Back to Journals » OncoTargets and Therapy » Volume 16
Interleukin-33 as a Potential Therapeutic Target in Gastric Cancer Patients: Current Insights
Authors Chatterjee A, Azevedo-Martins JM, Stachler MD ![]()
Received 1 April 2023
Accepted for publication 6 August 2023
Published 10 August 2023 Volume 2023:16 Pages 675—687
DOI https://doi.org/10.2147/OTT.S389120
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Daniel Neureiter
Annesha Chatterjee, Jordana Maria Azevedo-Martins, Matthew D Stachler
University of California San Francisco, Department of Pathology, San Francisco, CA, USA
Correspondence: Matthew D Stachler, University of California San Francisco, 513 Parnassus Ave HSW450B, San Francisco, CA, 94143, Tel +1 514 419-1902, Email [email protected]
Abstract: Gastric cancer is a significant global health problem as it is the fifth most prevalent cancer worldwide and the fourth leading cause of cancer-related mortality. While cytotoxic chemotherapy remains the primary treatment for advanced GC, response rates are limited. Recent progresses, focused on molecular signalling within gastric cancer, have ignited new hope for potential therapeutic targets that may improve survival and/or reduce the toxic effects of traditional therapies. Carcinomas are generally initiated when critical regulatory genes get mutated, but the progression to malignancy is usually supported by the non-neoplastic cells that create a conducive environment for transformation and progression to occur. Interleukin 33 (IL-33) functions as a dual activity cytokine as it is also a nuclear factor. IL-33 is usually present in the nuclei of the cells. Upon tissue damage, it is released into the extracellular space and binds to its receptor, suppression of tumorigenicity 2 (ST2) L, which is expressed on the membranes of the target cells. IL-33 signalling activates the T Helper 2 (Th2) immune response among other responses. Although the studies on the role of IL-33 in gastric cancer are still in the early stages, they have revealed potentially important (though sometimes conflicting) functions or roles in cancer development and progression. The pro-tumorigenic roles include induction and the recruitment of tumor-associated immune cells, promoting metaplasia progression, and inducing stem cell like and EMT properties in gastric cancer cells. Therapeutic interventions to disrupt these functions may provide a unique strategy for gastric cancer prevention and treatment. This review aims to provide a summary of the role of IL-33 in GC, state its multiple functions in relation to GC, and show potential avenues for promising therapeutic investigation.
Keywords: gastric cancer, interleukin-33, therapeutic target
Introduction
Gastric cancer (GC) is one of the most prevalent types of cancer worldwide. GC is fifth in incidence and fourth in mortality, and is responsible for one million newly diagnosed cases and one fatal case in every 13 cancer-related deaths.1 Unfortunately, highly effective therapy for GC is lacking, and thus new approaches toward treatment are needed. In 2005, IL-33 was first identified as a member of the IL-1 family of cytokines and as the ligand for an orphan receptor, ST2L (Suppressor of Tumorigenicity 2).2 It is constitutively expressed in a wide array of cells and tissues. IL-33 acts as a proinflammatory cytokine and expression can be induced by various external stimuli in a variety of cell types such as epithelial cells, myofibroblasts, adipocytes, endothelial cells, smooth muscle cells, and macrophages.3,4 Studies on the role of IL-33 in GC show that IL-33 expression is high in the serum and tissues of GC patient samples.5 In addition, expression of the IL-33 receptor, ST2, is upregulated in GC tissue as compared to paired adjacent normal tissue.6 This review summarises the different functions of IL-33 in GC progression in an attempt to show possible signalling areas that can be targeted for IL-33 mediated therapy.
Gastric Cancer: Overview
Classification and Risk Factors of Gastric Cancer
GC can be classified anatomically into proximal (cardia region) and distal (non-cardia regions) cancers.7 The most frequently used classification is the Lauren’s classification, which describes two main histological types: intestinal (with cells resembling intestinal mucosa or cancer) and diffuse (without intercellular junctions) type adenocarcinoma,8 each with distinct histological and molecular characteristics.9 As each region has different cells and functions, so too does gastric cancer have several morphological, cytological, and molecular types.10 The Cancer Genome Atlas Network (TCGA) classified GC into four molecular subtypes: genomically stable, chromosomal instability, microsatellite instability (MSI) and induced by Epstein-Barr Virus (EBV). This classification aimed to provide a better possibility of diagnosis and therapeutic choice,11 however, there is no classification consensus due to controversies in which one has greater clinical relevance and better therapeutic approach (Cancer Genome Atlas Research Network, 2014). This review primarily focuses on intestinal type gastric cancers which arise from areas of pre-cancerous metaplasia and are enriched for the chromosomal instable TCGA classification.
The most widely associated risk factor for the development of non-cardia GC is chronic Helicobacter pylori (H. pylori) infection.12,13 Cancers of the gastric cardia are not generally associated with H. pylori infection, and can even be inversely associated in some populations.14,15 However, GC in the cardia is associated with gastroesophageal reflux disease and obesity, with characteristics similar to esophageal adenocarcinoma.16 Importantly, through either H. pylori or reflux, both are associated with chronic inflammation.
Present Therapy and Treatment Options in Gastric Cancer
Surgery is the primary treatment option for GC when diagnosed at an early stage.17 Chemotherapy or chemoradiation can be utilized either before or after surgery to diminish the tumor (neoadjuvant therapy) before surgery or to kill any remaining cancer cells (adjuvant therapy) after surgery.18 This treatment allows stage I patients a reasonable 5-year survival as 68–80%. However, the rate of early diagnosis is low due to the lack of specific symptoms, so most patients (>70%) are diagnosed at later stages. For those advanced cases, with unresectable cancer, chemotherapy is normally the first option or a combination of chemotherapy with targeted therapy, immunotherapy, or radiation therapy.19 Unfortunately, even with treatment, the prognosis drops dramatically as the diagnosed stage increases, to 46–60% for stage II, 8–30% for stage III, and only 5% for stage IV.20–22 Therefore, new more effective therapeutic strategies are greatly needed.23–26
IL-33/ ST2 Signalling: General Overview
IL-33 is a 30-kDa protein exerting dual functions of a cytokine and a transcription factor. The N-terminal domain of IL-33 contains a nuclear localization signal, a NA-binding homeodomain-like helix-turn-helix motif, and a chromatin binding domain.27,28 The C-terminus has an IL-1 like cytokine domain.27
As a cellular damage response, tissue injury or certain viral infections can cause IL-33 to be released from the nucleus into the extracellular space where it can bind to the membrane-bound ST2L receptor through its cytokine domain.29,30 Upon binding of IL-33 to its receptor, an inflammatory cascade is initiated. Therefore, IL-33 functions as an “alarmin” and is considered to be a damage-associated molecular pattern (DAMP).31
Suppression of tumorigenicity 2 (ST2) was first identified in mouse fibroblasts as an oncogene-induced gene encoded by IL1RL1.32,33 The alternative splicing of IL1RL1 gene produces four isoforms: ST2L (ligand), sST2 (soluble form), ST2V (variant), and ST2LV (ligand variant). ST2L is the membrane embedded receptor.34,35 sST2 is secreted in the form of a glycosylated protein and is a soluble form of ST2 that lacks a transmembrane domain but has an extracellular domain along with an additional stretch of nine amino acids at the C-terminal tail.34,36 ST2V is mostly similar to sST2 except in place of the third Ig-like domain, where there is a hydrophobic tail.37,38 ST2LV is the soluble, N-glycosylated isoform without the transmembrane domain as found in ST2L.39 ST2L primarily functions as a membrane bound receptor for IL-33 which is essential for the downstream IL-33 signalling cascade.36,40 The sST2 (soluble form) functions by sequestering extracellular IL-33.
As a nuclear factor, IL-33 functions in repressing the expression of inflammatory responses by binding to chromatin. In response to cell damage, it is released into the extra-cellular space as a cytokine.41 IL-33 then binds to the ST2L receptor, via its C-terminal IL-1 like cytokine domain, which triggers a conformational change and induces the recruitment of IL-1RAcP that results in the formation of a heterodimeric receptor complex on the cell membrane.42 This brings together the intracellular domains of the two transmembrane proteins, resulting in the recruitment of adaptor molecules through which IL-33 signalling occurs. These include myeloid differentiation primary response protein 88 (MyD88), IL-1 receptor-associated kinases (IRAK-1 and IRAK-4), and tumor necrosis factor (TNF) acceptor associated factor 6 (TRAF6).42 Pathway activation causes the degradation of the inhibitory protein IκB together with subsequent activation of NF-κB transcription factor. Along with this, the activation of mitogen activated protein (MAP) kinases p38, c-Jun N-terminal kinase (JNK), and extracellular signal-regulated kinase (ERK) results in the activation of downstream transcription factors like activator protein (AP)-1.4 These transcription factors direct the expression of cell-specific cytokines and chemokines, as well as a plethora of downstream effects.4
IL-33 can interact with and influence multiple inflammatory cells, including functioning as a key regulator of Th2 responses. The IL-33/ST2 axis triggers the production of Th2 cytokines such as IL-4, IL-5, and IL-13 in Th2 cells.43 In epithelial cells, IL-33/ST2 activation influences chemokine production44 and active signalling by IL-33/ST2 induces activation of immune effector cells.4,45 Recruitment of both pro- and anti-tumorigenic cells into the TME in cancer has been reported.45 IL-33 signalling has been found to be inhibited by multiple mechanisms. The pro-inflammatory activities of IL-33 are reduced by its nuclear localization, where it remains bound to chromatin, repressing transcription and finally undergoes degradation by pro-apoptotic proteases to prevent extracellular release in apoptotic cells.
Role of IL-33 Signalling in Different Cancers
Experimental evidence indicates that the IL-33/ST2 signalling pathway plays a critical role in influencing the tumor immune microenvironment. There is evidence to support both pro-tumoral and antitumoral functions and IL-33 differentially affects inflammatory microenvironment depending on the types of tumors, the various immune cell targets, and on separate cooperating microenvironmental factors.46
IL-33 acts as a pro-tumoral agent in breast cancer where in a mouse model it was shown that IL-33/ST2 signalling creates a favourable environment for the recruitment and accumulation of various immunosuppressive myeloid cells. In addition, it also increases the expression of the pro-angiogenic marker VEGF.47–50 In human breast cancer, this signalling axis can induce the transformation of epithelial cells and induces increased breast cancer progression. It is also involved in the induction of endocrine resistance and stem cell properties in the cancer cells.51,52 The pro-tumorigenic functions of IL-33 have also been implicated in colorectal cancer. In a murine model, it has been observed that IL-33 increases polyposis, is involved in proliferation and growth of tumor, enhances angiogenesis, and supports the accumulation of immunosuppressive myeloid cells.53–58 Increased expression of IL-33 has also been associated with metastasis, recruitment of tumor associated macrophages, and promotion of stemness in colorectal cancer cells.54,55,59 Studies also show that ST2 is involved in inducing tumor migration and invasion properties in human lung cancer and that IL-33 blockage results in decreased tumor metastasis, reduced M2 macrophage polarization, and accumulation of less numbers of Tregs.60–63 Studies have implicated IL-33 and ST2 in the formation and progression of multiple other cancers as well.64–66 Together, these studies point to multiple roles of IL-33 signalling in cancer, suggesting they should also be in gastric cancer.
On the contrary, IL-33 has been proposed to function as an anti-tumor cytokine and is suppressed in certain types of cancers. In melanoma, increased IL-33 expression is associated with hindered tumor growth and decreased pulmonary metastasis through the stimulation of different effector cells such as DCs, NK Cells, CD8+T cells, ILC2s and the eosinophils.67–71 Contrary to the other studies, overexpression of IL-33 in lung cancer has been found to reduce the tumor growth in in vivo models, along with the restoration of MHC-1 expression.68,72 The IL-33 signalling cascade has also been negatively associated with tumor progression by the induction of IFN gamma.44,73,74 In leukaemia and lymphoma, studies suggest an anti-tumor effect of IL-33.75,76 Literature studies thus indicate that IL-33 has pleiotropic roles in cancer having both protumor and antitumor activities.
Role of IL-33 in Gastric Cancer
Expression of IL-33 in Gastric Cancer Patients and Role as a Potential Biomarker
Early studies on the role of IL-33 in gastric cancer revealed that IL-33 expression is high in the serum and tissues of gastric cancer patient samples.5,6 In addition, expression of the IL-33 receptor, ST2, is upregulated in gastric cancer tissues compared to paired adjacent normal tissues.77 IL-33 is secreted as an alarmin as a response to damage in the gastric epithelium and is responsible for mucosal inflammation, defence against H.pylori infection, and is a healing mechanism in the gastric cells (Figure 1). The signalling results in recruiting tumor associated like macrophages at the sites of the damage. The IL-33/ST2 axis also contributes to inflammation in the gastric tumor microenvironment.78 IL-33 is highly expressed in the circulation of gastric cancer patients and has been associated with poor prognosis, invasion, and distant metastasis. In a study by Sun et al, they showed that expression of IL-33 is upregulated in the serum samples and tissues of GC patient samples as compared to healthy individuals and it is associated with factors that determine long-term prognoses such as depth of invasion and metastasis. However, the expression of IL-33 is not correlated to the expression of the classical tumor markers CEA and CA19-9. Thus, since the serum and gastric cancer tissue levels of IL-33 expression are independent of other tumor markers, it shows potential as a biomarker for the detection of GC and may predict disease advancement and prognosis.5
 |
Figure 1 Proposed functions of IL-33 in gastric cancer. Elevated levels of IL-33 can be identified in both gastric cancer tissue samples as well as in the serum of patients with gastric cancer. Schematic showing proposed pro-tumorigenic roles of IL-33 during gastric cancer formation and progression (black boxes) and potential anti-tumorigenic role in late-stage gastric cancer (grey box). |
Involvement of IL-33 After H. Pylori Infection and in the Early Stages of Metaplasia Induction Within the Gastric Mucosa
One of the main factors responsible for gastritis and thus gastric cancer is the colonization of the stomach by the bacteria Helicobacter pylori (H.pylori). Post H.pylori infection, there is an upregulation in the expression of both IL-33 and ST2. IL-33 binds to its receptor, and this triggers the recruitment of the IL-1 receptor accessory protein (IL-1RAP) followed by activation of the IL-33 signalling pathway (Figure 2A). The expression of IL-33 may itself then be regulated by IL-1RAP. In addition to induction of IL-33/ST2 signalling, there is also increased leukocyte recruitment in the gastric mucosa after H.pylori infection. Post infection, there is a mobilization of ST2 to membrane lipid rafts to initiate IL-33/ST2 signalling.79,80 Thus, IL-33 expression, triggered upon H.pylori infection, can increase inflammation in the stomach. In a study conducted on H.pylori infected patient samples, Kuo et al showed that IL-33 expression is elevated, and it is a pivotal mediator for generating the immune response and inflammation due to the erosion or damage caused by pathogens in the mucosa. Buzzelli et al also showed that IL-33 expression is high in gastric mucosal cells. Following H.pylori infection, there is an initial increase in the expression of IL-33 along with a Th2 immune response, increase in type II innate lymphoid cells, and increased recruitment of IL-33 secreting M2 macrophages. However, with prolonged chronic H.pylori infection, the expression level of IL-33 goes down suggestive of a response shift from a Th2 to a Th1 immune response.79–81
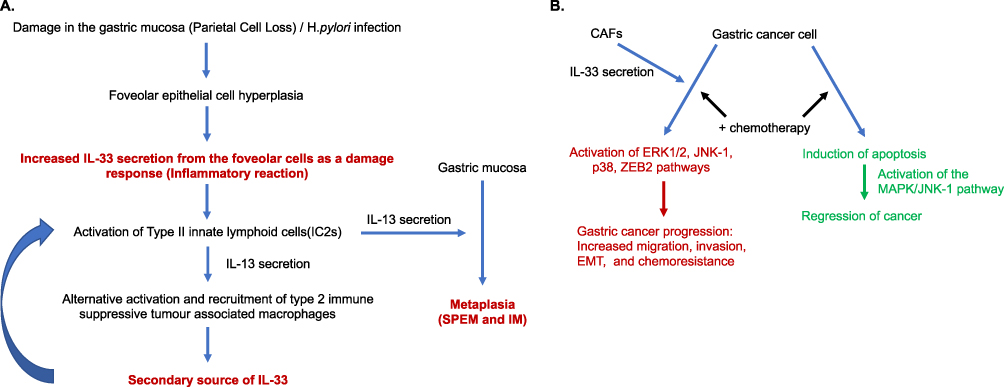 |
Figure 2 Pro-tumorigenic effects of IL-33 in early and late stages of gastric cancer progression. Opportunities may be present for therapeutic regulation of IL-33 at multiple stages of disease. (A) In the pre-cancerous stages of gastric cancer formation, damage of the gastric mucosa, such as through H.pylori infection and inflammation, results in foveolar cell hyperplasia and increased expression of IL-33 which can have a direct effect on the inflammatory background and promote metaplasia formation. (B) Potential effects of IL-33 on later stages of cancer progression and chemoresistance. IL-33 production either from CAFs or other cells increase activation of multiple pathways that can lead to cancer progression and chemoresistance. Abbreviations: SPEM Spasmolytic polypeptide-expressing metaplasia, IM intestinal metaplasia, CAF cancer associated fibroblast. |
Studies have also suggested that IL-33 is involved in some of the earlier events that leads to gastric cancer. It is thought that intestinal type gastric cancer arises out of a metaplastic transition in response to inflammation and tissue damage (for example from H. pylori infection) which can advance to dysplasia and eventually cancer. Both SPEM (Spasmolytic Polypeptide-Expressing Metaplasia) and intestinal metaplasia have been associated with intestinal type gastric cancer. It has been suggested that IL-33 is integral to the initiation and development of metaplasia.82 As a physiological damage response, studies suggest epithelial foveolar cells secrete IL-33, which stimulates the production of IL-13 from type II innate lymphoid cells (IC2s), and together this triggers the differentiation of chief cells into SPEM and results in alternative activation of macrophages. Thus, IL-33 signaling can create an immune suppressive microenvironment. These macrophages further secrete IL-33 themselves which accelerates metaplasia development.83 While this is thought of as a protective mechanism in the beginning, progression towards carcinoma can occur with a continued damage response, chronic inflammation, and cell reprogramming.82,83
In a gastric metaplasia mouse model study, Petersen et al evaluated candidate genes that are responsible for macrophage recruitment in SPEM. The macrophages isolated from the corpus region of the stomach revealed that in advanced SPEM, there was increased expression of both IL-33 and ST2. Further, using an IL-33 knockout, ST2 knockout, and IL-13 knockout mouse, it was observed that the induction of SPEM was absent in the knockout mice but was present in the wild-type mouse. However, the ST2 knockout mice post treatment with IL-13 did develop metaplasia, suggesting a combined cytokine activity of IL-33 and IL-13 in driving metaplasia formation in the gastric mucosa84 (Figure 2A).
Using the SAMP murine model of chronic gastritis, De Salvo et al further showed the importance of IL-33 in the development of SPEM. Over time, SAMP mice develop gastritis and eventually progress to SPEM. In these mice, there was recruitment of eosinophils and M2 macrophages into the inflamed stomach. Either IL-33 inhibition or eosinophil depletion causes a decrease in the infiltration of M2 macrophages and reverses the SPEM features within the stomach. Upon treatment with IL-33, control mice stomachs underwent inflammation and metaplasia development like SAMP mice. When IL-33 was neutralized in the already diseased SAMP mice, the SPEM features partially subsided.85 Thus, IL-33 is a critical cytokine that is involved in the pathway leading from inflammation to metaplasia. Inhibition of IL-33 at the earliest stages of gastric cancer carcinogenesis (formation of metaplasia) provides an interesting avenue for further study as a possible therapeutic preventative that may stop disease formation and progression.
Importance of IL-33 in Gastric Cancer Progression
Studies have illustrated the importance of mast cells in the progression of gastric cancer and have highlighted the involvement of IL-33 in the process. IL-33 is secreted by tumor cells in response to IL-11 and this activates mast cells present in the tumor milieu.86 These activated mast cells secrete the macrophage attracting factors CSF2, CCL3 and IL-6 and are responsible for recruiting tumor associated macrophages to the tumor site. Together, these cells can increase cell proliferation, enhance angiogenesis, and support the tumorigenesis process as well as progression.86 In support of this, in a murine model, tumor burden is stunted when the IL-33 receptor ST2 is inactivated, mast cells are genetically ablated, or the macrophages are depleted from accumulating at the tumor site.86 Taken together, this suggests that IL-33 is involved in the progression of gastric cancer via the activation of mast cells, which in turn attract tumor associated macrophages. Therefore, targeting this signalling cascade might be a strategy for future immune modulating therapies in gastric cancer.86
Supporting the above findings, in a study utilizing an IL-33 deficient mouse model of gastric cancer, Tran et al showed the importance of IL-33 overexpression in gastric cancer progression. Mice lacking IL-33 (gp130F/F/IL33−/−) were found to have low or reduced gastric tumor burden coupled with much less accumulation of myeloid cells, including certain mast cell subsets (pro tumorigenic cells) and immunosuppressive tumor associated macrophages (TAM). The authors showed that IL-33 is involved in mounting a pro-tumorigenic immune environment via mast cell expression of the IL-33 receptor and this is responsible for the recruitment of the alternate immune suppressive TAMs. The TAMs were a secondary source of IL-33 and they can also activate mast cells, so the macrophage population is reinforced at the tumor site. Disruption of the IL-33 signalling pathway either by genetically silencing the expression of IL-33 ligand from the epithelial cells, or by hindering the expression of the IL-33 receptor on the mast cells was sufficient to reduce the gastric tumor burden,87 suggesting a potential therapeutic approach. Yipin et al have also shown that mast cells infiltrating into the tumor sites are activated by upregulated IL-33 in gastric cancer and these mast cells in turn stimulate ICOS expressing T-reg cells, thus creating an immunosuppressive environment.88
IL-33 Mediates Interactions Between Cancer Associated Fibroblasts and GC Cells and is Involved in the Activation of Downstream Signalling Pathways That Promote GC Progression
Studies have suggested that tumor cells express increased levels of TNF-α, which is taken up by the TNF-α receptor present on CAFs, activating the NF-κβ/IRF-1 pathway and inducing CAF IL-33 production.89 Consequently, CAFs secrete high levels of IL-33 which can be internalized by the tumor cells overexpressing the IL-33 receptor. This can activate downstream pathway such as ERK1/2-SP1 and the EMT associated transcription factor ZEB2, potentially increasing the invasiveness and the metastatic potential of the gastric tumor cells89 (Figure 2B). Zhou et al have shown that by co-injection of CAFs with GC cells intraperitoneally into nude mice, the peritoneal metastatic rate (disseminations of gastric cancer cells) in-vivo is reduced upon downregulation of both IL-33 in the CAFs and TNF-α in tumor cells.
Studies have also shown that MAPK pathways (ERK1/2, JNK, p38) are activated by IL-33 signalling in gastric cancer progression and that this is reversed upon IL-33 signalling blockage, further pointing to the pivotal role of IL-33 in GC progression.6 Some functional effects suggested to be influenced by IL-33 signalling include inducing increased GC cell migration and invasion by signalling through ST2 and activating the ERK1/2 pathway, which increases the production of MMP-3 and IL-6. This results in augmented migration, invasion, and GC progression90 (Figure 2B). Blocking either ST2 or the ERK 1/2 pathway resulted in less production of MMP3 and IL-6 with decreased migration and invasion abilities of the GC cells, emphasising the importance of the role of IL-33 in GC progression.90
An in vitro study conducted on the GC cell lines AGS/MKN45 further demonstrated the involvement of IL-33 in the progression of gastric cancer. Treatment of the GC cell lines with IL-33 resulted in increased cell proliferation, migration and invasion, and reduced the apoptosis rate in the cells. Cell cycle progression was evident by the upregulation of CDK4, CDK6 and Cyclin D1. Conversely, all these features were reduced upon disruption of the IL-33 signalling cascade by silencing the expression of the IL-33 receptor ST2. It was also demonstrated that the MAPK pathways (ERK1/2, JNK, p38) are activated by IL-33 signalling in gastric cancer progression and that this is reversed upon IL-33 signalling blockage, further pointing to the pivotal role of IL-33 in GC progression and suggesting blocking of IL-33 may serve as a strategy to decrease GC growth, invasion, and metastasis.6
IL-33 Induces Properties of Stemness and Chemoresistance in Gastric Cancer
The role of IL-33 and its receptor ST2 has also been implicated in supporting cancer stem cell like activity and progression of GC. A subpopulation of cells expressing ST2 and IL-33 can arise spontaneously during GC progression, and this expression level was correlated clinically with cancer stage progression. This subpopulation of cells was found to increase the metastatic abilities of a tumor and confer chemoresistant properties to the tumor cells.91 This subpopulation was found to highly express Bcl-xl, which is regulated by increased MAPK signalling, and was proposed to account for increased survival and tumorigenicity. The authors suggest the IL-33/ST2 axis functions in maintaining self-renewal as well as ensures the survival of the gastric cancer stem cells. This study also revealed that the Wnt pathway and the ST2/IL-33 pathway co-operatively function to support cancer stem cell survival by activating Wnt associated transcription factors. However, upon inhibition of Bcl-xl, this activity of the cancer stem cells was diminished. This study suggested the role of ST2 as a possible cancer stem cell marker in GC, which induces Wnt signalling and presents a possible therapeutic intervention to inhibit GC progression through targeting the IL-33/ST2 and BCl-xl signalling cascade.91
In vitro studies performed on gastric cancer cell lines show that IL-33 protects them from chemotherapy-induced apoptosis. The gastric cancer cell line MGC-803 undergoes apoptosis when treated with cisplatin (a platinum-based drug), but upon administration of IL-33 together with cisplatin, the apoptosis rate of the GC cells is reduced. This suggests that IL-33 induces a protective effect against chemotherapy-based apoptosis in these cells.92 However, there was no change observed in the death rate of the normal GES-1 cell line. This effect was found to function through the MAPK JNK1 pathway and, like other studies, also induces increased GC migration and invasion capacities. Upon blocking this pathway, the protective role of IL-33 was reversed. This Suggests the expression level of IL-33 may be important when platinum-based chemotherapy is utilized for gastric cancer treatment and that inhibition of IL-33 may serve as a sensitizing agent for chemotherapy92 (Figure 2B).
Serum expression levels of IL-33 detected in pre-chemotherapy and post-chemotherapy samples from patients with GC as well as healthy controls revealed that the IL-33 expression was lower in the post-chemotherapy serum samples as compared to pre-chemotherapy samples. The expression level of IL-33 was similar in the post-chemotherapy treated groups and the healthy controls, thus suggesting that chemotherapy can reduce the tumor burden and thereby reducing the level of IL-33 secretion by the tumor.93 It was also found that the rate of decrease of IL-33 was associated with the progression-free survival (PFS) of the patients, with larger decreases of IL-33 being directly related to longer PFS.93 In the patients that were chemoresistant, there was no decrease in the expression of IL-33. Thus, while preliminary, declining expression of IL-33 could be a molecular marker that would indicate response to chemotherapy in the GC patients.93
IL-33 is thus involved in both the early stages of metaplasia induction as well as in gastric cancer progression. Given the effects of IL-33 seen in both metaplasia and gastric cancer, it is reasonable to suspect it may also play a role in progression of metaplasia into invasive cancer. However, future studies will need to further explore this as currently there is no definitive study supporting this hypothesis.
Regulation of IL-33 Expression in Gastric Cancer
IL-33 expression is regulated by the tumor suppressive miRNA miR-496 in gastric cancer and upstream, miR-496 is regulated by the lncRNA NORAD (Figure 3). Huang et al found that the expression of NORAD is upregulated and hence, the expression of its target miR-496 is downregulated in both the epithelial GC cells and CAFs in the tumor microenvironment within gastric cancer tissue samples.94 Upon overexpression of NORAD in CAFs, the proliferation, migration, invasion, and EMT phenotype are all increased, further indicating a tumor promoting role of the CAFs in GC through regulation of the NORAD/miR-496/IL-33 axis. An opposite effect is seen upon inhibition of NORAD, which reverses the tumor inducing effects of the CAFs, as miR-496 expression is upregulated, inhibiting IL-33 expression.94
 |
Figure 3 Regulation of IL-33 expression. Several cell types within the gastric metaplasia and gastric cancer microenvironment including the gastric cancer cells, fibroblasts, and inflammatory cells regulate the expression of IL-33 through multiple mechanisms. All may be potential areas for therapeutic intervention. Abbreviations: WFDC2, WAP-4 disulphide core domain protein 2; TTP, ARE binding protein-tristetraprolin. |
The gene encoding the human epididymis protein 4 called WAP 4-disulphide core domain protein 2 (WFDC2) has been found to be dysregulated in many different cancers. Experiments with gastric cancer models suggest its possible role as a tumor promoting gene by positively regulating the expression of IL-33.95 Using either wild type or WFDC2 knockout mice, three different SPEM models were generated and showed that in the WFDC2 knockout mice the SPEM phenotype appearance is attenuated along with a reduction of M2 macrophage infiltration compared to the wild-type models. However, this state was partially reversed showing SPEM development with WFDC2 supplements. Restoration of WFDC2 expression upregulated IL-33 expression and was associated with more recruitment of M2 macrophages and SPEM formation in the gastric mucosa, highlighting the regulation of IL-33 by WFCD2.95
Studies have also shown the expression of IL-33 is regulated by an ARE binding protein, Tristetraprolin (TTP) in gastric cancer. The expression of TTP and IL-33 is inversely correlated in gastric cancer samples and was associated with survival in gastric cancer patients.95 In in vitro models, overexpression of TTP downregulates the expression of IL-33 and thus reduces the migration, invasion, and proliferation of the GC cells.96
A recent study by Zeng et al suggested a mechanism that can lead to IL-33 overexpression during the early stages of metaplasia formation. In a murine model, they were able to show that loss of mitochondrial GRIM-19 in parietal cells drives the expression of IL-33 through aberrant NRF2 activation by increased oxidative stress, leading to increased gastritis and SPEM formation.97
In summary, multiple mechanisms appear to regulate IL-33 expression. Going forward, if inhibition of IL-33 is desired as a potential clinical therapeutic, further studies will need to determine which of the genes/pathways are best utilized for IL-33 regulation and how the other pathways may compensate to re-establish IL-33 signalling.
Effects of IL-33 Inhibition in Gastric Cancer- Potential Future Therapeutic Target?
IL-33 signalling and regulation is a complex system involving not just the tumor cells themselves, but also several cells within the tumor microenvironment. Of these cells, as reviewed above, CAFs appear to play a central role in releasing IL-33 into the local environment. IL-33 released from CAFs can be taken up by the GC tumor cells which activates the different downstream signalling pathways and enhances the invasion and migration abilities of the tumor cells. In the GC cells, IL-33 induces the activation of MAP kinase (MAPK) pathways (ERK1/2, JNK, p38) to promote the process of EMT and enhance the motility of the GC cells. Hence, to contain the progression of cancer, either neutralization of IL-33 itself or the inhibition of the critical factors involved in the IL-33 signalling cascade might be an option for therapy worth considering. Upon inhibition of IL-33 production or upon inhibition of expression of ST2 on the cancer cells, the entire signalling pathway gets disrupted with downstream effects on the migration and invasion abilities of the cancer cells. These findings credential the idea that inhibition of IL-33 in CAFs or inhibiting the activation of ST2 on the epithelial cells may present a route for therapy in gastric cancer. As studies have shown the pro-IL-33 signalling role of lncRNA NORAD through the inhibition of the IL-33 suppressing miR-496, for therapeutic purpose, this NORAD/miR-496/IL-33 axis is a potential option for targeting to dampen the tumor enhancing abilities of CAFs. In fact, Huang et al demonstrated decreasing the expression of NORAD or upregulating the expression of miR-496 decreased proliferation, migration and invasion in an in vitro model system. Upregulated IL-33 expression is also associated with a decreased rate of apoptosis and cell cycle progression. IL-33 was shown to induce chemoresistance to cisplatin in GC tumor cells. On top of the direct effects of inhibiting IL-33 signalling, sensitizing GC cells to platinum-based agents may be an additional benefit.
Mice lacking IL-33 (gp130F/F/IL33−/−) show low or reduced gastric tumor growth with fewer myeloid cells, including certain mast cell subsets (pro tumorigenic cells) and TAMs. IL-33 is involved in mounting a pro tumorigenic immune environment via mast cells, which attract the accumulation of alternate immune suppressive TAMs at the tumor site. These TAMs become a secondary source of IL-33, which further activates the mast cells, replenishing their own population at the tumor site. Disruption of the IL-33 signalling pathway either by blocking the activation/expression of IL-33 ligand from the epithelial cells, or by blocking the activation/expression of the IL-33 receptor on the mast cells may be beneficial. As discussed above, IL-33 also appears to play a critical role involved in the pathway leading from inflammation to metaplasia. Inhibition of IL-33 at the early stage may serve as a possible therapeutic/preventative treatment that would stop disease progression early before advanced cancer develops.
Potential Anti-Tumorigenic Role of IL-33 in Gastric Cancer
While this review has focused on the pro tumorigenic role IL-33 may play in gastric cancer, it should be noted that several studies have also suggested a possible anti-tumorigenic role. In a contrasting study, using an in vitro model, Pisani et al showed that IL-33 might be involved in inducing antitumorigenic and antiproliferative effects on GC cells.98
Another research study has proposed that IL-33 expression is low in patient samples taken from advanced gastric cancer tissues as compared to the paired adjacent normal tissues. Also, while IL-33 expression could be correlated with the age of the patients or the rate of invasion, its expression could not be correlated to the survival of the gastric cancer patients.99 Similarly, a separate clinical study also showed that the expression of IL-33 is low in gastric tumor tissue samples compared to the paired adjacent normal samples by immunohistochemistry (IHC). The expression of IL-33 was also found to be lower in late-stage disease.100 Importantly, these studies compared IL-33 expression within the gastric cancer to adjacent “normal” tissue. Given the studies suggesting IL-33 role in early gastritis and metaplasia, it is possible that this adjacent tissue is already altered and upregulating IL-33. It could be that while IL-33 is still elevated in advanced gastric cancers, it is less so than in the early stages of disease.
While further studies are needed to clarify the role of IL-33 in later stage disease compared to earlier stage disease, it is possible that IL-33 as well as other inflammatory mediators play different roles depending on the clinical context. If this is the case, it will be important to further clarify when inhibition of IL-33 signalling may be beneficial and when it may cause harm.
Conclusions
Gastric cancer is currently the 4th most common cause of cancer-related mortality worldwide. Given the less than ideal response to current therapies and the poor outcome of many gastric cancer patients, it is imperative to further develop new strategies and targets for improving the overall survival of these patients. It is clear gastric cancer is a complex and dynamic disease driven by a multitude of interactions between stromal, inflammatory, and epithelial cells (both neoplastic and non-neoplastic). To understand what role a singular molecule (IL-33) plays in the process, it is important to understand its temporal and spatial context. As discussed in this review, IL-33 can play multiple roles depending on where and what disease condition it is being expressed.
Immunotherapy is now considered to be an innovative approach for treating different cancers including melanoma, lung cancer, and gastric cancer. Many studies have shown fruitful results with better survival rates in the patients treated with immunotherapy. A better knowledge of immunogenomics of gastric cancer and identification of new immune checkpoint regulators could prove to be very important.101 Given the role of IL-33, targeting this pathway may serve as an additional route to target the tumor microenvironment. However, further research along with a thorough and clear understanding of the dysregulation of the immune system will be needed to enhance immunotherapy treatment in the future.
Interleukins are the crucial mediators which orchestrate the tumor microenvironment by inducing tumor–immune cell crosstalk and they are the key players in large cytokine networks. Interleukin-targeted therapy still has many hurdles that it has to overcome in order to be used as a therapy for gastric cancer in the clinic. However, the intensive foundational research that has been or is ongoing on interleukins in cancer biology shall certainly help to get a better understanding of how interleukin therapy could be applied, carving a path for the discovery of new strategies of treating cancer. Importantly, clinical trials targeting IL-33 have been performed for other non-cancer conditions. Therefore, there are therapies potentially available for trials if/when the pre-clinical research supports their use.
The current review has summarised the general involvement and the therapeutic potentials of IL-33 in gastric cancer progression. Studies have now revealed that IL-33 is involved in regulating early metaplasia formation and gastric cancer progression. IL-33 regulates different cancer promoting signalling cascades that can drive the progression of gastric cancer as well as mediating the interactions and crosstalk between the epithelial cells and CAFs. Studies also suggest IL-33 is involved in induction of resistance to platinum-based chemotherapeutic drugs. Thus, because of this important functional role in driving gastric cancer progression, identifying ways in which to alter IL-33 signalling may be a promising target for gastric cancer therapy.
Funding
This work was supported by NCI R37CA269649 (MDS) and by Brazilian National Council for Scientific and Technological Development - CNPq 154866/2019-9 and 200995/2022-6 (JMAM).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Larsen KM, Minaya MK, Vaish V, Peña MMO. The role of IL-33/ST2 pathway in tumorigenesis. Int J Mol Sci. 2018;19(9):2676. doi:10.3390/ijms19092676
3. Afferni C, Buccione C, Andreone S, et al. The pleiotropic immunomodulatory functions of IL-33 and its implications in tumor immunity. Front Immunol. 2018;9:2601. doi:10.3389/fimmu.2018.02601
4. Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol. 2016;16(11):676–689. doi:10.1038/nri.2016.95
5. Sun P, Ben Q, Tu S, Dong W, Qi X, Wu Y. Serum interleukin-33 levels in patients with gastric cancer. Dig Dis Sci. 2011;56(12):3596–3601. doi:10.1007/s10620-011-1760-5
6. Huang N, Cui X, Li W, Zhang C, Liu L, Li J. IL-33/ST2 promotes the malignant progression of gastric cancer via the MAPK pathway. Mol Med Rep. 2021;23(5):361. doi:10.3892/mmr.2021.12000
7. Siewert JR, Stein HJ. Classification of adenocarcinoma of the oesophagogastric junction. Br J Surg. 1998;85(11):1457–1459. PMID: 9823902. doi:10.1046/j.1365-2168.1998.00940.x
8. Lauren P. The two histological main types of gastric carcinoma: diffuse and so-called intestinal-type carcinoma. An attempt at a histo-clinical classification. Acta Pathol Microbiol Scand. 1965;64:31–49. doi:10.1111/apm.1965.64.1.31
9. Smyth EC, Verheij M, Allum W, Cunningham D, Cervantes A, Arnold D; ESMO Guidelines Committee. Gastric cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2016;27(suppl 5):v38–v49. doi:10.1093/annonc/mdw350
10. Hu B, El Hajj N, Sittler S, Lammert N, Barnes R, Meloni-Ehrig A. Gastric cancer: classification, histology and application of molecular pathology. J Gastrointest Oncol. 2012;3(3):251–261. PMID: 22943016; PMCID: PMC3418539. doi:10.3978/j.issn.2078-6891.2012.021
11. Berlth F, Bollschweiler E, Drebber U, Hoelscher AH, Moenig S. Pathohistological classification systems in gastric cancer: diagnostic relevance and prognostic value. World J Gastroenterol. 2014;20(19):5679. doi:10.3748/wjg.v20.i19.5679
12. Limburg P, Qiao Y-L, Mark SD, et al. Helicobacter pylori seropositivity and subsite-specific gastric cancer risks in Linxian, China. J Natl Cancer Inst. 2001;93:226–233. doi:10.1093/jnci/93.3.226
13. Plummer M, Franceschi S, Vignat J, Forman D, de Martel C. Global burden of gastric cancer attributable to Helicobacter pylori. Int J Cancer. 2015;136(2):487–490. doi:10.1002/ijc.28999
14. Kamangar F, Dawsey SM, Blaser MJ, et al. Opposing risks of gastric cardia and noncardia gastric adenocarcinomas associated with Helicobacter pylori seropositivity. J Natl Cancer Inst. 2006;98:1445–1452. doi:10.1093/jnci/djj393
15. Koshiol J, Wei WQ, Kreimer AR, et al. The gastric cardia is not a target for human papillomavirus-induced carcinogenesis. Cancer Epidemiol Biomarkers Prev. 2010;19(4):1137–1139. doi:10.1158/1055-9965.EPI-10-0089
16. de Martel C, Ferlay J, Franceschi S, et al. Global burden of cancers attributable to infections in 2008: a review and synthetic analysis. Lancet Oncol. 2012;13(6):607–615. doi:10.1016/S1470-2045(12)70137-7
17. Coburn N, Cosby R, Klein L, et al. Staging and surgical approaches in gastric cancer: a systematic review. Cancer Treat Rev. 2018;63:104–115. doi:10.1016/j.ctrv.2017.12.006
18. Allen CJ, Pointer DT, Blumenthaler AN, et al. Chemotherapy versus chemotherapy plus chemoradiation as neoadjuvant therapy for resectable gastric adenocarcinoma: a multi-institutional analysis. Ann Surg. 2021;274:544–548. doi:10.1097/SLA.0000000000005007
19. Ajani JA, D’Amico TA, Bentrem DJ, et al. Gastric cancer, version 2. 2022, NCCN clinical practice guidelines in oncology. J Natl Compr Canc Netw. 2022;20:167–192.
20. In H, Solsky I, Palis B, et al. Validation of the 8th edition of the AJCC TNM staging system for gastric cancer using the national cancer database. Ann Surg Oncol. 2017;24:3683–3691. doi:10.1245/s10434-017-6078-x
21. Janjigian YY, Sanchez-Vega F, Jonsson P, et al. Genetic predictors of response to systemic therapy in esophagogastric cancer genomic biomarkers in esophagogastric adenocarcinoma. Cancer Discov. 2018;8:49–58. doi:10.1158/2159-8290.CD-17-0787
22. Zhang S-X, Liu W, Ai B, et al. Current advances and outlook in gastric cancer chemoresistance: a review. Recent Pat Anticancer Drug Discov. 2022;17:26–41. doi:10.2174/1574892816666210929165729
23. Song Z, Wu Y, Yang J, Yang D, Fang X. Progress in the treatment of advanced gastric cancer. Tumor Biol. 2017;39(7):1010428317714626. doi:10.1177/1010428317714626
24. Chang AY, Foo KF, Koo W, et al. Phase II study of neo-adjuvant chemotherapy for locally advanced gastric cancer. BMJ Open Gastroenterol. 2016;3:e000095. doi:10.1136/bmjgast-2016-000095
25. Yoshikawa T, Rino Y, Yukawa N, Oshima T, Tsuburaya A, Masuda M. Neoadjuvant chemotherapy for gastric cancer in Japan: a standing position by comparing with adjuvant chemotherapy. Surg Today. 2014;44:11–21. doi:10.1007/s00595-013-0529-1
26. Lei ZN, Teng QX, Tian Q, et al. Signaling pathways and therapeutic interventions in gastric cancer. Signal Transduct Target Ther. 2022;7(1):358. doi:10.1038/s41392-022-01190-w
27. Schmitz J, Owyang A, Oldham E, et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity. 2005;23(5):479–490. doi:10.1016/j.immuni.2005.09.015
28. Carriere V, Roussel L, Ortega N, et al. IL-33, the IL-1-like cytokine ligand for ST2 receptor, is a chromatin-associated nuclear factor in vivo. Proc Natl Acad Sci USA. 2007;104(1):282–287. doi:10.1073/pnas.0606854104
29. Zhao W, Hu Z. The enigmatic processing and secretion of interleukin-33. Cell Mol Immunol. 2010;7(4):260–262. doi:10.1038/cmi.2010.3
30. Cayrol C, Girard JP. The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1. Proc Natl Acad Sci USA. 2009;106(22):9021–9026. doi:10.1073/pnas.0812690106
31. Cayrol C, Girard JP. IL-33: an alarmin cytokine with crucial roles in innate immunity, inflammation and allergy. Curr Opin Immunol. 2014;31:31–37. doi:10.1016/j.coi.2014.09.004
32. Tominaga S. A putative protein of a growth specific cDNA from BALB/c-3T3 cells is highly similar to the extracellular portion of mouse interleukin 1 receptor. FEBS Lett. 1989;258(2):301–304. doi:10.1016/0014-5793(89)81679-5
33. Tominaga S, Yokota T, Yanagisawa K, Tsukamoto T, Takagi T, Tetsuka T. Nucleotide sequence of a complementary DNA for human ST2. Biochim Biophys Acta. 1992;1171(2):215–218. doi:10.1016/0167-4781(92)90125-j
34. Yanagisawa K, Takagi T, Tsukamoto T, Tetsuka T, Tominaga S. Presence of a novel primary response gene ST2L, encoding a product highly similar to the interleukin 1 receptor type 1. FEBS Lett. 1993;318(1):83–87. doi:10.1016/0014-5793(93)81333-u
35. Hardman C, Ogg G. Interleukin-33, friend and foe in type-2 immune responses. Curr Opin Immunol. 2016;42:16–24. doi:10.1016/j.coi.2016.05.004
36. Bergers G, Reikerstorfer A, Braselmann S, Graninger P, Busslinger M. Alternative promoter usage of the Fos-responsive gene Fit-1 generates mRNA isoforms coding for either secreted or membrane-bound proteins related to the IL-1 receptor. EMBO J. 1994;13(5):1176–1188. doi:10.1002/j.1460-2075.1994.tb06367.x
37. Tominaga S, Kuroiwa K, Tago K, Iwahana H, Yanagisawa K, Komatsu N. Presence and expression of a novel variant form of ST2 gene product in human leukemic cell line UT-7/GM. Biochem Biophys Res Commun. 1999;264(1):14–18. doi:10.1006/bbrc.1999.1469
38. Tago K, Noda T, Hayakawa M, et al. Tissue distribution and subcellular localization of a variant form of the human ST2 gene product, ST2V. Biochem Biophys Res Commun. 2001;285(5):1377–1383. doi:10.1006/bbrc.2001.5306
39. Iwahana H, Hayakawa M, Kuroiwa K, et al. Molecular cloning of the chicken ST2 gene and a novel variant form of the ST2 gene product, ST2LV. Biochim Biophys Acta. 2004;1681(1):1–14. doi:10.1016/j.bbaexp.2004.08.013
40. Oshikawa K, Yanagisawa K, Tominaga S, Sugiyama Y. Expression and function of the ST2 gene in a murine model of allergic airway inflammation. Clin Exp Allergy. 2002;32(10):1520–1526. doi:10.1046/j.1365-2745.2002.01494.x
41. Millar NL, O’Donnell C, McInnes IB, Brint E. Wounds that heal and wounds that don’t - the role of the IL-33/ST2 pathway in tissue repair and tumorigenesis. Semin Cell Dev Biol. 2017;61:41–50. doi:10.1016/j.semcdb.2016.08.007
42. Funakoshi-Tago M, Tago K, Hayakawa M, et al. TRAF6 is a critical signal transducer in IL-3 signalling pathway. Cell Signal. 2008;20(9):1679–1686. doi:10.1016/j.cellsig.2008.05.013
43. Milovanovic M, Volarevic V, Radosavljevic G, et al. IL-33/ST2 axis in inflammation and immunopathology. Immunol Res. 2012;52(1–2):89–99. doi:10.1007/s12026-012-8283-9
44. O’Donnell C, Mahmoud A, Keane J, et al. An antitumorigenic role for the IL-33 receptor, ST2L, in colon cancer. Br J Cancer. 2016;114(1):37–43. doi:10.1038/bjc.2015.433
45. Akimoto M, Takenaga K. Role of the IL-33/ST2L axis in colorectal cancer progression. Cell Immunol. 2019;343:103740. doi:10.1016/j.cellimm.2017.12.014
46. Lu B, Yang M, Wang Q. Interleukin-33 in tumorigenesis, tumor immune evasion, and cancer immunotherapy. J Mol Med. 2016;94(5):535–543. doi:10.1007/s00109-016-1397-0
47. Jovanovic I, Radosavljevic G, Mitrovic M, et al. ST2 deletion enhances innate and acquired immunity to murine mammary carcinoma. Eur J Immunol. 2011;41(7):1902–1912. doi:10.1002/eji.201141417
48. Jovanovic IP, Pejnovic NN, Radosavljevic GD, et al. Interleukin-33/ST2 axis promotes breast cancer growth and metastases by facilitating intratumoral accumulation of immunosuppressive and innate lymphoid cells. Int J Cancer. 2014;134(7):1669–1682. doi:10.1002/ijc.28481
49. Xiao P, Wan X, Cui B, et al. Interleukin 33 in tumor microenvironment is crucial for the accumulation and function of myeloid-derived suppressor cells. Oncoimmunology. 2015;5(1):e1063772. doi:10.1080/2162402X.2015.1063772
50. Blatner NR, Bonertz A, Beckhove P, et al. In colorectal cancer mast cells contribute to systemic regulatory T-cell dysfunction. Proc Natl Acad Sci USA. 2010;107(14):6430–6435. doi:10.1073/pnas.0913683107
51. Kim JY, Lim SC, Kim G, Yun HJ, Ahn SG, Choi HS. Interleukin-33/ST2 axis promotes epithelial cell transformation and breast tumorigenesis via upregulation of COT activity. Oncogene. 2015;34(38):4928–4938. doi:10.1038/onc.2014.418
52. Hu H, Sun J, Wang C, et al. IL-33 facilitates endocrine resistance of breast cancer by inducing cancer stem cell properties. Biochem Biophys Res Commun. 2017;485(3):643–650. doi:10.1016/j.bbrc.2017.02.080
53. He Z, Chen L, Souto FO, et al. Epithelial-derived IL-33 promotes intestinal tumorigenesis in Apc Min/+ mice. Sci Rep. 2017;7(1):5520. doi:10.1038/s41598-017-05716-z
54. Akimoto M, Maruyama R, Takamaru H, Ochiya T, Takenaga K. Soluble IL-33 receptor sST2 inhibits colorectal cancer malignant growth by modifying the tumour microenvironment. Nat Commun. 2016;7:13589. doi:10.1038/ncomms13589
55. Fang M, Li Y, Huang K, et al. IL33 promotes colon cancer cell stemness via JNK activation and macrophage recruitment. Cancer Res. 2017;77(10):2735–2745. doi:10.1158/0008-5472.CAN-16-1602
56. Maywald RL, Doerner SK, Pastorelli L, et al. IL-33 activates tumor stroma to promote intestinal polyposis. Proc Natl Acad Sci USA. 2015;112(19):E2487–E2496. doi:10.1073/pnas.1422445112
57. Mertz KD, Mager LF, Wasmer MH, et al. The IL-33/ST2 pathway contributes to intestinal tumorigenesis in humans and mice. Oncoimmunology. 2015;5(1):e1062966. doi:10.1080/2162402X.2015.1062966
58. Zhang Y, Davis C, Shah S, et al. IL-33 promotes growth and liver metastasis of colorectal cancer in mice by remodeling the tumor microenvironment and inducing angiogenesis. Mol Carcinog. 2017;56(1):272–287. doi:10.1002/mc.22491
59. Liu X, Zhu L, Lu X, et al. IL-33/ST2 pathway contributes to metastasis of human colorectal cancer. Biochem Biophys Res Commun. 2014;453(3):486–492. doi:10.1016/j.bbrc.2014.09.106
60. Wang K, Shan S, Yang Z, et al. IL-33 blockade suppresses tumor growth of human lung cancer through direct and indirect pathways in a preclinical model. Oncotarget. 2017;8(40):68571–68582. doi:10.18632/oncotarget.19786
61. Akimoto M, Hayashi JI, Nakae S, Saito H, Takenaga K. Interleukin-33 enhances programmed oncosis of ST2L-positive low-metastatic cells in the tumour microenvironment of lung cancer. Cell Death Dis. 2016;7(1):e2057. doi:10.1038/cddis.2015.418
62. Yang Z, Gao X, Wang J, Xu L, Zheng Y, Xu Y. Interleukin-33 enhanced the migration and invasiveness of human lung cancer cells. Onco Targets Ther. 2018;11:843–849. doi:10.2147/OTT.S155905
63. Wang C, Chen Z, Bu X, et al. IL-33 signalling fuels outgrowth and metastasis of human lung cancer. Biochem Biophys Res Commun. 2016;479(3):461–468. doi:10.1016/j.bbrc.2016.09.081
64. Chen SF, Nieh S, Jao SW, et al. The paracrine effect of cancer-associated fibroblast-induced interleukin-33 regulates the invasiveness of head and neck squamous cell carcinoma. J Pathol. 2013;231(2):180–189. doi:10.1002/path.4226
65. Yamada D, Rizvi S, Razumilava N, et al. IL-33 facilitates oncogene-induced cholangiocarcinoma in mice by an interleukin-6-sensitive mechanism. Hepatology. 2015;61(5):1627–1642. doi:10.1002/hep.27687
66. Li J, Razumilava N, Gores GJ, et al. Biliary repair and carcinogenesis are mediated by IL-33-dependent cholangiocyte proliferation. J Clin Invest. 2014;124(7):3241–3251. doi:10.1172/JCI73742
67. Gao K, Li X, Zhang L, et al. Transgenic expression of IL-33 activates CD8(+) T cells and NK cells and inhibits tumor growth and metastasis in mice. Cancer Lett. 2013;335(2):463–471. doi:10.1016/j.canlet.2013.03.002
68. Gao X, Wang X, Yang Q, et al. Tumoral expression of IL-33 inhibits tumor growth and modifies the tumor microenvironment through CD8+ T and NK cells. J Immunol. 2015;194(1):438–445. doi:10.4049/jimmunol.1401344
69. Dominguez D, Ye C, Geng Z, et al. Exogenous IL-33 restores dendritic cell activation and maturation in established cancer. J Immunol. 2017;198(3):1365–1375. doi:10.4049/jimmunol.1501399
70. Kim J, Kim W, Moon UJ, et al. Intratumorally establishing type 2 innate lymphoid cells blocks tumor growth. J Immunol. 2016;196(5):2410–2423. doi:10.4049/jimmunol.1501730
71. Lucarini V, Ziccheddu G, Macchia I, et al. IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. Oncoimmunology. 2017;6(6):e1317420. doi:10.1080/2162402X.2017.1317420
72. Saranchova I, Han J, Huang H, et al. Discovery of a metastatic immune escape mechanism initiated by the loss of expression of the tumour biomarker interleukin-33. Sci Rep. 2016;6(1):30555. doi:10.1038/srep30555
73. Eissmann MF, Dijkstra C, Wouters MA, et al. Interleukin 33 signaling restrains sporadic colon cancer in an interferon-γ-dependent manner. Cancer Immunol Res. 2018;6(4):409–421. doi:10.1158/2326-6066.CIR-17-0218
74. Malik A, Sharma D, Zhu Q, et al. IL-33 regulates the IgA-microbiota axis to restrain IL-1α-dependent colitis and tumorigenesis. J Clin Invest. 2016;126(12):4469–4481. doi:10.1172/JCI88625
75. Qin L, Dominguez D, Chen S, et al. Exogenous IL-33 overcomes T cell tolerance in murine acute myeloid leukemia. Oncotarget. 2016;7(38):61069–61080. doi:10.18632/oncotarget.11179
76. Duault C, Betous D, Bezombes C, et al. IL-33-expanded human Vγ9Vδ2 T cells have anti-lymphoma effect in a mouse tumor model. Eur J Immunol. 2017;47(12):2137–2141. doi:10.1002/eji.201747093
77. Bergis D, Kassis V, Radeke HH. High plasma sST2 levels in gastric cancer and their association with metastatic disease. Cancer Biomark. 2016;16(1):117–125. doi:10.3233/CBM-150547
78. Chang CP, Hu MH, Hsiao YP, Wang YC. ST2 signaling in the tumor microenvironment. Adv Exp Med Biol. 2020;1240:83–93. doi:10.1007/978-3-030-38315-2_7
79. Kuo CJ, Chen CY, Lo HR, et al. Helicobacter pylori induces IL-33 production and recruits ST-2 to lipid rafts to exacerbate inflammation. Cells. 2019;8(10):1290. doi:10.3390/cells8101290
80. Lv Q, Xia Q, Li A, Wang Z. The potential role of IL1RAP on tumor microenvironment-related inflammatory factors in stomach adenocarcinoma. Technol Cancer Res Treat. 2021;20:1533033821995282. doi:10.1177/1533033821995282
81. Buzzelli JN, Chalinor HV, Pavlic DI, et al. IL33 is a stomach alarmin that initiates a skewed Th2 response to injury and infection. Cell Mol Gastroenterol Hepatol. 2015;1(2):203–221.e3. doi:10.1016/j.jcmgh.2014.12.003
82. Meyer AR, Goldenring JR. Injury, repair, inflammation and metaplasia in the stomach. J Physiol. 2018;596(17):3861–3867. doi:10.1113/JP275512
83. Bie Q, Zhang P, Su Z, et al. Polarization of ILC2s in peripheral blood might contribute to immunosuppressive microenvironment in patients with gastric cancer. J Immunol Res. 2014;2014:923135. doi:10.1155/2014/923135
84. Petersen CP, Meyer AR, De Salvo C, et al. A signalling cascade of IL-33 to IL-13 regulates metaplasia in the mouse stomach. Gut. 2018;67(5):805–817. doi:10.1136/gutjnl-2016-312779
85. De Salvo C, Pastorelli L, Petersen CP, et al. Interleukin 33 triggers early eosinophil-dependent events leading to metaplasia in a chronic model of gastritis-prone mice. Gastroenterology. 2021;160(1):302–316.e7. doi:10.1053/j.gastro.2020.09.040
86. Eissmann MF, Dijkstra C, Jarnicki A, et al. IL-33-mediated mast cell activation promotes gastric cancer through macrophage mobilization. Nat Commun. 2019;10(1):2735. doi:10.1038/s41467-019-10676-1
87. Tran CP, Scurr M, O’Connor L, et al. IL-33 promotes gastric tumour growth in concert with activation and recruitment of inflammatory myeloid cells. Oncotarget. 2022;13:785–799. doi:10.18632/oncotarget.28238
88. Lv Y, Tian W, Teng Y, et al. Tumor-infiltrating mast cells stimulate ICOS+ regulatory T cells through an IL-33 and IL-2 axis to promote gastric cancer progression. J Adv Res. 2023. PMID: 37086778. doi:10.1016/j.jare.2023.04.013
89. Zhou Q, Wu X, Wang X, et al. The reciprocal interaction between tumor cells and activated fibroblasts mediated by TNF-α/IL-33/ST2L signaling promotes gastric cancer metastasis. Oncogene. 2020;39(7):1414–1428. doi:10.1038/s41388-019-1078-x
90. Yu XX, Hu Z, Shen X, Dong LY, Zhou WZ, Hu WH. IL-33 promotes gastric cancer cell invasion and migration via ST2-ERK1/2 pathway. Dig Dis Sci. 2015;60(5):1265–1272. doi:10.1007/s10620-014-3463-1
91. Kwon JW, Seok SH, Kim S, et al. A synergistic partnership between IL-33/ST2 and Wnt pathway through Bcl-xL drives gastric cancer stemness and metastasis. Oncogene. 2023;42(7):501–515. doi:10.1038/s41388-022-02575-5
92. Ye XL, Zhao YR, Weng GB, et al. IL-33-induced JNK pathway activation confers gastric cancer chemotherapy resistance. Oncol Rep. 2015;33(6):2746–2752. doi:10.3892/or.2015.3898
93. Hu W, Wu C, Li X, et al. Serum IL-33 level is a predictor of progression-free survival after chemotherapy. Oncotarget. 2017;8(21):35116–35123. doi:10.18632/oncotarget.16627
94. Huang C, Liu J, He L, et al. The long noncoding RNA noncoding RNA activated by DNA damage (NORAD)-microRNA-496-Interleukin-33 axis affects carcinoma-associated fibroblasts-mediated gastric cancer development. Bioengineered. 2021;12(2):11738–11755. doi:10.1080/21655979.2021.2009412
95. Jeong H, Lee B, Kim KH, et al. WFDC2 promotes spasmolytic polypeptide-expressing metaplasia through the up-regulation of IL33 in response to injury. Gastroenterology. 2021;161(3):953–967.e15. doi:10.1053/j.gastro.2021.05.058
96. Deng K, Wang H, Shan T, et al. Tristetraprolin inhibits gastric cancer progression through suppression of IL-33. Sci Rep. 2016;6:24505. doi:10.1038/srep24505
97. Zeng X, Yang M, Ye T, et al. Mitochondrial GRIM-19 loss in parietal cells promotes spasmolytic polypeptide-expressing metaplasia through NLR family pyrin domain-containing 3 (NLRP3)-mediated IL-33 activation via a reactive oxygen species (ROS) -NRF2- Heme oxygenase-1(HO-1)-NF-кB axis. Free Radic Biol Med. 2023;202:46–61. PMID: 36990300. doi:10.1016/j.freeradbiomed.2023.03.024
98. Pisani LF, Tontini GE, Gentile C, et al. Proinflammatory interleukin-33 induces dichotomic effects on cell proliferation in normal gastric epithelium and gastric cancer. Int J Mol Sci. 2021;22(11):5792. doi:10.3390/ijms22115792
99. Hu W, Li X, Li Q, et al. Interleukin-33 expression does not correlate with survival of gastric cancer patients. Pathol Oncol Res. 2017;23(3):615–619. doi:10.1007/s12253-016-0167-1
100. Liu QH, Zhang JW, Xia L, et al. Clinical implications of interleukins-31, 32, and 33 in gastric cancer. World J Gastrointest Oncol. 2022;14(9):1808–1822. doi:10.4251/wjgo.v14.i9.1808
101. Patel TH, Cecchini M. Targeted therapies in advanced gastric cancer. Curr Treat Options Oncol. 2020;21(9):70. doi:10.1007/s11864-020-00774-4
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.