two genetic models (dominant and recessive models) were also calculated. P < 0.05 was considered to be statistically significant. Bonferroni correction was used for multiple testing in SNP analysis, and P < 0.0055 (0.05/9) was considered to be statistically significant.
Results
Subject Characteristics
In this study, the mean ages of 476 PTB patients and 473 controls were 45.09 ± 17.79 years and 43.18 ± 13.68 years, respectively. The PTB group included 163 females and 313 males, while the control group consisted of 268 females and 205 males. In the PTB group, 129 (27.10%) patients had smear-positive sputum, the proportion of other clinical features were pulmonary infection (104, 21.84%), fever (83, 17.44%), drug resistance (73, 15.34%), DILI (68, 14.29%), hypoproteinemia (34, 7.14%), leukopenia (31, 6.51%).
Association of IL-13, IL-13RA1, and IL-13RA2 Genes Polymorphisms with PTB Susceptibility
The genotype distributions and allele frequencies of all SNPs in IL-13, IL-13RA1, and IL-13RA2 genes are shown in Table 1, and all the SNPs frequencies among in controls were conformed to HWE. The results demonstrated that IL-13 rs20541 AA genotype frequency appeared to be increased in PTB patients in comparison to normal controls (AA vs GG: P = 0.017 after adjustment of sex and age), while the difference was not statistically significant after Bonferroni correction (P > 0.0055). In addition, rs20541 variant was found to be associated with PTB risk under recessive mode (AA vs GG+GA: P = 0.001 after adjustment of sex and age). However, we did not find any association of rs2066960 and rs2069744 polymorphisms with the predisposition to PTB.
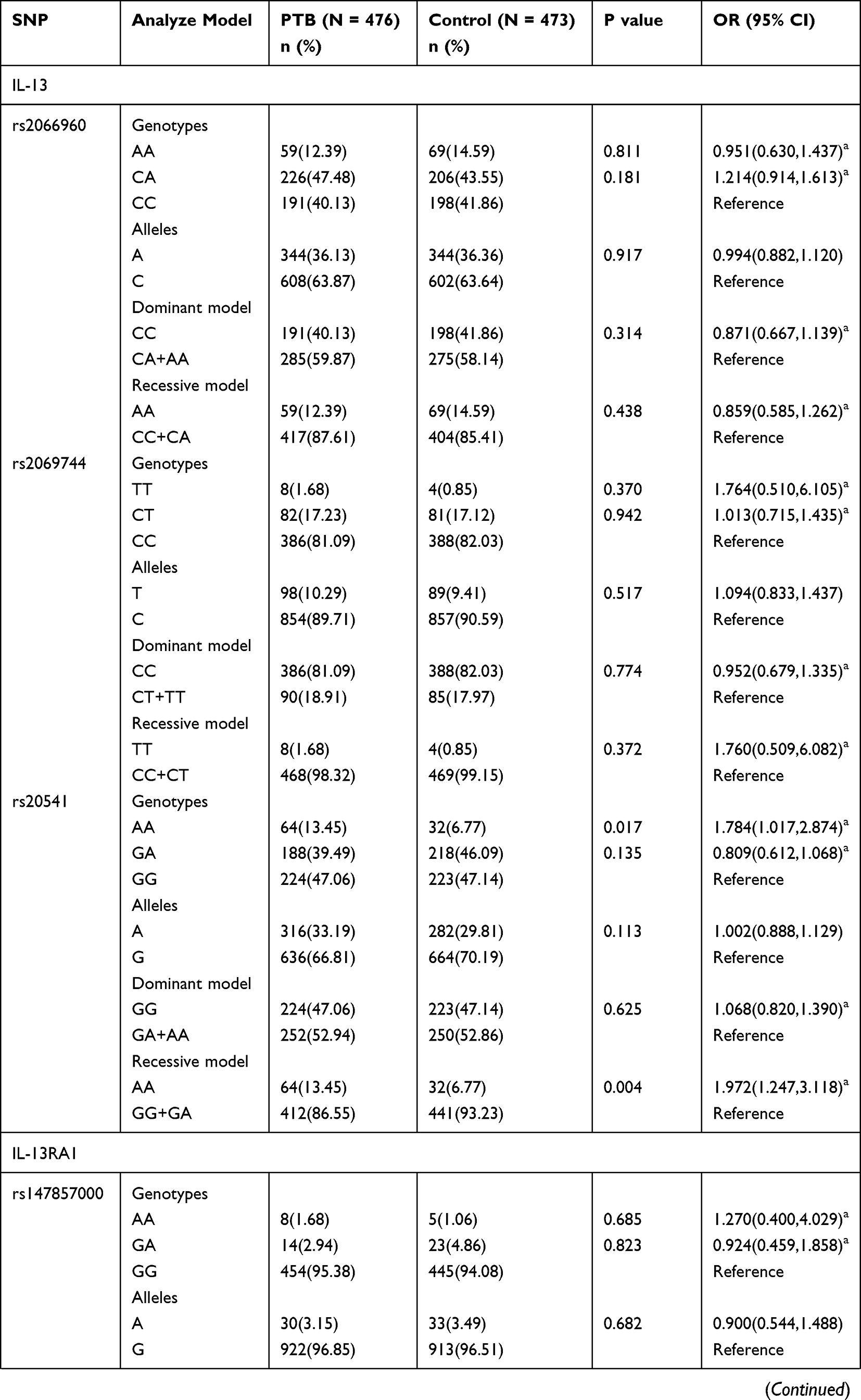 | 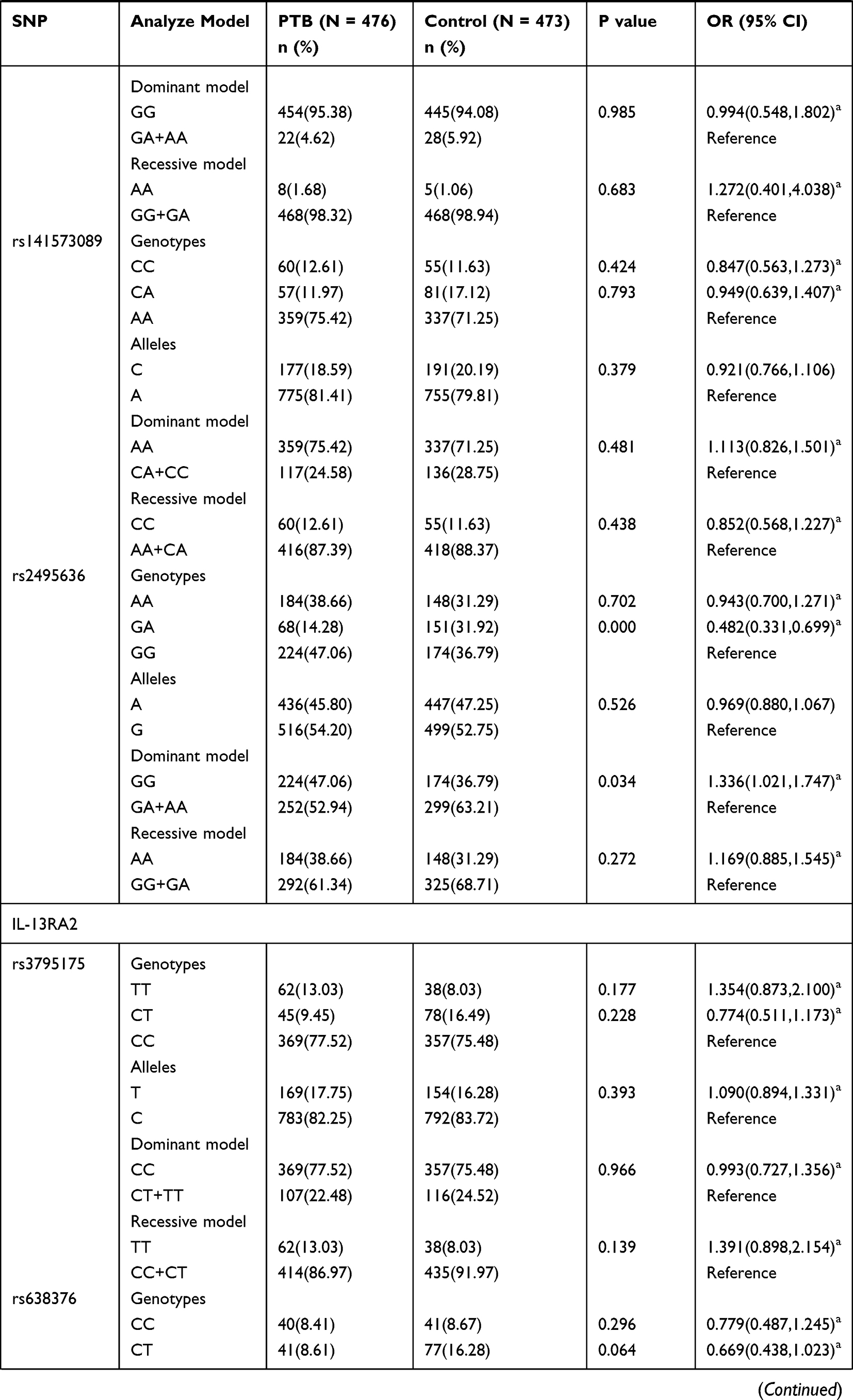 | 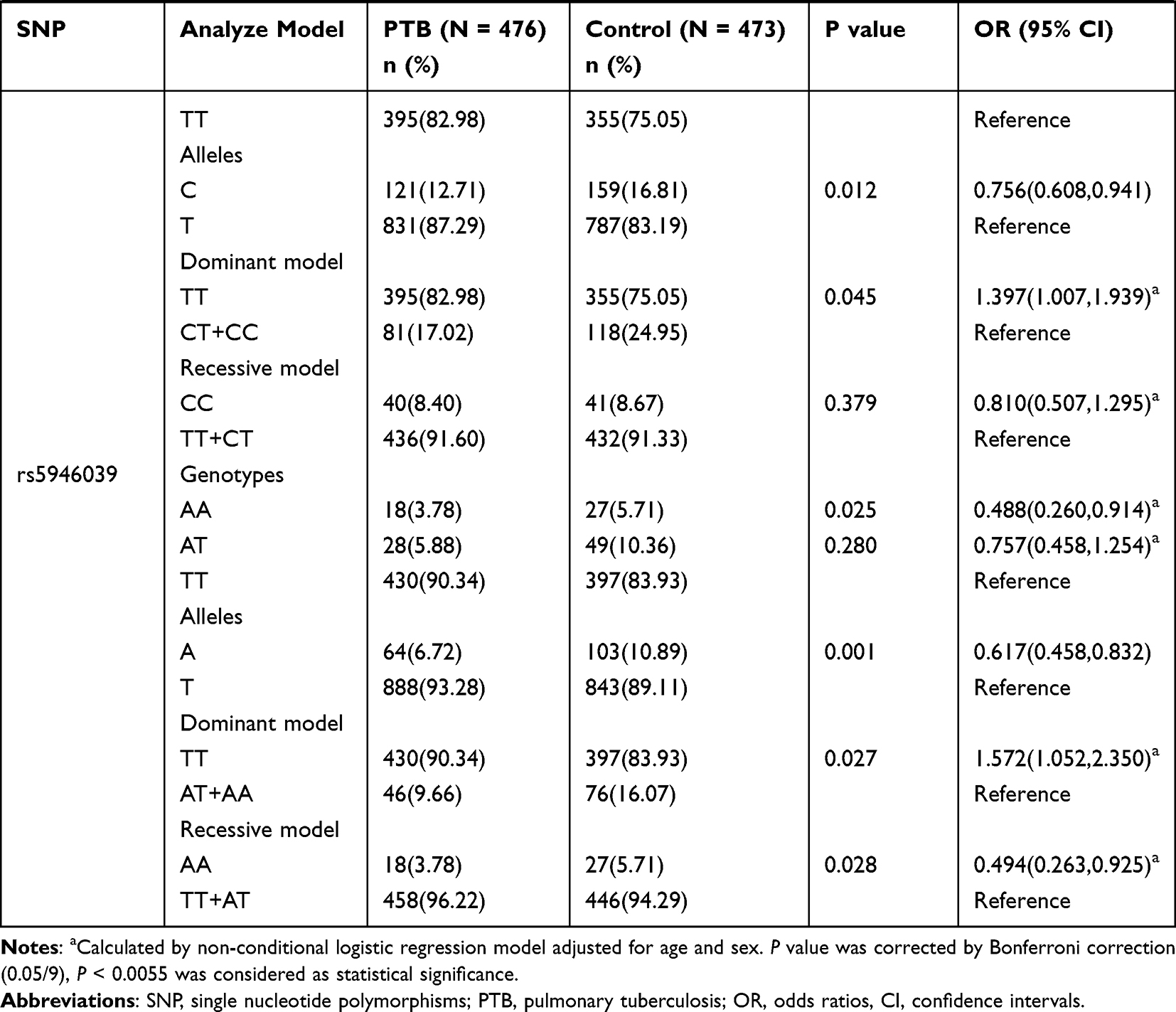 |
Table 1 Genotypes and Alleles Frequencies of IL-13, IL-13RA1, and IL-13RA2 Genes in PTB Patients and Normal Controls |
The IL-13RA1 rs2495636 GA genotype frequency in PTB patients was significantly decreased (GA vs GG: P = 0.000 after adjustment of sex and age), while the association between rs2495636 variant and PTB susceptibility under dominant mode was not reached statistically insignificant after Bonferroni correction (GG vs GA+AA: P > 0.0055). Regarding IL-13RA2 gene variations, we found that rs5946039 A allele was related to the lower risk of PTB (A vs T: P = 0.001 after adjustment of sex and age). The rs638376 CT genotype, C allele, rs5946039 AA genotype frequencies seemed to be associated with PTB susceptibility; however, no statistically significant association was found after Bonferroni correction (P > 0.0055).
Association of IL-13, IL-13RA1, and IL-13RA2 Genes SNPs with Clinical Features Among PTB Patients
A case-only analysis was performed to investigate the potential relationship between IL-13, IL-13RA1, and IL-13RA2 genes polymorphism and several common clinical features of PTB patients (Table S1). We noted that there was no significant relationship between IL-13 gene rs2066960, rs2069744, rs20541 and any clinical features in PTB patients.
In IL-13RA1 gene, the rs141573089 C allele frequency was significantly lower in PTB patients with fever, drug resistance when compared to the patients without these features, respectively (P = 0.009, P = 0.032, respectively), and the rs141573089 CC genotype frequency was significantly decreased in PTB patients with leukopenia (P = 0.015) (Table 2). Furthermore, the AA genotype and A allele frequencies of rs2495636 in PTB patients with drug resistance were both significantly higher than that in PTB patients without drug resistance (P = 0.049, P = 0.001, respectively), and the rs2495636 A allele frequency was also increased in PTB patients with pulmonary infection (P = 0.041).
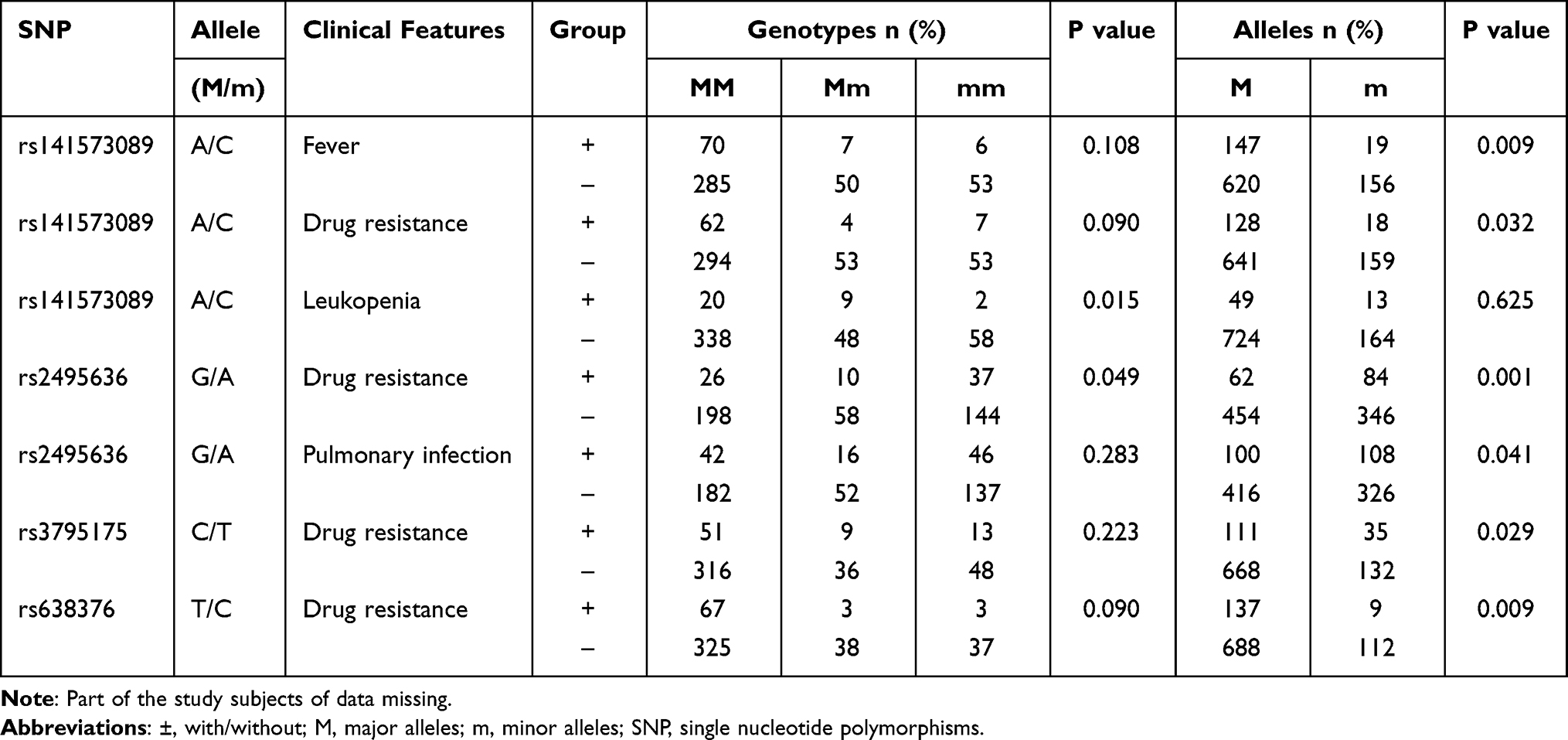 |
Table 2 The Positive Findings of Associations Between IL-13, IL-13RA1, and IL-13RA2 Genes Polymorphisms and Several Clinical Features of PTB Patients |
For the IL-13RA2 gene, when compared to PTB patients without drug resistance, the increased frequency of rs3795175 T allele, as well as the decreased frequency of rs638376 C allele, was found in PTB patients with drug resistance (P = 0.029, P = 0.009, respectively).
Haplotype Analysis
Five main haplotypes (ACA, ACG, CCA, CCG, CTA) for IL-13, four main haplotypes (AAG, GAA, GAG, GCG) for IL-13RA1, four main haplotypes (CCA, CCT, CTT, TTT) for IL-13RA2 were detected using SHEsis software. These haplotype frequency distributions between PTB patients and controls are summarized in Table 3.
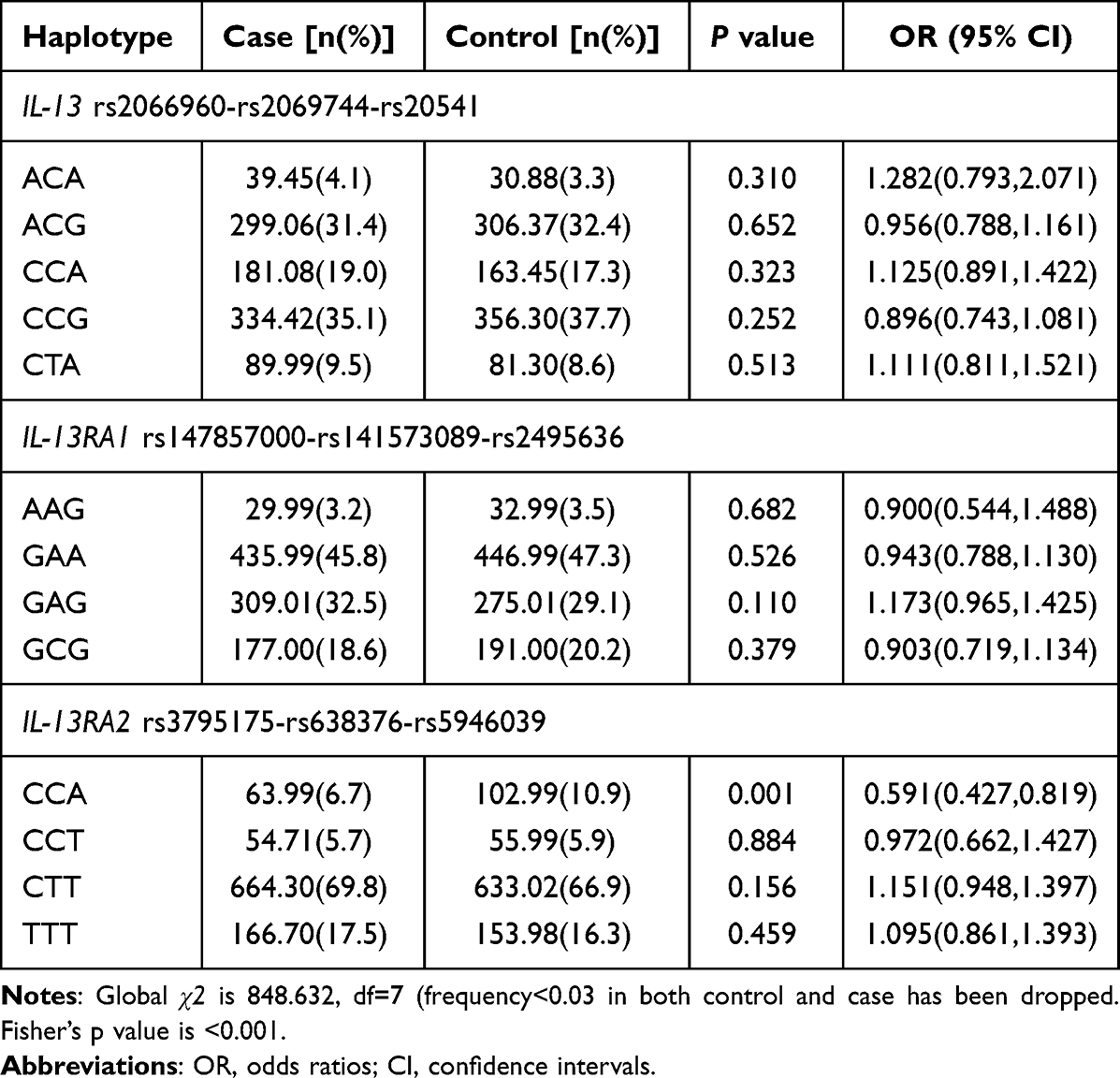 |
Table 3 Haplotype Analysis of IL-13, IL-13RA1, IL-13RA2 Genes in PTB Patients and Controls |
The results suggested that the frequency of IL-13RA2 CCA haplotype was significantly lower in PTB patients than normal controls, while there was no significant difference regarding the IL-13, IL-13RA1 haplotype frequencies between PTB patients and controls.
Discussion
Studies had increasingly focused on the genetic factors that affected the PTB risk, and many new genetic markers had been identified in previous studies.24,25 Additionally, since the susceptibility and severity of PTB were determined by complex immune interactions, discussing the role of immune-modulatory gene variants in PTB development was essential. Considering that T cell-derived IL-13 participated in promoting inflammation, regulating cytokine networks, and determining the outcome of PTB patients,26 we analyzed the association between three SNPs in IL-13 gene and susceptibility to PTB. Meanwhile, our study also examined the association between several SNPs of two IL-13 receptors (IL-13RA1, IL-13RA2) and PTB susceptibility. Finally, we found the IL-13 rs20541 variant was associated with PTB susceptibility, and significant associations were also observed for IL-13RA1 rs2495636, IL-13RA2 rs5946039 variants with PTB.
Abnormal IL-13 level in PTB patients suggested that IL-13 was closely related to the pathogenesis of PTB. Heitmann et al demonstrated that IL-13/IL-4Ra mediated mechanisms were involved in PTB-associated tissue pathology by regulating the expression level of arginase-1 in an experimental PTB model.15 SNPs in IL-13 were also associated with various human diseases, including renal cell carcinoma, colorectal cancer, breast cancer, and bladder cancer.27–29 Hence, the potential association between IL-13 gene variation and PTB susceptibility also attracted more attention. Sun et al explored the association of IL-13 gene polymorphisms (rs2066960, rs1295686, rs20541, rs2069757 and rs2243248) with PTB susceptibility in a Western Chinese Han population and did not find any associations.16 Consistent with these results, we observed that the allele and genotype frequencies of rs20541 were not associated with the increased risk of PTB. Meanwhile, genetic model analysis showed that rs20541 was associated with an increased PTB susceptibility in recessive mode, suggesting that rs20541 variant might be involved in the risk of PTB. We selected two other SNPs (rs2066960 and rs2069744) of IL-13 for genotyping, but no correlation was found. Similarly, the authors found that IL-13 rs2066960, rs2069744 had no relationship with the risk of renal cell carcinoma in another study.30 Currently, research on the association between IL-13 and PTB was very limited, and our results contributed to further understanding of the role of IL-13 gene variation in the pathogenesis of PTB. It was important to noted that increased sample sizes and additional SNP might need to be explored.
IL-13 mediated its effects through interacting with its cognate receptors (IL-13RA1, IL-13RA2) on a variety of biological processes, and the influence of several SNPs in IL-13RA1, IL-13RA2 genes had also been studied in some diseases, such as asthma, atopic dermatitis, and systemic sclerosis.19,31,32 The results by Namkung et al identified an association between IL-13RA1 rs2265753, rs2254672 and atopic dermatitis phenotype.19 In another study, no significant association was found between IL-13RA1 +1398 genotype frequencies in asthma, allergic rhinitis patients when compared to in controls.32 Granel et al suggested that IL-13RA2 rs638376 was related to the risk of systemic sclerosis.31 Whereas no studies had been conducted to explore the association between IL-13RA1, IL-13RA2 gene variation and PTB susceptibility, our study was the first to analyze the association between several SNPs in IL-13RA1, IL-13RA2 genes and PTB susceptibility. We found that IL-13RA1 rs2495636 GA genotype was significantly decreased in PTB patients, suggesting that this SNP might contribute to susceptibility to PTB. Moreover, the results demonstrated that the reduced frequency of rs5946039 A allele existed in PTB patients. In haplotype analysis, IL-13RA2 CCA haplotype was also proved to be associated with PTB risk. We hypothesized that IL-13RA1, IL-13RA2 gene polymorphism might be involved in the pathogenesis of PTB by modulating IL-13 function, and repetitive studies were necessary to verify our experimental results. Furthermore, the functional role of IL-13RA1, IL-13RA2 genes polymorphism in the development of PTB is worth further investigation.
As a complex disease, PTB usually has multiple clinical manifestations and complications, such as drug resistance, pulmonary infection, fever, hypoproteinemia, and leukopenia, which may greatly impact treatment and rehabilitation. Many variations were found to be associated with clinical features.33,34 Song et al found that lncRNA RP11-37B2.1 rs218916 and rs160441 were associated with thrombocytopenia in PTB patients.33 Our recent study found a significant association between CYP27A1 rs933994 T allele frequency and drug resistance in PTB patients.4 Similarly, IL-13RA1 gene rs141573089 polymorphism was found to be related to fever, drug resistance, and leukopenia, while rs2495636 polymorphisms were closely associated with pulmonary infection and drug resistance in PTB patients. These results indicated that the variants of IL-13RA1, IL-13RA2 contributed to host response to drug treatment and played an active role in the development of appropriate treatment choices for PTB patients. However, the mechanism underlying how they affect the drug treatment response was unclear; hence, more rigorous studies at the molecular genetic level were needed.
Some limitations should be noted in our study. First, this study was conducted in a single center; therefore, the sample size might not be sufficient. Second, the potential influence of some confounding factors, such as environmental factors, treatment regimen, and so on, was not excluded. Finally, we only studied several SNPs within IL-13, IL-13RA1, IL-13RA2, other functional SNPs of these genes warrant further investigation. Further studies with large-scale samples and other ethnic backgrounds were warranted to determine the precise role of IL-13, IL-13RA1, and IL-13RA2 gene variations in PTB.
Conclusion
Our study demonstrated that IL-13 rs20541, IL-13RA1 rs2495636, IL-13RA2 rs5946039 polymorphisms might be associated with PTB susceptibility in Chinese population. Moreover, several SNPs in IL-13RA1 and IL-13RA2 genes were related to some clinical features among PTB patients, including leukopenia, pulmonary infection, and drug resistance.
Abbreviations
TB, tuberculosis; IL, interleukin; MAF, minor allele frequency; IMLDR, improved multiple ligase detection reaction; HWE, Hardy-Weinberg equilibrium; SNPs, single nucleotide polymorphisms; MTB, Mycobacterium tuberculosis; LD, linkage disequilibrium.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (82003515) and Anhui Provincial Medical and Health Key Specialty Construction Project (No.[2021]273).
Author Contributions
All authors made a significant contribution to study design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically; agreed to submit to the current journal; gave final approval for the version to be published; and agreed to be accountable for all aspects of the work.
Hong-Miao Li and Fei Tang should be considered as co-first authors.
Disclosure
The authors declare that they have no conflict of interest.
References
1. Morishita F, Viney K, Lowbridge C, et al. Epidemiology of tuberculosis in the western pacific region: progress towards the 2020 milestones of the end TB strategy. Western Pac Surveill Response J. 2020;11:10–23. doi:10.5365/wpsar.2020.11.3.002
2. World Health Organization. Global tuberculosis report; 2021. Available from: https://www.who.int/tb/publications/global_report/en/.
3. Wu S, Wang MG, Wang Y, He JQ. Polymorphisms of cytokine genes and tuberculosis in two independent studies. Sci Rep. 2019;9:2507. doi:10.1038/s41598-019-39249-4
4. Zhang TP, Chen SS, Zhang GY, Shi SJ, Wei L, Li HM. Association of vitamin D pathway genes polymorphisms with pulmonary tuberculosis susceptibility in a Chinese population. Genes Nutr. 2021;16:6. doi:10.1186/s12263-021-00687-3
5. Longhi RM, Zembrzuski VM, Basta PC, Croda J. Genetic polymorphism and immune response to tuberculosis in indigenous populations: a brief review. Braz J Infect Dis. 2013;17:363–368. doi:10.1016/j.bjid.2012.11.001
6. Fouad NA, Saeed AM, Mahedy AW. Toll like receptor-4 gene polymorphism and susceptibility to pulmonary tuberculosis. Egypt J Immunol. 2019;26:1–10.
7. Moideen K, Kumar NP, Bethunaickan R, Banurekha VV, Nair D, Babu S. Heightened systemic levels of anti-inflammatory cytokines in pulmonary tuberculosis and alterations following anti-tuberculosis treatment. Cytokine. 2020;127:154929. doi:10.1016/j.cyto.2019.154929
8. Kumar NP, Moideen K, Banurekha VV, Nair D, Babu S. Plasma Proinflammatory cytokines are markers of disease severity and bacterial burden in pulmonary tuberculosis. Open Forum Infect Dis. 2019;6:ofz257. doi:10.1093/ofid/ofz257
9. Yim JJ, Selvaraj P. Genetic susceptibility in tuberculosis. Respirology. 2010;15:241–256. doi:10.1111/j.1440-1843.2009.01690.x
10. Yu ZG, Wang BZ, Li J, Ding ZL, Wang K. Association between interleukin-17 genetic polymorphisms and tuberculosis susceptibility: an updated meta-analysis. Int J Tuberc Lung Dis. 2017;21(12):1307–1313. doi:10.5588/ijtld.17.0345
11. Hu Y, Wu L, Li D, Jiang W, Xu B, Xu B. Association between cytokine gene polymorphisms and tuberculosis in a Chinese population in Shanghai: a case-control study. BMC Immunol. 2015;16:8. doi:10.1186/s12865-015-0071-6
12. Wu SQ, Wang Y, Zhang MM, Shrestha SS, He JQ, He J-Q. Genetic polymorphisms of IL1B, IL6, and TNF α in a Chinese han population with pulmonary tuberculosis. Biomed Res Int. 2018;2018:3010898. doi:10.1155/2018/3010898
13. Rengarajan J, Szabo SJ, Glimcher LH. Transcriptional regulation of Th1/Th2 polarization. Immunol Today. 2000;21:479–483. doi:10.1016/S0167-5699(00)01712-6
14. Roy S, Schmeier S, Kaczkowski B, et al. Transcriptional landscape of Mycobacterium tuberculosis infection in macrophages. Sci Rep. 2018;8:6758. doi:10.1038/s41598-018-24509-6
15. Heitmann L, Abad Dar M, Schreiber T, et al. The IL-13/IL-4Ralpha axis is involved in tuberculosis-associated pathology. J Pathol. 2014;234:338–350. doi:10.1002/path.4399
16. Sun W, Jiao L, Liu T, et al. No significant effects of IL-6 and IL-13 gene variants on tuberculosis susceptibility in the Chinese population. DNA Cell Biol. 2020;39:1356–1367. doi:10.1089/dna.2020.5404
17. Seyfizadeh N, Seyfizadeh N, Gharibi T, Babaloo Z. Interleukin-13 as an important cytokine: a review on its roles in some human diseases. Acta Microbiol Immunol Hung. 2015;62:341–378. doi:10.1556/030.62.2015.4.2
18. Hilton DJ, Zhang JG, Metcalf D, Alexander WS, Nicola NA, Willson TA. Cloning and characterization of a binding subunit of the interleukin 13 receptor that is also a component of the interleukin 4 receptor. Proc Natl Acad Sci U S A. 1996;93:497–501. doi:10.1073/pnas.93.1.497
19. Namkung JH, Lee JE, Kim E, et al. Association of polymorphisms in genes encoding IL-4, IL-13 and their receptors with atopic dermatitis in a Korean population. Exp Dermatol. 2011;20:915–919. doi:10.1111/j.1600-0625.2011.01357.x
20. Aguirre CA, Concetta Morale M, Peng Q, et al. Two single nucleotide polymorphisms in IL13 and IL13RA1 from individuals with idiopathic Parkinson’s disease increase cellular susceptibility to oxidative stress. Brain Behav Immun. 2020;88:920–924. doi:10.1016/j.bbi.2020.04.007
21. Li HM, Wang LJ, Tang F, Pan HF, Zhang TP. Association of leptin and leptin receptor genes variants and pulmonary tuberculosis susceptibility, clinical manifestations in a Chinese population. Microb Pathog. 2022;165:105499. doi:10.1016/j.micpath.2022.105499
22. Qi Y, Wei Y, Yu F, et al. Association study of a genetic variant in the long intergenic noncoding RNA (linc01080) with schizophrenia in Han Chinese. BMC Psychiatry. 2021;21:613. doi:10.1186/s12888-021-03623-2
23. Li Z, Zhang Z, He Z, et al. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: update of the SHEsis. Cell Res. 2009;19:519–523. doi:10.1038/cr.2009.33
24. Harishankar M, Selvaraj P, Bethunaickan R. Influence of genetic polymorphism towards pulmonary tuberculosis susceptibility. Front Med. 2018;5:213. doi:10.3389/fmed.2018.00213
25. Li M, Hu Y, Zhao B, et al. A next generation sequencing combined genome-wide association study identifies novel tuberculosis susceptibility loci in Chinese population. Genomics. 2021;113:2377–2384. doi:10.1016/j.ygeno.2021.05.035
26. Corren J. Role of interleukin-13 in asthma. Curr Allergy Asthma Rep. 2013;13:415–420. doi:10.1007/s11882-013-0373-9
27. Walczak A, Przybyłowska K, Trzciński R, et al. Association of −1112 c/t promoter region polymorphism of the interleukin 13 gene with occurrence of colorectal cancer. Pol Przegl Chir. 2011;83:27–31. doi:10.2478/v10035-011-0004-x
28. Chu H, Ma L, Wang M, et al. The polymorphisms of IL-4, IL-4R and IL-13 genes and bladder cancer risk in a Chinese population: a case-control study. Mol Biol Rep. 2012;39:5349–5357. doi:10.1007/s11033-011-1334-9
29. Erdei E, Kang H, Meisner A, et al. Polymorphisms in cytokine genes and serum cytokine levels among new Mexican women with and without breast cancer. Cytokine. 2010;51:18–24. doi:10.1016/j.cyto.2010.03.014
30. Lin Y, Yuan Q, Qian F, et al. Polymorphism rs4787951 in IL-4R contributes to the increased risk of renal cell carcinoma in a Chinese population. Gene. 2019;685:242–247. doi:10.1016/j.gene.2018.11.070
31. Granel B, Allanore Y, Chevillard C, et al. IL13RA2 gene polymorphisms are associated with systemic sclerosis. J Rheumatol. 2006;33:2015–2019.
32. Hussein YM, El-Tarhouny SA, Shalaby SM, et al. Interleukin-13 receptor A1 gene polymorphism and IL-13 serum level in atopic and non-atopic Egyptian children. Immunol Invest. 2011;40:523–534. doi:10.3109/08820139.2011.565106
33. Song J, Liu T, Zhao Z, et al. Genetic polymorphisms of long noncoding RNA RP11-37B2.1 associate with susceptibility of tuberculosis and adverse events of antituberculosis drugs in west China. J Clin Lab Anal. 2019;33:e22880. doi:10.1002/jcla.22880
34. Robert J, Morvan VL, Smith D, Pourquier P, Bonnet J. Predicting drug response and toxicity based on gene polymorphisms. Crit Rev Oncol Hem. 2005;54:
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Methylation in B7-H4 and BTLA Genes are Associated with the Risk of Pulmonary Tuberculosis
Cai XQ, Huang Q, Zhang TP
ImmunoTargets and Therapy 2023, 12:149-163
Published Date: 24 November 2023