Back to Journals » Drug Design, Development and Therapy » Volume 16
Immunotherapy and CRISPR Cas Systems: Potential Cure of COVID-19?
Received 1 November 2021
Accepted for publication 16 March 2022
Published 30 March 2022 Volume 2022:16 Pages 951—972
DOI https://doi.org/10.2147/DDDT.S347297
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Tin Wui Wong
Xuesong He,1 Xiao Xue Zeng2
1Department of Cardiology, Changzhou Jintan First People’s Hospital, Changzhou City, Jiangsu Province, 213200, People’s Republic of China; 2Department of Health Management, Centre of General Practice, The Seventh Affiliated Hospital, Southern Medical University, Foshan City, Guangdong Province, 528000, People’s Republic of China
Correspondence: Xiao Xue Zeng, Department of Health Management, Centre of General Practice, The Seventh Affiliated Hospital, Southern Medical University, No. 28, Desheng Road Section, Liguan Road, Lishui Town, Nanhai District, Foshan City, Guangdong Province, 528000, People’s Republic of China, Email [email protected]
Abstract: The COVID-19 has plunged the world into a pandemic that affected millions. The continually emerging new variants of concern raise the question as to whether the existing vaccines will continue to provide sufficient protection for individuals from SARS-CoV-2 during natural infection. This narrative review aims to briefly outline various immunotherapeutic options and discuss the potential of clustered regularly interspaced short palindromic repeat (CRISPR Cas system technology against COVID-19 treatment as specific cure. As the development of vaccine, convalescent plasma, neutralizing antibodies are based on the understanding of human immune responses against SARS-CoV-2, boosting human body immune responses in case of SARS-CoV-2 infection, immunotherapeutics seem feasible as specific cure against COVID-19 if the present challenges are overcome. In cell based therapeutics, apart from the high costs, risks and side effects, there are technical problems such as the production of sufficient potent immune cells and antibodies under limited time to treat the COVID-19 patients in mild conditions prior to progression into a more severe case. The CRISPR Cas technology could be utilized to refine the specificity and safety of CAR-T cells, CAR-NK cells and neutralizing antibodies against SARS-CoV-2 during various stages of the COVID-19 disease progression in infected individuals. Moreover, CRISPR Cas technology are proposed in hypotheses to degrade the viral RNA in order to terminate the infection caused by SARS-CoV-2. Thus personalized cocktails of immunotherapeutics and CRISPR Cas systems against COVID-19 as a strategy might prevent further disease progression and circumvent immunity escape.
Keywords: COVID-19, SARS-CoV-2, immunotherapy, CRISPR Cas
Introduction
Since December 2019, a novel coronavirus name of “SARS‐CoV‐2”, caused the outbreak of COVID‐19 pandemic and scientists from all over the world have been racing to find a cure for COVID-19.1 The SARS-CoV-2 virus bears strong similarity to SARS-CoV and mainly enter cells by utilizing its spike(S) glycoproteins which is complementary to the angiotensin converting enzyme 2 (ACE-2) receptors in the epithelial cells of the human lungs.2 SARS-CoV-2 hijacks the human body cells and releases its RNA genome into the cytoplasm, and then initiates the replication of its viral RNA to generate a new virus to infect other healthy cells.3 Transmission dynamics include two categories, one is the transmission between humans and the other is from air pollution to human which is also the airborne viral infectivity. Pollution increase the diffusion rate of viral infectivity which might have stronger infectivity than human-to-human transmission. Industrialized regions with high levels of air pollution have a significant impact on higher infection and mortality rate.4–8 A recent systematic review suggests that some ethnic factors might also influence COVID-19 outcome.9 Non-pharmaceutical interventions such as wearing masks, hand hygiene, disinfection of surfaces, social distancing and self isolation are educated to the general public to slow down or decrease the spreading of COVID-19.10,11 Countries implemented lockdown policies, public event cancellations, placing restrictions on private gatherings, closing schools and workplaces, quarantine or extensive testing in order to curb COVID-19 transmissions, and additionally the real-time monitoring of transmissibility mortality is vital.12–14 Moreover, speedy optimal vaccination rollout for the population is a useful strategy to decrease infection rate and mortality before the take off of a next pandemic wave.15 The factors of a relative better preparedness of countries to cope with COVID-19 pandemic crisis could be associated with a relative smaller population or better governance and higher expenditures in health systems. Experience drawn from the comparison between first and second wave of the COVID-19 pandemic in Italy can promote effective policy responses to decrease the negative impact on people’s health in recurring waves and in potential future pandemics.16–20 Conventional COVID-19 management include supportive care, eg oxygenation, and fluid management with are provided widely for mild and more severe cases. In severe COVID-19 cases, patients with ARDs are put under ventilation or endotracheal intubation, or extracorporeal membrane oxygenation (ECMO) according to circumstances and availability.21 The antiviral remdisivir was approved for emergency use by US Food and Drug Administration (FDA) on 8th May 2020. Moreover, a study in Italy concludes that the use of remdesivir was associated with positive effect on the survival of severe patients under mechanical ventilation.22 However, in March 2020, WHO coordinated a WHO solidarity trial on remdesivir, hydrochloroquine, lopinavir and interferon-β1a.2, concluding that they have little or no effect on COVID-19 inpatients on death rate, ventilation or on the length of hospitalization days.23 Tumour necrosis factor regulator 2 is proposed as tregs-based therapy in the management of COVID-19 to overcome hyperinflammation in severe cases of COVID-19.24 Baricitinib as an immunomodulatory treatment is suggested as an approach to lower mortality rate.25 Then, dexamethasone used intravenously with standard care was concluded to have effects in shortening ventilator days in COVID-19 patients with moderate to severe ARDS.26 However a study revealed that the there was no synergistic effect between tocilizumab and dexamethasone on the hospital mortality.27 Despite the implementation of preventative measures, supportive care, and various therapeutic approaches to contain COVID-19,28 the SARS-CoV-2 associated infection nevertheless occur, therefore alternative approaches should be taken into account. Thus this review aims to briefly outline various immune response boosting strategies and discuss the immunotherapeutics and CRISPR Cas technology as potential specific cure for COVID-19 and the challenges to be overcome.
Methods
We searched the PubMed for relevant literature in subject area including COVID-19 vaccine, convalescent plasma, antibody therapy, T cell based therapy, CAR-T cell therapy, CAR-NK cell therapy and CRISPR Cas technology against COVID-19. The search terms included “COVID-19 vaccine”, “Vaccine against COVID-19 variants”, “convalescent plasma COVID-19”, “neutralizing antibodies against COVID-19”, “T cell COVID-19”, “CAR-T COVID-19”, “CAR-NK COVID-19”, “CRISPR Cas COVID-19”. 645 full text articles were read and assessed for elegibility. The final choice of literature and references included, among which 84 are identified via reference search or other sources, are based on our judgement of the relevance of their contents in providing a review mainly focused on the potential of immunotherapeutics and CRISPR Cas technology against the complex disease of COVID-19 and a very brief outline of COVID-19 targeting prophylactic vaccine and immunotherapeutic aspects of COVID-19. We have also stated some of our opinions formed based on the literature search and reading. As for the tables we searched the www.clinicaltrials.gov for neutralizing antibodies, T cell, CAR-T cell, NK cell, CAR-NK cell against COVID-19. See Figure 1.
 |
Figure 1 Strategies to tackle SARS-CoV-2. |
COVID-19 Outcome
Some patients suffering from COVID-19 merely experienced mild symptoms and recovered without hospitalization treatment29 whereas others experienced more severe symptoms which required hospitalization, or developed into critically ill cases and were admitted into ICU. People could also die from COVID-19.30,31 Risk factors for patients susceptible to critically ill COVID-19 cases could be old age, male gender, obesity, pregnancy, immune deficiencies, chronic illnesses such as hypertension, diabetes, chronic diseases of the lung, heart, kidney or liver or even tumors.32 In a survey it is concluded that even the majority of patients who were discharged six months after COVID-19 hospitalization still felt that their health have not returned to their normal state before infection and the most of them suffered from fatigue, with 9% of them still requiring oxygen therapy.33
Immune Responses in COVID-19 Patients
Development of vaccines and immune-based therapeutics against SARS-CoV-2 are based on the understanding of the immunopathogenic basis of human immune responses against SARS-CoV-2.34 Spike specific CD19+IgG+ memory B cells were detected in the blood samples of eight COVID-19 patients around the time of 9 to 28 days since the onset of symptoms. Monoclonal antibodies were separated from single B cells from 8 COVID-19 patients, some with the capacity to neutralize SARS-CoV-2.35 Studies indicate lymphopenia of T lymphocytes, with drastically decreased amount of CD4+ T cells and CD8+ T cells in moderate and severe cases of COVID-19.36 A study showed that severe cases of COVID-19 have decreased numbers of total lymphocytes, CD4+ T cells, CD8+ T cells, B cells, and natural killer cells in their blood plasma than mild cases.37 In recovered patients from mild COVID-19, reactive specific N, M, and S protein targeting T cells were detected with IFN-γ ELISPOT.38 Follicular helper T cells and Th2 CD4+ T cells both promote for B cell antibody generation.39 Bcl-6+ follicular helper cells and germinal centers has been reported to be missing in COVID-19 patients.40
Impact of SARS-CoV-2 Mutations
During the COVID-19 pandemic SARS-CoV-2 has undergone evolution developing into variants such as B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma) and B.1.617 divided into B.1.617.1 or (Kappa), and B.1.617.2 (Delta) with mutations on the N-terminal domain or receptor binding domain of the spike protein.41 Mutations occur in viruses such as modifications in genetic sequences that could lead to higher infectivity, or more efficient replication. Examples such as evolved S proteins, include B.1.1.7 (Alpha variant), B.1.351 (Beta variant), and B.1.1.28 (Gamma variant). In B.1.1.7 four deletions occurred of the spike protein such as N501Y, P681H, H69–V70, and Y144/145. B.1.351 include three mutations in the spike protein such as N501Y, K417N, and E484K.42 B.1.1.28 included N501Y, K417T, and E484K mutations in the gene coding spike protein and the P.1 include mutations such as 20J/501Y.V3. 20J/501Y.V3. In addition, B.1.617 include E484Q and L452R mutations. It is notable that the N439K mutation raises the binding affinity of the spike protein of SARS-CoV-2 for the ACE2 receptor and decreases the effect some monoclonal antibodies and polyclonal antibodies against the infection in the sera of patients recovered from COVID-19.43,44 B.1.1.529 (Omicron) included 30 signature mutations, and shares some mutations with the previous variants of concern mentioned.45 It is possible that future mutations could pose challenges to the development of effective prophylactics and therapeutics.
Vaccines
Pfizer (BNT162b2), Moderna (mRNA-1273) are RNA based vaccines approved by the Food and Drug Administration (FDA). Adenovirus vaccines include AZD1222 from AstraZeneca/Oxford, Ad26.COV2.S from Johnson & Johnson/Janssen, Ad5-nCoV from iCanSino Biologics. Moreover, some companies used the typical vaccine platform to make inactivated vaccines which include BBIBP-CorV from the Beijing Institute of Biological Products/Sinopharm, CoronaVac from the Wuhan Institute of Biological Products/Sinopharm, and BBV152 from Bharat Biotech in India.46 Pfizer-BioNTech and Moderna were able to gain the first FDA approval for modified mRNA-based vaccines under Emergency Use Authorization (EUA).The synthetic modified mRNA has been already approved as a vaccination-based method for targeting SARS-CoV-2 virus as mRNA does not enter the nucleus or integrate into the host’s genome and it does not cause mutagenicity, which is the major disadvantage of the DNA-based delivery systems.47 BNT162b2 is a Lipid nanoparticle formulated mRNA COVID-19 vaccine, comprised of two doses (Pfizer-BioNTech, Mainz, Germany) which was applied after its approval by the authorities. The purpose of the lipid nanoparticles are to prevent the mRNA from breaking down before it reaches the human body cells after vaccination. The mRNA is translated into S protein inside the host cells causing immune responses, inducing antibody and CD4+ and CD8+ T cell responses. BNT162b2 is reported to be capable of preventing symptomatic manifestation of SARS-CoV-2 infections after the completion of two doses.48–53 However, in a study, obvious antibody decline was reported after 3 months post BNT162b2 vaccination but did not vanish entirely,54 but immunocompromised patients respond less well to COVID-19 vaccine despite its protective purposes.55,56 In a cohort study, three patients who underwent dialysis contracted COVID-19 after the first dose of vaccine, then one patient deceased, indicating the necessity of intervention post vaccinations, the monitoring of SARS-CoV-2-S1 IgA in blood serum of such patients.57 In Goes et al Six subjects in Brazil were vaccinated with either one or two doses of CoronaVac or AZD1222 before suffering from mild symptoms of COVID-19, for some of them it was even a reinfection, suggesting the possibility of infection irrespective of vaccine category and completion of vaccine doses.58 COVID-19 vaccine may lower the possibility of infection upon exposure to the virus or might decrease the possibility of infection symptoms leading to asymptomatic infection with viral shedding, still contagious, but probably lower the contagiousness of SARS-CoV-2 infected people. However, if the vaccine does not decrease contagiousness merely causing asymptomatic infections, it might potentially lead to increased transmission of SARS-CoV-2 since asymptomatic infections of COVID-19 are more difficult to detect.59 Moreover, SARS-CoV-2 variants could also influence vaccine efficacy. Although BNT162b2 mRNA vaccine is an effective prophylactic against SARS-CoV-2, a cohort study of BNT162b2 assessed 813 viral genome sequences from nasopharyngeal swabs suggesting lowered vaccine effectiveness against B.1.351 and B.1.1.7. A greater proportion of B.1.351 was detected in the group vaccinated with BNT162b2, in 7–14 days after the second dose, in comparison to the matched unvaccinated controls. Moreover, an increased proportion of B.1.1.7 was found in individuals between 14 days after the first dose and 6 days after the second dose, in comparison to the matched unvaccinated control, however there was no increased breakthrough rate of B.1.1.7 were detected a week or after the second dose, but not enough data were available to assess vaccine immune escape of B.1.351 in the one dose group.60 However, a recent report shows that BNT162b2 mRNA vaccines induced neutralizing antibodies against the SARS-CoV-2 virus, and also against 5 variants of concern Alpha (B.1.1.7) Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Kappa (B.1.617.1) at a week post vaccination but dropped in levels at 12 weeks and six months, but the level of spike antibodies were detected six months after the second dose are the close to those after receiving the first dose and from the convalescent plasma of people infected by COVID-19.61 Moreover, The BNT162b2 mRNA COVID-19 vaccine induced spike protein-specific neutralizing antibodies for the B.1.1.7 variant, although the neutralization for the B.1.351 variant is reduced but they still provide some level of protection.62 In another study the mRNA-1273 vaccine elicit high neutralizing responses against B.1.1.7 and B.1.351 infections, whether symptomatic or asymptomatic, providing protection against mortality even after a single dose.63 A study showed that a two-dose regimen of the AZD1222 from AstraZeneca/Oxford (ChAdOx1 nCoV-19) vaccine did not show sufficient efficacy in preventing against mild-to-moderate COVID-19 caused the B.1.351 variant. (ClinicalTrials.gov number, NCT04444674).64 A cohort study of COVID-19 immune responses in 585 patients with cancer after two doses of BNT162b2 (153 out of 585) or AZD1222 (430 out of 585) vaccines. Seroconversion rates followed by two doses were 85% and 59% in patients with solid (447 out of 585) and hematological cancer (138 out of 585) although 29 out of 585 did not receive a second dose of vaccine due to various reasons. Post-first dose, 49% of patients with no infection had detectable neutralizing antibodies to wildtype SARS-CoV-2, with significantly lower proportion having detectable NAb to variants of concerns (Alpha 15%, Beta 9%, Delta 9%). Post-second dose, the proportion of patients with detectable neutalizing antibodies against all strains increased, but less so against variants of concern versus wild type (wild type 83%, Alpha 61%, Beta 53%, Delta 54%).65 A 1.617.2 live virus was used in a study to analyze its sensitivity to vaccine induced serum neutralizing antibodies post two doses of AZD1222 or BNT162b2 in vitro, which demonstrated diminished neutralizing effect against B.1.617.2 (Delta variant) relative to wild type Wuhan-1 bearing D614G of approximately an eightfold.66 A study researched on the immune response of twenty Ad26.COV2.S vaccinated individuals, analyzing the vaccine induced neutralizing effect against the original SARS-CoV-2 virus WA1/2020 as well as against the B.1.1.7, CAL.20C, P.1 and B.1.351 variants of concern, suggesting reduced neutralizing effect when targeting the B.1.351 and P.1 variants, although other antibody responses and T cell responses were preserved, moreover natural killer cell responses were preserved against the B.1.351 variant.67 In another study and serum from 50 individuals in 2 to 3 weeks after receiving of the second dose vaccines — BBIBP-CorV (Sinopharm) or CoronaVac (Sinovac), which showed a low antibody response. In the CoronaVac vaccinee serum samples, there was a remarkable decrease in the serum neutralization of B.1.1.7 and B.1.351.68 While a single dose of BBIBP-CorV (Sinopharm)or CoronaVac (Sinovac) do not generate sufficient protection against the B.1.617.2 variant among study participants between 19 and 59 years, post two doses the neutralizing activity reached an vaccine efficacy of 59% in a study, which surpassed the World Health Organization minimal threshold of 50%.69 Moreover in a study it was suggested that the vaccine BBV152/COVAXIN effectively neutralizes recently emerged B.1.1.7 variant of SARS-CoV-2. BBV152 has equivalent neutalizing antibody titres to B.1.1.7,70 However, BBV152 induced neutralisation activity against B.1.351 variant diminished but demonstrated effectiveness in protection against the B.1.617.2 variant.71 SARS-CoV-2 Omicron variant (BA.1/B.1.1.529) has up to 36 mutations in the spike protein. A study show that Omicron variant escapes neutralizing antibodies elicited by BNT162b2 or Coronavac. A group measured the neutralization potency of sera from 88 mRNA-1273, 111 BNT162b, and 40 Ad26.COV2.S vaccinees against wild-type, B.1.617.2, and Omicron SARS-CoV-2 pseudoviruses. However, results show that the participants who received additional booster doses of mRNA vaccines presented vaccine induced potent antibody neutralization of Omicron, merely 4 to 6-fold lower than wild type.72,73 In another study, six months after the 2nd dose the BNT162b2 and mRNA-1273 vaccines, there was no neutralizing activity against the Omicron variant detected in the sera of the study participants, but after a booster shot (3rd dose), neutralizing responses was detected in over 90% of the participants against the omicron variant.74,75 More detail of vaccines against SARS-CoV-2 variants are beyond the scope of this paper and reviewed else elsewhere.76 Alternative strategies include vaccines not merely designed to target the spike protein of SARS-CoV-2 virus to prevent immune escape of the future novel variants. A hAd5 S-Fusion+N-ETSD vaccine was generated to induce T cells and antibodies that target both S protein and N protein of SARS-CoV-2 virus in mice in an effort to overcome the shortcomings of vaccine that merely elicit immunity against spike protein as T cells play a role in long term immunity in inducing antibody responses and in the annihilation of SARS-CoV- 2 infected cells.77
Convalescent Plasma Therapy
Monoclonal antibodies extracted and developed from convalescent plasma show the capacity to deter progression of mild to severe COVID-19 and can be suggested as a therapeutic during the early stage of COVID-19.78 A trial of applying convalescent plasma in treating COVID-19 came to the conclusion that this approach is not effective in preventing the mild cases from developing into serious cases, however due to the limitations in the trial this conclusion has yet to be confirmed by more researches.79 In another study in China the results could lead to the conclusion that convalescent plasma could relieve the symptoms and lower mortality in severe cases of COVID-19.80 One concern is whether convalescent plasma is a safe enough method to apply. In a case study the results suggested that the transfer of convalescent plasma is even safe enough with a SARS-CoV-2 infected patient with immunoglobulin G(IgG)K multiple myeloma although she was treated for her disease.81 Another multicenter cohort study concluded that convalescent plasma can reduce in-hospital mortality of COVID-19 patients.82 The risk of mortality is decreased with patients hospitalized with COVID-19 who did not undergo mechanical ventilation by the transfusion of convalescent plasma with higher anti-SARS-CoV-2 IgG antibody levels rather than transfusion of plasma with lower antibody levels. ClinicalTrials.gov number, NCT04338360.83 In a randomized trial, hospitalized adult patients with severe COVID-19 pneumonia in a 2:1 ratio to receive convalescent plasma or placebo, but the outcome showed no difference between convalescent plasma and placebo, PlasmAr ClinicalTrials.gov number, NCT04383535.84 Moreover, in Kemp et al an immunocompromised patient who had B cell lymphoma underwent convalescent plasma therapy for COVID-19 and the results indicated evolution of the culprit virus producing spike double mutation with both ΔH69/ΔV70 and D796H which enabled SARS-CoV-2 to evade neutralizing antibodies with increased infectivity, suggests an evaluation on the suitability of the individual’s health condition before convalescent plasma therapy as a lack of support of T cell and B cell for the antibodies administered result in reduction of viral clearance could potentially lead to viral evolution and immune escape.85 One individual transfusion of convalescent plasma was ceased after 2–3 mL were administered, due to diffuse erythema and nausea.86 A therapeutic strategy based on the level of antibodies against the SARS-CoV-2 virus, in which patients with positive serology receive high-dose dexamethasone and/or tocilizumab and patients with negative serology receive early convalescent plasma therapy that suggest that early treatment of convalescent plasma and high-dose dexamethasone may benefit patients with SARS-CoV-2 infection by reducing the rate of lung damage or intensive care unit transfer rate. However, T cell depletion occurred in patients who received high-dose dexamethasone, and B cell depletion occurred in those who received Tocilizumab.87 A study documented in vivo microevolution of SARS-CoV-2 observed by analyzing SARS-CoV-2 genetic sequence derived from tracheal aspirates from day 0, day 7, day 21, and day 27 of a patient infected by B.1.369 with immunodeficiencies who underwent convalescent plasma therapy. 5 mutations were detected in B.1.369 over 27 days demonstrating that previously undetected variants of concern can appear in immunocompromised patient despite the intervention against COVID-19 provided.88 According to Wang et al, convalescent plasma from patients infected with SARS-CoV-2 earlier in the pandemic demonstrate no particular difference in its ability to neutralize B.1.1.7, but its neutralizing capacity against B.1.351 decreased. Also there was a decrease in neutralizing activity against P.1, but less compromised than with B.1.315. The convalescent plasma were collected from 20 of SARS-CoV-2 patients infected during the spring of 2020 and showed a 3.4-fold loss of activity against authentic P.1 virus variant.89 A research compared the neutralization of B.1.351 using plasma drawn from hospitalized COVID-19 patients during the two waves of infection in South Africa, the second wave dominated by the B.1.351 variant. The first-wave virus variant could be neutralized by using the plasma of the first-wave infected donors, but the B.1.351 variant was poorly cross-neutralized by plasma from individuals with first-wave infections, the efficacy was reduced by 15.1-fold relative to neutralization of B.1.351 by plasma from individuals infected in the second wave. However, cross-neutralization of first-wave virus variants using plasma from donors of second-wave infections demonstrated merely a 2.3-fold decrease in comparison to neutralization of first-wave virus variants by plasma from donors infected in the first wave.90 Since humans generate antibodies against the viral species with which they are infected, and these antibodies are found in convalescent plasma. As any individual who recovers from variant SARS-CoV-2 has antibodies against that variant, convalescent plasma is an antibody-based therapy that locally keeps up with variants.91
SARS-CoV-2 Neutralizing Antibodies
In a research report, 206 receptor domain specific monoclonal antibodies were obtained from single B cells from 8 COVID-19 patients. Human antibodies that can neutralize SARS-CoV-2 are found which with competes with angiotensin-converting enzyme 2 (ACE2) in binding to receptor-binding domain (RBD), thus impair viral entry and these antibodies might bring advantage developing antibody therapies for COVID-19.92 Researches on monoclonal antibodies for neutralization include tocilizumab, sarilumab, and siltuximab.93 Additionally the use of a combination of some single monoclonal antibodies, so called a “cocktail of antibodies” instead of one type of antibody, with other antibodies acting as backup antibodies in case the spike proteins should escape notice. Trials were run in monkeys although we have wait for more evidence about effectiveness.94 In a study, SARS-CoV-2 neutralizing antibodies are detected. The majority of the antibodies target the spike protein—the receptor-binding domain and N-terminal domain (NTD), antibodies directed against the receptor binding domain (RBD) are 10 −100 times more effective than antibodies that target the N-terminal domain. The neutralizing antibodies can be classified into four groups, group one include S1 RBD-specific neutralizing antibodies (J08, I14, F05, G12, C14, and B07), group two consists of S1-specific neutralizing antibodies that did not bind the RBD (I21, J13, and D14).The third group consists of antibodies able to bind to the S-protein only in its entire trimeric form (H20, I15, F10, and F20), the fourth group consists of antibodies that are specific to S2 domain.95,96 Although antibody therapy is used as a immune activity based therapeutic with COVID-19, a combination of IgG monoclonal antibodies against the receptor-binding domain of the spike protein, Immunoglobulin A dimers and Immunoglobulin M pentamers also seem to demonstrate favorable preliminary treatment results.97 The FDA provided authorization for emergency use for two monoclonal antibody cocktails, one consisting of casirivimab and imdevimab (REGEN-COV), and the other is made up of bamlanivimab and etesevimab. Bamlanivimab is authorized as a prophylactic measure for those in risk after exposure to SARS-CoV-2.98 A clinical trial of 275 of COVID-19 outpatients with symptoms put into groups of those who received 2.4g or 8g of a antibody cocktail that consists of Casirivimab and Imdevimab and those who received placebo. The results demonstrate that the antibody cocktail decreased the viral load.99 Furthermore another study evaluation of findings from over 6500 outpatient with monoclonal antibodies (mAbs) including bamlanivimab/etesevimab, and casirivimab/imdevimab led to the conclusion that they protect individuals against mild to moderate COVID-19, inhibits further progression of the disease in most cases.100 A study showed that Imdevimab neutralized B.1.351 lineage pseudoviruses containing N501Y, K417N, and E484K mutations, while Casirivimab showed partial neutralizing effect against them.101 In another study it was concluded that bamlanivimab, casirivimab, and imdevimab possess the capacity to effectively neutralize authentic SARS-CoV-2, including variant B.1.1.7 (Alpha), however variants B.1.351 (Beta) and P.2 (Zeta) were resistant to bamlanivimab and partly resistant to casirivimab as they carry the E484K mutation. The capacity of NTD-specific monoclonal antibody 4A8 targeting effect decreased with the B.1.351 variant in South Africa that includes deletion of NTD residues 242–244.102 Antibody cocktails that consists of S2M11, S2E12, or S309 demonstrated neutralizing capacities against a panel of circulating SARS-CoV-2 and produced effector functions.103 An investigation into potent antibodies targeting the ACE-2 receptor found that they usually possess poor breadth and are easily escaped by SARS-CoV-2 variants but the S2E128 antibody demonstrated properties to circumvent viral escape.104 In a study researchers used neutralization assays against P.1 including 20 antibodies such as S309 Vir AstraZeneca (AZD) 8895, AZD1061, and AZD7442 (a mixture of AZD8895 and AZD1061); REGN10987 and REGN10933 (Regeneron),LY-CoV555 and LY-CoV16 (Lilly), and ADG10ADG20, and ADG30 (Adagio). LY-CoV16 and LY-CoV555 were almost incapable neutralizing P.1 and B.1.351 and LY-CoV16 also showed decreased ability in neutralization of B.1.1.7. Moreover, P.1 escaped neutralization by REGN10933 and there was a mild decrease in neutralization of P.1 by AZD8895, while AZD1061 and AZD 7442 showed same neutralization capacity of all three SARS-CoV-2 variants including B.1.1.7, P.1 and B. 1.351. In addition the ADG10, ADG20, and ADG30 antibodies neutralized all three variants, monoclonal antibody (mAb) 222 neutralized all three variants despite interacting with two of the ACE2-binding site mutations.105 In Zhou et al capacity of antibodies 175 and 158 to neutralize P.1 decreased remarkably, and P.1 escaped neutralization by 269 almost completely. Antibody 150 neutralization against P.1 is less decreased than against B.1.351.106 Some studies have suggested a moderate decrease in B1.617.2 (Delta variant) neutralization by the convalescent plasma and the sera of recipients of mRNA vaccines and resistance to monoclonal antibodies approved for COVID-19 treatment. B.1.617 is resistant against bamlanivimab, an authorized antibody. B.1.617 evades antibodies induced by infection or vaccination though less decreased by the B.1.351 variant.107,108 As Omicron RBD binds to human ACE2 with increased affinity in comparison to the Wuhan-Hu-1 RBD. It is observed by a group that most of the receptor-binding motif (RBM)-directed monoclonal antibodies (mAbs) in a study lost in vitro neutralizing capacity against Omicron and whereas 3 out of 29 monoclonal antibodies, S2K146 mAb 1 and sotrovimab 2, that included S2X259 3 and S2H97 4 which also possess the capacity to target antigenic sites outside the RBM.109 See Table 1.
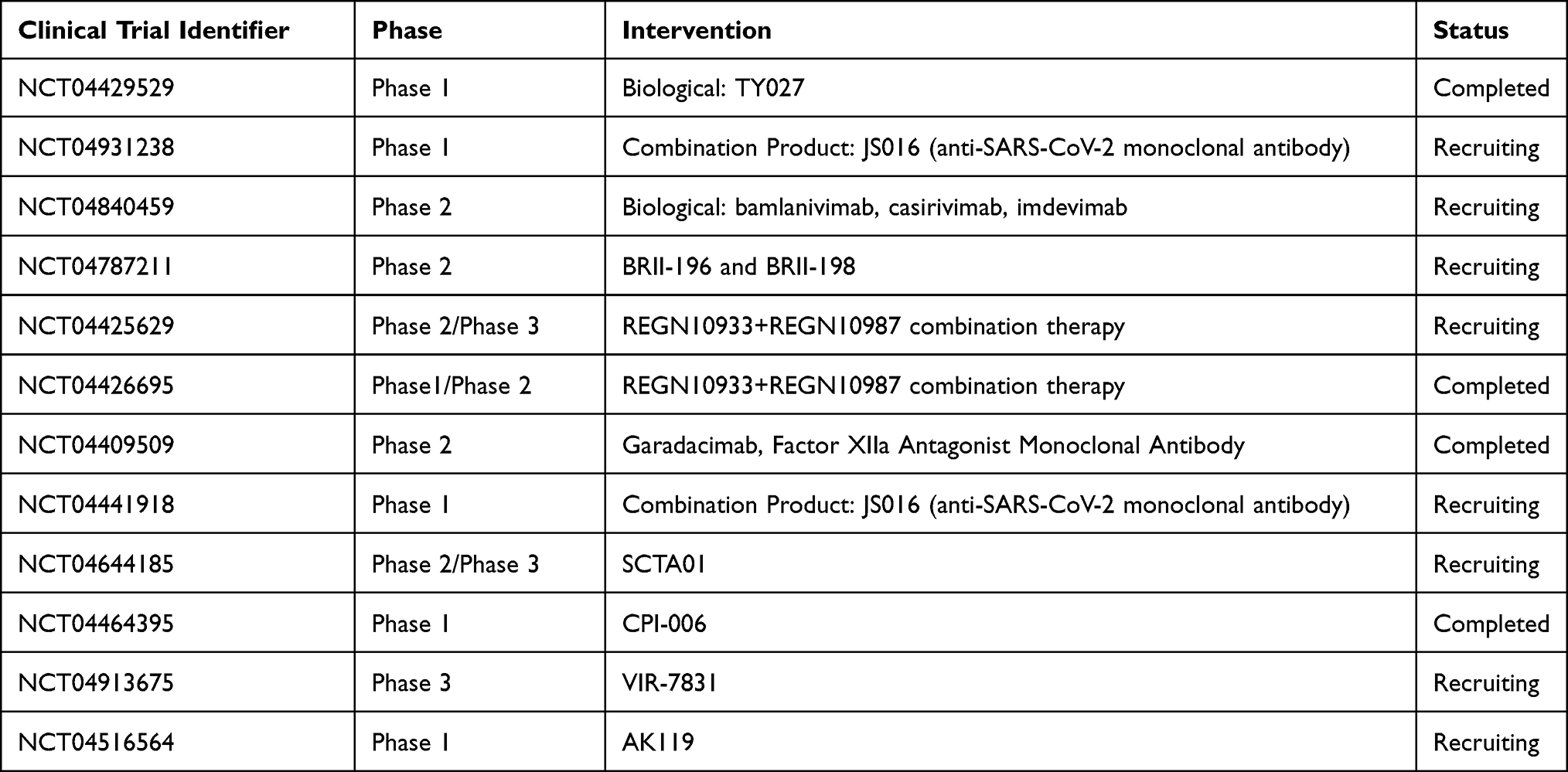 |
Table 1 Neutralizing Antibody Therapy |
T Cell Based Therapy
Patients with immunodeficiency may suffer from more negative prolonged effects of COVID-19 since T cells play a vital role in recovery.110 A study suggests that cultured SARS-CoV-2 membrane protein targeting specific T cell therapy would be applicable in prevention or early treatment of COVID-19 patients with immune deficiency after bone marrow transplantation while relieving inflammation.111 Furthermore in a study it was reported that receivers of SARS‐CoV‐2 spike glycoprotein‐based mRNA COVID‐19 vaccine introduced immune response against SARS‐CoV‐2 spike glycoprotein in healthy vaccinees who were not infected by SARS‐CoV‐2. SARS‐CoV‐2 spike glycoprotein‐targeting CD4+ and CD8+ T cells were extracted from the vaccinated individuals and largely expanded ex vivo in eleven days via cell cultures. Moreover, two of the vaccinees caught COVID-19 and showed that the BNT162b2 vaccine does not induce a immune reaction to nucleocapsid and membrane protein of SARS‐CoV‐2 whereas infection with the virus itself does.112 Appropriate T cell counts and adequate T cell activation and expansion in numbers are recorded in COVID-19 convalescent patients.113 Convalescent plasma of COVID-19 recovered patients contain CD4 and CD8 memory T cells which remember the SARS-CoV-2 spike, nucleocapsid and membrane antigens and can be isolated for multiple dose expansion to be used as SARS-CoV-2 specific immunotherapeutics.114 The study from Clinicaltrials.gov registration: NCT04578210 demonstrate the transfusion of convalescent CD45RA memory T cells to COVID-19 patients with moderate to severe symptoms are safe and applicable.115 In a study it is suggested that alone the utilization of immune checkpoint inhibitors or combined application of immune checkpoint inhibitors with anti-inflammatory drugs would enhance T-cell responses against the culprit virus that causes the COVID-19 crisis.116 T cells from recovered patient specific to both spike and nucleocapsid protein of the SARS-CoV-2 virus were discovered and cloned. The highly effective CD4+ T cells isolated from donors and expanded and suggested as a potential treatment option.117 Another study shows that T cells isolated from COVID-19-recovered donors which are specific in targeting against several structural SARS-CoV-2 proteins, among which include the Spike protein, then engineered genetically modified to equip them with resistance to corticosteroids.118 See Table 2.
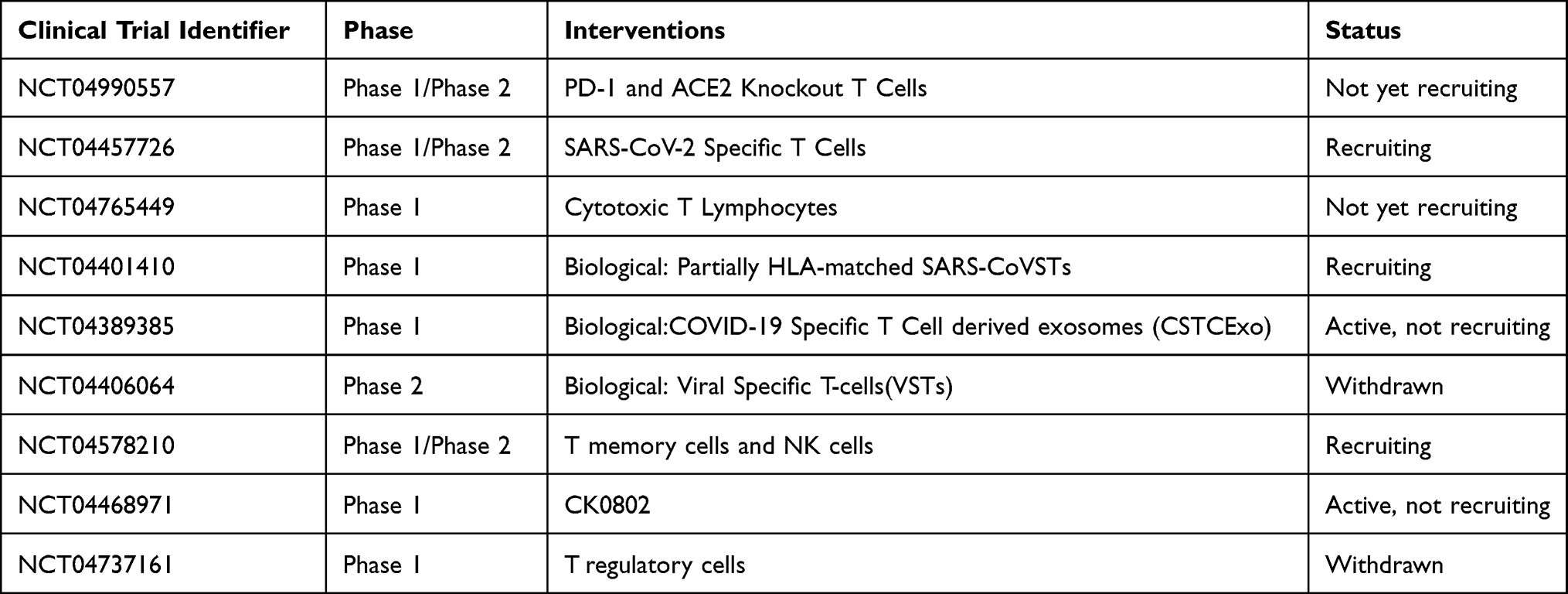 |
Table 2 T Cell Based Therapy |
CAR-T Cell Therapy
CAR-T cells are engineered from autologous or allogeneic sources of T cells to express a chimeric antigen receptor (CAR). CAR-T sparked an evolution the cancer therapy research. CAR consists of an extracellular single-chain variable fragment (scFv) complementary to the target antigens by the stimulation of the intracellular costimulatory signaling domain, followed by T cell proliferation, thereby annihilating tumor cells via cytolysis and cytokine release.119 CAR-T cells can be generated by isolation of relevant T cells from patients’ or donors’ blood, T cells are transduced by lentiviral or retroviral vector with the gene encoding a CAR to equip it with particular antigen specificity and then cultivated into larger quantities and then transfused into patients, to eliminate the cells with target antigens which suggests similar approach could be used to target the SARS-CoV-2 in order to assist patient’s immune system in their defence during COVID-19 onset.120 The CAR-T has gone under evolution and there are currently three generations of CAR, with low efficacy on solid tumours and recurrence of malignancies, and side effects.121 In 2017, the FDA approved Kymriah as cure for acute lymphoblastic leukemia and Yescarta for large B cell lymphoma which are quite expensive.122 Apart from application of CAR-T in cancer therapy, CAR-T cell therapy is also researched in chronic viral infectious diseases.123,124 Scientists are inspired likewise to design CAR-T cells specific to the SARS-CoV-2 spike, nucleocapsid and membrane antigens and have them re-infused into COVID-19 patient. There is also a hypothesis is based on the the CAR-T cells, that are generated to target the N protein of SARS-CoV-2, as the N protein binds to the SARS-2 viral RNA and is found in this virus.125 However a recent study that demonstrated the CAR-T technique as a feasible approach against COVID-19 applied the injection of a combination of 2×106NIH/3T3-S1 cells and 4×106 CR3014-28Z/ CR3022-28Z CAR-T cells into NOD-SCID IL2Rγ mice via the intraperitoneal route to conclude that CAR-T cells are able to eradicate the S1 expressing cells in vivo in mice.126
Obstacles of CAR-T Cell Therapy
Graft versus host disease is the primary challenge for treatments with allogeneic immune cells where the infused allogeneic T cells target the host cell antigens and attack the patient’s own cells of various organs such as skin, lungs, gastrointestinal tract, but leaving out kidneys, muscle, and brain, might implicate that the donor immune cells reach some organs more easily than others and there are obstructions that deters them from reaching some of the other organs.127 The infused allogeneic immune cells need to be competent and the recipient must express antigens on their cell surface that are foreign to the allogeneic transfused cells and the body of the patient receiving the treatment must be unable to deliver a successful immune response in killing the infused cells.128 The production of autologous CAR-T cells can be time consuming and cost great expenditure curbing their clinical implementation on large scale. Not all patients are able to produce enough functional T cells in numbers or at the time they are needed. Autologous T cell grafts is an obstacle for immunodeficient patients as they have difficulty in generating healthy T cell to be expanded and then reintroduced into the body. The challenge could be overcome potentially by the allogeneic T cell graft.129 Universal bank of CAR-T cells derived from allogeneic healthy donors would provide T cells to be genetically modified into off-the-shelf CAR-T cell based medications, simultaneously overcoming the potential graft-versus-host disease.130,131 The development of useful quantities of effective CAR immune cells treat COVID-19 patients is a race against time as the cultivation of modified immune cells usually requires at least several weeks.132 Also CAR-T cell therapy could cause off-target effects in cases when the healthy body cells coincidentally express the same target antigen intended for recognition for instance it was reported that CD19 is expressed on normal and malignant B cells.133,134 Thus, similar considerations must be taken into account when developing CAR-T therapeutics to treat COVID-19. Moreover CAR-T immunotherapy causes cytokine release a problem as seen in COVID-19.135,136 Hence, excessive cytokine/chemokine release can cause hyperinflammation, which leads to severe complications of the disease including pneumonitis, acute lung injury, acute respiratory distress syndrome, shock, vascular hyper permeability, organ failure, and death.137 Cytokine release syndrome (CRS) could lead to multiorgan dysfunction, which can occur in COVID-19, and after chimeric antigen receptor T (CAR-T) therapy, which is clinically characterized high fever, hyperinflammation, and severe organ dysfunction.138 According to the website clinicaltrials.gov there are currently at least 16 clinical trials ongoing to assess the efficacy of blocking IL-6R in COVID-19 patients with cytokine storm syndrome.139 Tocilizumab, is used for rheumatic conditions and is also co-labelled by the US Food and Drug Administration (FDA) for treatment with CAR-T therapy, which might be repurposed for treatment for COVID-19.140 Hemodialysis is suggested in a study that suggests by the use of cytokine filter to decrease the cytokine storm in an effort to preserve the life of a severe COVID-19 patient in the study and it came to the conclusion that it was effective, but implementation at earlier stages of the disease would prevent permanent damages.139 Tumor necrosis factor receptor 2 (TNFR2), to be targeted in Tregs-based therapies for management of COVID-19 is suggested as an approach to overcome cytokine storm to prevent multi organ failure.141
Natural Killer Cell Based Therapy
Natural Killer (NK) cell mediated antiviral activity which could include targeting the virus infected cells bring hope in developing specific COVID-19 treatment.142 IgG1 and IgG3 induce NK cell mediated cytotoxicity through the IgGFc-receptor IIIa during the onset of COVID-19 and NK cells utilize the Fc domain of bound IgG to target antigens on infected host cells, and kill these cells.143–145 NK cells have activating receptors such as natural cytotoxic receptors, NKG2D, CD16 that initiate NK cell activation.146 Natural killer cells could target virus-infected cells or cancer cells, without former sensitization whereas cytotoxic T lymphocytes needs sensitization whereas T cells requires a recognition of antigen to kill the virus infected cells. However, the NK cells are regulated by a mixture of stimulatory and inhibitory receptors.147 NK cells function without matching human leukocyte antigen (HLA), can serve as allogeneic effectors and do not need to be initially isolated from a human leukocyte antigen matching donor.148 CAR-NK elicit less severe side effects of graft-versus-host disease than CAR-T as the donor NK cells normally do not bring harm to non-hematopoietic cells and due to the reason NK cells possess a shorter life span than T cells, thus they normally do not harm cells from organs such as kidney, muscle, liver and lung.149 CAR-NK cell consists of isolating NK cells and modify these cells to express CAR that targets a tumor-specific protein for instance and then re-infuse them back into the recipient.150,151 CAR-modified human pluripotent stem cells might hold the capacity to produce functional CAR NK cells continuously, a potential solution to deal with the relative shorter life span of NK cells.152 Although NK92 cell line is often considered for application in NK cell therapy based clinical trials, they have to be irradiated due to the reason they are aneuploid.153 Moreover, a study reported of CAR-engineered hematopoietic stem/progenitor cells turning into NK cells and T cells that exhibit anti viral activities in humanized mice.154 CAR-NK cell therapy provide prospects for the prevention and cure of COVID-19 as CAR-NK cells might be designed to combat virus infected cells. CAR-NK cells may also utilized as an off-The-shelf tool alone or in combination with other therapeutic methods for viruses.155 However, an allogeneic and off-The-shelf NK cell which are cryopreserved, CYNK-001, derived from placental stem cells for treating leukemia and multiple myeloma is manufactured.156 Also scientists carried out trials to evaluate the effect of CYNK-001 against mild COVID-19. Moreover NKG2D-ACE2 CAR-NK cells are generated to treat COVID-19. The study from Clinicaltrials.gov registration: NCT04324996157 A study group developed CAR-NK cells to target the SARS-CoV-2 spike protein with the CR3022 scFv domain, a robust neutralizing antibody that targets the S protein of SARS-CoV-2. The results demonstrated that CR3022-CAR-NK cells hold the capacity to kill SARS-CoV-2 infected cells in vitro.158 For more NK cell based therapy clinical trials for COVID-19, see Table 3.
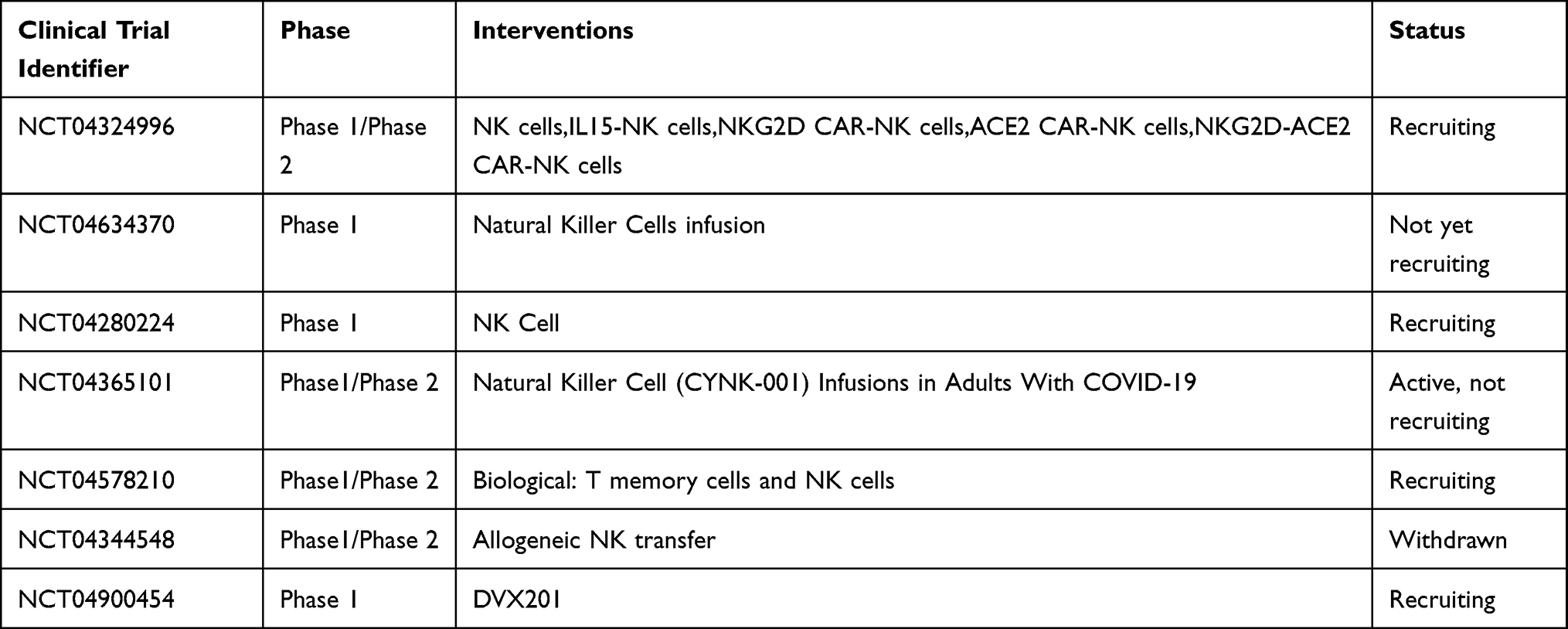 |
Table 3 Natural Killer Cell Based Therapy |
Clustered Regularly Interspaced Short Palindromic Repeat Cas System
Clustered regularly interspaced short palindromic repeat (CRISPR) Cas was discovered acting as defender of bacteria cells from foreign genetic material invasion.159 Bacteria cells defend themselves against virus foreign genomic sequences by producing a specific CRISPR RNA (crRNA) with a target sequence that binds and cleaves one or multiple regions in the invading viral genetic material.160 The CRISPR system includes two classes, class 1 and class 2. Class 1 is further subdivided into type I,III,and IV systems whereas class two is subdivided into type II, V, and VI. The class 1 CRISPR system consists of multiple Cas proteins that mediate various functions including the processing of pre crRNA, binding to crRNA, and cleavage whereas Class 2 consists of single, multi-domain crRNA binding Cas proteins.Cas 13 is categorized under class 2 type VI is utilizes to target RNA and is the unique RNA targeting-resistant mechanism in bacteria in addition to the type III CRISPR system in class 1.161–163 As CRISPR and CRISPR-associated Cas proteins possess many functions including, knock-in and knock-out of genes, inspiring scientists to develop therapeutics, to cure diseases by correction of genetic defects.164 Systems include CRISPR-Cas9,CRISPR-Cas12a and CRISPR-13 are technologies that can be applied as tools for genetic modification.165,166
CRISPR Cas System with Potential to Develop COVID-19 Therapeutics
A few studies consider CRISPR-Cas13 as an option for developing novel therapeutics against COVID-19. Immunotherapy focuses on providing assistance to the body immune system to combat SARS-CoV-2 or prevent viral entry into cells. The CRISPR-Cas13d system is associated with the RNA guided endonuclease obtained from Ruminococcus flavefaciens XPD3002 with the ability to target RNA which could offer opportunities to SARS-CoV-2 therapeutic development.167 The average size of Cas13d is 930 aa allowing CasRx effector domain fusions to be paired with a CRISPR array encoding multiple guide RNAs can still fit into the adeno-associated virus delivery vehicle for in vivo delivery.168 Cas13d could block expression of the viral gene by cleavage of the specific RNA sequence of the virus. Cas13 is small in size, possesses high specificity with catalytic activity which bears advantage over other Cas13 proteins, in targeting and inhibiting viruses such as SARS-CoV-2.169 A strategy is proposed to eliminate SARS-CoV-2 by the CRISPR-Cas13d system with gRNAs. As a maximum of three guide RNAs to target various peptide-encoding regions of SARS-CoV-2 RNA genetic material can be packaged into one adeno-associated virus vector, in order reduce the chance of resistance. Moreover, adeno-associated virus has serotypes highly specific to the lung which is a major organ that can be infected by SARS-CoV-2.170 Another study aimed to use the ability of the CRISPR-Cas13a system to edit RNA to the advantage of combating SARS-CoV-2. Cas13a functions when activated by a particular crRNA that aims at a specific sequence of single-stranded RNA to cleave the target RNA and causing collateral cleavage, resulting in the cleavage of the SARS-CoV-2 RNA sequence thus preventing viral protein synthesis and viral replication.171
Prophylactic antiviral CRISPR in huMAN cells (PAC-MAN) strategy is developed as an option acting as intervention to inhibit SARS-CoV-2 and other corona-viruses in a study. Cas13 and CRISPR-associated RNA (crRNA) can be utilized to cleave or degrade SARS-CoV-2 RNA genome and inhibit intracellular replication. The success of SARS-CoV-2 RNA degradation is assessed by the measurement of reporter signal in the research. As a pool of crRNA could suppress around 70% of reporter signal. Moreover, multiple crRNAs targeting the regions of the RdRP and N protein genes of SARS-CoV-2 resulted in over 80 and 90% viral RNA degradation.172 Three methods have been proposed by scientists for the CRISPR Cas technology to carry out its genetic material cleavage function inside living cells: 1. Introduce CRISPR components in the form of DNA to express Cas13 and crRNA shortly inside the cells. 2. Introduce Cas13 and crRNA in the form of RNA and allow it to be expressed inside host cells. 3. Introduce CRISPR components as a ribonucleoprotein complex which have a high editing efficiency transiently, with limited off-target effects.173,174 The technical problem lies in finding highly specific delivery system that transports CRISPR components uniquely into infected cells, otherwise there would be an equal opportunity that they get delivered into healthy cells.
Antibody And Cas fusion (ABACAS) is another strategy to gain specificity to SARS-CoV-2 infected cells where CRISPR nuclease is attached to a major virus structural protein for instance the S protein, to facilitate the entry of Cas13 into infected cells together. A promising antiviral therapeutic termed as fused antibody in the Antibody And Cas fusion (ABACAS) works by attaching Cas13 to the specific antibody of SARS-CoV-2 S protein for instance CR3022. The (ABACAS) approach possesses the ability to detect SARS-CoV-2 S protein and facilitates the entry of the Cas13 alongside the virus into the virus infected cell.170–177 SARS-CoV-2 enters human cells by the binding of S protein with ACE2 receptor therefore Peptidase Domain of ACE2 and Cas13 (PDCas13) has been suggested as a potential alternative method. The antibody utilized in ABACAS is replaced with the peptidase domain of ACE2 or with a simple peptide of ACE2 to bind with the S protein of SARS-CoV-2. The delivery of PDCas13 alongside with virus particles into the infected cells induces the antiviral impact of PDCas13 via degradation of viral genome RNA.178
More Hypotheses of CRISPR Cas Systems Against SARS-CoV-2
A review suggests that CRISPR/Cas13 system aims to cleave the viral RNA genome inside the infected cells or deactivate the expression of protein-coding genes of the SARS-CoV-2, such as spike protein coding genes, making this approach RNA specific instead of viral surface protein specific. Several target sites are recommended for it to cleave and to disable genome NSP1-NSP16, accessory proteins, and ORF1-10. In addition adeno-associated virus vector is suggested as a vehicle for transferring the CRISPR/Cas13d system into the infected cells.179 Moreover, another review proposed that if CRISPR/Cas13 gene constructs enters the SARS-CoV-2 infected cells by a virosomal delivery system, with the spike protein embedded on the membrane surface of the virosomes the virus genome sequence will be cleaved and thus the virus will be deactivated.180 Furthermore, it has been suggested as a hypothesis that CRISPR in human cells (PAC-MAN) along with CRISPR/Cas13d could break down the specific RNA sequence to inhibit viral replication as a prophylactic strategy.181
CRISPR Cas Technology to Enhance Cell Based Therapeutics
Due to the capacity of CRISPR Cas technology to modify genetic sequences, it has been used in a variety of applications to enhance immunotherapy. In an effort to investigate the engineering of universal anti tumour CAR-T cells for off-the-shelf purpose, Cas9 ribonucleoprotein electroporation was utilized to develop T cells deficient in human LEUKOCYTE antigen class I, human leukocyte antigen class II, and T-cell receptor via deletion of the beta 2-microglobulin (B2M), class II transactivator (CIITA), and T cell receptor alpha constant (TRAC) genes all at the same time and then transduced such T cells with CD19 chimeric antigen receptor. However they still could be killed by natural killer cells, although they survive better than T cells with TRAC and B2M double knockout in the allogeneic peripheral blood mononuclear cells in vivo of immunocompromised mice and did not cause graft versus host disease which indicates the potential of engineering SARS-Cov-2 specific off-the-shelf CAR-T cells for various variants.182 CRISPR/Cas9 system has been utilized to knock out T cell receptors (TCR), beta-2 microglobulin and PD1 simultaneously, to produce universal CAR-T cells with a lack of T cell receptors (TCR), human leukocyte antigens (HLA) class I molecule and programmed cell death protein 1(PD1) to treat infectious diseases. These CAR-T cells show the same potency as non genetically modified CAR-T cells and demonstrate the capacity to avoid graft-versus-host disease.183 Another study also reports that CRISPR/Cas 9 technology engineered PD1 deficient anti-CD19 CAR-T cells demonstrated improved anti tumour effects in vivo.184 CRISPR Cas technology can be utilized to knock out CD7 on CAR-T cells for the survival of CAR-T cells as they might kill among themselves due to the CAR recognition of CD7.185 There is another report about “off-the-shelf ”universal T cells generated with CRISPR /Cas9 system which is against CD7+ T cell malignancies and express neither CD7 or T cell receptor alpha chain (TRAC). These CAR-T cells demonstrated efficacy against human T cell acute lymphoblastic leukemia and without the occurrence of graft-versus-host disease in vivo or in vitro.186 A team led by oncologist Lu You at Sichuan University in Chengdu injected CRISPR Cas9 modified autologous T cell back into a person with aggressive lung cancer as part of a clinical trial for cancer therapy ClinicalTrials.gov Identifier: NCT02793856).187 These above examples could inspire scientists to develop universal T cells against COVID-19. In addition CRISPR/Cas9 technology shows great potential in promoting the CAR-NK cell therapy for instance by induced pluripotent stem cell -derived CAR NK cells188 as it has the capacity to knock in and out genes or cell surface receptors already researched in CAR-T cell engineering or correct disease causing genetic mutations.189,190
Limitation of CRISPR Cas Technology
Safety concerns such as off-target effects needs to be overcome before CRISPR Cas technology can be applied widely and clinically.The CRISPR/Cas mediated genome editing technique is still facing obstacles and requires optimization. The gRNA/Cas9 complex demonstrate tolerance of 1–3 or even more mismatches in their targets, as Cas9 could target many unintended sites in human genome other than aimed at. Thus, genomic sequences that harbors similarity to the intended target site could potentially become the undesired target site. A research group suggested that most of these off-target effects can be avoided by the careful control of CAS9 mRNA concentration.191 CRISPR/Cas9 systems targeting the human hemoglobin β and C-C chemokine receptor type 5 genes caused off-target cleavage, resulting in gross chromosomal deletions.192 A study indicated that the application of CRISPR gene editing could introduce hundreds of random mutations in the genome when whole-genome sequencing was carried out on a CRISPR–Cas9-edited mouse.193 Another shortcoming of CRISPR Cas technology is that it may result in unexpected translocations between double-strand breaks when multiple genes are modified.194 Off-target genetic mutations are relatively harder to detect than on-target modifications, as off- target mutations are random and could be anywhere in the genome. The most common method to counter these off-target effects is to design guide RNA sequences with more precision and there are tools are publicly available for that.195 By altering the length, secondary structure, or chemical composition of single guide RNAs, a few groups suggested that off-target effects can be minimized.196–198 Although, these modifications may also decrease their efficiency, studies showed that partial replacement of RNA nucleotides with DNA could further minimize the off-target impact while retaining on-target efficacy.199 See Figures 2.
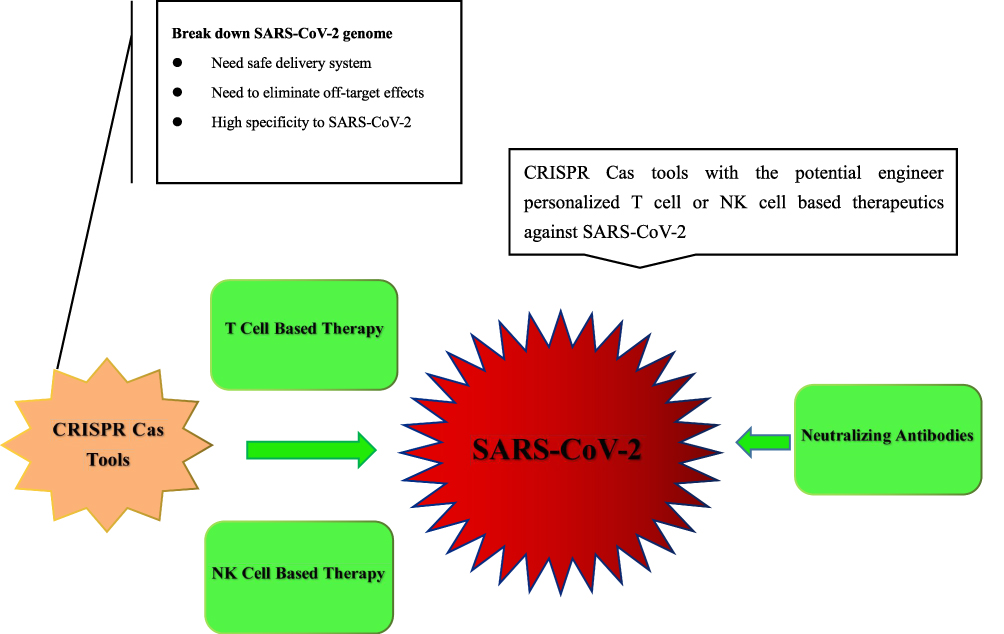 |
Figure 2 Combined strategy of CRISPR Cas tools and immunotherapeutics during the course of COVID-19. |
Results and Discussion
The current literature suggests that most of the current side effects of vaccines, convalescent plasma and neutralizing antibodies, are mainly mild or moderate and non-life threatening, Table 4. However, factors including mobility, environment, ethnicity, pollution, vaccinations among countries, and risk factors in society, that can accelerate diffusion of SARS-CoV-2 also affect the effectiveness of vaccines, therapies. Usually, vaccines are utilized for prophylactic purposes and therefore they are given to the population after various clinical trials to ensure safety before mass production whereas during natural exposure to authentic SARS-CoV-2, factors such as the quantity of viruses, the authenticity and viability of viruses, the type of variant and the precise viral immunity of the individuals exposed cannot be artificially controlled, but vaccines still provide protection for the population from SARS-CoV-2, although development of COVID-19 cannot be entirely prevented. Neutralizing antibodies and immune cell based therapeutics boost the body immune responses when administered to the immunosuppressed or infected individuals with highly specificity to the SARS-CoV-2 variants or any future novel variants detected in the body before the viral attack gains the upper hand, it is possible that they could promote a quicker recovery or prevent further progression of the disease. From the literature reviewed, it seems that the monitoring of blood and serological parameters and flow cytometry analysis of peripheral blood mononuclear cells to screen viral activities and immune activities in the body of COVID-19 patients could enable on-going personalized immunotherapeutic treatment and allow adjustment of regimen against the variants detected closing gates for immune escape. However, the development of immunotherapeutics such as CAR-T cell therapy or CAR-NK cell therapy are expensive and time consuming. In addition there are still technical difficulties to solve such as generating the appropriate immune cells and antibodies in sufficient quantities in a short period to treat the patient in order to prevent serious disease progression. Off-the-shelf immunotherapeutics might seem ideal as the physicians could prescribe immunotherapeutics according to the exact SARS-CoV-2 variant or variants. Both immunotherapy and CRISPR Cas based therapy have their limitations, but immunotherapy show some more advantage over CRISPR Cas technique currently as authorized convalescent plasma and neutralizing antibodies have already been utilized clinically with certain extent of efficacy, already having their safety evaluated in clinical trials, although immune cell based therapeutics are not yet mass produced for large scale clinical implementation.
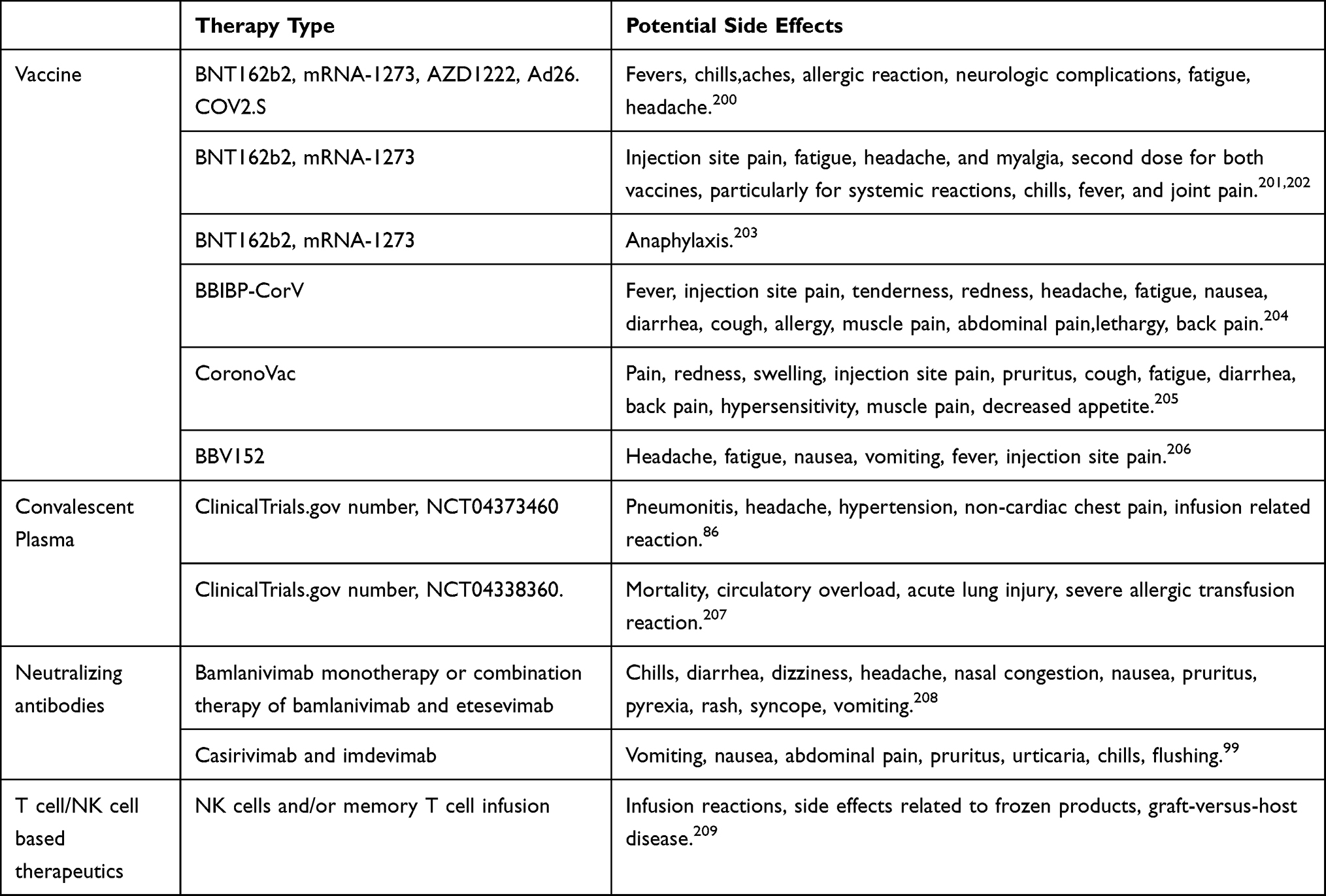 |
Table 4 Potential Side Effects of Vaccines and Immunotherapeutics |
Limitations
T cell based therapy, CAR-T cell therapy, NK cell based therapy or CAR-NK cell therapy and CRISPR Cas techniques are not yet therapeutics clinically conventionally used against COVID-19, and the data are currently rather insufficient in comparison to studies on the currently authorized vaccines, convalescent plasma and neutralizing antibodies, and studies of their effects against various current variants of concern Of SARS-CoV-2. We hope this review will inspire more future researches on CRISPR Cas based approaches and immune cell based therapeutics as specific cure against COVID-19.
Conclusion and Future Perspectives
The CRISPR Cas technology could be utilized to refine the combined cocktails of CAR-T cells, CAR-NK cells and neutralizing antibodies against SARS-CoV-2. In addition, CRISPR Cas technology are proposed in hypotheses to degrade the viral RNA in order terminate the SARS-CoV-2 activities. Thus personalized cocktails of immunotherapeutics and CRISPR Cas systems against COVID-19 as a strategy might be an option to inhibit further disease progression and circumvent immunity escape taking into account the genetic factors. CAR-T cell therapy, NK cell based therapy and CAR-NK cells are still under development whereas CRISPR Cas tool still require research and development before it can be applied clinically and safely to break down the SARS-CoV-2 genome in vivo with high efficacy, however challenges needs to be overcome in order to discover suitable delivery system for the CRISPR Cas tools, and it is necessary to eliminate any potential off-target effects to ensure precision and safety for it’s utilization against SARS-CoV-2 in vivo, and there are moral issues to consider before clinical application, although currently it has shown capacity to improve immune cell based therapeutics by knocking-in and knocking-out of genes and cell receptors as they have already been used in researches to refine cancer cell targeting CAR-T cells. SARS-CoV-2 could affect individuals uniquely, thus the application of immunotherapeutics and CRISPR Cas systems should be adjusted to every individual viewing their condition in a whole picture along with their unique behaviour, environment and genetic profiles.210
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rodríguez-Morales AJ, MacGregor K, Kanagarajah S, Patel D, Schlagenhauf P. Going global – travel and the 2019 novel coronavirus. Travel Med Infect Dis. 2020;33:101578. PMID: 32044389; PMCID: PMC7128681. doi:10.1016/j.tmaid.2020.101578
2. Perrotta F, Matera MG, Cazzola M, Bianco A. Severe respiratory SARS-CoV2 infection: does ACE2 receptor matter? Respir Med. 2020;168:105996. PMID: 32364961; PMCID: PMC7194970. doi:10.1016/j.rmed.2020.105996
3. Perlman S, Netland J. Coronaviruses post-SARS: update on replication and pathogenesis. Nat Rev Microbiol. 2009;7(6):439–450. PMID: 19430490; PMCID: PMC2830095. doi:10.1038/nrmicro2147
4. Coccia M. Effects of the spread of COVID-19 on public health of polluted cities: results of the first wave for explaining the dejà vu in the second wave of COVID-19 pandemic and epidemics of future vital agents. Environ Sci Pollut Res Int. 2021;28(15):19147–19154. PMID: 33398753; PMCID: PMC7781409. doi:10.1007/s11356-020-11662-7
5. Coccia M. How do low wind speeds and high levels of air pollution support the spread of COVID-19? Atmos Pollut Res. 2021;12(1):437–445. PMID: 33046960; PMCID: PMC7541047. doi:10.1016/j.apr.2020.10.002.
6. Coccia M. The effects of atmospheric stability with low wind speed and of air pollution on the accelerated transmission dynamics of COVID-19. Int J Environ Stud. 2021;78(1):1–27. doi:10.1080/00207233.2020.1802937
7. Coccia M. An index to quantify environmental risk of exposure to future epidemics of the COVID-19 and similar viral agents: theory and practice. Environ Res. 2020:110155. doi:10.1016/j.envres.2020.110155
8. Coccia M. Factors determining the diffusion of COVID-19 and suggested strategy to prevent future accelerated viral infectivity similar to COVID. Sci Total Environ. 2020. PMID: 32498152; PMCID: PMC7169901. doi:10.1016/j.scitotenv.2020.138474
9. Pan D, Sze S, Minhas JS, et al. The impact of ethnicity on clinical outcomes in COVID-19: a systematic review. EClinicalMedicine. 2020;23:100404. PMID: 32632416; PMCID: PMC7267805. doi:10.1016/j.eclinm.2020.100404
10. Askitas N, Tatsiramos K, Verheyden B. Estimating worldwide effects of non-pharmaceutical interventions on COVID-19 incidence and population mobility patterns using a multiple-event study. Sci Rep. 2021;11(1):1972. PMID: 33479325; PMCID: PMC7820317. doi:10.1038/s41598-021-81442-x
11. Flaxman S, Mishra S, Gandy A, et al.; Imperial College COVID-19 Response Team. Estimating the effects of non-pharmaceutical interventions on COVID-19 in Europe. Nature. 2020;584(7820):257–261. PMID: 32512579. doi:10.1038/s41586-020-2405-7
12. Ansumali S, Kaushal S, Kumar A, Prakash MK, Vidyasagar M. Modelling the covid-19 pandemic: asymptomatic patients, lockdown and herd immunity. IFAC-PapersOnLine. 2020;53(5):823–828. doi:10.1016/j.ifacol.2021.04.223
13. Coccia M. The relation between length of lockdown, numbers of infected people and deaths of Covid-19, and economic growth of countries: lessons learned to cope with future pandemics similar to Covid-19 and to constrain the deterioration of economic system. Sci Total Env. 2021;775:145801. doi:10.1016/j.scitotenv.2021.145801
14. Yuan J, Li M, Lv G, Lu ZK. Monitoring transmissibility and mortality of COVID-19 in Europe. Int J Infect Dis. 2020;95:311–315. doi:10.1016/j.ijid.2020.03.050
15. Coccia M. Optimal levels of vaccination to reduce COVID-19 infected individuals and deaths: a global analysis. Environ Res. 2022;204(Pt C):112314. PMID: 34736923; PMCID: PMC8560189. doi:10.1016/j.envres.2021.112314
16. Coccia M. The impact of first and second wave of the COVID-19 pandemic in society: comparative analysis to support control measures to cope with negative effects of future infectious diseases. Environ Res. 2021;197:111099. PMID: 33819476; PMCID: PMC8017951. doi:10.1016/j.envres.2021.111099
17. Coccia M. High health expenditures and low exposure of population to air pollution as critical factors that can reduce fatality rate in COVID-19 pandemic crisis: a global analysis. Environ Res. 2021;199:111339. PMID: 34029545; PMCID: PMC8139437. doi:10.1016/j.envres.2021.111339
18. Coccia M. Preparedness of countries to face COVID-19 pandemic crisis: strategic positioning and factors supporting effective strategies of prevention of pandemic threats. Environ Res. 2022;203:111678. PMID: 34280421; PMCID: PMC8284056. doi:10.1016/j.envres.2021.111678
19. Coccia M. Pandemic Prevention: lessons from COVID-19. Encyclopedia. 2021;1:433–444. doi:10.3390/encyclopedia1020036
20. Coccia M. Comparative critical decisions in management. In: Farazmand A, editor. Global Encyclopedia of Public Administration, Public Policy, and Governance. Cham: Springer Nature; 2021. doi:10.1007/978-3-319-31816-5_3969-1
21. Sarwar Z, Ahmad T, Kakar S. Potential approaches to combat COVID-19: a mini-review. Mol Biol Rep. 2020;47(12):9939–9949. PMID: 33185828; PMCID: PMC7662020. doi:10.1007/s11033-020-05988-1
22. Pasquini Z, Montalti R, Temperoni C, et al. Effectiveness of remdesivir in pa tients with COVID-19 under mechanical ventilation in an Italian ICU. J Antimicrob Chemother. 2020;75(11):3359–3365. doi:10.1093/jac/dkaa321
23. Pan H, Peto R, Henao-Restrepo AM, Preziosi MP; WHO Solidarity Trial Consortium. Repurposed antiviral drugs for COVID-19 - Interim WHO solidarity trial results. N Engl J Med. 2021;384(6):497–511. PMID: 33264556; PMCID: PMC7727327. doi:10.1056/NEJMoa2023184
24. Ahmad S, Hatmal MM, Lambuk L, MAI A-H, Alshaer W, Mohamud R. The role of TNFR2+ Tregs in COVID-19: an overview and a potential therapeutic strategy. Life Sci. 2021;286:120063. PMID: 34673116; PMCID: PMC8523334. doi:10.1016/j.lfs.2021.120063
25. Kalil AC, Stebbing J. Baricitinib: the first immunomodulatory treatment to reduce COVID-19 mortality in a placebo-controlled trial. Lancet Respir Med. 2021;9(12):1349–1351. PMID: 34480862; PMCID: PMC8409093. doi:10.1016/S2213-2600(21)00358-1
26. Tomazini BM, Maia IS, Cavalcanti AB, et al.; COALITION COVID-19 Brazil III Investigators. Effect of dexamethasone on days alive and ventilator-free in patients with moderate or severe acute respiratory distress syndrome and COVID-19: the CoDEX randomized clinical trial. JAMA. 2020;324(13):1307–1316. PMID: 32876695; PMCID: PMC7489411. doi:10.1001/jama.2020.17021
27. AlQahtani H, AlBilal S, Mahmoud E, et al. Outcomes associated with tocilizumab with or without corticosteroid versus dexamethasone for treatment of patients with severe to critical COVID-19 pneumonia. J Infect Public Health. 2022;15(1):36–41. PMID: 34883296; PMCID: PMC8611824. doi:10.1016/j.jiph.2021.11.017
28. Dash P, Mohapatra S, Ghosh S, Nayak B. A scoping insight on potential prophylactics, vaccines and therapeutic weaponry for the ongoing novel coronavirus (COVID-19) pandemic- a comprehensive review. Front Pharmacol. 2021;11:590154. PMID: 33815095; PMCID: PMC8015872. doi:10.3389/fphar.2020.590154
29. Cohen PA, Hall LE, John JN, Rapoport AB. The early natural history of SARS-CoV-2 infection: clinical observations from an urban, ambulatory COVID-19 clinic. Mayo Clin Proc. 2020;95:1124.
30. Wang D, Hu B, Hu C, et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA. 2020;323:1061–1069.
31. Ramírez P, Gordón M, Martín-Cerezuela M, et al. Acute respiratory distress syndrome due to COVID-19. Clinical and prognostic features from a Medical critical care unit in Valencia, Spain. Med Intensiva. 2021;45:27–34.
32. Gao YD, Ding M, Dong X, et al. Risk factors for severe and critically ill COVID-19 patients: a review. Allergy. 2021;76(2):428–455. PMID: 33185910. doi:10.1111/all.14657
33. Horwitz LI, Garry K, Prete AM, et al. Six-month outcomes in patients hospitalized with severe COVID-19. J Gen Intern Med. 2021:1–6. PMID: 34355349; PMCID: PMC8341831. doi:10.1007/s11606-021-07032-9
34. Shahgolzari M, Yavari A, Arjeini Y, et al. Immunopathology and Immunopathogenesis of COVID-19, what we know and what we should learn. Gene Rep. 2021;25:101417. PMID: 34778602; PMCID: PMC8570409. doi:10.1016/j.genrep.2021.101417
35. Ju B, Zhang Q, Ge J, et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature. 2020;584(7819):115–119. PMID: 32454513. doi:10.1038/s41586-020-2380-z
36. Chen G, Wu D, Guo W, et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest. 2020;130(5):2620–2629. PMID: 32217835; PMCID: PMC7190990. doi:10.1172/JCI137244
37. Wang F, Nie J, Wang H, et al. Characteristics of peripheral lymphocyte subset alteration in COVID-19 pneumonia. J Infect Dis. 2020;221:1762–1769.
38. Ni L, Ye F, Chen ML, et al. Detection of SARS-CoV-2-specific humoral and cellular immunity in COVID-19 convalescent individuals. Immunity. 2020;52:971–977.
39. Gustine JN, Jones D. Immunopathology of hyperinflammation in COVID-19. Am J Pathol. 2021;191:4–17.
40. Kaneko N, Kuo HH, Boucau J, et al.; Massachusetts Consortium on Pathogen Readiness Specimen Working Group. Loss of Bcl-6-expressing T follicular helper cells and germinal centers in COVID-19. Cell. 2020;183(1):143–157.e13. PMID: 32877699; PMCID: PMC7437499. doi:10.1016/j.cell.2020.08.025
41. Chen L, Zody MC, Di Germanio C, et al. Emergence of multiple SARS-CoV-2 antibody escape variants in an immunocompromised host undergoing convalescent plasma treatment. mSphere. 2021;6(4):e0048021. PMID: 34431691; PMCID: PMC8386433. doi:10.1128/mSphere.00480-21
42. Harvey WT, Carabelli AM, Jackson B, et al.; COVID-19 Genomics UK (COG-UK) Consortium. SARS-CoV-2 variants, spike mutations and immune escape. Nat Rev Microbiol. 2021;19(7):409–424. PMID: 34075212; PMCID: PMC8167834. doi:10.1038/s41579-021-00573-0
43. Thomson EC, Rosen LE, Shepherd JG, et al.; ISARIC4C Investigators; COVID-19 Genomics UK (COG-UK) Consortium. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell. 2021;184(5):1171–1187.e20. PMID: 33621484; PMCID: PMC7843029. doi:10.1016/j.cell.2021.01.037
44. Cevik M, Grubaugh ND, Iwasaki A, Openshaw P. COVID-19 vaccines: keeping pace with SARS-CoV-2 variants. Cell. 2021;184(20):5077–5081. PMID: 34534444; PMCID: PMC8445744. doi:10.1016/j.cell.2021.09.010
45. Kannan SR, Spratt AN, Sharma K, Chand HS, Byrareddy SN, Singh K. Omicron SARS-CoV-2 variant: unique features and their impact on pre-existing antibodies. J Autoimmun. 2022;126:102779. PMID: 34915422; PMCID: PMC8666303. doi:10.1016/j.jaut.2021.102779
46. Li Q, Lu H. Latest updates on COVID-19 vaccines. Biosci Trends. 2021;14(6):463–466. PMID: 33390384. doi:10.5582/bst.2020.03445
47. Elkhalifa D, Rayan M, Negmeldin AT, Elhissi A, Khalil A. Chemically modified mRNA beyond COVID-19: potential preventive and therapeutic applications for targeting chronic diseases. Biomed Pharmacother. 2022;145:112385. PMID: 34915673; PMCID: PMC8552589. doi:10.1016/j.biopha.2021.112385
48. Polack Fernando P, Thomas Stephen J, Kitchin N, et al.; C4591001 Clinical Trial Group. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N Engl J Med. 2020;383(27):2603–2615. doi:10.1056/NEJMoa2034577
49. Mulligan MJ, Lyke KE, Kitchin N, et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Erratum in: Nature. 2021 Feb;590(7844):E26. Nature. 2020. 586(7830):589–593. PMID: 32785213. doi:10.1038/s41586-020-2639-4
50. Sahin U, Muik A, Vogler I, et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature. 2021;595(7868):572–577. doi:10.1038/s41586-021-03653-6
51. Afonin KA, Dobrovolskaia MA, Ke W, Grodzinski P, Bathe M. Critical review of nucleic acid nanotechnology to identify gaps and inform a strategy for accelerated clinical translation. Adv Drug Deliv Rev. 2021;181:114081. PMID: 34915069. doi:10.1016/j.addr.2021.114081
52. Wang C, Zhang Y, Dong Y. Lipid nanoparticle-mRNA formulations for therapeutic applications. Acc Chem Res. 2021;54(23):4283–4293. PMID: 34793124. doi:10.1021/acs.accounts.1c00550
53. Ma KS, Lee CC, Liu KJ, Wei JC, Lee YT, Wang LT. Safety and seroconversion of immunotherapies against SARS-CoV-2 infection: a systematic review and meta-analysis of clinical trials. Pathogens. 2021;10(12):1537. PMID: 34959492; PMCID: PMC8706687. doi:10.3390/pathogens10121537
54. Favresse J, Jean-Louis. B, Mullier F, et al. Antibody titres decline 3-month post-vaccination with BNT162b2. Emerg Microbes Infect. 2021;10(1):1495–1498. doi:10.1080/22221751.2021.1953403
55. Schramm R, Costard-Jäckle A, Rivinius R, et al. Poor humoral and T-cell response to two-dose SARS-CoV-2 messenger RNA vaccine BNT162b2 in cardiothoracic transplant recipients. Clin Res Cardiol. 2021;110(8):1142–1149. PMID: 34241676; PMCID: PMC8267767. doi:10.1007/s00392-021-01880-5
56. Gupta S, Su H, Narsai T, Agrawal S. SARS‐CoV‐2‐associated T‐cell responses in the presence of humoral immunodeficiency. Int Arch Allergy Immunol. 2021;182:
57. Strugnell RA, Wijburg OL. The role of secretory antibodies in infection immunity. Nat Rev Microbiol. 2010;8(9):656–667. doi:10.1038/nrmicro2384
58. Goes LR, Siqueira JD, Garrido MM, et al.; INCA COVID-19 Task Forceˆ. New infections by SARS-CoV-2 variants of concern after natural infections and post-vaccination in Rio de Janeiro, Brazil. Infect Genet Evol. 2021;94:104998. PMID: 34252616; PMCID: PMC8270730. doi:10.1016/j.meegid.2021.104998
59. Swan DA, Bracis C, Janes H, et al. COVID-19 vaccines that reduce symptoms but do not block infection need higher coverage and faster rollout to achieve population impact. Sci Rep. 2021;11(1):15531. PMID: 34330945; PMCID: PMC8324774. doi:10.1038/s41598-021-94719-y
60. Kustin T, Harel N, Finkel U, et al. Evidence for increased breakthrough rates of SARS-CoV-2 variants of concern in BNT162b2-mRNA-vaccinated individuals. Nat Med. 2021;27(8):1379–1384. PMID: 34127854; PMCID: PMC8363499. doi:10.1038/s41591-021-01413-7
61. Naaber P, Tserel L, Kangro K, et al. Dynamics of antibody response to BNT162b2 vaccine after six months: a longitudinal prospective study. Lancet Reg Health Eur. 2021:100208. PMID: 34514454; PMCID: PMC8418937. doi:10.1016/j.lanepe.2021.100208
62. Jalkanen P, Kolehmainen P, Häkkinen HK, et al. COVID-19 mRNA vaccine induced antibody responses against three SARS-CoV-2 variants. Nat Commun. 2021;12(1):3991. PMID: 34183681; PMCID: PMC8239026. doi:10.1038/s41467-021-24285-4
63. Chemaitelly H, Yassine HM, Benslimane FM, et al. mRNA-1273 COVID-19 vaccine effectiveness against the B.1.1.7 and B.1.351 variants and severe COVID-19 disease in Qatar. Nat Med. 2021;27(9):1614–1621. PMID: 34244681. doi:10.1038/s41591-021-01446-y
64. Madhi SA, Baillie V, Cutland CL, et al.; NGS-SA Group; Wits-VIDA COVID Group. Efficacy of the ChAdOx1 nCoV-19 Covid-19 vaccine against the B.1.351 variant. N Engl J Med. 2021;384(20):1885–1898. PMID: 33725432; PMCID: PMC7993410. doi:10.1056/NEJMoa2102214
65. Fendler A, Shepherd STC, Au L, et al. Adaptive immunity and neutralizing antibodies against SARS-CoV-2 variants of concern following vaccination in patients with cancer: the CAPTURE study. Nat Cancer. 2021;2:1321–1337. PMID: 34950880; PMCID: PMC7612125. doi:10.1038/s43018-021-00274-w
66. Mlcochova P, Kemp SA, Dhar MS, et al.; Indian SARS-CoV-2 Genomics Consortium (INSACOG); Genotype to Phenotype Japan (G2P-Japan) Consortium; CITIID-NIHR BioResource COVID-19 Collaboration. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature. 2021;599(7883):114–119. PMID: 34488225; PMCID: PMC8566220. doi:10.1038/s41586-021-03944-y
67. Alter G, Yu J, Liu J, et al. Immunogenicity of Ad26.COV2.S vaccine against SARS-CoV-2 variants in humans. Nature. 2021;596(7871):268–272. PMID: 34107529; PMCID: PMC8357629. doi:10.1038/s41586-021-03681-2
68. Wang GL, Wang ZY, Duan LJ, et al. Susceptibility of circulating SARS-CoV-2 variants to neutralization. N Engl J Med. 2021;384(24):2354–2356. PMID: 33822491; PMCID: PMC8063885. doi:10.1056/NEJMc2103022
69. Li XN, Huang Y, Wang W, et al. Effectiveness of inactivated SARS-CoV-2 vaccines against the Delta variant infection in Guangzhou: a test-negative case-control real-world study. Emerg Microbes Infect. 2021;10(1):1751–1759. PMID: 34396940; PMCID: PMC8425710. doi:10.1080/22221751.2021.1969291
70. Sapkal GN, Yadav PD, Ella R, et al. Inactivated COVID-19 vaccine BBV152/COVAXIN effectively neutralizes recently emerged B.1.1.7 variant of SARS-CoV-2. J Travel Med. 2021;28(4):taab051. PMID: 33772577; PMCID: PMC8083765. doi:10.1093/jtm/taab051
71. Yadav PD, Sapkal GN, Ella R, et al. Neutralization against B.1.351 and B.1.617.2 with sera of COVID‐19 recovered cases and vaccines of BBV152. bioRxiv. 2021. doi:10.1101/2021.06.05.447177
72. Garcia-Beltran WF, St Denis KJ, Hoelzemer A, et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. Cell. 2022. doi:10.1016/j.cell.2021.12.033
73. Lu L, Mok BW, Chen LL, et al. Neutralization of SARS-CoV-2 Omicron variant by sera from BNT162b2 or Coronavac vaccine recipients. Clin Infect Dis. 2021:ciab1041. PMID: 34915551; PMCID: PMC8754807. doi:10.1093/cid/ciab1041
74. Edara VV, Manning KE, Ellis M, et al. mRNA-1273 and BNT162b2 mRNA vaccines have reduced neutralizing activity against the SARS-CoV-2 Omicron variant. bioRxiv. 2021. PMID: 34981056; PMCID: PMC8722593. doi:10.1101/2021.12.20.473557
75. Rzymski P, Camargo CA, Fal A, et al. COVID-19 vaccine boosters: the good, the bad, and the ugly. Vaccines. 2021;9(11):1299. PMID: 34835230; PMCID: PMC8623745. doi:10.3390/vaccines9111299
76. Hadj Hassine I. Covid-19 vaccines and variants of concern: a review. Rev Med Virol. 2021:e2313. PMID: 34755408; PMCID: PMC8646685. doi:10.1002/rmv.2313
77. Rice A, Verma M, Shin A, et al. Intranasal plus subcutaneous prime vaccination with a dual antigen COVID-19 vaccine elicits T-cell and antibody responses in mice. Sci Rep. 2021;11(1):14917. PMID: 34290317; PMCID: PMC8295250. doi:10.1038/s41598-021-94364-5
78. Menichetti F, Falcone M, Tiseo G. Management of COVID patients with convalescent plasma: do we have the final word? Eur J Intern Med. 2022;95:13–16. PMID: 34895816; PMCID: PMC8630688. doi:10.1016/j.ejim.2021.10.029
79. Anup A, Aparna M, Gunjan K, et al. Convalescent plasma in the management of moderate covid-19 in adults in India: open label phase II multiCentre randomised controlled trial (PLACID Trial). BMJ. 2020;371:m3939.
80. Xinyi X, Kening L, Lingxiang W, et al. Improved clinical symptoms and mortality among patients with severe or critical COVID-19 after convalescent plasma transfusion. Blood. 2020;136:755–759.
81. Tim L, Ryan M, Vicente P, et al. Successful transfer of anti-SARS-CoV-2 immunity using convalescent plasma in an MM patient with hypogammaglobulinemia and COVID-19. Blood Adv. 2020;4:4864–4868.
82. Arnold Egloff SA, Junglen A, Restivo JS, et al. Convalescent plasma associates with reduced mortality and improved clinical trajectory in patients hospitalized with COVID-19. J Clin Invest. 2021:151788. PMID: 34464352. doi:10.1172/JCI151788
83. Joyner MJ, Carter RE, Senefeld JW, et al. Convalescent plasma antibody levels and the risk of death from Covid-19. N Engl J Med. 2021;384(11):1015–1027. PMID: 33523609; PMCID: PMC7821984. doi:10.1056/NEJMoa2031893
84. Simonovich VA, Burgos Pratx LD, Scibona P, et al.; PlasmAr Study Group. A randomized trial of convalescent plasma in Covid-19 severe pneumonia. N Engl J Med. 2021;384(7):619–629. PMID: 33232588; PMCID: PMC7722692. doi:10.1056/NEJMoa2031304
85. Kemp SA, Collier DA, Datir RP, et al.; CITIID-NIHR BioResource COVID-19 Collaboration; COVID-19 Genomics UK (COG-UK) Consortium. SARS-CoV-2 evolution during treatment of chronic infection. Nature. 2021;592(7853):277–282. PMID: 33545711; PMCID: PMC7610568. doi:10.1038/s41586-021-03291-y
86. Sullivan DJ, Gebo KA, Shoham S, et al. Randomized controlled trial of early outpatient COVID-19 treatment with high-titer convalescent plasma. medRxiv. 2021. PMID: 34981068; PMCID: PMC8722611. doi:10.1101/2021.12.10.21267485
87. Garcia-Muñoz R, Farfán-Quiroga G, Ruiz-de-lobera N, et al. Serology-based therapeutic strategy in SARS-CoV-2-infected patients. Int Immunopharmacol. 2021;101(Pt B):108214. PMID: 34649116; PMCID: PMC8491979. doi:10.1016/j.intimp.2021.108214
88. Dionne A, Sperotto F, Chamberlain S, et al. Association of myocarditis with BNT162b2 messenger RNA COVID-19 vaccine in a case series of children. JAMA Cardiol. 2021:e213471. PMID: 34374740; PMCID: PMC8356143. doi:10.1001/jamacardio.2021.3471
89. Wang P, Nair MS, Liu L, et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature. 2021;593(7857):130–135. PMID: 33684923. doi:10.1038/s41586-021-03398-2
90. Cele S, Gazy I, Jackson L, et al.; Network for Genomic Surveillance in South Africa; COMMIT-KZN Team. Escape of SARS-CoV-2 501Y.V2 from neutralization by convalescent plasma. Nature. 2021;593(7857):142–146. PMID: 33780970. doi:10.1038/s41586-021-03471-w
91. Casadevall A, Henderson JP, Joyner MJ, Pirofski LA. SARS-CoV-2 variants and convalescent plasma: reality, fallacies, and opportunities. J Clin Invest. 2021;131(7):e148832. PMID: 33621214; PMCID: PMC8011895. doi:10.1172/JCI148832
92. Monir E, Li Q, Shurong H, et al. A cross-reactive human IgA monoclonal antibody blocks SARS-CoV-2 spike-ACE2 interaction. Nat Commun. 2020;11(1):4198. doi:10.1038/s41467-020-18058-8
93. Ucciferri C, Vecchiet J, Falasca K. Role of monoclonal antibody drugs in the treatment of COVID-19. World J Clin Cases. 2020;8:4280.
94. Matthews Dean B. A cocktail of antibodies for COVID-19 therapy. Nat Rev Immunol. 2020;20:591.
95. Andreano E, Rappuoli R. SARS-CoV-2 escaped natural immunity, raising questions about vaccines and therapies. Nat Med. 2021;27(5):759–761. PMID: 33972793. doi:10.1038/s41591-021-01347-0
96. Piccoli L, Park YJ, Tortorici MA, et al. Mapping neutralizing and immunodominant sites on the SARS-CoV-2 spike receptor-binding domain by structure-guided high-resolution serology. Cell. 2020;183(4):1024–1042.e21. PMID: 32991844; PMCID: PMC7494283. doi:10.1016/j.cell.2020.09.037
97. Parums DV. Editorial: post-exposure prophylactic neutralizing monoclonal antibodies to SARS-CoV-2 for individuals at high risk for COVID-19. Med Sci Monit. 2021;27:e934393. PMID: 34393218; PMCID: PMC8378223. doi:10.12659/MSM.934393
98. Hammarström L, Marcotte H, Piralla A, Baldanti F, Pan-Hammarström Q. Antibody therapy for COVID-19. Curr Opin Allergy Clin Immunol. 2021;21(6):553–558.
99. Weinreich DM, Sivapalasingam S, Norton T, et al.; Trial Investigators. REGN-COV2, a neutralizing antibody cocktail, in outpatients with Covid-19. N Engl J Med. 2021;384(3):238–251. PMID: 33332778; PMCID: PMC7781102. doi:10.1056/NEJMoa2035002
100. Goldin L, Elders T, Werhane L, Korwek K, Poland R, Guy J. Reactions and COVID-19 disease progression following SARS-CoV-2 monoclonal antibody infusion. Int J Infect Dis. 2021;112:73–75. PMID: 34508863; PMCID: PMC8425746. doi:10.1016/j.ijid.2021.09.007
101. Kim YJ, Jang US, Soh SM, Lee J-Y, Lee H-R. The impact on infectivity and neutralization efficiency of SARS-CoV-2 lineage B.1.351 pseudovirus. Viruses. 2021;13(4):633. PMID: 33917138; PMCID: PMC8067879. doi:10.3390/v13040633
102. Wibmer CK, Ayres F, Hermanus T, et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat Med. 2021;27(4):622–625. PMID: 33654292. doi:10.1038/s41591-021-01285-x.
103. Tortorici MA, Beltramello M, Lempp FA, et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science. 2020;370(6519):950–957. PMID: 32972994; PMCID: PMC7857395. doi:10.1126/science.abe3354
104. Starr TN, Czudnochowski N, Liu Z, et al. SARS-CoV-2 RBD antibodies that maximize breadth and resistance to escape. Nature. 2021;597(7874):97–102. PMID: 34261126. doi:10.1038/s41586-021-03807-6
105. Dejnirattisai W, Zhou D, Supasa P, et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell. 2021;184(11):2939–2954.e9. PMID: 33852911; PMCID: PMC8008340. doi:10.1016/j.cell.2021.03.055
106. Zhou D, Dejnirattisai W, Supasa P, et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell. 2021;184(9):2348–2361.e6. PMID: 33730597; PMCID: PMC7901269. doi:10.1016/j.cell.2021.02.037
107. Edara -V-V, Pinsky BA, Suthar MS, et al. Infection and vaccine-induced neutralizing-antibody responses to the SARS-CoV-2 B.1.617 variants. N Engl J Med. 2021;385(7):664–666. PMID: 34233096; PMCID: PMC8279090. doi:10.1056/NEJMc2107799
108. Hoffmann M, Hofmann-Winkler H, Krüger N, et al. SARS-CoV-2 variant B.1.617 is resistant to bamlanivimab and evades antibodies induced by infection and vaccination. Cell Rep. 2021;36(3):109415. PMID: 34270919; PMCID: PMC8238662. doi:10.1016/j.celrep.2021.109415
109. Cameroni E, Saliba C, Bowen JE, et al. Broadly neutralizing antibodies overcome SARS-CoV-2 Omicron antigenic shift. bioRxiv. 2021. PMID: 34931194; PMCID: PMC8687478. doi:10.1101/2021.12.12.472269
110. Dai M, Liu D, Liu M, et al. Patients with cancer appear more vulnerable to SARS-CoV-2: a multicenter study during the COVID-19 outbreak. Cancer Discov. 2020;10(6):783–791. PMID: 32345594; PMCID: PMC7309152. doi:10.1158/2159-8290.CD-20-0422
111. Keller MD, Harris KM, Jensen-Wachspress MA, et al. SARS-CoV-2–specific T cells are rapidly expanded for therapeutic use and target conserved regions of the membrane protein. Blood. 2020;136(25):2905–2917. PMID: 33104167. doi:10.1182/blood.2020008488
112. Taborska P, Lastovicka J, Stakheev D, Strizova Z, Bartunkova J, Smrz D. SARS-CoV-2 spike glycoprotein-reactive T cells can be readily expanded from COVID-19 vaccinated donors. Immun Inflamm Dis. 2021;9(4):1452–1467. PMID: 34314576. doi:10.1002/iid3.496
113. Braun J, Loyal L, Frentsch M, et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature. 2020;587(7833):270–274. PMID: 32726801. doi:10.1038/s41586-020-2598-9
114. Cooper RS, Fraser AR, Smith L, et al. Rapid GMP-compliant expansion of SARS-CoV-2–Specific T cells from convalescent donors for use as an allogeneic cell therapy for COVID-19. Front Immunol. 2021;11:598402. PMID: 33488592; PMCID: PMC7819874. doi:10.3389/fimmu.2020.598402
115. Pérez-Martínez A, Mora-Rillo M, Ferreras C, et al. Phase I dose-escalation single centre clinical trial to evaluate the safety of infusion of memory T cells as adoptive therapy in COVID-19 (RELEASE). EClinicalMedicine. 2021;39:101086. PMID: 34405140; PMCID: PMC8361305. doi:10.1016/j.eclinm.2021.101086
116. Toor SM, Saleh R, Sasidharan Nair V, Taha RZ, Elkord E. T-cell responses and therapies against SARS-CoV-2 infection. Immunology. 2021;162(1):30–43. PMID: 32935333; PMCID: PMC7730020. doi:10.1111/imm.13262
117. Verhagen J, Van der Meijden ED, Lang V, et al. Human CD4+ T cells specific for dominant epitopes of SARS-CoV-2 Spike and Nucleocapsid proteins with therapeutic potential. Clin Exp Immunol. 2021;205(3):363–378. PMID: 34061349; PMCID: PMC8239517. doi:10.1111/cei.13627
118. Basar R, Uprety N, Ensley E, et al. Generation of glucocorticoid-resistant SARS-CoV-2 T cells for adoptive cell therapy. Cell Rep. 2021;36(3):109432. doi:10.1016/j.celrep.2021.109432
119. Li J, Li W, Huang K, Zhang Y, Kupfer G, Zhao Q. Chimeric antigen receptor T cell (CAR-T) immunotherapy for solid tumors: lessons learned and strategies for moving forward. J Hematol Oncol. 2018;11(1):22. doi:10.1186/s13045-018-0568-6
120. Ma T, Shi J, Liu H. Chimeric antigen receptor T cell targeting B cell maturation antigen immunotherapy is promising for multiple myeloma. Ann Hematol. 2019;98(4):813–822. doi:10.1007/s00277-018-03592-9
121. Zhang C, Liu J, Zhong JF, Zhang X. Engineering CAR-T cells. Biomark Res. 2017;5(1):22. PMID: 28652918; PMCID: PMC5482931. doi:10.1186/s40364-017-0102-y
122. Aamir A, Shahab U, Martin S. CAR-T cell therapies: an overview of clinical studies supporting their approved use against acute lymphoblastic leukemia and large B-cell lymphomas. Int J Mol Sci. 2020;21:3906.
123. Bohne F, Chmielewski M, Ebert G, et al. T cells redirected against hepatitis B virus surface proteins eliminate infected hepatocytes. Gastroenterology. 2008;134(1):239–247. PMID: 18166356. doi:10.1053/j.gastro.2007.11.002
124. Hale M, Mesojednik T, Romano Ibarra GS, et al. Engineering HIV-resistant, anti-HIV chimeric antigen receptor T cells. Mol Ther. 2017;25(3):570–579. PMID: 28143740; PMCID: PMC5363191. doi:10.1016/j.ymthe.2016.12.023
125. Al-Utaibi KA, Nutini A, Sohail A, Arif R, Tunc S, Sait SM. Forecasting the action of CAR-T cells against SARS-Corona virus-II infection with branching process. Model Earth Syst Environ. 2021:1–9. PMID: 34667828; PMCID: PMC8516624. doi:10.1007/s40808-021-01312-3
126. Guo X, Kazanova A, Thurmond S, Saragovi HU, Rudd CE. Effective chimeric antigen receptor T cells against SARS-CoV-2. iScience. 2021;24(11):103295. PMID: 34693218; PMCID: PMC8520176. doi:10.1016/j.isci.2021.103295
127. Ferrara JL, Levine JE, Reddy P, Holler E. Graft-versus-host disease. Lancet. 2009;373(9674):1550–1561. doi:10.1016/S0140-6736(09)60237-3
128. Billingham RE. The biology of graft-versus-host reactions.. Harvey Lect. 1966;62:21–78.
129. Torikai H, Cooper LJ. Translational implications for off-the-shelf immune cells expressing chimeric antigen receptors. Mol Ther. 2016;24(7):1178–1186. PMID: 27203439; PMCID: PMC5088751. doi:10.1038/mt.2016.106
130. Ramos CA, Heslop HE, Brenner MK. CAR-T Cell Therapy for Lymphoma. Annu Rev Med. 2016;67(1):165–183. doi:10.1146/annurev-med-051914-021702
131. Zhao J, Lin Q, Song Y, Liu D. Universal CARs, universal T cells, and universal CAR T cells. J Hematol Oncol. 2018;11(1):132. doi:10.1186/s13045-018-0677-2
132. Maiti SN, Huls H, Singh H, et al. Sleeping beauty system to redirect T-cell specificity for human applications. J Immunother. 2013;36(2):112–123. PMID: 23377665; PMCID: PMC3568214. doi:10.1097/CJI.0b013e3182811ce9
133. Schuster SJ, Svoboda J, Chong EA, et al. Chimeric antigen receptor T cells in refractory B-cell lymphomas. N Engl J Med. 2017;377(26):2545–2554. PMID: 29226764; PMCID: PMC5788566. doi:10.1056/NEJMoa1708566
134. Kalos M, Levine BL, Porter DL, et al. T cells with chimeric antigen receptors have potent antitumor effects and can establish memory in patients with advanced leukemia. Sci Transl Med. 2011;3(95):95ra73. PMID: 21832238; PMCID: PMC3393096. doi:10.1126/scitranslmed.3002842.
135. Agarwal S, June CH. Harnessing CAR T-cell insights to develop treatments for hyperinflammatory responses in patients with COVID-19. Cancer Discov. 2020;10(6):775–778. doi:10.1158/2159-8290.CD-20-0473
136. Henderson LA, Canna SW, Schulert GS, et al. On the alert for cytokine storm: immunopathology in COVID −19. Arthritis Rheumatol. 2020;72(7):1059–1063. PMID: 32293098; PMCID: PMC7262347. doi:10.1002/art.41285
137. Kang S, Tanaka T, Narazaki M, Kishimoto T. Targeting interleukin-6 signaling in clinic. Immunity. 2019;50(4):1007–1023. doi:10.1016/j.immuni.2019.03.026
138. Li X, Shao M, Zeng X, Qian P, Huang H. Signaling pathways in the regulation of cytokine release syndrome in human diseases and intervention therapy. Signal Transduct Target Ther. 2021;6(1):367. doi:10.1038/s41392-021-00764-4
139. Rizvi S, Danic M, Silver M, LaBond V. Cytosorb filter: an adjunct for survival in the COVID-19 patient in cytokine storm? A case report. Heart Lung. 2021;50(1):44–50. PMID: 33041058; PMCID: PMC7500900. doi:10.1016/j.hrtlng.2020.09.007
140. Schulert GS, Grom AA. Pathogenesis of macrophage activation syndrome and potential for cytokine- directed therapies. Annu Rev Med. 2015;66(1):145–159.
141. Ahmad S, Hatmal MM, Lambuk L, Al-Hatamleh MAI, Alshaer W, Mohamud R. The role of TNFR2+ Tregs in COVID-19: an overview and a potential therapeutic strategy. Life Sci. 2021;286:120063. PMID: 34673116; PMCID: PMC8523334. doi:10.1016/j.lfs.2021.120063
142. Soleimanian S, Yaghobi R. Harnessing memory NK cell to protect against COVID-19. Front Pharmacol. 2020;11:1309. doi:10.3389/fphar.2020.01309
143. Dokun AO, Kim S, Smith HRC, Kang H-SP, Chu DT, Yokoyama WM. Specific and nonspecific NK cell activation during virus infection. Nat Immunol. 2001;2(10):951–956. doi:10.1038/ni714
144. Bonam SR, Kaveri SV, Sakuntabhai A, Gilardin L, Bayry J. Adjunct immunotherapies for the management of severely ill COVID-19 patients. Cell Rep Med. 2020;1(2):100016. doi:10.1016/j.xcrm.2020.100016
145. Pinto D, Park Y-J, Beltramello M, et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature. 2020;583(7815):s290–s295. doi:10.1038/s41586-020-2349-y
146. Lanier LL. NK cell recognition. Annu Rev Immunol. 2005;23(1):225–274. doi:10.1146/annurev.immunol.23.021704.115526
147. Bakker ABH, Wu J, Phillips JH, Lanier LL. NK cell activation: distinct stimulatory pathways counterbalancing inhibitory signals. Hum Immunol. 2000;61(1):18–27. doi:10.1016/S0198-8859(99)00160-3
148. Morvan MG, Lanier LL. NK cells and cancer: you can teach innate cells new tricks. Nat Rev Cancer. 2016;16(1):7–19. doi:10.1038/nrc.2015.5
149. Curti A, Ruggeri L, D’Addio A, et al. Successful transfer of alloreactive haploidentical KIR ligand-mismatched natural killer cells after infusion in elderly high risk acute myeloid leukemia patients. Blood. 2011;118(12):3273–3279. PMID: 21791425. doi:10.1182/blood-2011-01-329508
150. Xia J, Minamino S, Kuwabara Kazuma K. CAR-expressing NK cells for cancer therapy: a new hope. Biosci Trends. 2020;14(5):354–359. doi:10.5582/bst.2020.03308
151. Ghaffari S, Kazerooni H, Salehi-Najafabadi A. An overview of the recent findings of cell-based therapies for the treatment and management of COVID-19. Int Immunopharmacol. 2021;101(Pt B):108226. PMID: 34634685; PMCID: PMC8492917. doi:10.1016/j.intimp.2021.108226
152. Liu D, Tian S, Zhang K, et al. Chimeric antigen receptor (CAR)-modified natural killer cell-based immunotherapy and immunological synapse formation in cancer and HIV. Erratum in: Protein Cell. 2017 Jun 23. Protein Cell. 2017. 8(12):861–877. PMID: 28488245; PMCID: PMC5712291. doi:10.1007/s13238-017-0415-5
153. Klingemann H, Boissel L, Toneguzzo F. Natural killer cells for immunotherapy – advantages of the NK-92 cell line over blood NK cells. Front Immunol. 2016;7:91. doi:10.3389/fimmu.2016.00091
154. Zhen A, Kamata M, Rezek V, et al. HIV-specific immunity derived from chimeric antigen receptor-engineered stem cells. Mol Ther. 2015;23(8):1358–1367. PMID: 26050990; PMCID: PMC4817874. doi:10.1038/mt.2015.102
155. Mazarzaei A, Vafaei M, Ghasemian A, Mirforughi SA, Rajabi Vardanjani H, Alwan NA. Memory and CAR-NK cell-based novel approaches for HIV vaccination and eradication. J Cell Physiol. 2019;234(9):14812–14817. doi:10.1002/jcp.28280
156. Ilic D, Liovic M. Industry updates from the field of stem cell research and regenerative medicine in January 2020: Industry News. Regen Med. 2020;14(5):345–351. doi:10.2217/rme-2020-0016
157. Jeyaraman M, Muthu S, Bapat A, et al. Bracing NK cell based therapy to relegate pulmonary inflammation in COVID-19. Heliyon. 2021;7(7):e07635. PMID: 34312598; PMCID: PMC8294777. doi:10.1016/j.heliyon.2021.e07635
158. Ma M, Badeti S, Geng K, Liu D. Efficacy of targeting SARS-CoV-2 by CAR-NK cells. bioRxiv. 2020. doi:10.1101/2020.08.11.247320
159. Wright AV, NuñezJ K, Doudna JA. Biology and applications of CRISPR systems:harnessing nature’s toolbox for genome engineering. Cell. 2016. doi:10.1016/j.cell.2015.12.035
160. Labrie SJ, Samson JE, Moineau S. Bacteriophage resistance mechanisms. Nat Rev Microbiol. 2010;8:317–327.
161. Makarova KS, Wolf YI, Alkhnbashi OS, et al. An updated evolutionary classification of CRISPR−Cas systems. Nat Rev Microbiol. 2015;13(11):722–736.
162. Shmakov S, Smargon A, Scott D, et al. Diversity and evolution of class 2 CRISPR−Cassystems. Nat.Rev.Microbiol. 2017;15(3):169–182.
163. Abudayyeh OO, Gootenberg JS, Konermann S, et al. C2c2 is a single-component programmable RNAguided RNA-targeting CRISPR effector. Science. 2016;353:aaf5573.
164. Safari F, Afarid M, Rastegari B, Borhani-Haghighi A, Barekati-Mowahed M, Behzad-Behbahani A. CRISPR systems: novel approaches for detection and combating COVID-19. Virus Res. 2021;294:198282. PMID: 33428981; PMCID: PMC7832022. doi:10.1016/j.virusres.2020.198282
165. Horvath P, Romero DA, Coûté-Monvoisin AC, et al. Diversity, activity, and evolution of CRISPR loci in Streptococcus thermophilus. J Bacteriol. 2008;190(4):1401–1412. PMID: 18065539; PMCID: PMC2238196. doi:10.1128/JB.01415-07
166. Horvath P, Coûté-Monvoisin AC, Romero DA, Boyaval P, Fremaux C, Barrangou R. Comparative analysis of CRISPR loci in lactic acid bacteria genomes. Int J Food Microbiol. 2009;131(1):62–70. PMID: 18635282. doi:10.1016/j.ijfoodmicro.2008.05.030
167. Konermann S, Lotfy P, Brideau NJ, Oki J, Shokhirev MN, Hsu PD. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell. 2018;173(3):665–676.e14. PMID: 29551272; PMCID: PMC5910255. doi:10.1016/j.cell.2018.02.033
168. Naldini L. Gene therapy returns to centre stage. Nature. 2015;526:351–360.
169. Yan WX, Chong S, Zhang H, et al. Cas13d is a compact RNA-targeting type VI CRISPR effector positively modulated by a WYL-domain-containing accessory protein. Mol Cell. 2018;70(2):327–339.e5. PMID: 29551514; PMCID: PMC5935466. doi:10.1016/j.molcel.2018.02.028
170. Nguyen TM, Zhang Y, Pandolfi PP. Virus against virus: a potential treatment for 2019-nCov (SARS-CoV-2) and other RNA viruses. Cell Res. 2020;30(3):189–190. PMID: 32071427; PMCID: PMC7054296. doi:10.1038/s41422-020-0290-0
171. Wang L, Zhou J, Wang Q, Wang Y, Kang C. Rapid design and development of CRISPR-Cas13a targeting SARS-CoV-2 spike protein. Theranostics. 2021;11(2):649–664. PMID: 33391497; PMCID: PMC7738867. doi:10.7150/thno.51479
172. Abbott TR, Dhamdhere G, Liu Y, et al. Development of CRISPR as an antiviral strategy to combat SARS-CoV2 and influenza. Cell. 2020;181(4):865–876 e812.
173. Zuris JA, Thompson DB, Shu Y, et al. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat Biotechnol. 2015;33(1):73–80. PMID: 25357182; PMCID: PMC4289409. doi:10.1038/nbt.3081
174. Kim S, Kim D, Cho SW, Kim J, Kim JS. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 2014;24(6):1012–1019. PMID: 24696461; PMCID: PMC4032847. doi:10.1101/gr.171322.113
175. Yuan M, Wu NC, Zhu X, et al. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science. 2020;368(6491):630–633.
176. Safari F, Farajnia S, Ghasemi YN. New developments in CRISPR technology: improvements in specificity and efficiency. Curr Pharm Biotechnol. 2017;18(13):1038–1054.
177. Joyce MG, Sankhala RS, Chen WH, et al. Site of vulnerability on the receptor binding domain of the SARS-CoV-2 spike glycoprotein. bioRxiv. 2020. PMID: 32511298; PMCID: PMC7217142. doi:10.1101/2020.03.15.992883
178. Nalawansha DA, Samarasinghe KTG. Double-barreled CRISPR technology as a novel treatment strategy for COVID-19. ACS Pharmacol Transl Sci. 2020;3(5):790–800. PMID: 33062949; PMCID: PMC7469881. doi:10.1021/acsptsci.0c00071
179. Lotfi M, Rezaei N. CRISPR/Cas13: a potential therapeutic option of COVID-19. Biomed Pharmacother. 2020;131:110738.
180. Abbaszadeh-Goudarzi K, Nematollahi MH, Khanbabaei H, et al. Targeted delivery of CRISPR/Cas13 as a promising therapeutic approach to treat SARS-CoV-2. Curr Pharm Biotechnol. 2021;22(9):1149–1155. PMID: 33038909. doi:10.2174/1389201021666201009154517
181. Banerjee A, Mukherjee S, Maji BK. Manipulation of genes could inhibit SARS-CoV-2 infection that causes COVID-19 pandemics. Exp Biol Med. 2021;246(14):1643–1649. PMID: 33899542; PMCID: PMC8090827. doi:10.1177/15353702211008106
182. Kagoya Y, Guo T, Yeung B, et al. Genetic ablation of HLA class I, class II, and the T-cell receptor enables allogeneic T cells to be used for adoptive T-cell therapy. Cancer Immunol Res. 2020;(7):926–936. PMID: 32321775. doi:10.1158/2326-6066.CIR-18-0508
183. Ren J, Liu X, Fang C, Jiang S, June CH, Zhao Y. Multiplex genome editing to generate universal CAR T cells resistant to PD1 inhibition. Clin Cancer Res. 2017;23(9):2255–2266. PMID: 27815355; PMCID: PMC5413401. doi:10.1158/1078-0432.CCR-16-1300
184. Rupp LJ, Schumann K, Roybal KT, et al. CRISPR/Cas9-mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Sci Rep. 2017;Apr(1):737. PMID: 28389661; PMCID: PMC5428439. doi:10.1038/s41598-017-00462-8
185. Gomes-Silva D, Srinivasan M, Sharma S, et al. CD7-edited T cells expressing a CD7-specific CAR for the therapy of T-cell malignancies. Blood. 2017;130(3):285–296. PMID: 28539325; PMCID: PMC5520470. doi:10.1182/blood-2017-01-761320
186. Cooper ML, Choi J, Staser K, et al. An “off-The-shelf” fratricide-resistant CAR-T for the treatment of T cell hematologic malignancies. Leukemia. 2018;32(9):1970–1983. PMID: 29483708; PMCID: PMC6102094. doi:10.1038/s41375-018-0065-5
187. Cyranoski D. CRISPR gene-editing tested in a person for the first time. Nature. 2016;539:479.
188. Li Y, Hermanson DL, Moriarity BS, Kaufman DS. Human iPSC-derived natural killer cells engineered with chimeric antigen receptors enhance anti-tumor activity. Cell Stem Cell. 2018;23(2):181–192.e5. PMID: 30082067; PMCID: PMC6084450. doi:10.1016/j.stem.2018.06.002
189. Ureña-Bailén G, Lamsfus-Calle A, Daniel-Moreno A, et al. CRISPR/Cas9 technology: towards a new generation of improved CAR-T cells for anticancer therapies. Brief Funct Genomics. 2020;19(3):191–200. PMID: 31844895. doi:10.1093/bfgp/elz039
190. Schumann K, Lin S, Boyer E, et al. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proc Natl Acad Sci U S A. 2015;112(33):10437–10442. PMID: 26216948; PMCID: PMC4547290. doi:10.1073/pnas.1512503112
191. Fujii W, Kawasaki K, Sugiura K, Naito K. Efficient generation of large-scale genome-modified mice using gRNA and CAS9 endonuclease. Nucleic Acids Res. 2013;41(20):e187. PMID: 23997119; PMCID: PMC3814358. doi:10.1093/nar/gkt772
192. Manghwar H, Li B, Ding X, et al. CRISPR/Cas systems in genome editing: methodologies and tools for sgRNA design, off-target evaluation, and strategies to mitigate off-target effects. Adv Sci. 2020;7(6):1902312. PMID: 32195078; PMCID: PMC7080517. doi:10.1002/advs.201902312
193. Schaefer KA, Wu WH, Colgan DF, Tsang SH, Bassuk AG, Mahajan VB. Unexpected mutations after CRISPR-Cas9 editing in vivo. Nat Methods. 2017;14:547–548. doi:10.1038/nmeth.4293
194. Poirot L, Philip B, Schiffer-Mannioui C, et al. Multiplex genome-edited T-cell manufacturing platform for “Off-The-Shelf” adoptive T-cell immunotherapies. Cancer Res. 2015;75(18):3853–3864. PMID: 26183927. doi:10.1158/0008-5472.CAN-14-3321
195. Listgarten J, Weinstein M, Kleinstiver BP, et al. Prediction of off-target activities for the end-to-end design of CRISPR guide RNAs. Nat Biomed Eng. 2018;(1):38–47. PMID: 29998038; PMCID: PMC6037314. doi:10.1038/s41551-017-0178-6
196. Fu Y, Sander JD, Reyon D, Cascio VM, Joung JK. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat Biotechnol. 2014;32(3):279–284. PMID: 24463574; PMCID: PMC3988262. doi:10.1038/nbt.2808
197. Hendel A, Bak RO, Clark JT, et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nat Biotechnol. 2015;33(9):985–989. PMID: 26121415; PMCID: PMC4729442. doi:10.1038/nbt.3290
198. Kocak DD, Josephs EA, Bhandarkar V, Adkar SS, Kwon JB, Gersbach CA. Increasing the specificity of CRISPR systems with engineered RNA secondary structures. Nat Biotechnol. 2019;37(6):657–666. PMID: 30988504; PMCID: PMC6626619. doi:10.1038/s41587-019-0095-1
199. Yin H, Song CQ, Suresh S, et al. Structure-guided chemical modification of guide RNA enables potent non-viral in vivo genome editing. Nat Biotechnol. 2017;35(12):1179–1187. PMID: 29131148; PMCID: PMC5901668. doi:10.1038/nbt.4005
200. Epstein S, Xia Z, Lee AJ, Farber R, Wesley SF. Vaccination against SARS-CoV-2 in neuroinflammatory disease: early safety/ tolerability data. Mult Scler Relat Disord. 2022;57:103433.
201. Chapin-Bardales J, Gee J, Myers T. Reactogenicity following receipt of mRNA-based COVID-19 vaccines. JAMA. 2021;325(21):2201–2202. PMID: 33818592. doi:10.1001/jama.2021.5374
202. Finsterer J. Neurological side effects of SARS-CoV-2 vaccinations. Acta Neurol Scand. 2022;145(1):5–9. PMID: 34750810; PMCID: PMC8653194. doi:10.1111/ane.13550
203. Shimabukuro TT, Cole M, Su JR. Reports of anaphylaxis after receipt of mRNA COVID-19 vaccines in the US-December 14, 2020-January 18, 2021. JAMA. 2021;325(11):1101–1102. PMID: 33576785. doi:10.1001/jama.2021.1967
204. Saeed BQ, Al-Shahrabi R, Alhaj SS, Alkokhardi ZM, Adrees AO. Side effects and perceptions following Sinopharm COVID-19 vaccination. Int J Infect Dis. 2021;111:219–226. PMID: 34384899; PMCID: PMC8351310. doi:10.1016/j.ijid.2021.08.013
205. Zhang Y, Zeng G, Pan H, et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: a randomised, double-blind, placebo-controlled, Phase 1/2 clinical trial. Lancet Infect Dis. 2021;21(2):181–192. PMID: 33217362; PMCID: PMC7832443. doi:10.1016/S1473-3099(20)30843-4
206. Ella R, Vadrevu KM, Jogdand H, et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: a double-blind, randomised, phase 1 trial. Erratum in: Lancet Infect Dis. 2021 Apr;21(4):e81. Lancet Infect Dis. 2021. 21(5):637–646. PMID: 33485468; PMCID: PMC7825810. doi:10.1016/S1473-3099(20)30942-7
207. Joyner MJ, Wright RS, Fairweather D, et al. Early safety indicators of COVID-19 convalescent plasma in 5000 patients. J Clin Invest. 2020;130(9):4791–4797. PMID: 32525844; PMCID: PMC7456238. doi:10.1172/JCI140200
208. Gottlieb RL, Nirula A, Chen P, et al. Effect of bamlanivimab as monotherapy or in combination with etesevimab on viral load in patients with mild to moderate COVID-19: a randomized clinical trial. JAMA. 2021;325(7):632–644. PMID: 33475701; PMCID: PMC7821080. doi:10.1001/jama.2021.0202
209. García-García I, Guerra-García P, Ferreras C, et al. A phase I/II dose-escalation multi-center study to evaluate the safety of infusion of natural killer cells or memory T cells as adoptive therapy in coronavirus pneumonia and/or lymphopenia: RELEASE study protocol. Trials. 2021;22(1):674. PMID: 34600562; PMCID: PMC8487326. doi:10.1186/s13063-021-05625-7
210. Zeng BZ. On the holographic model of human body, 1st National Conference of Comparative Studies Traditional Chinese Medicine and West Medicine, Medicine and Philosophy; 1992.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.