Back to Journals » OncoTargets and Therapy » Volume 13
Interactions Between lncRNA TUG1 and miR-9-5p Modulate the Resistance of Breast Cancer Cells to Doxorubicin by Regulating eIF5A2
Authors Wang S, Cheng M, Zheng X, Zheng L, Liu H, Lu J ![]() , Liu Y, Chen W
, Liu Y, Chen W ![]()
Received 9 July 2020
Accepted for publication 1 December 2020
Published 23 December 2020 Volume 2020:13 Pages 13159—13170
DOI https://doi.org/10.2147/OTT.S255113
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Nicola Silvestris
Shuqian Wang,1,* Mengjing Cheng,2,* Xiaoxiao Zheng,2,* Li Zheng,2 Hao Liu,2 Jianju Lu,3 Yu Liu,1 Wei Chen2
1Department of Breast Surgery, First Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou 310003, People’s Republic of China; 2Cancer Institute of Integrated Traditional Chinese and Western Medicine, Key Laboratory of Cancer Prevention and Therapy Combining Traditional Chinese and Western Medicine, Zhejiang Academy of Traditional Chinese Medicine, Tongde Hospital of Zhejiang Province, Hangzhou, Zhejiang 310012, People’s Republic of China; 3Department of Breast Surgery, The First Hospital of Jiaxing, Affiliated Hospital of Jiaxing College, Jiaxing 314000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yu Liu
Department of Breast Surgery, First Affiliated Hospital, School of Medicine, Zhejiang University, 79 Qingchun Road, Hangzhou, Zhejiang 310006, People’s Republic of China
Tel +8613516805637
Email [email protected]
Wei Chen
Cancer Institute of Integrated Traditional Chinese and Western Medicine, Zhejiang Academy of Traditional Chinese Medicine, Tongde Hospital of Zhejiang Province, 234 Gucui Road, Hangzhou, Zhejiang 310012, China
Tel +8618668097597
Email [email protected]
Purpose: Breast cancer (BC) is one of the leading causes of cancer-related deaths. Chemoresistance of BC remains a major unmet clinical obstacle. TUG1 (taurine-upregulated gene 1), a long noncoding RNA (lncRNA), and microRNAs (miRNA) are implicated in therapeutic resistance. However, the interactions between TUG1 and miRNAs that regulate doxorubicin (Dox) resistance in BC remain elusive.
Materials and Methods: Expression of TUG1 and miR-9 was measured by real-time PCR. EIF5A2 (eukaryotic translation initiation factor 5A-2) was detected by Western blot. Transfection of siRNAs or miRNA inhibitors was applied to silence lncRNA TUG1, eIF5A2 or miR-9. Cell viability, proliferation, and apoptosis were determined by CCK-8 (cell counting kit-8), flow cytometry, and EdU (5-ethynyl-2ʹ-deoxyuridine) assays, respectively. The regulatory relationship between TUG1 and miR-9 was determined by a luciferase assay.
Results: LncRNA TUG1 was highly expressed in BC tissues and positively associated with Dox resistance in BC cell lines. SiRNA knockdown of TUG1 reversed Dox resistance in MCF-7/ADR cells. Mechanistically, TUG1 acted as a “sponge” for miR-9 and downregulated miR-9. Treatment with a miR-9 inhibitor blocked the effect of TUG1 siRNA, and knockdown of TUG1 inhibited the effects of miR-9. Furthermore, TUG1 inhibition of apoptosis induced by Dox involved miR-9 targeting of eIF5A2.
Conclusion: TUG1 modulates the susceptibility of BC cells to Dox by regulating the expression of eIF5A2 via interacting with miR-9. These results indicate that the lncRNA TUG1 may be a novel therapeutic target in breast cancer.
Keywords: TUG1, microRNA-9-5p, breast cancer, doxorubicin resistance, eIF5A2
Introduction
Breast cancer (BC) is one of leading cause of cancer-related mortality globally, has the highest incidence of malignant tumors among women, and is a significant public health concern.1 Doxorubicin (Dox) is an anthracycline drug that is commonly used in the effective treatment of breast cancer, and Dox resistance is a major barrier to BC therapy.2,3 Therefore, strategies that enable clinicians and researchers to explore the mechanisms of Dox resistance in BC and to prevent the chemoresistance are urgently needed.
Long noncoding RNAs (LncRNAs) are an abundant and functionally diverse species of noncoding RNAs (ncRNAs).4,5 LncRNAs play key roles in regulating gene expression related to drug resistance, growth, differentiation, and development.6,7 MicroRNAs (miRNAs) are small ncRNAs that negatively modulate gene expression.8,9 LncRNAs communicate with and regulate miRNAs by acting as competing endogenous RNAs (ceRNAs), or natural miRNA sponges.10
The lncRNA taurine-upregulated gene 1 (TUG1) directly binds to polycomb repressive complex 2 (PRC2) or PRC1 represses gene expression, and has been reported to participate in oncogenic processes.11,12 TUG1 acts as a key regulator of drug resistance by sponging miRNAs and involved in the control of some cancer-related genes.13–15
Here, using comparative profiling of lncRNAs between breast cancer tissues and peritumor tissues, we identified lncRNA TUG1 (LncTUG1) as an upregulated lncRNA in BC. Using Starbase, we found the presence of a consensus-binding site for miR-9-5p in the TUG1 lncRNA. Meanwhile, a number of studies have reported dysregulation of miR-9-5p was implicated in the occurrence and development of breast cancer.16–18 Our previous study indicated that miR-9 was closely related to the sensitivity of chemotherapy drugs.19 We also identified a negative correlation between TUG1 and miR-9-5p, and the luciferase assay confirmed this result. However, the interactions between TUG1 and miRNAs that regulate Dox resistance in BC remains elusive.
We hypothesized that lncTUG1 might bind to miR-9-5p and interact with miR-9-5p, which may be associated with the Dox resistance in BC. We further investigated the regulatory role of the lncTUG1/miR-9-5p interaction in doxorubicin resistance. Our results indicate that lncTUG1 may be a novel therapeutic target for breast cancer.
Materials and Methods
Cell Culture
The BC cell lines MCF-7/ADR, MDA-MB-231, HCC1937, and MCF-7 were obtained from ATCC (Rockville, USA). Cells were cultured in RMPI 1640 medium (Lonza, Switzerland) supplemented with 10% (v/v) fetal bovine serum, 1% penicillin (Gibco, USA), and 1% streptomycin (Gibco, USA). Cells were cultivated in standard conditions. Dox was obtained from Sigma-Aldrich (Merck, Germany), and diluted Dox with dimethyl sulfoxide for later use.
Cell Transfection
2×105 cells were evenly spread on the 6-well plate. After the cells were attached to the wall, Lipofectamine 2000 was mixed with siRNA or inhibitors and then added to the cells cultured in serum-free culture. After 6 hours, they were replaced with normal medium for subsequent experiments. All reagents were from Ribobio (Guangzhou, China), Fulengen (Guangzhou, China), and Thermo Scientific (Waltham, MA, USA).
miR-9-5p mimic: 5ʹ-UCUUUGGUUAUCUAGCUGUAUGA-3ʹ; 5ʹ-AUACAGCUAGAUAACCAAAGAUU-3ʹ; miR-9-5p inhibitor: 5ʹ-UCAUACAGCUAGAUAACCAAAGA-3ʹ; negative control: 5ʹ-CAGUACUUUUGUGUAGUACAA-3ʹ.
The lentiviral particles of shTUG1 were also designed and purchased from GenePharma Co., Ltd. To generate the lentiviruses, shRNA plasmids were co-transfected into MCF7/ADR cells along with envelope (VSVG) and packaging (pGag/Pol, pRev) plasmids using lipofectamine 2000 (Invitrogen). The viral supernatants were harvested and filtered after 48 h transfection. Cells were infected in the presence of a serum-containing medium supplemented with 8 μg/mL polybrene. Following infection for 48 h, cells were selected with 2.0 μg/mL puromycin (Sigma). Knockdown efficiencies were examined by qRT-PCR.
Cell Counting Kit-8 (CCK-8) Assay
1×104 cells were evenly spread on the 96-well plate. After the cells were treated with Dox of different concentrations for 48 h, 10% cck-8 reagent was added to each well, incubated for 1 h, and its absorbance was detected at 450nm. Dox concentrations up to 50% growth inhibition (IC50) were calculated using a dose response curve. The manufacturer of cck8 reagent and microplate reader were Dojindo Laboratories (Tokyo, Japan) and Multiskan Sky (Thermo Fisher Scientific, USA).
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA was extracted from BC cells using TRIzol™ Reagent (Invitrogen, China), and was reverse-transcribed to cDNA using a TaqMan™ miRNA kit or PrimeScript™ RT kit. qRT-PCR assays were tested using a TB SYBR™ Premix Ex Taq™ kit (Cosmo Bio, Boppard, Guangzhou, China). The primers using in this assay were as follows:
eIF5A2: Forward: 5ʹ-TATGCAGTGCTCGGCCTTG-3ʹ;
Reverse: 5ʹ-TTGGAACATCCATGTTGTGAGTAGA-3ʹ;
TUG1: Forward: 5ʹ-GACCGTCCAATGACCTTCCT-3ʹ;
Reverse: 5ʹ-TGGCTGAATGCTTCTTGGGT-3ʹ;
miR-9-5p: 5ʹ-TCTTTGGTTATCTAGCTGTATGA-3ʹ.
EdU Assay
BC cells were seeded in 24-well plates at 1x105 cells per well. Cell proliferation was measured using a Cell LightTM EdU Apollo 567 in vitro kit (Ribobio, China), according to manufacturer instructions. Cell nuclei were stained blue, and EdU-positive cells were green.
Apoptosis Assay
The collected cells were washed with PBS, centrifuged at 400g for 5min, and then resuspended in conjugation buffer. An appropriate amount was added with FITC labeled annexin V and PI (Sigma-Aldrich, Merck, Germany), and incubated at room temperature in dark for 15min, and then tested with CANTO™ II flow cytometer (BD Biosciences), and data were analyzed by FlowJo (Ashland, OR, USA).
Western Blotting
Proteins were extracted from BC cell lines using RIPA lysis buffer (Solarbio, USA). Lysates were isolated by sodium dodecyl sulfate polyacrylamide gels (12%). Gel imprinting was transferred to PVDF Membrane (Millipore, Merck, Germany) by transfer buffer. Membranes were incubated with antibodies recognizing eIF5A2 (1:1000; Abcam, #ab150439, USA) or GAPDH (1:2000; Abcam, #ab181602, USA), then oscillated overnight at 4°C. Next, membranes were incubated with the secondary antibodies of Goat anti-rabbit IgG (Abcam, Cambridge, UK), which was diluted at 1:2000.
Luciferase Activity Assays
For the luciferase assay, BC cells were seeded in 24-well plates at 1.5 x 105 cells per well. Dual-Luciferase Reporter Assay Kit (Promega, USA) was performed to measure the luciferase activity as described previously [1]. Results represent independent experiments performed in triplicate.
Tumor Xenograft Experiments
Male BALB/c nude mice aged 3–4 weeks were purchased from the Experimental Animal Center of Sun Yat-sen University (Guangzhou, China). Cells were harvested and re-suspended in serum free medium at a concentration of 1 × 107 cells/0.2 mL. Each mouse was inoculated subcutaneously in the right flank with MCF7/ADR cells stably transduced with shTUG1 or shControl. Tumor size was monitored every 2 days, and mice were euthanized after 4 weeks. In vivo chemosensitivity assays, the animals were treated with Dox or PBS via tail intravenous injection (2 mg/kg body weight Dox [once every 2 days]).
Statistical Analyses
Statistical analysis was performed using SPSS v18.0 (IBM, Armonk, NY, USA). All data were presented as the mean ± SEM. Two-tailed Student’s t-tests was used to analyze the two groups difference. For multiple group comparisons, one-way ANOVA was used to determine statistically significant differences between samples. Any P-value of P < 0.05 was considered to be statistically significant.
Results
TUG1 is Upregulated in Breast Cancer
We first measured the expression levels of cancer-related lncRNAs in 4 human non-triple-negative BC tissues and peripheral normal tissues. The TUG1 was highly expressed and significantly increased in non-triple-negative BC tissues (Figure 1A). To further verify the result, we explored the levels of TUG1 in additional non-triple-negative breast cancer patients, and found that TUG1 was overexpressed in non-triple-negative BC tissues compared to peritumor tissues (Figure 1B and Supplementary Table 1). We evaluated Dox sensitivity in four BC cell lines (MCF-7/ADR, MDA-MB-231, HCC1937 and MCF-7) using the CCK-8 assay, and found that MCF-7 cells were the most sensitive to Dox, while MCF-7/ADR cells were the least sensitive (Figure 1C). The expression of TUG1 was different in these four BC cell lines, and TUG1 was most highly expressed in MCF-7/ADR cells, demonstrating an association of higher TUG1 expression with lower sensitivity to Dox (Figure 1D and E).
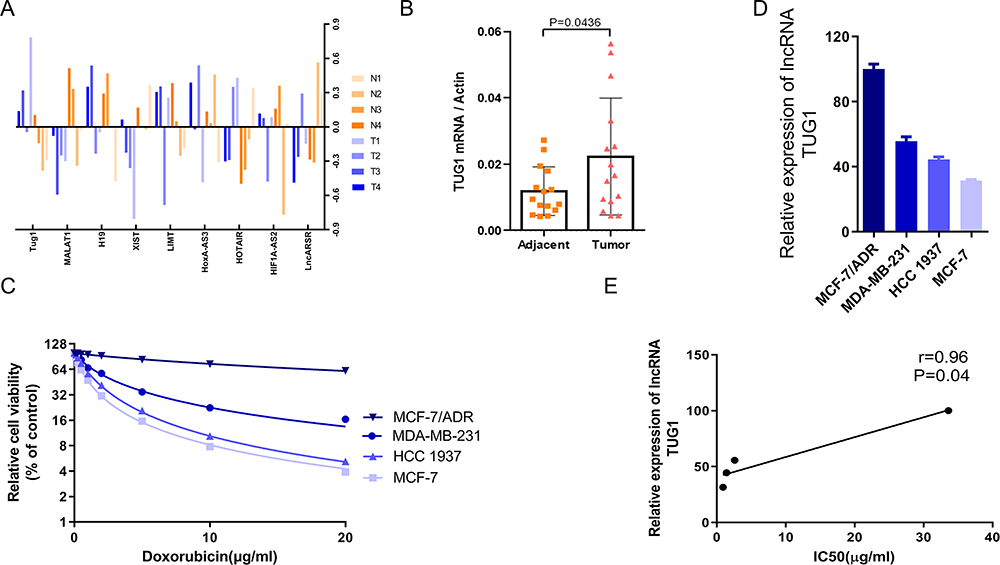 |
Figure 1 TUG1 is upregulated in breast cancer. (A) Real-time PCR analysis was used to determine the expression levels of lncRNAs in 4 human breast cancer tissues and adjacent normal tissues. (B) Validation of the expression of lncRNA TUG1 expression. The lncRNA TUG1 expression levels in breast cancer tissues and adjacent normal tissues (n = 15), as measured by real-time PCR. P = 0.0436 vs the adjacent group. (C) Viability of breast cancer cell lines under different concentrations of Dox, according to the CCK-8 assay. The IC50 of MCF-7/ADR, MDA-MB-231, HCC1937, and MCF-7 were 33.6 μg/mL, 2.6 μg/mL, 1.4 μg/mL, 0.9 μg/mL, respectively. (D) LncRNA TUG1 expression in breast cancer cell lines, as examined by qPCR. (E) The correlation between the relative expression of lncRNA TUG1 and the IC50 of Dox. |
Knockdown of TUG1 Inhibits Dox Resistance in Breast Cancer
We the explored the role of the lncRNA TUG1 as a mediator of breast cancer Dox resistance in vitro. Treatment of the four BC cell lines with Dox (IC50) significantly increased the expression of the lncRNA TUG1 (Figure 2A). To investigate the function of TUG1 in regulating Dox sensitivity in BC cells, we knocked down TUG1 using siRNA. The transfection efficiencies of three si-TUG1 oligos in these four BC cell lines were detected using real-time PCR (Figure 2B). We quantified cell viability in the presence of different concentrations of Dox in BC cells. Suppression of TUG1 inhibited Dox resistance in MCF-7/ADR and MCF-7 cells, but not the triple-negative BC lines of MDA-MB-231 or HCC1937 (Figure 2C and Supplementary Figure 1). The EdU assay performed on MCF-7/ADR and MCF-7 cells confirmed these results (Figure 2D and E). We then investigated whether the growth inhibition was caused by an increase in apoptosis. TUG1 siRNA treatment enhanced apoptosis in MCF-7/ADR and MCF-7 cells, as determined by PI/Annexin V-FITC assay (Figure 2F and G). These data demonstrate that lncTUG1 mediates Dox resistance, and may serve as a potential therapeutic target to overcome Dox resistance and enhance the benefits of Dox therapy in breast cancer.
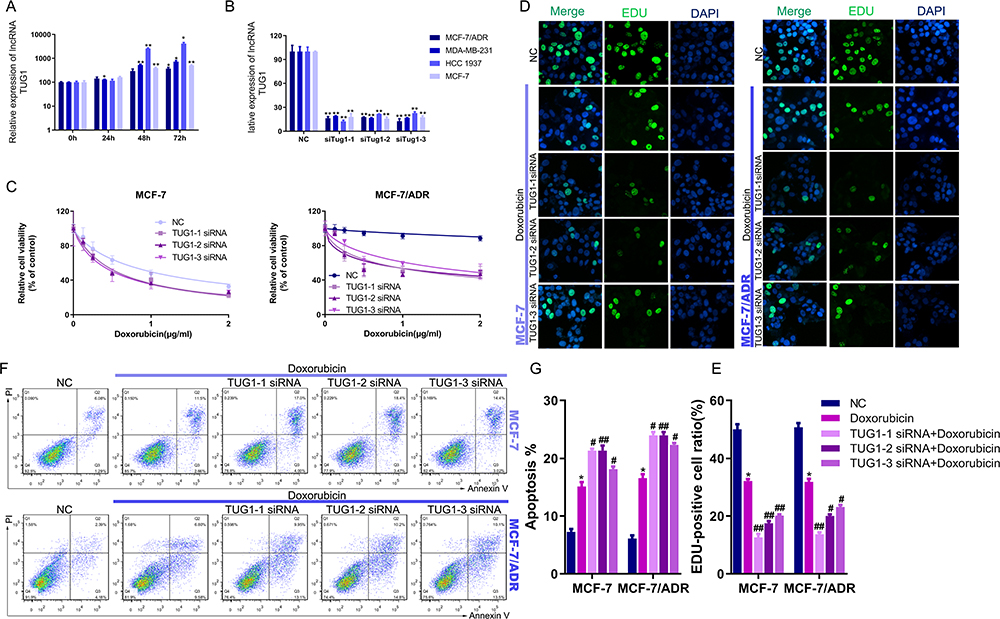 |
Figure 2 Knockdown of TUG1 inhibits Dox resistance in breast cancer. (A) Relative expression of lncRNA TUG1 in breast cancer cell lines after Dox treatment. (B) Transfection efficacies of four si-TUG1 oligos in breast cancer cell lines. (C) The viability of MCF-7 and MCF-7/ADR cells transfected with si-TUG1 or negative control under different concentrations of Dox. (D) Proliferation of MCF-7 and MCF-7/ADR cells transfected with si-TUG1 or negative control and treated with Dox, according to the EdU assay. (E) The number of EdU-positive cells was counted. (F) Apoptosis of MCF-7 and MCF-7/ADR cells transfected with si-TUG1 or negative control and treated with Dox, according to the PI/Annexin V-FITC assay. (G) The quantification of apoptosis ratio of MCF-7 and MCF-7/ADR cells. *P < 0.05, Dox group vs negative control group, #P < 0.05, ##P < 0.01, TUG1 siRNA+ Dox group vs Dox group. |
LncRNA TUG1 Binds miR-9-5p
Having determined the role of lncRNA TUG1 in Dox resistance, we next explored mechanisms of TUG1 regulation by miRNAs. Using Starbase, we found the presence of a consensus-binding site for miR-9-5p in the TUG1 lncRNA. Co-transfection of miR-9-5p mimics with the reporter psiCK-wt-TUG1 was significantly downregulated luciferase activity compared with mutated psiCK-mut TUG1 vector (Figure 3A). Consistent with this finding, miR-9-5p was increased after TUG1 was knocked down in MCF-7/ADR, MDA-MB-231, HCC1937, and MCF-7 breast cancer cell lines (Figure 3B). In addition, knockdown of miR-9-5p in MCF-7 cells promoted Dox resistance, while overexpression of miR-9-5p in resistant MCF-7/ADR cells inhibited Dox resistance (Figure 3C and D). Moreover, cell proliferation was accelerated after miR-9-5p was inhibited in MCF-7 cells. In contrast, the overexpression of miR-9-5p inhibited breast cancer proliferation (Figure 3E). Furthermore, Inhibition or overexpression of miR-9-5p suppressed or promoted apoptosis in MCF-7 or MCF-7/ADR cells treated with Dox (Figure 3F), respectively. Thus, overexpression of miR-9-5p can enhance Dox sensitivity in breast cancer cells.
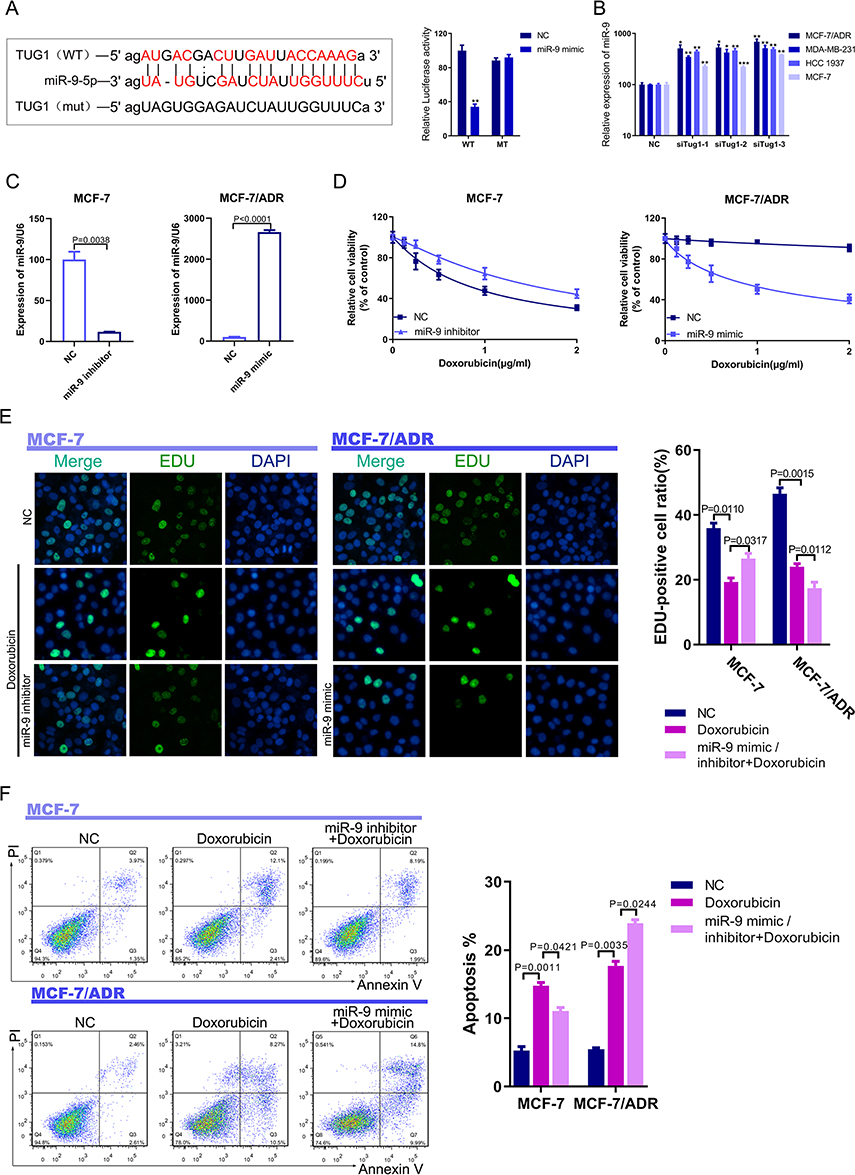 |
Figure 3 LncRNA TUG1 binds miR-9-5p. (A) Starbase predicted binding between lncRNA TUG1 and miR-9-5p. Luciferase activity decreased in the lncRNA TUG1 WT group. (B) LncRNA TUG1 knockdown upregulated miR-9-5p expression in breast cancer cell lines. *P < 0.05, **P < 0.01, ***P < 0.001, vs negative control group. (C) Transfection efficacies of miR-9-5p inhibitor or miR-9-5p mimics in MCF-7 and MCF-7/ADR cells. P = 0.0038, miR-9-5p inhibitor group vs negative control group, P < 0.0001, miR-9-5p mimics group vs negative control group. (D) Viability of MCF-7 or MCF-7/ADR cells transfected with miR-9-5p inhibitor or miR-9-5p mimics, under different concentrations of Dox. (E) Proliferation of MCF-7 or MCF-7/ADR cells transfected with miR-9-5p inhibitor or miR-9-5p mimics and treated with Dox according to the EdU assay. The number of EdU-positive cells was counted. P = 0.0110, negative control group vs Dox group in MCF-7 cell, P = 0.0317, miR-9-5p inhibitor group vs Dox group in MCF-7 cell. P = 0.0015, negative control group vs Dox group in MCF-7/ADR cell, P = 0.0112, miR-9-5p mimics group vs Dox group in MCF-7/ADR cell. (F) Apoptosis of MCF-7 and MCF-7/ADR cells transfected with miR-9-5p inhibitor or miR-9-5p mimics and treated with Dox according to the PI/Annexin V-FITC assay. The quantification of apoptosis in MCF-7 and MCF-7/ADR cells. P = 0.0011, negative control group vs Dox group in MCF-7 cell, P = 0.0421, miR-9-5p inhibitor group vs Dox group in MCF-7 cell. P = 0.0035, negative control group vs Dox group in MCF-7/ADR cell, P = 0.0244, miR-9-5p mimics group vs Dox group in MCF-7/ADR cell. |
TUG1 Inhibits Dox Resistance by Targeting miR-9-5p in vitro
To determine whether the TUG1-mediated functional effects specifically depend on the expression of miR-9-5p, we used a miR-9-5p inhibitor to examine whether the antitumor effect of TUG1 silencing could be blocked by miR-9-5p knockdown. Transfection of breast cancer cells with the miR-9-5p inhibitor significantly inhibited apoptosis induced by TUG1 siRNA (Figure 4A and B). Moreover, miR-9-5p inhibitor significantly blocked the TUG1 knockdown-mediated enhancement of breast cancer cells proliferation (Figure 4C and D). Additionally, the transduction of TUG1 siRNA resulted in enhanced susceptibility of breast cancer cells to Dox; this effect was reversed by pretreatment with the miR-9-5p inhibitor (Figure 4E). TUG1 siRNA and miR-9-5p inhibitor dramatically decreased the expression of TUG1 and miR-9-5p, respectively (Figure 4F and G).
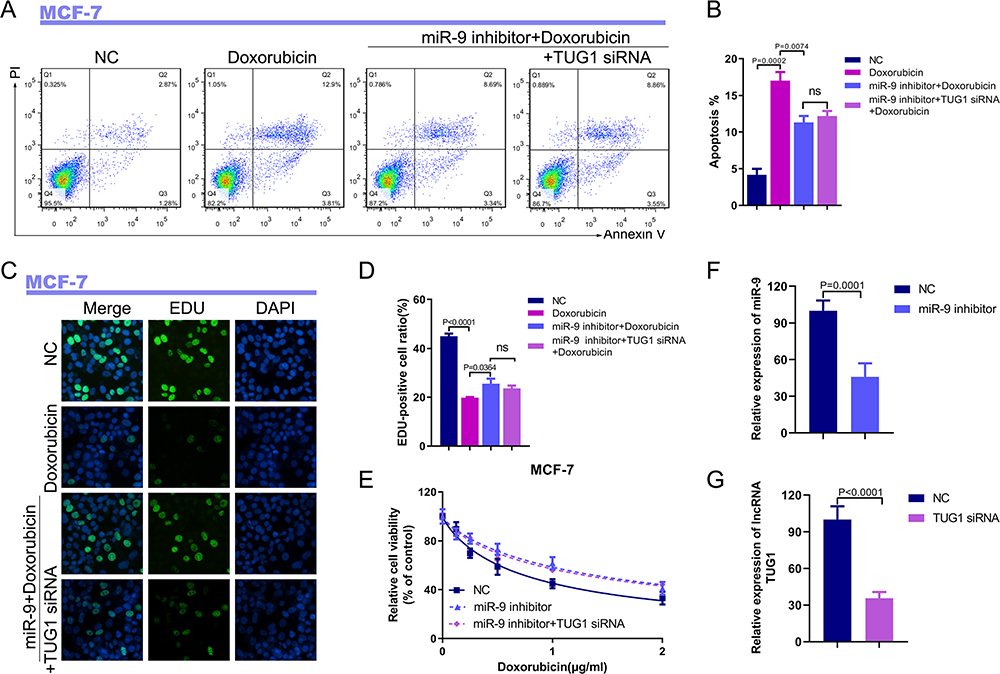 |
Figure 4 TUG1 inhibits Dox resistance by targeting miR-9-5p in vitro. (A) Apoptosis of MCF-7 cell transfected with miR-9-5p inhibitor plus a TUG1 siRNA, or miR-9-5p inhibitor plus a negative control, under different concentrations of Dox according to the PI/Annexin V-FITC assay. (B) The quantification of apoptosis in MCF-7 cells. P = 0.0002, negative control group vs Dox group, P = 0.0074, miR-9-5p inhibitor + Dox group vs Dox group, P > 0.05, miR-9-5p inhibitor + Dox group vs miR-9-5p inhibitor + TUG1 siRNA + Dox group. (C) Proliferation of MCF-7 cells transfected with miR-9-5p inhibitor plus a TUG1 siRNA, or miR-9-5p inhibitor plus a negative control oligo, under different concentrations of Dox, according to the EdU assay. (D) The number of EdU-positive cells was counted. P < 0.0001, negative control group vs Dox group, P = 0.0364, miR-9-5p inhibitor + Dox group vs Dox group, P > 0.05, miR-9-5p inhibitor + Dox group vs miR-9-5p inhibitor + TUG1 siRNA + Dox group. (E) Viability of MCF-7 cell transfected with miR-9-5p inhibitor plus a TUG1 siRNA, or miR-9-5p inhibitor plus a negative control oligo under different concentrations of Dox, according to the CCK-8 assay. (F) Transfection efficacies of the miR-9-5p inhibitor in MCF-7 cells. P = 0.0001, miR-9-5p inhibitor group vs negative control group. (G) Transfection efficacies of the TUG1 siRNA in MCF-7 cells. P < 0.0001, TUG1 siRNA group vs negative control group. |
MiR-9-5p Influences Dox Resistance by Targeting the TUG1 lncRNA in vitro
In order to investigate whether the antitumor effects of miR-9-5p are affected by the sponge activity of the lncRNA TUG1, we examined Dox sensitivity in breast cancer cells with knockdown or overexpression of miR-9-5p after pre-treatment with TUG1 siRNA. The tumor-promoting effects of miR-9-5p silencing could be blocked by TUG1 siRNA, and the antitumor effect of miR-9-5p overexpression could be blocked by TUG1 knockdown (Figure 5A). Consistently, TUG1 siRNA significantly blocked miR-9-5p silencing-mediated proliferation of MCF-7 and MCF-7/ADR cells (Figure 5B and C). Furthermore, transduction of TUG1 siRNA rescued the suppression of apoptosis induced by miR-9-5p inhibitor in MCF-7 and MCF-7/ADR cells (Figure 5D and E). These findings demonstrate that miR-9-5p is involved in lncTUG1-mediated Dox resistance in breast cancer cells.
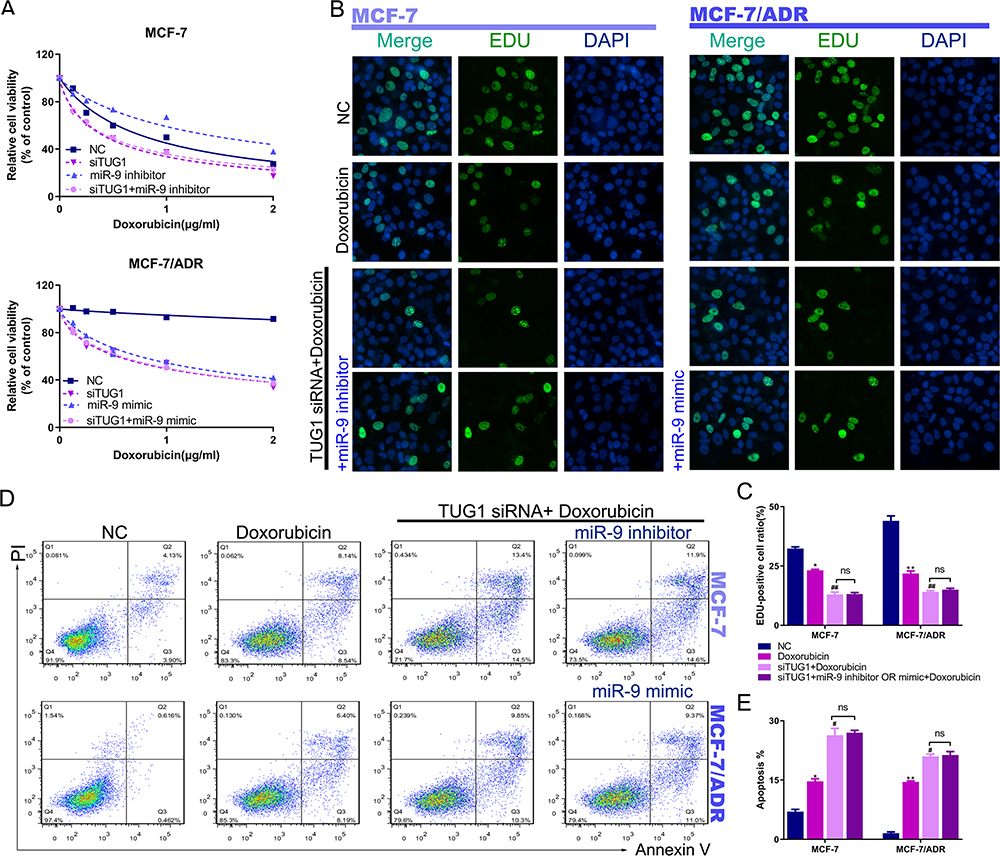 |
Figure 5 MiR-9-5p influences Dox resistance by targeting lncRNA TUG1 in vitro. (A) Transduction of MCF-7 or MCF-7/ADR cells with TUG1 siRNA significantly suppressed miR-9-5p inhibitor-inhibited Dox sensitivity, as determined by CCK-8 assay. (B) Transduction of MCF-7 or MCF-7/ADR cells with TUG1 siRNA significantly suppressed miR-9-5p inhibitor-promoted proliferation, as determined by EdU assay. (C) The number of EdU-positive cells was counted. *P < 0.05, negative control group vs Dox group in MCF-7 cell, ##P < 0.01, TUG1 siRNA + Dox group vs Dox group in MCF-7 cell, P > 0.05, TUG1 siRNA + Dox group vs TUG1 siRNA + miR-9-5p inhibitor + Dox group in MCF-7 cell. **P < 0.01, negative control group vs Dox group in MCF-7/ADR cell, ##P < 0.001, TUG1 siRNA + Dox group vs Dox group in MCF-7/ADR cell, P > 0.05, TUG1 siRNA + Dox group vs TUG1 siRNA + miR-9-5p mimics + Dox group in MCF-7/ADR cell. (D) Transduction of MCF-7 or MCF-7/ADR cells with TUG1 siRNA significantly suppressed miR-9-5p inhibitor-inhibited apoptosis, as determined by PI/Annexin V-FITC assay. (E) The quantification of apoptosis in MCF-7 or MCF-7/ADR cells. *P < 0.05, negative control group vs Dox group in MCF-7 cell, #P < 0.05, TUG1 siRNA + Dox group vs Dox group in MCF-7 cell, P > 0.05, TUG1 siRNA + Dox group vs TUG1 siRNA + miR-9-5p inhibitor + Dox group in MCF-7 cell. **P < 0.01, negative control group vs Dox group in MCF-7/ADR cell, #P < 0.05, TUG1 siRNA + Dox group vs Dox group in MCF-7/ADR cell, P > 0.05, TUG1 siRNA + Dox group vs TUG1 siRNA + miR-9-5p mimics + Dox group in MCF-7/ADR cell. |
EIF5A2 is the Downstream Target of the TUG1 lncRNA
Our previous work demonstrated that eIF5A2 is a direct target of miR-9. Therefore, we examined whether eIF5A2 lies downstream of the lncRNA TUG1 and is involved in regulating Dox resistance. We first examined whether lncRNA TUG1 can regulate the expression of eIF5A2. Real-time PCR and Western blot showed that the expression of eIF5A2 in MCF-7 or MCF-7/ADR cells was downregulated significantly by TUG1 siRNA (Figure 6A and B). CCK-8 assays revealed that eIF5A2 knockdown resulted in much lower viability compared with control treatments, and the effects of TUG1 siRNA were disrupted when eIF5A2 was knocked down (Figure 6C). Treatment with an eIF5A2 siRNA enhanced Dox-induced apoptosis in MCF-7/ADR and MCF-7 cells, and this effect was not seen when TUG1 was knocked down (Figure 6D and E). Our data suggest that the effects of the lncRNA TUG1 in breast cancer cells are mediated by eIF5A2.
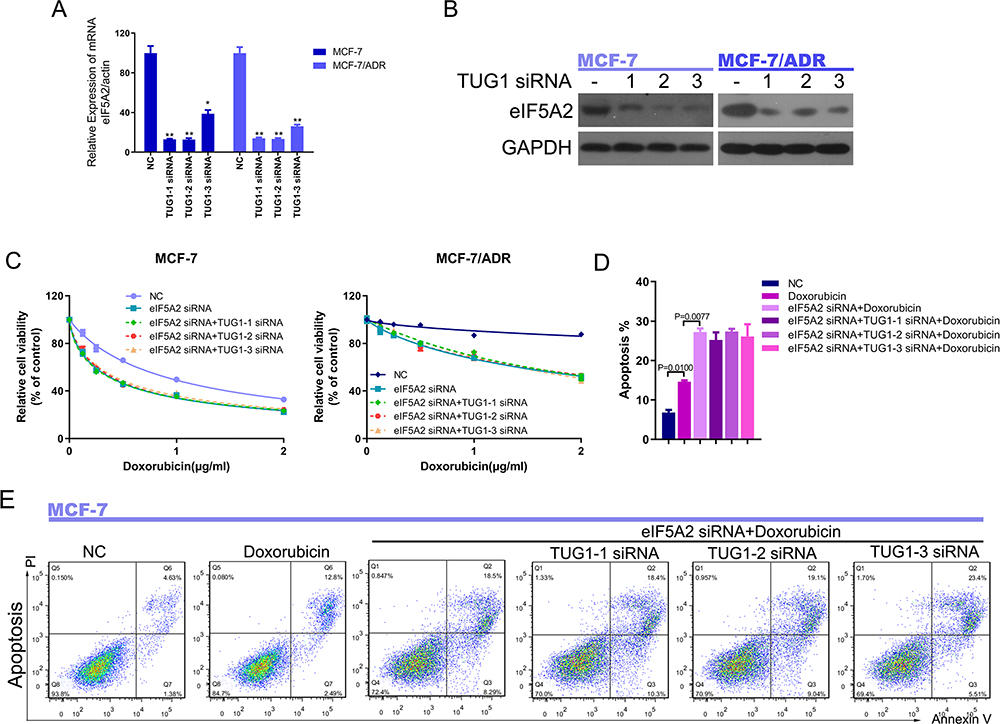 |
Figure 6 EIF5A2 is the downstream target of lncRNA TUG1. (A) LncRNA TUG1 knockdown downregulated eIF5A2 expression in MCF-7 and MCF-7/ADR cells. *P < 0.05, **P < 0.01, vs negative control group. (B) Expression of eIF5A2 protein in MCF-7 and MCF-7/ADR cells transfected with three TUG1 siRNAs or negative control oligo. (C) Viability of MCF-7 and MCF-7/ADR cells transfected with eIF5A2 siRNA plus three TUG1 siRNAs, or eIF5A2 siRNA plus a negative control, under different concentrations of Dox, according to the CCK-8 assay. (D) The quantification of apoptosis in MCF-7 cells. P = 0.0100, negative control group vs Dox group, P = 0.0077, eIF5A2 siRNA + Dox group vs Dox group. (E) Transduction of MCF-7 cells with eIF5A2 siRNA significantly suppressed TUG1 siRNA-promoted apoptosis, as determined by PI/Annexin V-FITC assay. |
TUG1 Regulates Dox Resistance in vivo
To further investigate the impact of TUG1 on Dox resistance, We then used a nude mouse xenograft model to further investigate the ability of TUG1 to confer chemoresistance in Dox. MCF7/ADR cells transfected with shTUG1 or shControl were subcutaneously injected into mice. As shown in Figure 7A, tumor growth was inhibited in the shTUG1 group treated with PBS or drugs (Dox) compared with the controls. Tumor grew significantly more slowly in mice following combined Dox treatment and TUG1 knockdown. Four weeks later, the mean tumor volume for the TUG1-knockdown group and the drugs group was obviously smaller than that of the control group (Figure 7B). Moreover, combined treatment with TUG1 knockdown and drugs led to an even further reduction in tumor volume. Similarly, the average tumor weight in shTUG1 group combined treatment with Dox showed a similar trend (Figure 7C). qRT-PCR analysis of TUG1 expression found it to be significantly lower in tumor tissues formed from shTUG1 group than those from controls (Figure 7D). These results suggested that downregulation of TUG1 increased the in vivo chemosensitivity of BC to Dox.
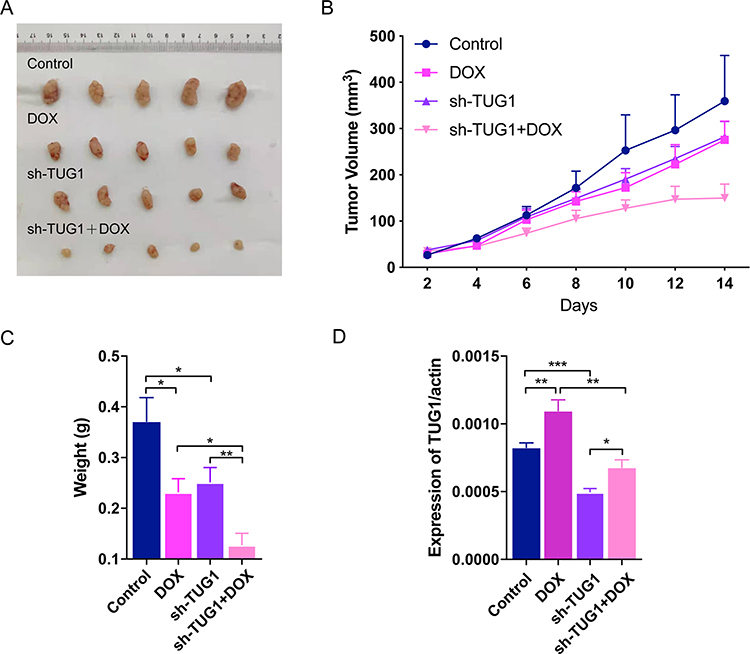 |
Figure 7 TUG1 regulates Dox resistance in vivo. (A) Tumors from all mice in each group (Each group has five mice). MCF7/ADR cells were transduced with shControl or shTUG1 as indicated. After cells (3x107) were injected into mice, Dox or PBS were injected tail intravenously as indicated. (B) Growth curve of tumor volumes. (C) Tumor weights were determined. (D) qRT-PCR was conducted to detect the average expression of TUG1. N=5, *P < 0.05, **P < 0.01, ***P < 0.001. |
Discussion
In this study, we discovered the role of lncTUG1 in regulating Dox sensitivity in BC cells, and we found that TUG1 modulates drug resistance of BC cells through interacting with miR-9-5p. These findings demonstrate that the interaction between the lncRNA TUG1 and miR-9-5p contributes to Dox resistance in BC, and indicate that the lncRNA TUG1 may be a novel therapeutic target in breast cancer.
LncRNAs are involved in many physiological and pathological processes. Here, we found that the lncRNA TUG1 exhibited high expression in clinical breast cancer tissues, suggesting that a correlation exists between lncTUG1 and the pathogenesis of breast cancer. Consistent with our findings, TUG1 has been shown to exert oncogenic effects in osteosarcoma, bladder cancer, esophageal cancer, and small cell lung cancer,15,20,21 whereas it is downregulated in non-small cell lung cancer.22 Tonghuai Li et al revealed that TUG1 promotes the proliferation of ovarian cancer cells through targeting Aurora kinase A (AURKA).23 In addition, a recent study demonstrated that expression of TUG1 is regulated by the Notch signaling pathway, and that TUG1 is highly expressed in GSCs and maintains stemness features of glioma cells.24
Our findings support previous studies on the role of the TUG1 in the regulation of chemoresistance.13,25 TUG1 has been shown to promote Dox-resistance in osteosarcoma by suppressing Akt signaling.26 However, whether TUG1 can regulate Dox sensitivity in breast cancer through miRNAs has not been previously investigated. In the present study, we uncovered the relation between TUG1 and miR-9-5p, confirmed a direct interaction between TUG1 and miR-9-5p, and revealed the role of this interaction in Dox resistance. We found three main indications that show the relationship between miR-9-5p and TUG1; first, we found that the expression of miR-9-5p are notably increased in TUG1 under-expressing BC cells; second, a dual-luciferase assay showed that miR-9-5p reduced the luciferase activity of the wild-type TUG1 vector, but not that of a mutant TUG1 vector; third, knockdown of TUG1 inhibited the effects of miR-9-5p, and miR-9-5p inhibitor treatment blocked the effects of TUG1 siRNA. Our study highlights the important regulatory relationships between miRNAs and lncRNAs in mediating Dox sensitivity in BC.
Our previous work demonstrated that miR-9 rescues daunorubicin resistance by mediating eIF5A2.[1] EIF5A2 is a small universally conserved acidic protein and plays a role in mRNA translation, cellular proliferation, cellular differentiation, and inflammation.27–30 Furthermore, eIF5A2 is a crucial factor in the proliferation, metastasis and aggressiveness of cancer cells.31–33 Previous studies reported that eIF5A2 acts as an oncogene and plays an important role in regulating drug resistance in BC.34–36 Here we report, for the first time, a new regulatory mechanism of eIF5A2 expression. TUG1 knockdown increased Dox sensitivity and TUG1 mediated the regulatory role of eIF5A2 in BC cells. This research proved that eIF5A2, as a downstream target of TUG1, regulates Dox sensitivity in BC, lending credence to our speculation that there is a lncTUG1-miR-9-5p-EIF5A2 axis that can be manipulated to improve the efficacy of Dox in BC. The mechanism by which eIF5A2 regulates Dox resistance may involve induction of epithelial–mesenchymal transition, cytoskeletal rearrangement, angiogenesis, and metabolic reprogramming as reported.29,37–39
Interestingly, we found that inhibition of TUG1 overcame the influences of miR-9-5p-dependent susceptibility of BC cells to Dox. These observations revealed that TUG1 binds to miR-9-5p, and that this interaction regulates the activity of both TUG1 and miR-9-5p. TUG1 functions as a miR-9-5p sponge, and competes for binding of miR-9-5p, directly interfering with the interaction of miR-9-5p and eIF5A2, which blocked miR-9-5p-regulated Dox resistance in BC cells. Furthermore, miR-9-5p may degrade TUG1 after binding, but the regulation of TUG1 by miR-9-5p merits additional study.
Conclusion
In short, our study revealed a previously unappreciated regulatory mechanism by which lncTUG1 mediates Dox resistance in BC, through targeting of the miR-9-5p-EIF5A2 pathway. Furthermore, we emphasized the interaction between lncTUG1 and miR-9-5p, which plays an important role in mediating Dox resistance in breast cancer. Specific blockage of lncTUG1 may be a potential therapeutic avenue to overcome Dox resistance in breast cancer treatment.
Abbreviations
AURKA, Aurora kinase A; EIF5A2, Eukaryotic translation initiation factor 5A-2; LncRNA, Long noncoding RNA; MiRNA, MicroRNA; NcRNAs, Noncoding RNAs; PRC2, Polycomb repressive complex 2; TUG1, Taurine-upregulated gene 1.
Data Sharing Statement
Data sets and materials used and/or analysed during the current study are available in the manuscript itself.
Ethics Approval and Informed Consent
This project was approved by Ethics Committee of Tongde Hospital of Zhejiang province. No informed consent was required because data were going to be analysed anonymously.
Acknowledgments
We sincerely acknowledge members in the Chen laboratory for their technical assistance and helpful advices.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Zhejiang Provincial Nature Science Foundation of China (LR20H160001 and LQ19H160040); Key Project Co-constructed by Zhejiang Province and Ministry (WKJ-ZJ-1916); the National Natural Science Foundation of China (81972693, 81802383, 81972674 and 31900543); the Zhejiang Provincial Traditional Chinese Medicine Science and Technology Project (2020ZZ004); Zhejiang province Public Welfare Projects (No.2017C33089 and LGF18H200001, LGF21H090011, and LGF18H200001).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. DeSantis CE, Ma J, Gaudet MM, et al. Breast cancer statistics, 2019. CA Cancer J Clin. 2019.
2. Young RC, Ozols RF, Myers CE. The anthracycline antineoplastic drugs. N Engl J Med. 1981;305(3):139–153. doi:10.1056/NEJM198107163050305
3. Arnason T, Harkness T. Development, maintenance, and reversal of multiple drug resistance: at the crossroads of TFPI1, ABC transporters, and HIF1. Cancers. 2015;7(4):2063–2082. doi:10.3390/cancers7040877
4. Iyer MK, Niknafs YS, Malik R, et al. The landscape of long noncoding RNAs in the human transcriptome. Nat Genet. 2015;47(3):199–208. doi:10.1038/ng.3192
5. Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26–46. doi:10.1016/j.cell.2013.06.020
6. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
7. Malek E, Jagannathan S, Driscoll JJ. Correlation of long non-coding RNA expression with metastasis, drug resistance and clinical outcome in cancer. Oncotarget. 2014;5(18):8027–8038. doi:10.18632/oncotarget.2469
8. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet. 2008;9(2):102–114. doi:10.1038/nrg2290
9. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
10. Tay Y, Rinn J, Pandolfi PP. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–352. doi:10.1038/nature12986
11. Khalil AM, Guttman M, Huarte M, et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A. 2009;106(28):11667–11672. doi:10.1073/pnas.0904715106
12. Yang L, Lin C, Liu W, et al. ncRNA- and Pc2 methylation-dependent gene relocation between nuclear structures mediates gene activation programs. Cell. 2011;147(4):773–788. doi:10.1016/j.cell.2011.08.054
13. Yu G, Zhou H, Yao W, Meng L, Lang B. lncRNA TUG1 promotes cisplatin resistance by regulating CCND2 via epigenetically silencing miR-194-5p in bladder cancer. Mol Ther Nucleic Acids. 2019;16:257–271. doi:10.1016/j.omtn.2019.02.017
14. Zhou Q, Hu T, Xu Y. Anticancer potential of TUG1 knockdown in cisplatin-resistant osteosarcoma through inhibition of MET/Akt signalling. J Drug Target. 2019:1–8.
15. Niu Y, Ma F, Huang W, et al. Long non-coding RNA TUG1 is involved in cell growth and chemoresistance of small cell lung cancer by regulating LIMK2b via EZH2. Mol Cancer. 2017;16(1):5. doi:10.1186/s12943-016-0575-6
16. Shen M, Dong C, Ruan X, et al. Chemotherapy-induced extracellular vesicle miRNAs promote breast cancer stemness by targeting ONECUT2. Cancer Res. 2019;79(14):3608–3621. doi:10.1158/0008-5472.CAN-18-4055
17. Barbano R, Pasculli B, Rendina M, et al. Stepwise analysis of MIR9 loci identifies miR-9-5p to be involved in Oestrogen regulated pathways in breast cancer patients. Sci Rep. 2017;7:45283.
18. Naorem LD, Muthaiyan M, Venkatesan A. Identification of dysregulated miRNAs in triple negative breast cancer: a meta-analysis approach. J Cell Physiol. 2019;234(7):11768–11779. doi:10.1002/jcp.27839
19. Liu Y, Lei P, Qiao H, et al. miR-9 enhances the chemosensitivity of AML cells to daunorubicin by targeting the EIF5A2/MCL-1 axis. Int J Biol Sci. 2019;15(3):579–586. doi:10.7150/ijbs.29775
20. Ma B, Li M, Zhang L, et al. Upregulation of long non-coding RNA TUG1 correlates with poor prognosis and disease status in osteosarcoma. Tumour Biol. 2016;37(4):4445–4455. doi:10.1007/s13277-015-4301-6
21. Han Y, Liu Y, Gui Y, Cai Z. Long intergenic non-coding RNA TUG1 is overexpressed in urothelial carcinoma of the bladder. J Surg Oncol. 2013;107(5):555–559. doi:10.1002/jso.23264
22. Zhang EB, Yin DD, Sun M, et al. P53-regulated long non-coding RNA TUG1 affects cell proliferation in human non-small cell lung cancer, partly through epigenetically regulating HOXB7 expression. Cell Death Dis. 2014;5:e1243. doi:10.1038/cddis.2014.201
23. Li T, Chen Y, Zhang J, LncRNA LS. TUG1 promotes cells proliferation and inhibits cells apoptosis through regulating AURKA in epithelial ovarian cancer cells. Medicine. 2018;97(36):e12131. doi:10.1097/MD.0000000000012131
24. Katsushima K, Natsume A, Ohka F, et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat Commun. 2016;7:13616. doi:10.1038/ncomms13616
25. Li Q, Song W, Wang J. TUG1 confers Adriamycin resistance in acute myeloid leukemia by epigenetically suppressing miR-34a expression via EZH2. Biomed Pharmacother. 2019;109:1793–1801. doi:10.1016/j.biopha.2018.11.003
26. Hu T, Fei Z, Su H, Xie R, Chen L. Polydatin inhibits proliferation and promotes apoptosis of doxorubicin-resistant osteosarcoma through LncRNA TUG1 mediated suppression of Akt signaling. Toxicol Appl Pharmacol. 2019;371:55–62. doi:10.1016/j.taap.2019.04.005
27. Mathews MB, Hershey JW. The translation factor eIF5A and human cancer. Biochim Biophys Acta. 2015;1849(7):836–844. doi:10.1016/j.bbagrm.2015.05.002
28. Zender L, Xue W, Zuber J, et al. An oncogenomics-based in vivo RNAi screen identifies tumor suppressors in liver cancer. Cell. 2008;135(5):852–864. doi:10.1016/j.cell.2008.09.061
29. Cao TT, Lin SH, Fu L, et al. Eukaryotic translation initiation factor 5A2 promotes metabolic reprogramming in hepatocellular carcinoma cells. Carcinogenesis. 2017;38(1):94–104. doi:10.1093/carcin/bgw119
30. Ganapathi M, Padgett LR, Yamada K, et al. Recessive rare variants in deoxyhypusine synthase, an enzyme involved in the synthesis of hypusine, are associated with a neurodevelopmental disorder. Am J Hum Genet. 2019;104(2):287–298.
31. Chen Z, Yu T, Zhou B, et al. Mg(II)-Catechin nanoparticles delivering siRNA targeting EIF5A2 inhibit bladder cancer cell growth in vitro and in vivo. Biomaterials. 2016;81:125–134. doi:10.1016/j.biomaterials.2015.11.022
32. Zhu W, Cai MY, Tong ZT, et al. Overexpression of EIF5A2 promotes colorectal carcinoma cell aggressiveness by upregulating MTA1 through C-myc to induce epithelial-mesenchymal transition. Gut. 2012;61(4):562–575. doi:10.1136/gutjnl-2011-300207
33. Li Y, Fu L, Li JB, et al. Increased expression of EIF5A2, via hypoxia or gene amplification, contributes to metastasis and angiogenesis of esophageal squamous cell carcinoma. Gastroenterology. 2014;146(7):1701–1713 e1709. doi:10.1053/j.gastro.2014.02.029
34. Liu Y, Du F, Chen W, Yao M, Lv K, Fu P. EIF5A2 is a novel chemoresistance gene in breast cancer. Breast Cancer. 2015;22(6):602–607. doi:10.1007/s12282-014-0526-2
35. Liu J, Wang P, Zhang P, et al. An integrative bioinformatics analysis identified miR-375 as a candidate key regulator of malignant breast cancer. J Appl Genet. 2019;60(3–4):335–346. doi:10.1007/s13353-019-00507-w
36. Post AEM, Bussink J, Sweep F, Span PN. Changes in DNA damage repair gene expression and cell cycle gene expression do not explain radioresistance in tamoxifen-resistant breast cancer. Oncol Res. 2019.
37. Liu Y, Liu R, Fu P, et al. N1-Guanyl-1,7-diaminoheptane sensitizes estrogen receptor negative breast cancer cells to doxorubicin by preventing epithelial-mesenchymal transition through inhibition of eukaryotic translation initiation factor 5A2 activation. Cell Physiol Biochem. 2015;36(6):2494–2503. doi:10.1159/000430209
38. Tang DJ, Dong SS, Ma NF, et al. Overexpression of eukaryotic initiation factor 5A2 enhances cell motility and promotes tumor metastasis in hepatocellular carcinoma. Hepatology. 2010;51(4):1255–1263. doi:10.1002/hep.23451
39. Li S, Ma Y, Xie C, et al. EphA6 promotes angiogenesis and prostate cancer metastasis and is associated with human prostate cancer progression. Oncotarget. 2015;6(26):22587–22597. doi:10.18632/oncotarget.4088
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.