Back to Journals » Journal of Hepatocellular Carcinoma » Volume 13
Intratumoral Tertiary Lymphoid Structures in Hepatocellular Carcinoma: Current Evidence and Future Directions – a Narrative Review
Authors Liu L, Wang Y, Ban J, Kang M, Li Y, Li Q, Zhang H ![]() , Cai P, Chen W, Li X, Li X
, Cai P, Chen W, Li X, Li X
Received 11 January 2026
Accepted for publication 14 March 2026
Published 21 March 2026 Volume 2026:13 595363
DOI https://doi.org/10.2147/JHC.S595363
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Mohamed Shaker
Lizhen Liu,1,* Yanfen Wang,2,* Jiawei Ban,3,* Manshu Kang,4,* Yiman Li,1 Qingrui Li,5 Huarong Zhang,5 Ping Cai,1 Wei Chen,1 Xinwei Li,6 Xiaoming Li1,6
1Department of Radiology, Southwest Hospital, Third Military Medical University (Army Military Medical University), Chongqing, People’s Republic of China; 2Department of Radiology, The Affiliated Hospital of the Non-Commissioned Officer (NCO) School, The Army Medical University, Shijiazhuang, Hebei, People’s Republic of China; 3Department of Radiology, the People’s Hospital of Lincang, Lincang, Yunnan, People’s Republic of China; 4Department of Radiology, Hailun City People’s Hospital, Hailun, Heilongjiang, People’s Republic of China; 5Institute of Pathology and Southwest Cancer Center, Third Military Medical University (Army Military Medical University), Chongqing, People’s Republic of China; 6School of Bioinformatics, Chongqing University of Posts and Telecommunications, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xinwei Li; Xiaoming Li, Email [email protected]; [email protected]
Abstract: Intratumoral tertiary lymphoid structures (iTLSs) have emerged as critical immune features in hepatocellular carcinoma (HCC). This narrative review critically synthesizes current evidence (sourced from PubMed, Embase, and Web of Science up to January 2026) on the biological mechanisms, pathological assessment, non-invasive imaging, and clinical implications of iTLSs. Moving beyond simple binary classifications, we emphasize that precise scoring based on morphological maturation is essential. Clinically, functionally mature iTLSs are strongly associated with favorable prognosis and immunotherapy benefits, though metabolic etiologies (e.g, NASH) can drive complex immunosuppression. Furthermore, while non-invasive radiomic models show high predictive accuracy, their clinical translation is hindered by mathematical overfitting and a “black box” lack of biological interpretability. Translating these biological insights into clinical practice, particularly through non-invasive imaging biomarkers and standardized pathological evaluations, holds great promise for guiding personalized immunotherapy. Ultimately, however, overcoming inter-study heterogeneity and conducting rigorous functional validations remain imperative before their routine clinical integration.
Keywords: hepatocellular carcinoma, tertiary lymphoid structures, formation mechanisms, imaging, prognosis
Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver malignancy, accounting for nearly 90% of all hepatic cancers worldwide. China bears almost half of the global HCC burden, mainly due to the high prevalence of chronic hepatitis B virus (HBV) infection, aflatoxin exposure, and metabolic liver disease.1 Despite progress in diagnosis and treatment, the prognosis of HCC remains poor, with a 5-year recurrence rate after curative resection reaching approximately 60–70%.2
Current treatments for HCC include surgery, liver transplantation, locoregional therapies, and systemic therapy, but long-term outcomes remain unsatisfactory for advanced disease. In recent years, combinations of immune checkpoint inhibitors and targeted agents-such as atezolizumab plus bevacizumab or durvalumab plus tremelimumab-have significantly improved survival and response rates, marking a new era of immunotherapy-based treatment,3–5 emphasizing the importance of understanding the tumor immune microenvironment in predicting therapeutic response and improving clinical outcomes.
With growing recognition of the immune microenvironment’s role in HCC, increasing attention has focused on tertiary lymphoid structures (TLSs). TLSs are ectopic immune aggregates composed of B cells, T cells, and dendritic cells, which develop in response to chronic inflammation or antigenic stimulation.6 TLSs in HCC can be categorized into intratumoral and peritumoral types, each showing distinct biological and clinical implications.7 Intratumoral TLSs (iTLSs) are operationally defined as organized lymphoid aggregates located directly within the tumor bed. Given the heterogeneity in the literature, it is crucial to recognize that iTLSs exist on a continuum of maturation. They are generally classified into immature iTLSs (simple lymphoid aggregates or primary follicles lacking germinal centers) and mature iTLSs (secondary follicle-like structures characterized by well-developed germinal centers and follicular dendritic cell networks). Emerging evidence presents conflicting findings regarding the prognostic role of peritumoral TLSs. Some have suggested that higher peritumoral TLSs density is associated with enhanced local immune activation and improved survival,7,8 whereas other studies have reported that peritumoral TLSs show either no significant association or even a negative correlation with prognosis,9,10 which may stem from methodological inconsistencies (eg, varying spatial definitions) and clinical heterogeneities (such as underlying etiologies and tumor stages). In contrast, based on current evidence predominantly derived from surgical resection and liver transplantation cohorts, viral-associated etiologies, and emerging immunotherapy trials, there is growing consensus on the roles of iTLSs in modulating the tumor microenvironment and guiding patient stratification.11–14
Understanding iTLSs may help refine immune-based prognostic assessment and guide individualized therapeutic strategies in HCC. Therefore, the primary purpose of this review is to critically synthesize current evidence on iTLSs in HCC and establish a multidimensional framework that explicitly separates their mere structural presence, morphological maturation state, and underlying immune functionality. The scope of this comprehensive review encompasses four key domains: (1) the biological mechanisms driving iTLSs formation across different etiologies; (2) precise histopathological definitions and maturity scoring; (3) the emerging role and limitations of non-invasive imaging predictions; and (4) their clinical implications for prognosis and immunotherapy response.
Search Strategy and Literature Selection
This review was conducted as a structured narrative review. To enhance methodological transparency and comprehensiveness, a literature search was performed across major electronic databases, including PubMed, Embase, and Web of Science. The search strategy utilized combinations of the following keywords and Medical Subject Headings (MeSH) terms: (“tertiary lymphoid structures” OR “tertiary lymphoid organs” OR “ectopic lymphoid structures” OR “TLS”) AND (“hepatocellular carcinoma” OR “liver cancer” OR “HCC”). The search encompassed articles published from database inception to January 2026, with a primary focus on recent high-impact studies from the last five years. Inclusion criteria were restricted to peer-reviewed original research articles, robust clinical trials, and comprehensive reviews published in English. Two authors independently screened the titles and abstracts to identify studies reporting on the formation mechanisms, pathological assessment, non-invasive imaging, and prognostic or predictive value of iTLSs in HCC. Selected articles were critically appraised, with particular attention paid to the study design, etiologic context, and the specific criteria used to define and score iTLSs maturation.
Formation Mechanisms of iTLSs
The formation and functional maturation of iTLSs within the tumor microenvironment (TME) are intricately linked to chronic inflammation, immune cell interactions, tumor microenvironment remodeling, and immune regulatory pathways. Emerging mechanistic studies have highlighted several key pathways driving the formation of iTLSs in HCC (Figure 1). However, it is important to note that while some pathways are supported by HCC-specific data, others are currently extrapolated from broader pan-cancer mechanistic literature.
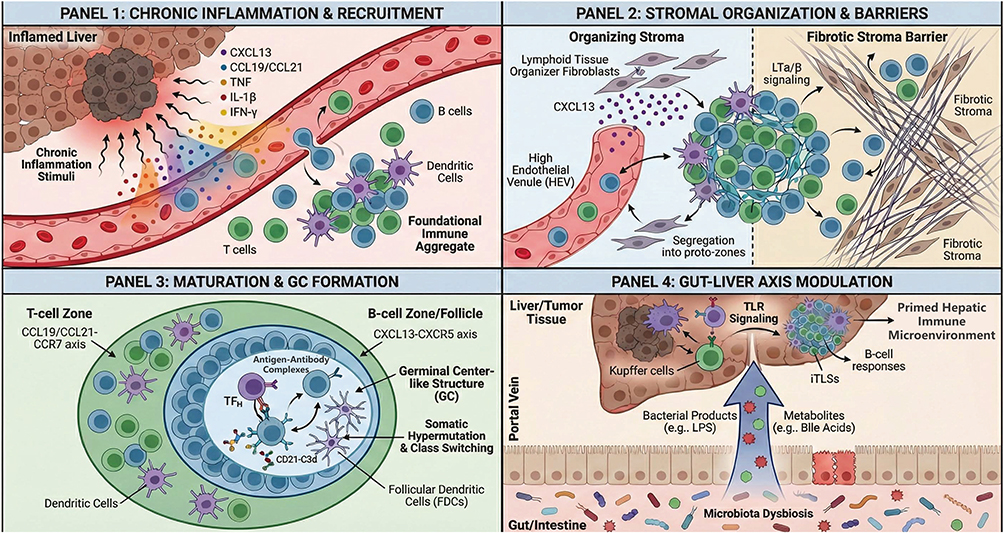 |
Figure 1 Schematic illustration of multiple parallel mechanisms regulating intratumoral tertiary lymphoid structures formation. It details the progression from chronic inflammation-driven immune recruitment and stromal-mediated organization to the establishment of functional germinal center-like structures. Additionally, the schematic highlights the critical immunomodulatory role of the gut-liver axis via portal vein microbial signaling. Abbreviations: CXCL, C-X-C motif chemokine ligand; CCL, C-C motif chemokine ligand; TNF, tumor necrosis factor; IL, interleukin; IFN-γ, interferon gamma; HEV, high endothelial venule; LT α/β, lymphotoxin alpha/beta; GC, germinal center; TFH, follicular helper T cell; FDC, follicular dendritic cell; TLR, Toll-like receptor; LPS, lipopolysaccharide; iTLSs, intratumoral tertiary lymphoid structures. |
Chronic Inflammation as a Proposed Driver of iTLSs Formation
The formation of iTLSs in HCC is frequently associated with chronic inflammation, a hallmark of liver diseases such as viral hepatitis and non-alcoholic steatohepatitis. These conditions result in persistent inflammatory stimuli that initiate the formation of TLSs by mimicking lymphoid organogenesis. Key cytokines, such as CXCL13, CCL19/21,15 and other inflammatory mediators like tumor necrosis factor (TNF), IL-1β, and IFN-γ, are upregulated in response to the chronic inflammatory environment. These cytokines are proposed to mediate the recruitment of B cells, T cells, and dendritic cells (DCs) to the tumor site, which then aggregate into the foundational structure for iTLSs.16–18 This cytokine-driven process supports the formation of immune cell-rich regions that mimic the early stages of secondary lymphoid tissue development.
Immune-Stromal Interactions and Structural Organization
Once immune cells are recruited to the tumor site, immune-stromal interactions play a critical role in organizing iTLSs. The lymphotoxin-α/β (LTα/β) signaling pathway is suggested to be a central mediator of this process, activating nuclear factor-κB (NF-κB) signaling pathways, which are essential for establishing the architecture of iTLSs.19 However, the stromal landscape in HCC is highly heterogeneous. While specific subsets of fibroblasts act as lymphoid tissue organizers to promote iTLSs assembly via CXCL13 secretion,20 others contribute to a dense fibrotic stroma. This fibrosis can act as a physical barrier, excluding lymphocytes and hindering the spatial organization required for iTLSs formation.21 Despite these barriers, successful stromal remodeling facilitates the segregation of immune cells into distinct T-cell and B-cell zones. Additionally, the formation of high endothelial venules (HEVs) supports continuous lymphocyte trafficking.22 This process leads to the stabilization of the iTLSs structure, ensuring that immune responses against tumor cells can be sustained over time.
Immunoregulatory Factors and iTLSs Maturation
The maturation and functional competence of iTLSs in HCC are critically dependent on the establishment of spatially organized T-cell and B-cell zones and the subsequent formation of germinal center-like (GC-like) structures, which represent the defining hallmark of mature iTLSs.23 Lymphocyte compartmentalization is orchestrated by coordinated chemokine axes, with the CCL19/CCL21-CCR7 pathway directing T-cell positioning and the CXCL13-CXCR5 axis driving B-cell recruitment and follicle formation.24,25 Following T/B cell segregation, GC development is initiated by the recruitment of follicular helper T (TFH) cells into the B-cell zone, thereby promoting B-cell proliferation, class-switch recombination, and somatic hypermutation.16 In parallel, follicular dendritic cell (FDC) differentiation, predominantly driven by TNFR1 signaling, establishes a reticular network capable of capturing and retaining antigen-antibody complexes through CD21-C3d-dependent mechanisms, ensuring sustained antigen presentation and GC stability.26 Collectively, TFH cells and FDCs create a specialized microenvironment that enables effective humoral immune responses within iTLSs. Conversely, iTLSs maturation can be constrained by immunosuppressive elements of the HCC microenvironment, including regulatory T cells, tumor-associated macrophages, and tumor-intrinsic metabolic pathways,27–29 contributing to the marked heterogeneity in iTLSs structure and function observed.
Role of the Tumor Microbiome in iTLSs Formation
Emerging evidence suggests that the microbiota serves as an important modulator of iTLSs formation. Locally, the intratumoral microbial community can activate pattern-recognition receptors (PRRs) on immune cells, triggering signaling pathways that promote immune recruitment.30,31 More importantly, given the unique anatomical link between the gut and the liver, the “Gut-Liver Axis” plays a pivotal role. Translocation of bacterial products (eg, LPS) and metabolites (eg, bile acids) via the portal vein can prime hepatic cells through Toll-like receptor (TLR) signaling. This priming modulates the hepatic immune microenvironment, suggesting that gut dysbiosis could be a critical distal regulator of intratumoral lymphoid neogenesis in HCC.32 Specifically, microbial components can enhance B-cell responses, potentially fostering the development of iTLSs.33 However, much of the current mechanistic understanding is extrapolated from other gastrointestinal cancers, and direct HCC-specific evidence remains nascent. Furthermore, evaluating the microbiome’s impact on iTLSs in HCC is heavily confounded by clinical variables such as prior antibiotic exposure, the severity of underlying cirrhosis, specific liver disease etiologies, and recent treatment histories. While promising, the precise causal mechanisms linking specific bacterial loads to iTLSs density remain an active area of investigation.
Histopathological Definition and Controversies of iTLSs
In HCC, iTLSs have attracted increasing attention as potential determinants of antitumor immunity and clinical outcome. However, despite rapidly expanding literature, the pathological definition, classification, and assessment of iTLSs remain subjects of ongoing debate, posing challenges to data interpretation and cross-study comparability.
Current literatures demonstrate that iTLSs display a spectrum of maturation states rather than a simple binary presence or absence.16,34,35 Pathologically, iTLSs are commonly classified into three subtypes according to their structural organization and maturation status. Lymphoid aggregates: consist of loosely arranged clusters of more than 100 lymphocytes with poorly defined borders, containing scattered CD3⁺ T cells and CD20⁺ B cells, with occasional high endothelial venules (identifiable by PNAd) but no follicular dendritic cell (FDC) networks. Primary follicle–like structures: display organized lymphoid architecture without GC, characterized by centrally located CD20⁺ B cells, peripherally distributed CD3⁺ T cells, and the presence of CD21⁺/CD23− FDC networks. Secondary follicle–like structures: represent fully mature iTLSs with active germinal centers (marked by BCL6 and Ki67 positivity) and well-developed FDC networks expressing both CD21 and CD2336,37 (Figure 2). To ensure reproducible and operational assessment across studies, we strongly recommend a core immunohistochemical or multiplex panel encompassing CD3, CD20, CD21/CD23, BCL6/Ki67, and PNAd.
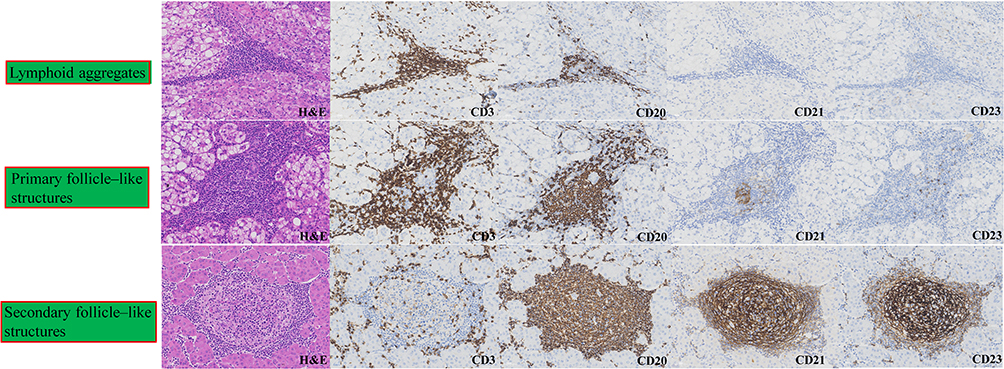 |
Figure 2 Representative histological and immunohistochemical profiles of intratumoral tertiary lymphoid structures (iTLSs) in hepatocellular carcinoma. iTLSs within the tumor microenvironment are categorized into three developmental stages based on their organizational complexity and cellular composition. Top row: Lymphoid aggregates characterized by an immature, disorganized collection of CD3+ T cells and CD20+ B cells. Middle row: Primary follicle-like structures showing initial compartmentalization of T and B cell zones with rudimentary follicular dendritic cell (FDC) networks (focal CD21+ and minimal CD23+ staining). Bottom row: Secondary follicle-like structures representing mature iTLSs with well-defined germinal centers, featuring a dense, organized meshwork of FDCs (strong and extensive CD21+ and CD23+ immunoreactivity). All images are original representative sections from our patient cohort. The leftmost column displays Hematoxylin and Eosin (H&E) staining, while subsequent columns display immunohistochemistry on sequential sections. Scale bars = 50 μm. Abbreviations: H&E, Hematoxylin and Eosin; CD, cluster of differentiation; FDC, follicular dendritic cell; iTLSs, intratumoral tertiary lymphoid structures. |
Although some studies broadly classify dense lymphoid aggregates within tumors as iTLSs,38–40 this definition remains controversial. Increasing evidence suggests that iTLSs represent a continuum of maturation rather than a binary presence or absence. In this framework, simple lymphoid aggregates are considered early, immature forms that lack clear architectural organization, B/T cell compartmentalization, or GC activity.37,41 Consequently, indiscriminately categorizing unstructured lymphoid aggregates as functional iTLSs may obscure the true biological significance of mature iTLSs and weaken their association with clinical outcomes, including tumor recurrence and patient survival.
A second controversy concerns how iTLSs are assessed. In early study, Hematoxylin and Eosin (H&E) staining has been the foundational method for diagnosing iTLSs, as they share some histological features with lymph nodes.42 In routine practice, it is used to identify and assess their basic morphology. However, H&E staining has inherent limitations, primarily its inability to accurately differentiate cellular subsets and functional structures within iTLSs.43 Challenging this limitation, a novel deep-learning approach utilizing H&E-stained slides demonstrated the ability to automatically segment and quantify the “iTLSs ratio”, which effectively predicts cancer prognosis and immunotherapy response across multiple cohorts. By leveraging matched multiplex immunohistochemistry (mIHC) images as ground truth, these AI-driven approaches reduce subjective human judgment. However, their clinical implementation faces several limitations, including the current inability to computationally separate mature germinal center-containing iTLSs from immature aggregates to refine predictive value, the challenge of using 2D histological sections to represent complex 3D immune structures, and potential vulnerability to domain shift across different scanning platforms44. To achieve a more precise characterization of iTLSs, such as identifying B/T cell zones, germinal centers, FDC networks, and specific immune cell population-IHC or multiplex immunofluorescence are essential complementary techniques34,45,46 (Table 1). However, there is no current consensus agreement on the definition of iTLSs, which limits the diagnostic and prognostic value. Developing a unified system for iTLSs identification, classification, and scoring, combining H&E and IHC, is essential for broader clinical application.
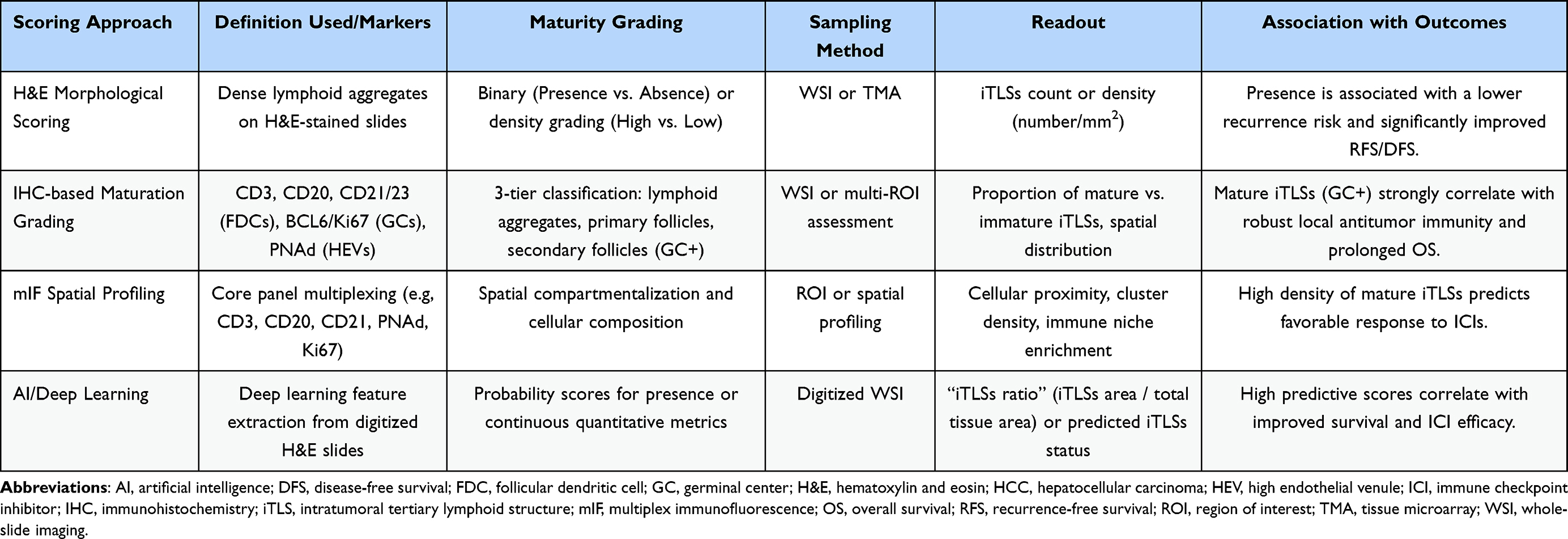 |
Table 1 Existing Histopathological iTLSs Scoring Approaches in HCC |
Non-Invasive Prediction of iTLSs in HCC
Accumulating evidence has increasingly underscored the clinical relevance of non-invasive, preoperative prediction of iTLSs in HCC, given their close association with tumor immune contexture, prognosis, and therapeutic responsiveness. At present, assessment of iTLSs relies largely on postoperative pathological evaluation, which is invasive and temporally delayed. This limitation restricts the use of iTLSs-related information for preoperative treatment stratification and may be influenced by sampling bias, thereby failing to capture intratumoral immune heterogeneity. Consequently, there is growing interest in imaging-based non-invasive approaches that enable a virtual biopsy of the tumor immune microenvironment.
Early studies using contrast-enhanced CT demonstrated the feasibility of preoperative, non-invasive prediction of iTLSs in HCC through nomogram- or machine learning–based frameworks. In a CT-based nomogram study, imaging-derived features were combined to enable preoperative discrimination of tumors with and without iTLSs, providing initial proof of concept for CT-driven iTLSs assessment.47 Subsequent machine learning-based CT radiomics studies further advanced this approach by integrating high-dimensional radiomic features extracted from multiphase CT images with clinical variables. Notably, one study identified the albumin-bilirubin score as an independent clinical predictor of iTLSs expression, and demonstrated that a combined clinical–radiomics model achieved superior predictive performance compared with clinical-only or radiomics-only models, with reported area under the curve values exceeding 0.90 in both training and validation cohorts.48 In addition, a multicenter study using multi-phase CT-based intratumoral and peritumoral radiomics confirmed the robustness of CT radiomic signatures for iTLSs prediction across independent cohorts, reporting good discriminative performance in both internal and external validation sets.49
Subsequent advances using MRI-based approaches further improved predictive performance and biological interpretability for non-invasive assessment of iTLSs. Several studies demonstrated that conventional MRI-derived features were associated with iTLSs status or maturity. In particular, MRI-based models identified imaging features such as intratumoral hemorrhage, tumor-in-vein, complete capsule, satellite nodules, and intratumoral arterial features as significant predictors when stratifying different iTLSs maturity stages. These feature-based models achieved moderate discriminative performance, with reported area under the curve values of approximately 0.75 and 0.76 for distinguishing adjacent iTLSs maturity categories, and maintained consistent performance in external validation cohorts.36
Beyond conventional imaging features, advanced MRI-based radiomic and transfer learning models substantially enhanced predictive accuracy. A multicenter transfer learning radiomic study based on contrast-enhanced MRI achieved excellent discrimination of iTLSs presence, with reported AUC values of 0.91 in the training cohort and approximately 0.85 in both internal and external validation cohorts, and further demonstrated associations between model-predicted iTLSs status and improved survival outcomes.50 In parallel, hybrid models integrating MRI radiomic signatures with clinical–radiologic variables achieved AUC values ranging from 0.82 to 0.88 across training and validation cohorts, and were shown to identify patient subgroups that derived survival benefit from postoperative targeted-immunotherapy.51
More recently, susceptibility-weighted imaging (SWI) has emerged as a complementary MRI technique for iTLSs prediction. An interpretable SWI-based radiomics model reported an AUC of 0.77 in an independent validation cohort, and identified specific radiomic features as significant predictors of iTLSs-positivity.52 In addition, diffusion-based MRI techniques, including intravoxel incoherent motion (IVIM) imaging, demonstrated added value, with radiomics-based IVIM models achieving an AUC of approximately 0.80, outperforming models based solely on IVIM parameters or clinical–radiologic features.53
While current non-invasive imaging models demonstrate high predictive accuracy, their clinical translation faces significant hurdles. A primary challenge is the “black box” nature of radiomics, where high-dimensional mathematical features often lack direct biological interpretability.54 Consequently, it remains difficult to definitively determine whether imaging-predicted iTLSs truly represent functionally active anti-tumor immune niches or merely reflect non-specific stromal alterations. Moreover, the heavy reliance on high-dimensional data extraction, coupled with heterogeneous imaging protocols, inherently increases the risk of mathematical overfitting and center-specific bias.55 Specifically, the frequently reported high AUCs must be carefully contextualized, as they are often derived from retrospective designs and can be heavily influenced by the pathological ground truth used for training—such as the inherent differences between comprehensive whole-slide imaging (WSI) and sampling-limited biopsy specimens. A structured summary of key imaging studies and their specific limitations is provided in Table 2.
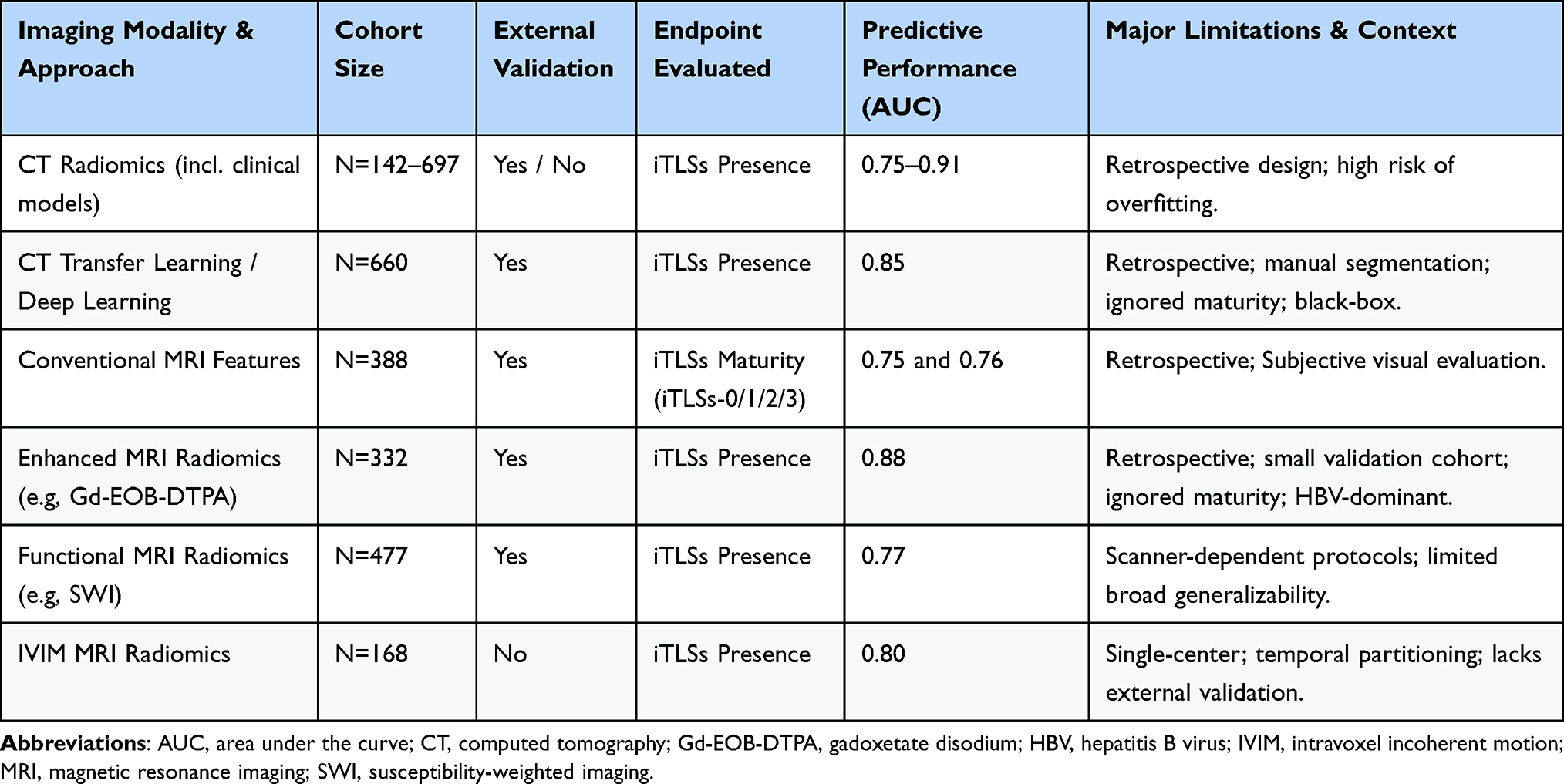 |
Table 2 Summary of Key Non-Invasive Imaging Studies for iTLSs Prediction in HCC |
iTLSs and Clinical Outcomes in HCC
Prognostic Association: Recurrence and Overall Survival
Accumulating evidence suggests that iTLSs in HCC are generally associated with a more favorable immune contexture and improved clinical outcomes, although their prognostic and therapeutic significance is heterogeneous and context dependent. Multiple studies have reported that the presence of iTLSs correlates with lower recurrence risk and improved survival, particularly in early-stage HCC,11,13,14 and is accompanied by increased infiltration of cytotoxic T cells, B cells, and other effector immune populations.56,57 Current evidence suggests that the prognostic association is particularly robust for recurrence-related outcomes, such as recurrence-free survival (RFS) or disease-free survival (DFS), following curative treatments like surgical resection.13 This strong correlation is biologically plausible, as the localized, primed immune microenvironment orchestrated by iTLSs actively suppresses micrometastases and delays local recurrence. While mature iTLSs are also generally associated with improved overall survival (OS), this relationship can sometimes appear less pronounced across studies. This is because OS in HCC is heavily confounded by competing risks over the long term, such as the progression of underlying liver cirrhosis, hepatic failure, or the development of de novo tumors.58,59 Therefore, explicitly distinguishing these endpoints is essential for accurately interpreting the clinical impact of iTLSs.
At the cellular level, single-cell transcriptomic analyses indicate that iTLSs-associated regions are enriched in memory T-cell subsets and organized immune niches,29,60 supporting the concept that iTLSs function as local platforms for sustained antitumor immune responses. However, emerging data emphasize that treating iTLSs as a simple binary variable (presence versus absence) risks conflating biologically distinct entities. The functional quality and maturation status of iTLSs critically determine their clinical relevance.57,61 Mature iTLSs, characterized by active germinal centers and robust FDC networks, are consistently associated with robust antitumor immunity and superior survival. In contrast, immature lymphoid aggregates often fail to mount effective humoral responses and may even harbor immunosuppressive cells.28 Consequently, the failure of earlier studies to clearly separate immature aggregates from germinal center-containing mature structures likely accounts for much of the inter-study variability regarding the prognostic value of iTLSs. Investigations focusing on spatial organization demonstrate that the presence of mature iTLSs, rather than mere immature lymphoid aggregates, is strongly associated with favorable patient outcomes.29,62 In addition, the prognostic value of iTLSs appears context dependent and is influenced by other tumor microenvironmental features. Evidence from studies that jointly evaluate iTLSs with microvascular invasion or incorporate neutrophil-related states, including N1/N2 heterogeneity and systemic neutrophil-to-lymphocyte ratio, shows that combined models can provide more granular prognostic stratification than iTLSs measures alone.8,63,64
Predictive Value for Immunotherapy Efficacy
From a therapeutic perspective, recent clinical evidence indicates that iTLSs-positive tumors exhibit enhanced responsiveness to adjuvant PD-1 inhibitor therapy following hepatectomy, with distinct immune cell profiles observed in iTLSs-rich tumors, suggesting a potential role for iTLSs as biomarkers of immunotherapy benefit.
Integrating iTLSs patterns with HCC molecular subtypes offers crucial translational insights. The WNT/β-catenin-activated phenotype represents a classic “immune-excluded” class, where pathway activation suppresses immune cell infiltration.65,66 This environment presumably hinders iTLSs neogenesis. Consequently, the absence of iTLSs may serve as a morphological surrogate for this immune-excluded subtype, identifying patients likely resistant to immune checkpoint inhibitors. Conversely, iTLSs-rich tumors likely align with the “immune-inflamed” subclass, suggesting better responsiveness to immunotherapy.
Furthermore, it is increasingly recognized that the functional variability of iTLSs is closely tied to the etiologic heterogeneity of HCC. iTLSs in virus-associated HCC (such as HBV or HCV infection) are generally linked to the classical activation of robust anti-tumor T-cell and B-cell responses, the continuous presence of viral antigens often provides strong immunogenic stimuli that support the maturation of iTLSs, correlating with favorable prognosis and enhanced responsiveness to immunotherapy. Metabolic etiologies present a fundamentally distinct immunopathology. Emerging data indicate that in non-alcoholic steatohepatitis (NASH)-associated HCC, the unique chronic inflammatory milieu can drive the formation of dysfunctional iTLSs. In these specific metabolic contexts, iTLSs may paradoxically entrap immunosuppressive IgA+ plasma cells or regulatory B cells, thereby dismantling immune surveillance and potentially blunting the efficacy of immunotherapies.67,68 Although literature specifically addressing iTLSs in toxin- and alcohol-related HCC remains scarce, non-viral insults such as chronic alcohol consumption are known to drive distinct immunosuppressive pathways. Mechanistically, alcohol exposure induces severe oxidative stress and promotes the intrahepatic accumulation of immunosuppressive MDSCs, profoundly suppressing T-cell recruitment and impeding effective adaptive immunity.69 Consequently, this highly compromised microenvironment hinders the de novo formation of mature iTLSs, leading to a poorer clinical prognosis and reduced responsiveness to immune checkpoint inhibitors compared to viral HCCs. Therefore, incorporating the underlying liver disease context is no longer optional but essential for accurately predicting the clinical trajectory and immunotherapy response based on iTLSs status.
Despite these strong prognostic associations, the exact causal relationship between iTLSs and clinical outcomes remains insufficiently addressed. A critical question is whether iTLSs are active drivers of antitumor immunity, or merely epiphenomenal biomarkers of a pre-existing, permissive immune microenvironment.39,70 Most current evidence in HCC relies on retrospective, observational studies, which establish correlation rather than direct causation. As highlighted in recent literature, it is important to realize that part of the observed prognostic value of iTLSs is likely explained by the fact that their formation indicates the presence of an ongoing immune response, rather than acting as the sole central driver.39 While the highly organized spatial architecture of mature iTLSs strongly implies an active role in educating tumor-reactive lymphocytes, the conflicting data from NASH-related HCC introduces significant complexity. In metabolic contexts, the aberrant formation of immunosuppressive iTLSs suggests they might sometimes merely reflect the underlying dysfunctional inflammatory milieu rather than orchestrating an effective defense. Therefore, while iTLSs are undoubtedly robust biomarkers, definitive proof of their causal role as indispensable drivers of anti-HCC immunity requires further functional validation using sophisticated in vivo models and prospective trials.
Key Confounders and Modifiers
The clinical utility and prognostic interpretation of iTLSs are significantly modulated by several key confounders reported in the current literature. First, underlying etiology fundamentally alters iTLSs functionality; while viral (HBV/HCV) HCCs typically harbor immune-active iTLSs, metabolic (NASH) backgrounds can drive paradoxically immunosuppressive configurations.67,68 Second, tumor stage and tissue sampling introduce critical variability. Given the high spatial heterogeneity of the tumor microenvironment, limited core-needle biopsies are highly susceptible to sampling errors compared to whole-tumor resections, often failing to accurately capture the true density and maturity of iTLSs.60 Third, prior locoregional therapies introduce significant variability depending on their specific modalities. Ischemia-inducing treatments (such as TACE or ablation) cause severe hypoxia and necrosis, which can physically disrupt the tumor environment. Conversely, non-embolizing treatments like hepatic arterial infusion chemotherapy (HAIC) can trigger immunogenic cell death and potentially induce de novo iTLSs formation.71 This modality-dependent remodeling of the immune microenvironment heavily confounds baseline iTLSs assessment. Finally, reducing iTLSs to a binary presence rather than evaluating functional germinal centers remains a major confounder limiting cross-cohort comparability.
Challenges and Future Perspectives
Future research should increasingly move beyond passive observation of iTLSs and toward therapeutic strategies that actively modulate iTLSs function and spatial distribution to optimize personalized immunotherapy for HCC. Despite the promising prognostic value of iTLSs in HCC, their clinical translation is currently impeded by critical challenges regarding functional heterogeneity, standardization, and safe induction. For instance, the lack of standardized scoring systems and biopsy sampling errors underscore the urgent need to validate non-invasive, AI-driven radiomic signatures for dynamic iTLSs assessment. Although recent advances in AI-driven radiomic models offer a promising non-invasive alternative, it must be explicitly acknowledged that the majority of current imaging-based models are retrospective in nature and often lack robust external validation. Furthermore, their clinical implementation is limited by variability in imaging protocols, scanner platforms, and tumor segmentation strategies across centers.
Overcoming these translational hurdles may require large-scale, prospective, multicenter studies with standardized imaging acquisitions to rigorously validate these radiomic signatures before they can be reliably integrated into routine clinical workflows. To advance this field beyond merely optimizing mathematical performance metrics, a standardization roadmap integrating pathological scoring, non-invasive imaging surrogates, and functional molecular signatures is essential. Furthermore, prospective study designs must be optimized: early-stage cohorts should directly link pre-operative imaging with whole-mount resection pathology mapping and clinical outcomes, while advanced cases require combining limited biopsies with mIHC and spatially resolved multi-omics to decode complex immune phenotypes and overcome sampling bias. In this regard, advanced single-cell and spatial RNA technologies, which have recently provided profound insights into the complex inflammatory mosaic of other digestive tract disorders such as inflammatory bowel disease,72 should be increasingly leveraged to decode the spatiotemporal heterogeneity within HCC-associated iTLSs.
Moreover, most existing imaging approaches are trained to predict pathologically defined iTLSs presence or maturation, rather than distinguishing functionally active from dysfunctional immune niches, underscoring the necessity of integrating imaging with immunophenotypic or molecular profiling. Future strategies must also navigate the distinct immunopathologies of viral versus metabolic etiologies to develop etiology-specific induction therapies (eg, HAIC or STING agonists) that promote mature iTLSs formation,12 while avoiding autoimmunity or vascular destruction, and inspired by successful lipid metabolism models in other cancers,73 integrating metabolic profiling with iTLSs transcriptomic signatures could significantly refine risk stratification, particularly for metabolic-associated HCC. However, safely translating these “iTLSs-inducing” approaches requires stringent safety evaluations. Actively hyper-stimulating ectopic lymphoid neogenesis carries significant inherent risks, including the triggering of severe off-target autoimmunity, excessive local inflammatory cascades, and severe vascular injury,74,75 thereby necessitating highly targeted, tumor-restricted delivery systems.
Conclusion
iTLSs appear to function as important immune hubs and are strongly associated with favorable antitumor response in HCC. Moving forward, standardizing the pathological assessment of iTLSs maturation and leveraging advanced, non-invasive radiomic imaging may significantly refine patient risk stratification. Ultimately, translating these biological and radiological insights into routine clinical workflows holds great promise for optimizing patient selection for immunotherapy and driving the development of personalized, etiology-specific therapeutic interventions for HCC.
Ethics Approval and Consent to Participate
Although this article is a narrative review, Figure 2 contains representative histological images from our patient cohort. Their use was approved by the Ethics Committee of Army Medical University (No. KY2023107), and informed consent was waived.
Consent for Publication
All authors approved the final manuscript.
Funding
This work was supported by the Program of the AI Medical Device Innovation Tasks: Unveiling the List and Taking the Lead (2025-AISJK-01008 to Wei Chen).
Disclosure
The authors declare no competing interests in this work.
References
1. Li X, Yao Q, Liu C, et al. Macrotrabecular-massive hepatocellular carcinoma: what should we know? J Hepatocell Carcinoma. 2022;9:379–13. doi:10.2147/jhc.S364742
2. Tsuge M, Kawaoka T, Oka S. Factors for the recurrence of hepatocellular carcinoma after hepatic resection. J Gastroenterol. 2023;58(3):292–293. doi:10.1007/s00535-023-01962-3
3. Finn RS, Qin S, Ikeda M, et al. Atezolizumab plus bevacizumab in unresectable hepatocellular carcinoma. New Engl J Med. 2020;382(20):1894–1905. doi:10.1056/NEJMoa1915745
4. Kelley RK, Rimassa L, Cheng AL, et al. Cabozantinib plus atezolizumab versus sorafenib for advanced hepatocellular carcinoma (COSMIC-312): a multicentre, open-label, randomised, Phase 3 trial. Lancet Oncol. 2022;23(8):995–1008. doi:10.1016/s1470-2045(22)00326-6
5. Qin S, Chen M, Cheng AL, et al. Atezolizumab plus bevacizumab versus active surveillance in patients with resected or ablated high-risk hepatocellular carcinoma (IMbrave050): a randomised, open-label, multicentre, phase 3 trial. Lancet. 2023;402(10415):1835–1847. doi:10.1016/s0140-6736(23)01796-8
6. Xu W, Lu J, Liu WR, et al. Heterogeneity in tertiary lymphoid structures predicts distinct prognosis and immune microenvironment characterizations of clear cell renal cell carcinoma. J ImmunoTher Cancer. 2023;11(12):e006667. doi:10.1136/jitc-2023-006667
7. Li H, Liu H, Fu H, et al. Peritumoral tertiary lymphoid structures correlate with protective immunity and improved prognosis in patients with hepatocellular carcinoma. Front Immunol. 2021;12:648812. doi:10.3389/fimmu.2021.648812
8. Wen S, Chen Y, Hu C, et al. Combination of tertiary lymphoid structure and neutrophil-to-lymphocyte ratio predicts survival in patients with hepatocellular carcinoma. Front Immunol. 2021;12:788640. doi:10.3389/fimmu.2021.788640
9. Zhang T, Lei X, Jia W, et al. Peritumor tertiary lymphoid structures are associated with infiltrating neutrophils and inferior prognosis in hepatocellular carcinoma. Cancer Med. 2023;12(3):3068–3078. doi:10.1002/cam4.5227
10. Hu L, Li X, Yang C, et al. Prognostic value of tertiary lymphoid structures in hepatocellular carcinoma: a meta-analysis and systematic review. Front Immunol. 2024;15:1390938. doi:10.3389/fimmu.2024.1390938
11. Li J, Zhang L, Xing H, et al. The absence of intra-tumoral tertiary lymphoid structures is associated with a worse prognosis and mTOR signaling activation in hepatocellular carcinoma with liver transplantation: a multicenter retrospective study. Adv Sci. 2024;11(21):e2309348. doi:10.1002/advs.202309348
12. Shu DH, Ho WJ, Kagohara LT, et al. Immunotherapy response induces divergent tertiary lymphoid structure morphologies in hepatocellular carcinoma. Nat Immunol. 2024;25(11):2110–2123. doi:10.1038/s41590-024-01992-w
13. Calderaro J, Petitprez F, Becht E, et al. Intra-tumoral tertiary lymphoid structures are associated with a low risk of early recurrence of hepatocellular carcinoma. J Hepatol. 2019;70(1):58–65. doi:10.1016/j.jhep.2018.09.003
14. Li H, Wang J, Liu H, et al. Existence of intratumoral tertiary lymphoid structures is associated with immune cells infiltration and predicts better prognosis in early-stage hepatocellular carcinoma. Aging. 2020;12(4):3451–3472. doi:10.18632/aging.102821
15. Khanal S, Wieland A, Gunderson AJ. Mechanisms of tertiary lymphoid structure formation: cooperation between inflammation and antigenicity. Front Immunol. 2023;14:1267654. doi:10.3389/fimmu.2023.1267654
16. Teillaud JL, Houel A, Panouillot M, et al. Tertiary lymphoid structures in anticancer immunity. Nat Rev Cancer. 2024;24(9):629–646. doi:10.1038/s41568-024-00728-0
17. Rodriguez AB, Peske JD, Woods AN, et al. Immune mechanisms orchestrate tertiary lymphoid structures in tumors via cancer-associated fibroblasts. Cell Rep. 2021;36(3):109422. doi:10.1016/j.celrep.2021.109422
18. Engelhard VH, Rodriguez AB, Mauldin IS, et al. Immune cell infiltration and tertiary lymphoid structures as determinants of antitumor immunity. J Immunol. 2018;200(2):432–442. doi:10.4049/jimmunol.1701269
19. Roozendaal R, Mebius RE. Stromal cell-immune cell interactions. Ann Rev Immunol. 2011;29:23–43. doi:10.1146/annurev-immunol-031210-101357
20. Zhang Q, He Y, Luo N, et al. Landscape and dynamics of single immune cells in hepatocellular carcinoma. Cell. 2019;179(4):829–45.e20. doi:10.1016/j.cell.2019.10.003
21. Affo S, Yu LX, Schwabe RF. The role of cancer-associated fibroblasts and fibrosis in liver cancer. Ann Rev Pathol. 2017;12:153–186. doi:10.1146/annurev-pathol-052016-100322
22. Allen E, Jabouille A, Rivera LB, et al. Combined antiangiogenic and anti-PD-L1 therapy stimulates tumor immunity through HEV formation. Sci Trans Med. 2017;9(385). doi:10.1126/scitranslmed.aak9679
23. Fridman WH, Meylan M, Pupier G, et al. Tertiary lymphoid structures and B cells: an intratumoral immunity cycle. Immunity. 2023;56(10):2254–2269. doi:10.1016/j.immuni.2023.08.009
24. Fleige H, Ravens S, Moschovakis GL, et al. IL-17-induced CXCL12 recruits B cells and induces follicle formation in BALT in the absence of differentiated FDCs. J Exp Med. 2014;211(4):643–651. doi:10.1084/jem.20131737
25. Luther SA, Lopez T, Bai W, et al. BLC expression in pancreatic islets causes B cell recruitment and lymphotoxin-dependent lymphoid neogenesis. Immunity. 2000;12(5):471–481. doi:10.1016/s1074-7613(00)80199-5
26. Vella G, Hua Y, Bergers G. High endothelial venules in cancer: regulation, function, and therapeutic implication. Cancer Cell. 2023;41(3):527–545. doi:10.1016/j.ccell.2023.02.002
27. Bugatti M, Bergamini M, Missale F, et al. A Population of TIM4+FOLR2+ Macrophages Localized in Tertiary Lymphoid Structures Correlates to an Active Immune Infiltrate Across Several Cancer Types. Cancer Immunol Res. 2022;10(11):1340–1353. doi:10.1158/2326-6066.Cir-22-0271
28. Joshi NS, Akama-Garren EH, Lu Y, et al. Regulatory T cells in tumor-associated tertiary lymphoid structures suppress anti-tumor T cell responses. Immunity. 2015;43(3):579–590. doi:10.1016/j.immuni.2015.08.006
29. Tang Z, Bai Y, Fang Q, et al. Spatial transcriptomics reveals tryptophan metabolism restricting maturation of intratumoral tertiary lymphoid structures. Cancer Cell. 2025;43(6):1025–1044.e14. doi:10.1016/j.ccell.2025.03.011
30. Yang L, Li A, Wang Y, et al. Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy. Signal Transduct Target Ther. 2023;8(1):35. doi:10.1038/s41392-022-01304-4
31. Overacre-Delgoffe AE, Bumgarner HJ, Cillo AR, et al. Microbiota-specific T follicular helper cells drive tertiary lymphoid structures and anti-tumor immunity against colorectal cancer. Immunity. 2021;54(12):2812–24.e4. doi:10.1016/j.immuni.2021.11.003
32. Ma C, Han M, Heinrich B, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018;360(6391). doi:10.1126/science.aan5931
33. Qu S, Jia W, Liu X, et al. Intratumoral bacterial load and tertiary lymphoid structure density in hepatocellular carcinoma: association and prognostic significance. Front Immunol. 2025;16:1652433. doi:10.3389/fimmu.2025.1652433
34. Vanhersecke L, Bougouin A, Crombé A, et al. Standardized pathology screening of mature tertiary lymphoid structures in cancers. Lab Invest. 2023;103(5):100063. doi:10.1016/j.labinv.2023.100063
35. Kinker GS, Vitiello GAF, Diniz AB, et al. Mature tertiary lymphoid structures are key niches of tumour-specific immune responses in pancreatic ductal adenocarcinomas. Gut. 2023;72(10):1927–1941. doi:10.1136/gutjnl-2022-328697
36. Li Y, Chen Y, Wu Z, et al. Noninvasive MRI imaging feature-based prediction of intratumoral tertiary lymphoid structure maturity in hepatocellular carcinoma: a multicenter retrospective study. Eur Radiol. 2025. doi:10.1007/s00330-025-11902-9
37. Le Rochais M, Hemon P, Ben-Guigui D, et al. Deciphering the maturation of tertiary lymphoid structures in cancer and inflammatory diseases of the digestive tract using imaging mass cytometry. Front Immunol. 2023;14:1147480. doi:10.3389/fimmu.2023.1147480
38. Li H, Ding JY, Zhang MJ, et al. Tertiary lymphoid structures and cytokines interconnections: the implication in cancer immunotherapy. Cancer Lett. 2023;568:216293. doi:10.1016/j.canlet.2023.216293
39. Schumacher TN, Thommen DS. Tertiary lymphoid structures in cancer. Science. 2022;375(6576):eabf9419. doi:10.1126/science.abf9419
40. Zhang X, Yao J, Xie M, et al. Tertiary lymphoid structures as potential biomarkers for cancer prediction and prognosis. Int Immunopharmacol. 2024;140:112790. doi:10.1016/j.intimp.2024.112790
41. Dieudé M, Kaci I, Hébert M-J. The impact of programmed cell death on the formation of tertiary lymphoid structures. Front Immunol. 2021;12:696311. doi:10.3389/fimmu.2021.696311
42. Sautès-Fridman C, Lawand M, Giraldo NA, et al. Tertiary lymphoid structures in cancers: prognostic value, regulation, and manipulation for therapeutic intervention. Front Immunol. 2016;7:407. doi:10.3389/fimmu.2016.00407
43. Deng S, Chen Y, Song B, et al. Tertiary lymphoid structures in cancer: spatiotemporal heterogeneity, immune orchestration, and translational opportunities. J hematol oncol. 2025;18(1):97. doi:10.1186/s13045-025-01754-7
44. Chen Z, Wang X, Jin Z, et al. Deep learning on tertiary lymphoid structures in hematoxylin-eosin predicts cancer prognosis and immunotherapy response. NPJ Precis Oncol. 2024;8(1):73. doi:10.1038/s41698-024-00579-w
45. Su X, Kang D, Wang J, et al. Tertiary lymphoid structures associated with improved survival and enhanced antitumor immunity in acral melanoma. NPJ Precis Oncol. 2025;9(1):103. doi:10.1038/s41698-025-00891-z
46. Rakaee M, Kilvaer TK, Jamaly S, et al. Tertiary lymphoid structure score: a promising approach to refine the TNM staging in resected non-small cell lung cancer. Br J Cancer. 2021;124(10):1680–1689. doi:10.1038/s41416-021-01307-y
47. Li P, Liang Y, Zeng B, et al. Preoperative prediction of intra-tumoral tertiary lymphoid structures based on CT in hepatocellular cancer. Eur J Radiol. 2022;151:110309. doi:10.1016/j.ejrad.2022.110309
48. Wu J, Zuo Z, Na L, et al. Machine learning-driven prediction of intratumoral tertiary lymphoid structures in hepatocellular carcinoma using contrast-enhanced CT imaging and integrated clinical data. Front Oncol. 2025;15:1652509. doi:10.3389/fonc.2025.1652509
49. Wang R, Li Y, Zhang X, et al. Multi-phase CT-based intratumoral and peritumoral radiomics for predicting tertiary lymphoid structures of hepatocellular carcinoma: a multi-center retrospective cohort study. Eur J Surg Oncol. 2025;52(2):111341. doi:10.1016/j.ejso.2025.111341
50. Long S, Li M, Chen J, et al. Transfer learning radiomic model predicts intratumoral tertiary lymphoid structures in hepatocellular carcinoma: a multicenter study. J ImmunoTher Cancer. 2025;13(3):e011126. doi:10.1136/jitc-2024-011126
51. Li Y, Li X, Xiao X, et al. A novel hybrid model for predicting tertiary lymphoid structures and targeted immunotherapy outcomes in hepatocellular carcinoma: a multicenter retrospective study. Eur Radiol. 2024;35(6):3206–3222. doi:10.1007/s00330-024-11255-9
52. Liu L, Gao F, Li Y, et al. An interpretable radiomics-based model using susceptibility-weighted imaging for non-invasive prediction of tertiary lymphoid structures in hepatocellular carcinoma. J Hepatocell Carcinoma. 2025;12:2197–2211. doi:10.2147/jhc.S551462
53. Ma L, Liao S, Zhang X, et al. Application of intravoxel incoherent motion in the prediction of intra-tumoral tertiary lymphoid structures in hepatocellular carcinoma. J Hepatocell Carcinoma. 2025;12:383–398. doi:10.2147/jhc.S508357
54. Tomaszewski MR, Gillies RJ. The biological meaning of radiomic features. Radiology. 2021;298(3):505–516. doi:10.1148/radiol.2021202553
55. Lambin P, Leijenaar RTH, Deist TM, et al. Radiomics: the bridge between medical imaging and personalized medicine. Nat Rev Clin Oncol. 2017;14(12):749–762. doi:10.1038/nrclinonc.2017.141
56. Su JY, Li JR, Pan LX, et al. Tertiary lymphoid structures in HCC: influence on immune cell profiles in tumors and on efficacy of adjuvant PD-1 inhibitor therapy after hepatectomy. Hepatology. 2025. doi:10.1097/hep.0000000000001433
57. Li J, Xu H, Han J, et al. Lymphocyte function in tertiary lymphoid structures predicts hepatocellular carcinoma outcome. Lab Invest. 2024;104(11):102144. doi:10.1016/j.labinv.2024.102144
58. Villanueva A. Hepatocellular Carcinoma. New Engl J Med. 2019;380(15):1450–1462. doi:10.1056/NEJMra1713263
59. Czauderna C, Schmidtmann I, Koch S, et al. High pretreatment static and dynamic alpha-fetoprotein values predict reduced overall survival in hepatocellular carcinoma. United European Gastroenterol J. 2021;9(3):388–397. doi:10.1177/2050640620972611
60. Lu Y, Yang A, Quan C, et al. A single-cell atlas of the multicellular ecosystem of primary and metastatic hepatocellular carcinoma. Nat Commun. 2022;13(1):4594. doi:10.1038/s41467-022-32283-3
61. Munoz-Erazo L, Rhodes JL, Marion VC, et al. Tertiary lymphoid structures in cancer - considerations for patient prognosis. Cell Mol Immunol. 2020;17(6):570–575. doi:10.1038/s41423-020-0457-0
62. Kurebayashi Y, Sugimoto K, Tsujikawa H, et al. Spatial dynamics of T- and B-cell responses predicts clinical outcome of resectable and unresectable hepatocellular carcinoma. Clin Cancer Res. 2024;30(24):5666–5680. doi:10.1158/1078-0432.Ccr-24-0479
63. Ma L, Liao S, Yuan S, et al. Refining risk stratification in hepatocellular carcinoma by integrating tertiary lymphoid structures and microvascular invasion: a multicenter retrospective study. Int J Surg. 2025;111(11):8212–8225. doi:10.1097/js9.0000000000003045
64. Lang Y, Fu W, Xu W, et al. Prognostic value of N1/N2 neutrophils heterogeneity and tertiary lymphoid structure in hepatocellular carcinoma patients. Cancer Med. 2024;13(24):e70551. doi:10.1002/cam4.70551
65. Aoki T, Nishida N, Kurebayashi Y, et al. Two distinct characteristics of immune microenvironment in human hepatocellular carcinoma with Wnt/β-catenin mutations. Liver Cancer. 2024;13(3):285–305. doi:10.1159/000533818
66. Harding JJ, Nandakumar S, Armenia J, et al. Prospective genotyping of hepatocellular carcinoma: clinical implications of next-generation sequencing for matching patients to targeted and immune therapies. J Clin Cancer Res. 2019;25(7):2116–2126. doi:10.1158/1078-0432.CCR-18-2293
67. Shalapour S, Lin X-J, Bastian IN, et al. Inflammation-induced IgA+ cells dismantle anti-liver cancer immunity. Nature. 2017;551(7680):340–345. doi:10.1038/nature24302
68. Pfister D, Núñez NG, Pinyol R, et al. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature. 2021;592(7854):450–456. doi:10.1038/s41586-021-03362-0
69. Llovet JM, Castet F, Heikenwalder M, et al. Immunotherapies for hepatocellular carcinoma. Nat Rev Clin Oncol. 2022;19(3):151–172. doi:10.1038/s41571-021-00573-2
70. Colbeck EJ, Ager A, Gallimore A, et al. Tertiary lymphoid structures in cancer: drivers of antitumor immunity, immunosuppression, or bystander sentinels in disease? Front Immunol. 2017;8:1830. doi:10.3389/fimmu.2017.01830
71. Xing R, Mei J, Zuo Z, et al. Enhanced formation of tertiary lymphoid structures shapes the anti-tumor microenvironment in hepatocellular carcinoma after FOLFOX-HAIC therapy. Cell Rep Med. 2025;6(9):102298. doi:10.1016/j.xcrm.2025.102298
72. Liu L, Davidorf B, Dong P, et al. Decoding the mosaic of inflammatory bowel disease: illuminating insights with single-cell RNA technology. Comput Struct Biotechnol J. 2024;23:2911–2923. doi:10.1016/j.csbj.2024.07.011
73. Miao X, Wang H, Fan C, et al. Enhancing prognostic accuracy in head and neck squamous cell carcinoma chemotherapy via a lipid metabolism-related clustered polygenic model. Can Cell Inter. 2023;23(1):164. doi:10.1186/s12935-023-03014-5
74. Pitzalis C, Jones GW, Bombardieri M, et al. Ectopic lymphoid-like structures in infection, cancer and autoimmunity. Nat Rev Immunol. 2014;14(7):447–462. doi:10.1038/nri3700
75. Johansson-Percival A, He B, Li Z-J, et al. De novo induction of intratumoral lymphoid structures and vessel normalization enhances immunotherapy in resistant tumors. Nat Immunol. 2017;18(11):1207–1217. doi:10.1038/ni.3836
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.