Back to Journals » Cancer Management and Research » Volume 9
Interleukin-1 receptor antagonist inhibits angiogenesis via blockage IL-1α/PI3K/NF-κB pathway in human colon cancer cell
Authors Ma J, Sun X, Guo T, Su H, Chen Q, Gong Z, Qi J, Zhao X
Received 29 July 2017
Accepted for publication 6 September 2017
Published 9 October 2017 Volume 2017:9 Pages 481—493
DOI https://doi.org/10.2147/CMAR.S147699
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Jiachi Ma,1 Xiaowen Sun,2 Tiankang Guo,1 He Su,1 Quan Chen,1 Zhenqiang Gong,3 Jianbo Qi,3 Xiaodan Zhao3
1Department of General Surgery, Gansu Provincial People’s Hospital, LanZhou, 2Department of Dermatology, The First Hospital of Tianshui, Tianshui, 3Ningxia Medical University Graduate School of Medical Sciences, Surgical Oncology, Yinchuan, People’s Republic of China
Purpose: This article investigates the relationship between cancer cells and stromal cells in carcinoma cell living microenvironment and elucidates the mechanism that interleukin-1 receptor antagonist (IL-1RA) blocks metastatic potential in colon cancer.
Methods: Western blot and RT-PCR assay were used to determine the expression of hepatocyte growth factor (HGF) and IL-1α in colon carcinoma cells and stromal cells. Effect of IL-1RA and HGF on metastatic potential of colon cancer cells were examined by proliferation, invasion, and angiogenesis assays. The interactional role of IL-1RA and HGF were detected by ELISA assay, invasion, and angiogenesis assay making up a co-culture system consisting of stromal and colon cancer cells in cells living microenvironment.
Results: IL-1α was expressed in human umbilical vein endothelial cells (HUVECs) and HT-29 and WiDr (colon cancer cell lines with higher liver metastatic potential). HGF was expressed only in fibroblast. HGF secretion from fibroblasts was significantly inhibited by IL-1RA (P<0.01). Furthermore, IL-1RA could significantly inhibit migration, proliferation, and angiogenesis of HUVECs (P<0.01). In the double co-culture system, there is a high liver metastatic potential of colon cancer cell line (HT-29) because it can secrete autocrine IL-1α, significantly enhanced angiogenesis compared with low liver metastatic cell line (CaCo-2) (P<0.01), which does not secrete IL-1α. On the contrary, blockage of autocrine IL-1α by IL-1RA might significantly decrease metastatic potential of colon carcinoma cells through downregulation of IL-1α/PI3K/NF-κB pathway.
Conclusion: IL-1 receptor antagonist (IL-1RA) is an important inhibitor in metastatic process of colon carcinoma cell. Based on the above results, we suggest that IL-1RA may be a promising new therapeutic approach in inhibiting colon cancer with IL-1-producing patients.
Keywords: interleukin receptor antagonist, hepatocyte growth factor, metastasis, angiogenesis, colon cancer, IL-1RA, HGF, angiogenesis, colon carcinoma
Introduction
Colorectal carcinoma is one of the deadliest cancers due to its high propensity to metastatic potential. It accounts for ~1 million new incidence of colon cancer cases and causes a half million deaths annually worldwide.1–3 The main treatment of colorectal cancer at present are surgical radical operation, chemotherapy, radiotherapy, and targeted therapy.4 There have not been highly effective approaches against metastasis of colorectal cancer so far. Recently, researching on the microenvironment of solid tumors shows that chemokines and their receptors act as key function in cancer metastatic process, and chemokines play their specific role in the regulation of angiogenesis, activation of a tumor-specific immune response, and inducing proliferation of tumor cell in an autocrine or paracrine fashion.5–8
Development of new blood vessels is the critical path in the progressive growth of solid tumor. Angiogenesis is critical for metastasis of carcinoma cell, which not only carries oxygen and essential nutrients to the tumor, but also facilitates the spread and proliferation of cancer cells.9 The states of angiogenesis are essential for both tumor cell growth and metastasis. Angiogenesis is a complicated and multistep process involving extracellular matrix remodeling, migration, and proliferation of endothelial cell and capillary vessel formation.10 These processes depend on the production of angiogenic factors secreted by both tumor and stromal cells.11 Angiogenesis provides a gateway for the tumor cells entering the circulation and metastasizing to distant organs, such as liver or lung. Thus, it is a critical step in tumor cell metastasis.
Interleukin (IL)-1α is an important inflammatory cytokine. It is produced by cancer cells, such as colon, pancreas, lung, ovary, and gastric cancer. IL-1α can induce the expression of adhesion molecules in vascular endothelial cells and enhance expression of proteases in pancreatic cancer.12,13 Recent research showed that IL-1α was highly expressed in tumor environment and was secreted by tumor cells or stromal cells. IL-1α binds to the IL-1 receptor 1 (IL-1R1), which then forms a complex with the IL-1 receptor accessory protein (IL-1RAcP).10,25 It can stimulate the development of cancers. However, the underlying mechanism that IL-1α influences the tumor behaviors remains poorly characterized. Therefore, the relationships between cells and IL-1 in tumor environment is not fully understood, especially critical is the exploration of the innate pathways of IL-1 regulating metastatic potential in tumor microenvironments.14
Hepatocyte growth factor (HGF), a kind of pleiotropic cytokine, acts as a highly specific mitogen for endothelial cells that regulates cell proliferation, invasiveness, survival, and tumor angiogenesis.15,16 MET proto-oncogene, receptor tyrosine kinase (c-Met) is the receptor for HGF. c-Met overexpression is associated with the proliferation, invasion and metastasis of cancer cells.17 HGF has been identified as a stromal cells-derived epithelial morphogen that induces branching tubular morphogenesis. The stromal cells-derived HGF plays an important role in cancer cell’s invasiveness and proliferation.18-20 Fibroblasts production of HGF has been regulated by IL-1, tumor necrosis factor (TNF), and basic fibroblast growth factor (bFGF), and leading to the enhanced invasion and proliferation ability of carcinoma cells.21 IL-1 receptor antagonist (IL-1RA) is an endogenous inhibitor of IL-1 that prevents the activation of IL-1 receptors by inhibiting. IL-1RA is connected with a variety of diseases, including cardiovascular disease, type II diabetes, carcinoma and joint diseases.22 IL-1RA has been shown to improve the symptoms of the disease in the treatment of pre-clinical animal PTOA (posttraumatic osteoarthritis) models.23 IL-1RA is also a promising drug for the treatment of stroke. It is reported that it may improve the immune response in traumatic brain injury and subarachnoid hemorrhage.24
The authors should describe more about chemical and/or physiological characteristics of IL-1RA. However, the underlying mechanisms that IL-1 or IL-1RA influences HGF biochemical reaction behind the tumor behavior and the relationship between cancer cells and stromal cells are still unknown, especially in tumor environment.
In order to clarify the relationship between cancer cells and stromal cells in the tumor living microenvironment and elucidate the mechanisms that IL-1α and IL-1RA influence the metastatic potential in colon cancer cells, we detected the expression of IL-1α and HGF in fibroblasts, human umbilical vein endothelial cells (HUVECs), and different liver metastatic potential of colon carcinoma cell lines. Then, we investigated whether IL-1RA affects HGF secretion from fibroblasts, and how IL-1RA influences metastatic potential of colon carcinoma.
Methods
Cell lines and culture conditions
Four types of human colon carcinoma cell lines (HT-29, WiDr, CaCo-2, and Colo320) were obtained from American Type Culture Collection (Rockville, MD, USA). HT-29 was incubated in McCoy’s supplemented with 10% fetal bovine serum (FBS). WiDr and CaCo-2 were cultured in minimum essential medium Eagle (Sigma Chemical Co., St. Louis, MO, USA) with 10% FBS and high glucose. Colo320 was maintained in RPMI-1640 medium (Sigma Chemical Co.) supplemented with 10% FBS. HUVEC was purchased from Kurabo Co. (Osaka, Japan). HUVECs were maintained in HuMedia-EG2 medium supplemented with 2% FBS, 5 ng/mL basic fibroblast growth factor, 10 µg/mL heparin, 10 ng/mL epidermal growth factor, and 1 µg/mL of hydrocortisone according to the supplier’s instruction (Kurubo Co.). Fibroblast was obtained from Lonza (Walkersville, MD, USA) and cultured in fibroblast medium-2 (FBM-2) medium supplemented with 2% FBS, 1 ng/mL basic fibroblast growth factor, and 1 mg/mL insulin, according to the supplier’s instructions. All the cells were cultured at 37°C in a humidified atmosphere of 5% CO2 in air.
Experiment reagents and antibodies
Recombinant human HGF and anti-HGF antibody were obtained from R&D systems (Minneapolis, MN, USA), and recombinant human IL-1α was provided by Diaclone (Beasancon, France), while recombinant human IL-1RA was purchased from Pepro Tech EC Ltd (London, UK).
Reverse transcription polymerase chain reaction (RT-PCR) analysis
The total RNA was extracted from all cell lines using the RNA OUT kit, and then the RNA concentration was determined by using DNA/RNA calculator. cDNA was synthesized by reverse transcription using 5 μg of RNA, and the cDNA was used as a template for PCR amplification of IL-1α, HGF, and c-Met gene. Reverse transcription-polymerase chain reaction (RT-PCR) system included 10× RT buffer, 25 mM MgCl2, 0.1 M dithiothreitol (DTT), RNaseOUT, and 200U SuperScript™ ІІІ RT, which was placed in the water temperature box at 50°C for 50 min and at 85°C for 5 min. About 1 μL of cDNA was used for amplification reaction, and the operations were performed, according to the supplier’s instructions. Primer sequence and PCR conditions are shown in Table 1.
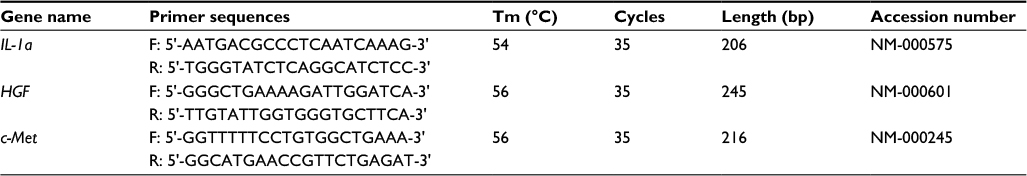 | Table 1 Primer sequence and PCR condition Abbreviations: HGF, hepatocyte growth factor; IL-1α, interleukin-1α; PCR, polymerase chain reaction; Tm, temperature. |
Western blot analysis
The colon cells and stromal cells were lysed by lysis buffer consisting of 25 mM Tris (pH 7.8) with H3PO, 2 mM EDTA (ethylene diamine tetraacetic acid), 10 mM DTT, 10% glycerol, 1% Triton® X-100, 2 mM phenylmethylsulfonyl fluoride, 1 mM sodium orthovanadate, and 10 µM leupeptin. The protein concentrations were measured by BCA protein assay kit (Pierce, Rockford, IL, USA). About 30 µg of protein samples per lane was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene membrane (Immobilo PVDF; Nihon Millipore Ltd, Tokyo, Japan). The membrane was incubated in blocking buffer for 60 minutes at room temperature. The blocking buffer was formed by dissolving 5% nonfat dry milk into Tris-buffered saline which contained 0.1% Tween 20 (TBS-T). The membranes were washed by TBS-T and then immunoblotted by each being diluted into 1:1000–2000 of primary antibody overnight at 4ºC. Then, the membranes were washed by TBS-T three times. The primary antibody was combined by horseradish peroxidase-conjugated secondary antibody in Tris-buffered saline for 60 minutes at room temperature. The complexes of protein antibody were visualized with an ECL Western blotting detection and analysis system (Amersham Biosciences, Buckinghamshire, UK). β-Actin served as a loading control.
Enzyme-linked immunosorbent assay
Fibroblast was implanted at a concentration of 2×105 cells/mL into a 24-well plate using the media with 2% FBS incubating overnight, and then the medium was exchanged to culture the cells for another 24 hours. The media were collected, and microcentrifuged at 1,500 rpm for 10 minutes to remove the particulates, and then the supernatants were obtained, the supernatants were frozen until ELISA assay by ELISA Kit (R&D Systems) in accordance with the instructions of the manufacturer. To evaluate the influence of IL-1α and IL-1RA on HGF production from fibroblasts, the culturing fibroblasts were stimulated by IL-1α (10 ng/mL) or IL-1RA (10 ng/mL), or fibroblasts were cultured for another 24 hours. The concentration of HGF was measured by ELISA assay. To investigate the co-operative role and interaction of tumor–stromal cells, we next examined the effect of IL-1RA on the production of HGF from fibroblasts by adopting a new technology that consisted of a double-chamber system containing fibroblasts and colon cancer cells in a 24-well plate. The fibroblasts were implanted at a concentration of 2×105 cells/mL into a 24-well plate containing medium with 10% FBS and cultured overnight. The medium was exchanged (0.5% FBS). Medium was exchanged (0.5% FBS), the medium was added or not added IL-1α or IL-1RA, then the 5×104 cells/mL of HT-29 and CaCo-2 were planted into inserts with 0.45-mm pores (Kurabo Co.), and the pores were placed into 24-well plates. The co-culture systems were cultured for another 24 hours, and subsequently the concentrations of HGF were measured according to the manufacturer’s protocol.
Proliferation of HUVEC in the presence of HGF or anti-HGF antibody
HUVECs were implanted at a concentration of 5×103 cells/100 mL in 96-well plates overnight, and the medium was exchanged with medium alone (control) or media supplemented with different concentration of HGF or 10 mg/mL of anti-HGF antibody. About 10 mL of WST-1 cell proliferation assay reagent was put in each well, and the cells were incubated for 72 hours at 37°C. HUVEC proliferation was determined by Premix WST-1 cell proliferation assay system (Takara Bio Ing, Shiga, Japan). We determined the absorbance by a microplate reader (Molecular Devices LLC, Sunnyvale, CA, USA) at a test wavelength of 450 nm and reference wavelength of 690 nm.
Proliferation of HUVEC treatment with fibroblasts or cancer cell-cultured supernatants
To prepare the conditioned media, 1×106/2 mL of fibroblast was seeded into a chamber of 24-well plate to culture with FBM-2 medium containing 2% FBS, while 5×104/mL of HT-29 or CaCo-2 cells was inoculated into trans-well chamber containing 0.45-mm pores of polycarbonate membranes with 2% FBS of McCoy’s or DMEM, and then the trans-well chambers were plated into 24-well chambers. The co-cultured system was cultured for 24 hours. The cultured medium was gathered and microcentrifuged at 1,500 rpm for 10 minutes. The supernatants were used in HUVEC proliferation assay. About 2×103 cells/100 mL of HUVECs were implanted in a 96-well plate with HuMedia-EB2 medium to incubate overnight, and then, the cells were incubated with HuMedia-EB2 medium only (control), conditioned media (supernatants from HT-29 and CaCo-2 co-cultured with fibroblast system mixed with HuMedia-EB2 medium [1:1]), and the media were exchanged every day. HUVECs were incubated for 72 hours, and the proliferation of HUVEC was measured by the Premix WST-1 cell proliferation assay as described earlier.
Migration of HUVEC pretreated with HGF or co-culture with fibroblasts or pretreated IL-1α or IL-1RA
We used the BioCoat Matrigel Invasion Chambers (Bencton Dickinson, Bedford, MA, USA) to confirm the migration of HUVECs in vitro. First, HUVEC was inoculated at the density of 1×105/mL cells into the Martrigel precoated trans-wells containing polycarbonate membranes with 8 mm pores. Tran-well chambers were placed in 24-well plate with basic medium alone (control), or medium pretreated with 1, 10, 100 ng/mL of HGF, or 10 mg/mL of anti-HGF antibody. After being cultured for 24 hours, the upper surfaces of trans-wells were wiped with cotton swab and the invaded cells were fixed and stained with Diff-Quik kit. The invaded cells were counted in five microscope fields (×200). To further examine the effect of fibroblast-derived HGF on migration of HUVEC, a double-chamber method was adopted to confirm the migration assay for HUVEC. We measured the invaded HUVECs as follows: fibroblast was implanted at a density of 1×105/mL cells into 24-well plate with FGM-2 medium with or without 10 μg/mL of anti-HGF Ab or 10 ng/mL of IL-1α or 10 ng/mL of IL-1α and 10 ng/mL of IL-1RA. About 5×104/mL of fibroblasts was inoculated into trans-well chamber containing of 0.45-mm pores of polycarbonate membranes and cultured in a medium with 2% FBS, and then the trans-well chambers were laid into 24-well plates. The co-cultured system was incubated for 24 hours, and then the invaded HUVECs were measured.
HGF influence angiogenesis activity in vitro
To research the effect of HGF on tubular formation by HUVEC, we co-cultured HUVEC and fibroblast in base medium only or in basal medium containing different concentration of HGF adopting an Angiogenesis Kit (Kurabo Co.). In brief, HUVECs and fibroblasts were co-cultured in 24-well plate chamber with base medium alone (control) or base medium with 1, 10, or 100 ng/mL of HGF or 10 mg/mL of anti-HGF Ab. HUVECs and fibroblasts were totally incubated for 12 days, and the medium was changed every 2 days. HUVEC tubular formation was stained with anti-CD31 antibody by the protocols of the manufacturer. The area of tubular formation was measured quantitatively over five different fields for each condition using an Angiogenesis Image Analyzer (Kurabo Co., Osaka, Japan).
Effect of co-cultured colon cancer cells on angiogenesis in vitro
We further investigated the influence of different colon cancer cells (HT-29 secreted IL-1α or CaCo-2 not secreted IL-1α) on tubule formation by HUVEC. HT-29 or CaCo-2, HUVECs, and fibroblasts were co-cultured using a double-chamber method. First, HUVEC/fibroblasts were cultured in a 24-well plate for 3 days, and then HT-29 or CaCo-2 cells (5×104 cells/mL) were inoculated into trans-well chambers in which the bottom of chamber consisted of polycarbonate membrane with 0.45-mm pores, and then trans-well chambers were placed in HUVEC/fibroblast cultured 24-well plate. Approximately 10 ng/mL of IL-1α, IL-1RA, or anti-HGF Ab was added to the cultured medium, and the medium was changed every 2 days. All the cells were totally incubated for 12 days, then the upper trans-wells were removed, and the 24-well plate chambers were washed with 100 mL of 1× PBS three times. Then the HUVEC tubular formation was stained with anti-CD31 antibody by the protocols of manufacturer, and the total area of tubular formation was measured as described earlier.
Measurement of IL-1α and HGF concentrations in angiogenesis supernatants by ELISA
In order to investigate the relationship between colon cancer cells and tumor microenvironment, we examined the concentrations of IL-1α and HGF protein in the supernatants of angiogenesis assay. Cultured media of angiogenesis assay on day 12 were gathered and microcentrifuged at 1,500 rpm for 10 minutes to get rid of the particles. The concentrations of IL-1α and HGF in the media were determined by ELISA assay.
Statistical analysis
All data are presented as means ± standard deviations (SD). Differences in the mean of two groups were analyzed by an unpaired t-test. Multiple group comparison was performed by one-way analysis of variance (ANOVA) with a post hoc test for subsequent individual group comparisons. P<0.05 was considered statistically significant. Mean values and SD were calculated for experiments performed in triplicate (or more).
Results
Expression of HGF, c-Met, and IL-1α in colon cancer cell lines, HUVEC, and fibroblast
Colon carcinoma cell lines were previously classified into two groups by intrasplenic liver metastatic assay; one is high liver metastatic potential cell lines (HT-29 and WiDr) and the other group is low liver metastatic potential cell lines (CaCo-2 and CoLo320).25 RT-PCR experiments showed that IL-1α mRNA was expressed by the higher liver metastatic colon cancer cell lines HT-29, WiDr, and HUVEC. HGF mRNA was expressed only by fibroblast. c-Met is the receptor for HGF. c-Met mRNA was shown in all cells, but HGF mRNA was discovered only by fibroblast (Figure 1A). Western blot assay showed that IL-1α protein was also revealed in HT-29, WiDr, and HUVEC. HGF protein was expressed only by fibroblast, and c-Met protein was detected in all cell lines (Figure 1B).
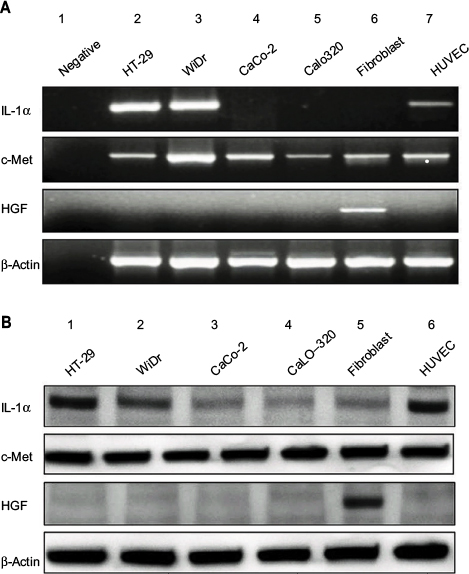 | Figure 1 Expression of IL-1a, HGF and c-Met. Notes: (A) Expression of IL-1α, HGF, and c-Met in colon cancer cell lines and stromal cells. PCR products stained with ethidium bromide were subjected to 1.5% agarose gel electrophoresis. (B) The protein expression of IL-1α, HGF, and c-Met in colon cancer cell lines and stromal cells were confirmed by Western blotting analysis. We used the method of 10% SDS-PAGE to dispose 30 µg of total cell lysate and then transferred them to polyvinylidenedifluoride membrane which was probed with anti-IL-1α, HGF, and c-Met antibodies. β-Actin served as a loading control. Abbreviations: HGF, hepatocyte growth factor; IL-1α, interleukin-1α; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis. |
The secretion levels of HGF from fibroblasts were influenced by recombinant human IL-1α, colon cancer cell-derived IL-1α and IL-1RA
The secretion levels of HGF by stromal cells were measured by ELISA assay. HGF production from fibroblast was increased by IL-1α (compared with control, P<0.01). Likewise, the secretion level of HGF from fibroblast was significantly enhanced when co-cultured with HT-29 cells (1.563-folds versus control, P<0.01), and the enhanced production of HGF by co-culturing with HT-29 cells was obviously decreased by the presence of IL-1RA (P<0.01, Figure 2A). The secretion level of HGF from fibroblast has no significant effect of co-culturing with CaCo-2, but the treatment of IL-1α significantly increased the secretion level of HGF in this co-culture system (P<0.01, Figure 2B).
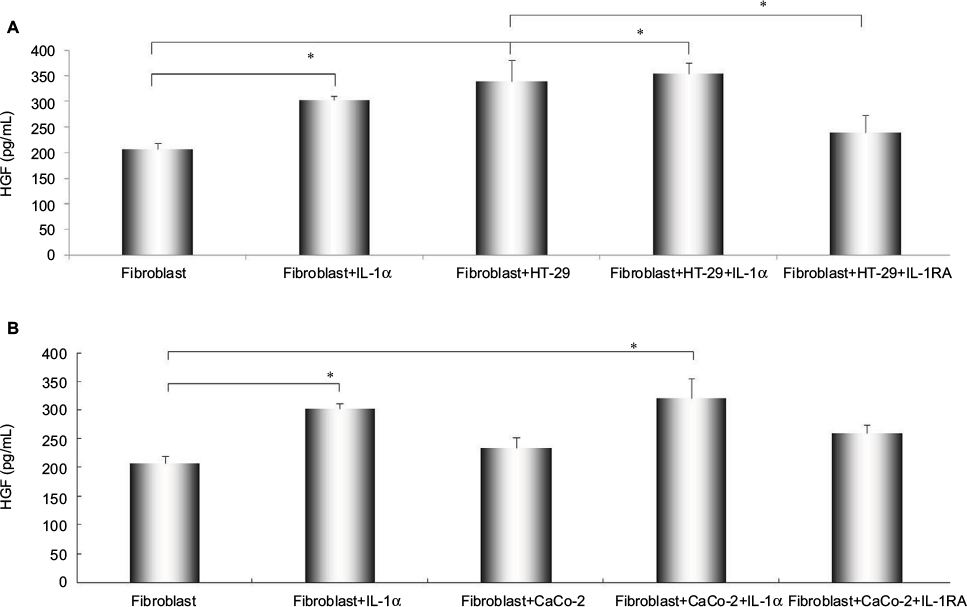 | Figure 2 The effect of IL-1RA and co-culture with different metastatic potential of colon cancer cells on the secretion of HGF from fibroblasts. Notes: Alteration of HGF production from fibroblasts were pretreated by recombinant human IL-1α (10 ng/mL), co-cultured with colon cancer cells [(A) HT-29 and (B) CaCo-2], and IL-1RA (10 ng/mL). Fibroblasts were cultured for 48 hours, and the concentration of HGF was measured by ELISA assay. Multiple comparisons were made by using the method of one-way ANOVA, followed by Student–Newman–Keuls test. Values are expressed as mean ± SD. Error bars indicated SD, *P<0.01. Abbreviations: ANOVA, analysis of variance; HGF, hepatocyte growth factor; HUVEC, human umbilical vein endothelial cell; IL-1α, interleukin-1α; SD, standard deviation. |
HGF promoted HUVEC proliferation
In order to investigate the effect of HGF on the proliferation of HUVEC, the proliferation assay was performed. The HUVEC proliferation was enhanced by HGF in a concentration-dependent manner. About 100 ng/mL of HGF significantly enhanced the proliferation of HUVECs (P<0.01). Interestingly, anti-HGF antibody significantly inhibited the enhanced proliferation in presence of HGF in culture media (Figure 3A).
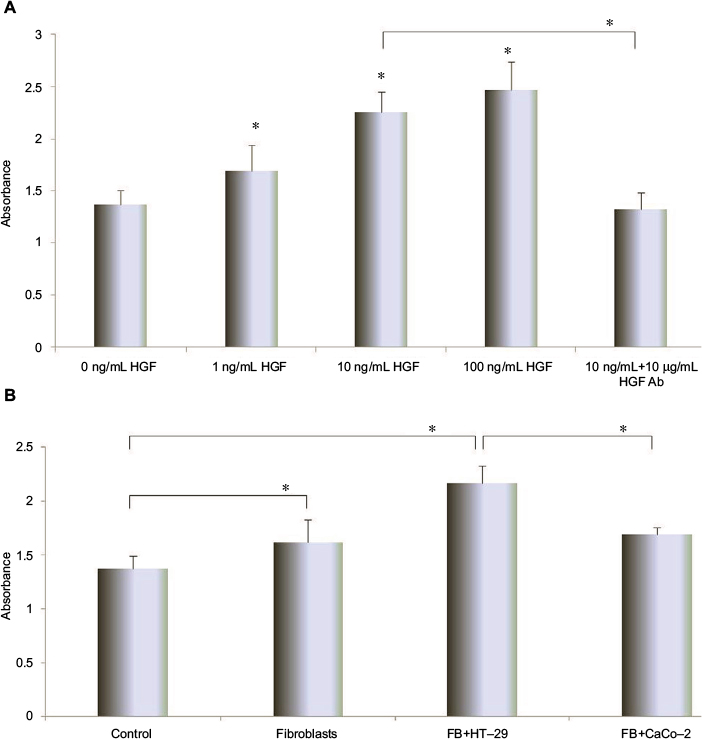 | Figure 3 The effect of increasing concentrations of HGF and anti-HGF antibody on HUVEC proliferation. Notes: (A) The different conditioned HGF or anti-HGF antibody influenced HUVEC proliferation. (B) The effect of conditioned cell media from HT-29 and CaCo-2 on HUVEC proliferation. HUVECs were pretreated with CM from fibroblasts or different metastatic potential of colon carcinoma cell lines. HUVECs were cultured for 72 hours, and then HUVEC proliferations were measured by the premix WST-1 cell proliferation assay. The absorbance was measured at 450 and 690 nm. Multiple comparisons were made by using one-way ANOVA, followed by Student–Newman–Keuls test. *P<0.01. Each experiment was performed for more than three times. Abbreviations: ANOVA, analysis of variance; CM, conditional media; HGF, hepatocyte growth factor; HUVEC, human umbilical vein endothelial cell; SD, standard deviation. |
The cell-cultured supernatants enhanced HUVEC proliferation
The condition medium effect of HUVEC proliferation was measured by WST-1 Cell Proliferation Assay. HUVEC proliferation was significantly enhanced by the addition of fibroblasts, fibroblast + HT-29, and fibroblast + CaCo-2 conditioned media (P<0.01 compared with control), and fibroblast + HT-29 most significantly increased the proliferation of HUVECs (P<0.01 compared with fibroblast and fibroblast + CaCo-2; Figure 3B).
Effect of HGF, co-cultured with fibroblast and IL-1RA on HUVEC migration
The migrating capability of HUVEC was promoted by HGF in a dose-dependent manner (P<0.01, compared with control), 100 ng/mL of HGF significantly enhanced HUVEC migration, and the enhanced migration was inhibited by anti-HGF antibody (P<0.01; Figure 4A). Co-culture with fibroblasts caused significant enhancement on HUVEC migration (P<0.01), and this enhanced role was inhibited by anti-HGF antibody (P<0.01). Moreover, the addition of IL-1α significantly promoted the migration of HUVEC when co-cultured with fibroblast system and inhibited in the presence of IL-1RA as well (P<0.01; Figure 4B).
 | Figure 4 HGF and co-culturing fibroblast influence on migration of HUVEC. Notes: (A) The effect of different concentrations of HGF on HUVEC migration. HUVECs were treated with different concentrations of HGF and/or anti-HGF antibody and incubated for 24 hours. The invading cells were fixed and stained with Diff-Quick stain. The invading cells were counted in five random microscopic fields (×200). Multiple comparisons were made by using one-way ANOVA, followed by Student–Newman–Keuls test. Columns indicate relative invading numbers. Error bars indicate SD. *P<0.01. (B) The effect of co-culturing with fibroblasts and IL-1RA on HUVEC migration. The HUVEC migration influenced by co-culturing with fibroblasts or IL-1RA was assessed by the BD Bio-Coat Matrigel invasion assay as described in “Migration of HUVEC pretreated with HGF or co-culture with Fibroblasts or pretreated IL-1α or IL-1RA” section. To assess the influence of fibroblast-derived HGF on HUVEC migration, HUVECs were co-cultured with fibroblast pretreated with or without IL-1α or anti-HGF antibody or IL-1RA. The invading cells were incubated for 24 hours and then fixed and stained with Diff-Quick stain. The invading cells were counted in five random microscopic fields (×200). Multiple comparisons were made by using one-way ANOVA, followed by Student–Newman–Keuls test. Bars indicate SD, *P<0.01. All experiments were performed at least three times. Abbreviations: ANOVA, analysis of variance; HGF, hepatocyte growth factor; HUVEC, human umbilical vein endothelial cell; IL-1RA, interleukin-1 receptor antagonist; IL-1α, interleukin-1α; SD, standard deviation. |
Influence of HGF on angiogenesis by HUVEC
To further examine the role of HGF in tube formation by HUVEC, we measured the influence of HGF on HUVEC tube formation by using angiogenesis assay. The tubular formation was obviously promoted by the presence of HGF in a dose-dependent manner (P<0.01, compared with control), and about 100 ng/mL of HGF most significantly increased the tube formation of HUVEC (P<0.01, compared with control). On the contrary, anti-HGF antibody could inhibit the tube formation by HUVEC (P<0.01, compared with 10 ng/mL of HGF group; Figure 5A).
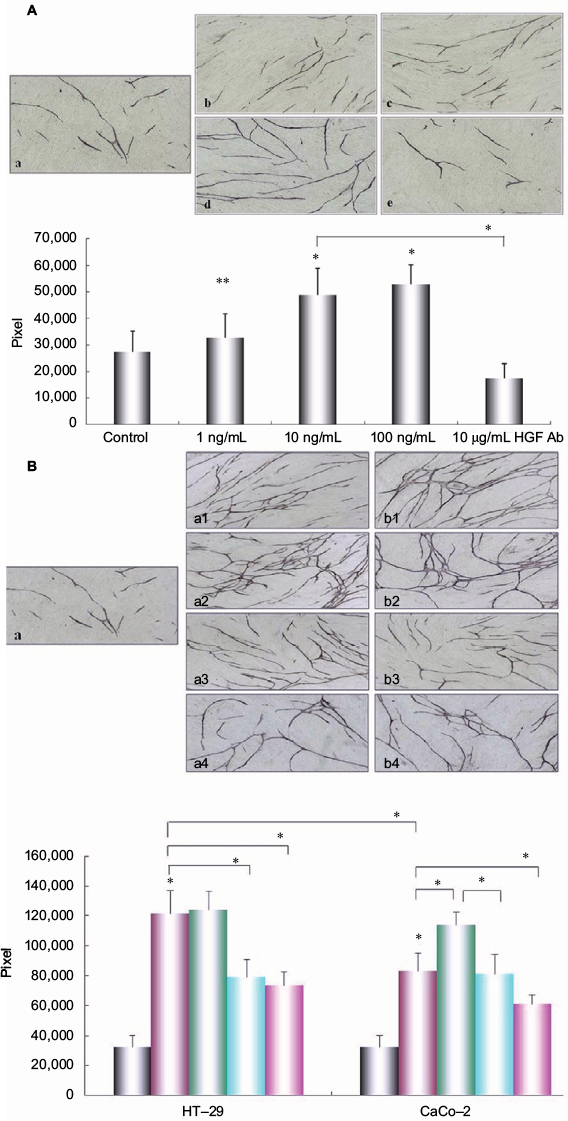 | Figure 5 Effect of HGF and co-cultured colon cancer cells on angiogenesis. Notes: (A) Different concentrations of HGF impacted the angiogenesis by HUVEC. The HUVEC/fibroblast co-culture system was incubated in the presence or absence of different concentrations of HGF or anti-HGF antibody for 11 days, and then the incubated system was stained by anti-CD31 antibody. An image analyzer was used to measure the tube formation area quantitatively. (a) Control; (b) 1 ng/mL of HGF; (c) 10 ng/mL of HGF; (d) 100 ng/mL of HGF; (e) 10 µg/mL of anti-HGF antibody. Columns represent mean pixels of HUVEC tube formation area; error bars represent SD. Multiple comparisons were made by using one-way ANOVA, followed by Student–Newman–Keuls test. *P<0.01 versus control. Magnification: ×40. (B) Effect of co-culture with different metastatic potential colon cancer cells (HT-29) and (CaCo-2) on HUVEC tubular formation. HUVECs/fibroblasts were co-cultured with HT-29 or CaCo-2 with or without IL-1α, anti-HGF antibody, or IL-1RA by using the double-chamber method. The co-cultured system was incubated for 11 days, and the tube formation was measured as described earlier. Magnification: ×40. (a) Control; (a1) co-cultured with HT-29 cells; (a2) co-cultured with HT-29 cells pretreated with 10 ng/mL of IL-1a; (a3) co-cultured with HT-29 cells pretreated with 10 ng/mL of IL-1RA; (a4) co-cultured with HT-29 cells pretreated with 10 ng/mL of anti-HGF antibody; (b1) Co-cultured with CaCo-2 cells; (b2) co-cultured with CaCo-2 cells pretreated with 10 ng/mL of IL-1α; (b3) co-cultured with CaCo-2 cells pretreated with 10 ng/mL of IL-1RA; (b-4) co-cultured with CaCo-2 cells pretreated with 10 ng/mL of anti-HGF antibody. Columns represent mean pixels of HUVEC tube formation area; error bars represent SD. Multiple comparisons were made by using one-way ANOVA, followed by Student–Newman–Keuls test. *P<0.01. Key to graph in (B): Gray, control; purple, co-culture with colon cancer cells; green, co-cultured with colon cancer cells pretreated with 10 ng/mL of IL-1a; blue, co-culture with colon cancer cells pretreated with 10 ng/mL of IL-1RA; lavender, co-culture with colon cancer cells pretreated with 10 ng/mL of anti-HGF Ab. Abbreviations: ANOVA, analysis of variance; HGF, hepatocyte growth factor; HUVEC, human umbilical vein endothelial cell; IL-1RA, interleukin-1 receptor antagonist; IL-1α, interleukin-1α; SD, standard deviation. |
Effect of colon cancer cells with or without IL-1α on tube formation
We also investigated the influence of various liver metastatic potentials of colon cancer cells on the tube formation by HUVEC. We co-cultured colon cancer cells and HUVECs/fibroblasts by adopting double-chamber methods to examine the relationship among them. The tube formation was significantly promoted when co-cultured with HT-29 cells compared with control (HUVECs and fibroblasts only) or co-cultured with CaCo-2 cells (P<0.01; Figure 5B). Furthermore, the presence of IL-1α could obviously promote the tubular formation when co-cultured with CaCo-2 cell system (P<0.01). By comparison, the enhanced tubular formation by HUVEC was obviously inhibited by the addition of IL-1RA when co-cultured with HT-29 cell system (P<0.01) and was significantly inhibited by anti-HGF antibody in HT-29 or CaCo-2 cell co-culture system (P<0.01).
Concentration of IL-1α and HGF in the angiogenesis assay supernatants
To investigate the interaction between colon cancer cells and stromal cells in microenvironment, the concentrations of IL-1α and HGF containing the angiogenesis assay of supernatants were examined by ELISA assay. IL-1α concentration in the supernatant of HUVEC/fibroblast co-cultured with HT-29 cells was higher (16.54±1.72 pg/mL) than HUVEC/fibroblast only or cultured with CaCo-2 cells (3.91±0.13 pg/mL, P<0.01). Similarly, HGF concentration in the supernatant of HUVEC/fibroblast co-cultured with HT-29 cells was high (382.89±31.05 pg/mL, P<0.01 compared with HUVEC/fibroblast only or co-cultured with CaCo-2). HGF protein concentration in HUVEC/fibroblast co-cultured with CaCo-2 and IL-1α was high (379.40±30.51 pg/mL, P<0.01 compared with HUVEC/fibroblast only or co-cultured with CaCo-2), but this promoted action was inhibited by IL-1RA (240.71±23.86 pg/mL, P<0.01 compared with HUVEC/fibroblast co-cultured with CaCo-2 + IL-1α group). The concentration of HGF was inhibited by IL-1RA co-cultured with HT-29 cell system (P<0.01). But there is no effect of IL-1RA co-cultured with CaCo-2 cell system (Table 2).
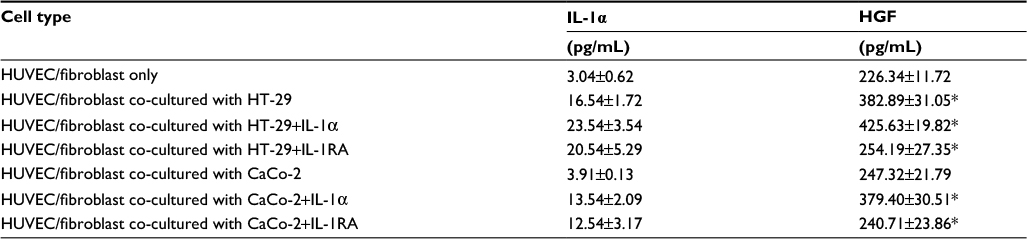 | Table 2 Concentrations of HGF and IL-1α proteins in angiogenesis assay supernatant Notes: In order to evaluate the interactions between colon cell lines and the tumor microenvironment, we measured IL-1a and HGF protein concentrations in the angiogenesis assay supernatants. Culture media from angiogenesis assays were collected on day11 and microfuged at 1500 rpm for 10 min to remove any particles, and the concentrations of IL-1a and HGF in the media were then measured by ELISA. Statistical significance was tested by one-way ANOVA followed by the Dunnett test. Statistical significance was indicated by P<0.05. All data are expressed as mean ± standard deviation.*P<0.01 compared with HUVEC/fibroblast only or co-cultured with CaCo-2 cells. Abbreviations: HGF, hepatocyte growth factor; HUVEC, human umbilical vein endothelial cell; IL-1RA, interleukin-1 receptor antagonist; IL-1α, interleukin-1α. |
Activation of PI3K/NF-κB signaling pathway after the stimulation of IL-1α in fibroblast
Stromal cell fibroblast was used to determine IL-1α/PI3K/NF-κB signaling pathway activated by IL-1α stimulation. The result showed that 100 ng/mL of IL-1α could increase PI3K and NF-κB (Figure 6A) phosphorylation in a time-dependent manner. To investigate the effect of IL-1RA and PI3K inhibitor on the activation of NF-κB in stromal cell after stimulation by IL-1α, fibroblasts were pretreated with 100 ng/mL of IL-1α for 15 minutes, then 10 µg/mL of IL-1RA or 50 µM of LY294002 (PI3K inhibitor) was added to the culture medium and stimulated for 15 minutes. The proteins were extracted and separated by SDS-PAGE and transferred to membranes, and the membranes were probed with antibody directed against phospho-NF-κB, phospho-PI3K, total NF-κB and total PI3K. We found that IL-1α-mediated phospho-NF-κB was inhibited by 10 µg/mL of IL-1Ra. Phospho-NF-κB was significantly blocked by 50 µM of LY294002. These data indicate that IL-1RA inhibited the metastatic potential of colon cancer depending on downregulation of IL-1α/PI3K/NF-κB signaling pathway (Figure 6B).
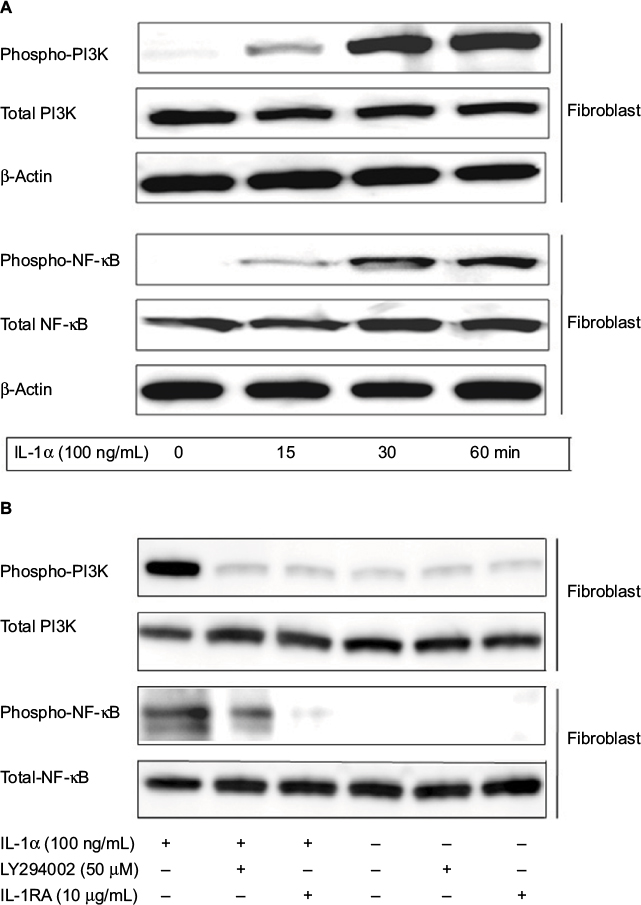 | Figure 6 IL-1α-induced phosphorylation of PI3K/NF-κB signaling in fibroblast. Notes: (A) Fibroblasts were stimulated with 100 ng/mL of IL-1α, and then fibroblasts were cultured for 5, 10, and 30 minutes. The cells were collected and lysed by lysis buffer. Approximately 30 μg of lysed protein was used to perform immunoblotting with a phospho-PI3K and phospho-NF-κB antibody. Detection of total PI3K or NF-κB levels served as a loading control. (B) Fibroblasts after being pretreated with 100 ng/mL of IL-1RA and 50 μM LY294002 for 1 hour were incubated with 100 ng/mL of IL-1α for 1 hour, and the cells were immunoblotted by using PI3K and NF-κB antibody. Detection of total NF-κB levels served as a loading control. Abbreviations: IL-1α, interleukin-1α; IL-1RA, interleukin-1 receptor antagonist. |
Discussion
Our previous studies elucidated that IL-1α expression was a feature in which mRNA and protein were determined only in high liver metastatic colon cancer cell lines HT-29 and WiDr but not detected in the low liver metastatic cell lines CaCo-2 and Colo320. IL-1α is one of the most important inflammatory cytokines, is involved in the metastatic process of colon cancer, and regulates the tumor growth, progression, and liver metastasis in gastric carcinoma and pancreatic cancer. Pancreatic carcinoma cell-derived IL-1α increases the secretion level of HGF by stromal cells with a paracrine manner, and enhanced HGF expression promotes invasion, proliferation, and angiogenesis of cancer cells.25–28 Present research offered a new viewpoint about cooperative interaction between cancer cells and stromal cells with respect to the biological effects of HUVEC/fibroblast co-culture cytokines. We investigated how IL-1RA influences the progression of colon cancer in living microenvironment, how it affects this, and what is the relationship between cancer cells and living microenvironment. Our results showed that colon carcinoma cell-derived IL-1α increased the secretion level of HGF from fibroblast in a paracrine manner and that enhanced HGF expression promoted HUVEC proliferation, migration, and tube formation. On the contrary, these enhanced roles were significantly inhibited by IL-1RA through the blockage of IL-1/PI3K/NF-κB pathway.
The metastasis of colon carcinoma is associated with the abundant stromal cells, and the tumor–stromal cell interaction is known to play a critical role in tumor progression. Recent studies have proved that tumor growth and metastasis formation are influenced by the tumor microenvironment, which consists of cancer cells, HUVECs, fibroblasts, and a group of cytokines.29,30 Chemokines are highly expressed in tumor microenvironment and have a close connection with tumor behaviors, such as tumorigenesis, proliferation, angiogenesis, and metastasis.31–33 Presently, a study showed that IL-1α and c-Met were overexpressed in colon cancer tissues, and the overexpression was related to poor prognosis.26 Meanwhile, patients with IL-1α and c-Met overexpression exhibited a worse prognosis compared to patients with either IL-1α or c-Met overexpression alone or decreased expression of these proteins. Correlation analysis revealed that IL-1α expression was significantly associated with c-Met expression in colon cancer tissues.34 We found that IL-1α could enhance the expression of HGF secreted from fibroblasts, which is an upstream signal molecule in c-Met signaling pathway in tumor microenvironment.
As the results of our experiments showed that IL-1α was confirmed in HT-29, WiDr, and HUVEC, HGF was expressed only by fibroblasts, and c-Met was expressed in both colon cell lines and stromal cells (HUVECs and fibroblasts). The ELISA results demonstrated that secretion level of HGF from fibroblast was enhanced by IL-1α and when co-cultured with HT-29 cells, which could secrete IL-1α, and the enhanced HGF secretion was significantly blocked by IL-1RA. We also examined the effect of IL-1RA on the proliferation of colon cancer cells, but were not able to confirm a significant relationship between IL-1RA and carcinoma cell proliferation (data not shown). However, endogenous (condition supernatants from fibroblast-cultured medium) and exogenous HGF could enhance HUVEC proliferation. These results indicate that IL-1RA could inhibit the proliferation of HUVECs in tumor living microenvironment. Therefore, we further investigated the effect of HGF and IL-1RA on the migration of HUVECs using a double-chamber cultured method to examine the effect of HGF and IL-1RA on the interactions between cancer cells and stromal cells in the tumor living microenvironment. Our results indicate that HUVEC migration was significantly enhanced by HGF and when co-cultured with fibroblasts, and this ability was inhibited by IL-1RA. This enhanced migration could be inhibited through blocking IL-1α signaling to decrease HGF secretion. These results suggest that vascular endothelial cells located near high liver metastatic tumors that secreted high levels of inflammatory cytokine (IL-1α) are able to proliferate and migrate faster than endothelial cells near tumors that do not secrete IL-1α and that these functions are blocked by IL-1RA. Furthermore, we also investigated capillary tube formation by HUVEC by adopting a HUVEC/fibroblast or colon cancer cell co-cultured system. The tube formation of HUVECs was significantly promoted when co-cultured with HT-29 cells, and this effect was decreased. In contrast, no significant difference was detected between HUVECs co-cultured with and without CaCo-2 cells by IL-1RA. For a more comprehensive understanding of the interactions between carcinoma cancer cells and tumor living microenvironment, we examined IL-1α and HGF concentrations with angiogenesis assay supernatants. The secreted IL-1α and HGF proteins with the supernatants of HUVEC/fibroblast co-cultured with HT-29 cells were higher than those with the supernatants from control HUVEC/fibroblasts or from those co-cultured with CaCo-2 cells. These results suggest that colon cancer cells with high liver metastatic potential also have a high angiogenic potential and that fibroblast-derived HGF plays an important role in angiogenesis and metastasis. IL-1RA blocked the IL-1α/PI3K/NF-кB signaling to inhibit HUVEC angiogenesis that was enhanced by HT-29 cells, but the inhibited levels did not completely return to a basal level. One might conjecture that colon cancer cells produce other angiogenic cytokines, such as IL-6 and IL-8. IL-1 signaling is also associated with tumor growth, angiogenesis, metastasis, and cancer-associated wasting syndrome.35 Activation of IL-1 signaling pathway leads to the expression of numerous inflammatory cytokines involved in tumor survival and in the alterations of various immune cells to the tumor, which further promotes the inflammatory and pro-survival microenvironment of the tumor.36
HGF, which was secreted by several types of carcinoma cells and stromal cells, is a growth-promoting factor that plays an important role in cancer metastasis. HGF was overexpressed in colon cancer cells. Recent molecular profiling has shown that activation of PI3K/NF-κB signaling is associated with aggressive disease and poor prognosis. These data suggested that enhanced HGF secreted by the stromal cells might be responsible for the metastasis of colon cancer in the microenvironment. Both HGF and c-Met were upstream signal molecular target that activate and enhance phosphatidylinositol 3-kinase (PI3K), and the activation of PI3K/NF-κB or MAPK pathway is involved in multiple mechanisms responsible for cancer progression, including apoptosis, proliferation, migration, and angiogenesis.37–40 In our next study, we have planned to use carcinoma cells with shRNA knockout of the cytokines or its receptor genes to investigate the relationship between AKT/NF-κB or MAPK signaling pathway and cytokines in cancer metastatic process and further search for more potent angiogenesis-modulating agents to be used in the clinical treatment of colon cancers as a new therapeutic technique.
In summary, we have demonstrated that IL-1RA not only decreases the proliferation and invasion of HUVEC but also suppresses angiogenesis, and this role is involved in the interaction between colon cancer cells and the tumor living microenvironment depending on the inhibition of secreted HGF from fibroblast by downregulating IL-1α/HGF/PI3K signaling pathway. Blockage of the IL-1 receptor system has recently been shown to be a critical event in the development of several human tumors. Our results indicate that blockage of IL-1α/PI3K/NF-κB signaling phosphorylation may be a major mechanism by which IL-1RA inhibits cancer cell metastatic potential and induce apoptosis. Based on our findings, it is suggested that IL-1RA may be a new beneficial clinical therapeutic approach to inhibit the aggressive spread of patients with IL-1-producing cancers.
Conclusion
IL-1RA could downregulate the secretion of HGF in a paracrine manner, and this role is involved in the interaction between colon cancer cells and tumor living microenvironment. IL-1RA blocking IL-1α/HGF signaling pathway significantly inhibited the metastatic potential in colon cancer. Our data suggest that IL-1RA may be a new beneficial clinical therapeutic approach to inhibit the aggressive spread of patients with IL-1-producing cancers.
Acknowledgment
This study was supported by National Natural and Science Foundation of People’s Republic of China (81260325).
Author contributions
All authors contributed to the conception and design of the study, acquisition of data, or analysis and interpretation of data. JCM, XWS, TKG, HS, QC, ZQG, JBQ and XDZ collected data; JCM, XWS, TKG, HS, QC, ZQG, JBQ and XDZ contributed to the analysis and the interpretation of data; and JCM, XWS, TKG and HS drafted the manuscript. All authors contributed to critically revising the manuscript for important intellectual content and approved the final manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
Parkin DM. Epidemiology of cancer: global patterns and trends. Toxicol Lett. 1998;102:227–234. | ||
Akouchekian M, Houshmand, M, Hemati, S, Shafa M. Occurrence of large-scale mitochondrial DNA deletions in human colorectal cancer. Arch Med Sci. 2008;4:249–253. | ||
Akouchekian M, Houshmand, M. Mitochondrial haplogroup analysis in colorectal cancer: identification of a high-risk population. Arch Med Sci. 2008;4:397–400. | ||
Kim GR, Ha GH, Bae JH. Metastatic colon cancer cell populations contain more cancer stem-like cells with a higher susceptibility to natural killer cell-mediated lysis compared with primary colon cancer cells. Oncol Lett. 2015;9:1641–1646. | ||
Ning Y, Manegold PC, Hong YK, et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int J Cancer. 2011;128:2038–2049. | ||
Balkwill, F. Cancer and the chemokine network. Nat Rev Cancer. 2004;4:540–550. | ||
Araki S, Omori Y, Lyn D, et al. Interleukin-8 is a molecular determinant of androgen independence and progression in prostate cancer. Cancer Res. 2007;67:6854–6862. | ||
Waugh DJ, Wilson C. The interleukin-8 pathway in cancer. Clin Cancer Res. 2008;14:6735–6741. | ||
Liotta LA, Kohn EC. The microenvironment of the tumour-host interface. Nature. 2001;411:375–379. | ||
Ma J, Sawai H, Matsuo Y, et al. Interleukin-1 enhances angiogenesis and is associated with liver metastatic potential in human gastric cancer lines. J Surg Res. 2008;148:197–204. | ||
Folkman J. What is the evidence that tumors are angiogenesis dependent? J Natl Cancer Inst. 1990;82:4–6. | ||
Kaji M, Ishikura H, Kishimoto T, et al. E-selectin expression induced by pancreas carcinoma-derived interleukin-1α results in enhanced adhesion of pancreas-carcinoma cells to endothelial cells. Int J Cancer. 1995;60:712. | ||
Guo B, Fu S, Zhang J, Liu B, Li Z. Targeting inflammasome/IL-1 pathways for cancer immunotherapy. Sci Rep. 2016;12:6. | ||
Weber A, Wasiliew P, Kracht M. Interleukin-1 (IL-1) Pathway. Sci Signal. 2010;3:1–7. | ||
Cecchi F, Rabe DC, Bottaro DP. Targeting the HGF/Met signaling pathway in cancer therapy. Expert Opin Ther Targets. 2012;16:553–572. | ||
Sakai K, Aoki S, Matsumoto K. Hepatocyte growth factor and Met in drug discovery. J Biochem. 2015;157:271–284. | ||
Birchmeier C, Birchmeier W, Gherardi E, Woude GF Vande. Met, metastasis, motility and more. Nat Rev Mol Cell Biol. 2003;4:915–925. | ||
Bussolino F, DiRenzo, MF, Ziche M. Hepatocyte growth factor is a potent angiogenic factor which stimulates endothelial cell motility and growth. J Cell Biol. 1992;119:629–641. | ||
Montesano R, Matsumoto K, Nakamura T, Orci L. Identification of a fibroblast-derived epithelial morphogen as hepatocyte growth factor. Cell. 1991;67:901–908. | ||
Qian LW, Mizumoto K, Maehara N, et al. Co-cultivation of pancreatic cancer cells with orthotopic tumor-derived fibroblasts: fibroblasts stimulate tumor cell invasion via HGF secretion whereas cancer cells exert a minor regulative effect on fibroblasts HGF production. Cancer Lett. 2003;190:105–112. | ||
Nakamura T, Matsumoto K, Kiritoshi A, Tano Y, Nakamura T. Induction of hepatocyte growth factor in fibroblasts by tumor-derived factors affects invasive growth of tumor cells: in vitro analysis of tumor-stromal interactions. Cancer Res. 1997;57:3305–3313. | ||
Bell S, Mehta G, Moore K, Britton A. Ten-year alcohol consumption typologies and trajectories of C-reactive protein, interleukin-6 and interleukin-1 receptor antagonist over the following 12 years: a prospective cohort study. J Intern Med. 2017;281:75–85. | ||
Morshedy SA, Elsaadany HF, Ibrahim HE, Sherif AM, Farghaly MA, Allah MA. Interleukin-1beta and interleukin-1receptor antagonist polymorphisms in Egyptian children with febrile seizures: a case-control study. Medicine (Baltimore). 2017;96:e6370. | ||
McCann SK, Cramond F, Macleod MR, Sena ES. Systematic review and meta-analysis of the efficacy of interleukin-1 receptor antagonist in animal models of stroke: an update. Transl Stroke Res. 2016;7:395–406. | ||
Matsuo Y, Sawai H, Ma J, et al. IL-1alpha secreted by colon cancer cells enhances angiogenesis: the relationship between IL-1alpha release and tumor cells’ potential for liver metastasis. J Surg Oncol. 2009;99(6):361–367. | ||
Xu D, Matsuo Y, Ma J, et al. Cancer cell-derived IL-1α promotes HGF secretion by stromal cells and enhances metastatic potential in pancreatic cancer cells. J Surg Oncol. 2010;102:469–477. | ||
Jiachi M, Hirozumi S, Yoichi M, et al. IGF-1 and PTEN regulate the proliferation and invasiveness of colon cancer cells through opposite effects on PI3K/Akt signalling. J Clin Res. 2009;5(2):195–206. | ||
JiaChi M, XiaoWen S, He S, Quan C, TianKang G. Fibroblast-derived CXCL12/SDF-1α promotes CXCL6 secretion and co-operatively enhances metastatic potential through the PI3K/Akt/mTOR pathway in colon cancer. World J Gastroenterol. 2017;23(28):5167–5178. | ||
Goubran HA, Kotb RR, Stakiw J. Regulation of tumor growth and metastasis: the role of tumor microenvironment. Cancer Growth Metastasis. 2014;2:9–18. | ||
Simpson KD, Templeton DJ, Cross JV. Macrophage migration inhibitory factor promotes tumor growth and metastasis by inducing myeloid-derived suppressor cells in the tumor microenvironment. J Immunol. 2012;189:5533–5540. | ||
Hara Y, Ogata Y, Shirouzu K. Early tumor growth in metastatic organs influenced by the microenvironment is an important factor which provides organ specificity of colon cancer metastasis. J Exp Clin Cancer Res. 2000;19:497–504. | ||
Nasser MW, Elbaz M, Ahirwar DK, Ganju RK. Conditioning solid tumor microenvironment through inflammatory chemokines and S100 family proteins. Cancer Lett. 2015;365:11–22. | ||
O’Hayre M, Salanga CL, Handel TM, Allen SJ. Chemokines and cancer: migration, intracellular signalling and intercellular communication in the microenvironment. Biochem J. 2008;409:635–649. | ||
Bonfil R, Chinni S, Fridman R, Kim HR, Cher ML. Proteases, growth factors, chemokines, and the microenvironment in prostate cancer bone metastasis. Urol Oncol. 2007;25:407–411. | ||
Voronov E, Apte RN. IL-1 in colon inflammation, colon carcinogenesis and invasiveness of colon cancer. Cancer Microenviron. 2015;8:187–200. | ||
Dinarello CA. An expanding role for interleukin-1 blockade from gout to cancer. Mol Med. 2014;20(1):S43–S58. | ||
Voronov E, Carmi Y, Apte RN. The role IL-1 in tumor-mediated angiogenesis. Front Physiol. 2014;5:114. | ||
Atif F, Yousuf S, Stein DG. Anti-tumor effects of progesterone in human glioblastoma multiforme: role of PI3K/Akt/mTOR signaling. J Steroid Biochem Mol Biol. 2014;146:62–73. | ||
Chatterjee M, Andrulis M, Stuhmer T, et al. The PI3K/Akt signaling pathway regulates the expression of Hsp70, which critically contributes to Hsp90- chaperone function and tumor cell survival in multiple myeloma. Haematologica. 2013;98:1132–1141. | ||
Samartzis EP, Noske A, Dedes KJ, Fink D. ARID1A mutations and PI3K/AKT pathway alterations in endometriosis and endometriosis-associated ovarian carcinomas. Int J Mol Sci. 2013;14:18824–18849. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.