Back to Journals » Journal of Inflammation Research » Volume 17
Immune-Related Genes and Immune Cell Infiltration Characterize the Maturation Status of Arteriovenous Fistulas: An Integrative Bioinformatics Study and Experimental Validation Based on Transcriptome Sequencing
Authors Lu P, Wang T, Wan Z, Wang M, Zhou Y, He Z, Liao S, Liu H, Shu C
Received 2 August 2023
Accepted for publication 31 October 2023
Published 8 January 2024 Volume 2024:17 Pages 137—152
DOI https://doi.org/10.2147/JIR.S433525
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Peng Lu,1,2,* Tun Wang,1,2,* Zicheng Wan,1,2 Mo Wang,1,2 Yang Zhou,1,2 Zhenyu He,1,2 Sheng Liao,1,2 Haiyang Liu,3 Chang Shu1,2,4
1Department of Vascular Surgery, the Second Xiangya Hospital, Central South University, Changsha, People’s Republic of China; 2Institute of Vascular Diseases, Central South University, Changsha, People’s Republic of China; 3Department of Geriatrics, the Second Xiangya Hospital, Central South University, Changsha, People’s Republic of China; 4Center of Vascular Surgery, Fuwai Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, 100037, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chang Shu, Department of Vascular Surgery, the Second Xiangya Hospital, Central South University, Changsha, People’s Republic of China, Tel +86 136 0744 4222, Email [email protected]
Purpose: Arteriovenous fistula (AVF) is the preferred vascular access for hemodialysis, but the low maturation rate is concerning. Immune cells’ impact on AVF maturation lacks bioinformatics research. The study aims to investigate the potential predictive role of immune-related genes and immune cell infiltration characteristics in AVF maturation.
Patients and Methods: We analyzed the high-throughput sequencing dataset to identify differentially expressed genes (DEGs). Then, we performed enrichment analyses (GO, KEGG, GSEA) on immune-related genes and pathways in mature AVF. We focused on differentially expressed immune-related genes (DEIRGs) and constructed a PPI network to identify hub genes. These hub genes were validated in other databases and experiments, including qPCR and immunohistochemistry (IHC). The immune cell infiltration characteristics in native veins, failed AVFs, and matured AVFs were analyzed by cibersortX. Partial experimental validation was conducted using clinical samples.
Results: Our results showed that immune-related genes and signaling pathways are significantly enriched in mature AVF. We validated this in other databases and ultimately identified three hub genes (IL1B, IL6, CXCR4) in combination with experiments. Significant differences in immune cell infiltration characteristics were observed among native veins, failed AVFs, and matured AVFs. Immune cell infiltration analysis revealed that accumulation of CD4+ T cells, dendritic cells, mast cells and M2 macrophages contribute to AVF maturation. These immune-related genes and immune cells have the potential to serve as predictive factors for AVF maturation. We partially validated this experimentally.
Conclusion: From a bioinformatics perspective, our results have identified, for the first time, a set of immune-related genes and immune cell infiltration features that can characterize the maturation of AVF and significantly impact AVF maturation. These features hold potential as predictive indicators for AVF maturation outcomes.
Keywords: AVF maturation, bioinformatics, immune-related genes, inflammation
Introduction
Chronic kidney disease (CKD) affects between 8% and 16% of the population worldwide.1 End-stage renal disease (ESRD) is the terminal state of CKD. Patients with ESRD rely on renal replacement therapy for life, and native arteriovenous fistula (AVF) is the preferred vascular access.2 AVF maturation clinically means that 6 weeks after surgery, the blood flow in the AVF is ≥600 mL/min, the depth from the skin surface is ≤6 mm, and the lumen diameter of the venous outflow is >6 mm2.Successful AVF maturation requires venous wall thickening and outward remodeling.3 However, approximately 60% of AVFs will lose function after surgery, also known as failed AVFs, which causes a huge loss to the patients.4,5 We still have a limited understanding of the mechanisms that influence AVF maturation.
Immune response and inflammation have intersections and overlaps in the occurrence and development of disease, both consistently accompany with dialysis in ESRD patients. Some studies have found an increase in inflammatory markers primarily in dialysis patients.6,7 It is clear that dialysis can prime blood lymphocytes to release IL-1 and IL-2.8 The plasma levels of IL-6 and IL-1B have been found to increase during dialysis.9,10 Furthermore, Immune response and inflammation also play an important role in tissue repair and regeneration. For instance, inflammatory monocytes and tissue-resident macrophages are key regulators of tissue repair.11 Macrophages undergo marked phenotypic and functional changes during the phases of tissue repair.12 Research has found that regulating genes related to vascular remodeling, such as ST2,13 can improve vascular remodeling, and vascular remodeling and tissue repair clearly exist after AVF.14,15 However, the effects of immunity and inflammation on AVF are still poorly understood.
High-throughput RNA sequencing (RNA-seq) is commonly used to find key genes associated with specific biological processes and to build predictive models in conjunction with a patient’s clinical prognosis. However, failed AVFs or AVFs that fail after a period of use often undergo interventional procedures due to the requirements of the surgical approach to AVF, making it difficult to obtain unintervened failed AVF tissue.2 Since failed AVFs or those that fail after a period of use typically do not lead to acute patient mortality, and they often require interventional treatment due to the demands of AVF surgical techniques, it becomes challenging to obtain the patient’s accurate tissue at the follow-up endpoint in clinical settings. Therefore, unlike in the field of oncology, there have been no large sample size studies of AVF to predict AVF maturation or failure. The search for molecular or cellular features that predict or characterize the maturation state of AVF is a viable alternative.
Due to ethical constraints, researchers often face difficulties in obtaining samples twice from the same patient at the same anatomical location to establish optimal control experiments and minimize the impact of baseline differences in patient characteristics. Owing to the unique two-stage AVF creation surgeries, the surgeon can potentially collect samples at 2 times: a segment of the vein used to create the arteriovenous anastomosis during the 1st stage surgery and a sample of the juxta-anastomotic area of both mature and failed AVFs during the second-stage transposition. Thus, the high-throughput RNA sequence data (GSE119296) we used for analysis collected samples of native veins (at the time of the first-stage procedure) and their corresponding AVFs (at the time of the second-stage procedure after one month) from the same anatomical location in the same individual (Figure 1).16 To understand the differences in immune gene expression and immune cell characteristics of native veins, failed AVFs and mature AVFs, we performed bioinformatics analysis and validated the results using clinical samples.
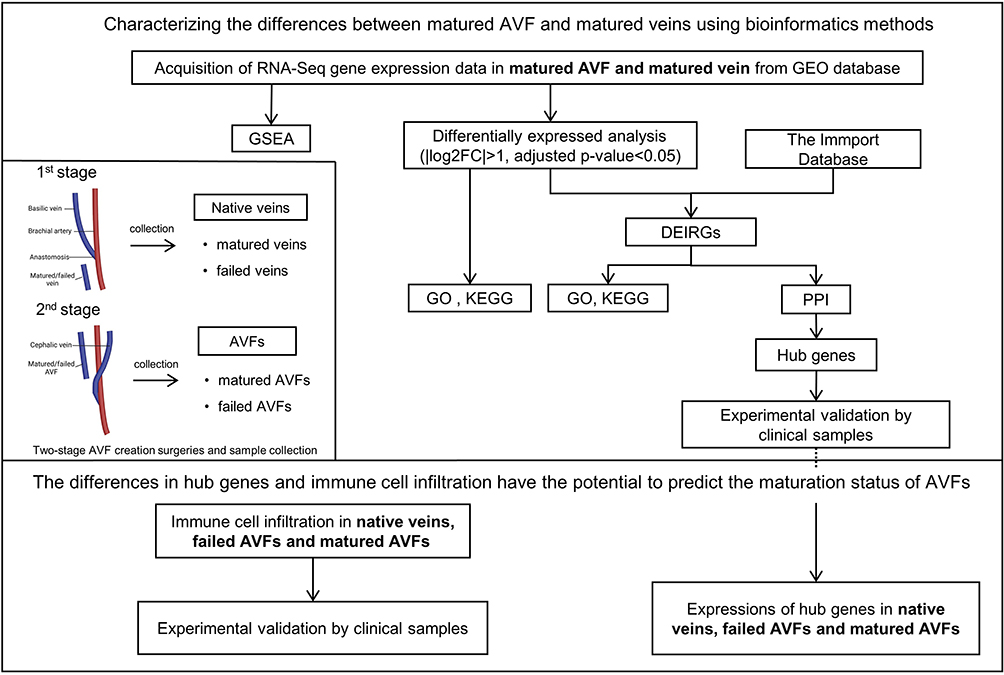 |
Figure 1 The flow diagram of the entire study. In the 1st stage surgery, native veins were collected, including matured veins and failed veins. Matured or failed vein refers to a vein that generated an arteriovenous fistula (AVF) that matured or failed, respectively. In the 2nd stage surgery, AVFs were obtained from the patients, including matured AVFs and failed AVFs. Matured or failed AVF refers to an AVF that matured or failed, respectively. Abbreviations: GEO, gene expression omnibus; GSEA, the gene set enrichment analysis; GO, gene ontology; KEGG, Kyoto encyclopedia of genes and genomes; DEIRGs, the differentially expressed immune-related genes; PPI, protein–protein interaction network. |
Materials and Methods
Data Source
We downloaded the datasets (GSE119296, GSE39488) from the GEO database (www.ncbi.nlm.nih.gov/geo). GSE119296 contained 38 samples’ RNA expression profiling, in which the native vein and AVF samples are related because they originate from the same individual and same anatomical location. We selected 10 matured AVF samples and 10 paired native vein samples from the GSE119296 profiling to perform differential expression analysis. GSE39488 profiling was utilized as the validation set, which includes six samples collected from the venous segments of matured AVF and four control samples collected from normal cephalic veins in hemodialysis patients.
Identification of Differentially Expressed Immune-Related Genes
Gene count data was input into DESeq2 package in R to standardize the number of counts in each sample (using the BaseMean value to estimate the expression level), calculate the FoldChange, and use negative binomial distribution test to test the significance of the difference in the number of reads. The criteria of statistically significant Differentially Expressed Genes (DEGs) were set as follows: adjusted p-value < 0.05 and |log2 fold change| >1. A volcano plot was mapped to present all of the DEGs using “ggplot2” packages. The immune-related genes (IRGs) were exported from the ImmPort database. The Differentially Expressed immune-related Genes (DEIRGs) are derived from the intersection of DEGs and IRGs.
Enrichment Analysis
Gene Ontology (GO) analysis was applied to identify the common functions of DEGs and DEIRGs, which consisted of biological process, cellular component and molecular function. The Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was applied to identify the significant pathways for DEGs and DEIRGs. The P < 0.05 was the criteria for statistically significant enriched GO term and KEGG pathways.
Gene Set Enrichment Analysis
The Gene Set Enrichment Analysis (GSEA) was investigated to explore the enriched pathways in two risk groups using the GSEA software provided by the Broad Institute. False discovery rate (FDR) q < 0.05 with p < 0.05 after performing 1000 permutations was determined to be statistically significant.
Analysis of Immune Cells Infiltration Using the CIBERSORTx
The ratio of the 22 immune cell subsets was calculated stepwise using CIBERSORTx according to the mRNA expression of immune signature genes in the AVFs and native veins.
Construction of Protein–Protein Interaction Network
We selected 40 up-regulated DEIRGs with the smallest adjusted p-value and submitted them into the online search tool in the STRING database (http://string-db.org) to create the protein–protein interaction (PPI) networks. The confident score ≥0.7 was set as the cutoff to obtain significant PPI networks.
Validation of Hub Genes in AVFs and Native Veins
This study was conducted in accordance with the Declaration of Helsinki, and approved by the Medical Ethics Committee of the Second Xiangya Hospital (NO. 2019-Clinical Research-82), and patients gave informed consent to use the samples. AVF samples were collected from patients undergoing AVF revision (Table S1). Native veins were pieces of the cephalic vein collected from patients who underwent first operation of AVF creation (Table S1). All clinical samples were from patients treated at the Second Xiangya Hospital.
Real-Time Quantitative Polymerase Chain Reaction
Total RNA was isolated from tissue by using trizol. RNA quality was confirmed using a NanoDrop spectrophotometer at the 260/280-nm ratio (Thermo Scientific, Wilmington, DE). Reverse transcription was performed using Evo M-MLV RT premix (Accurate Biotechnology, Hunan). Real-time polymerase chain reaction was performed using SYBR Green supermix (Bio-Rad) and LightCycler® 96 System (Roche).
Immunohistochemistry
Tissue sections were de-paraffined using xylene and a graded series of alcohols. Sections were heated in 0.01M citric acid buffer (pH 6.0) at 100°C for 20 minutes and cooled to room temperature. After washing three times by PBS, sections were blocked with 5% normal goat serum in PBS (pH 7.4) for 1 hour at room temperature. Sections were then incubated at 4°C overnight with the primary antibody anti-IL-6 (ab6672, abcam), anti-CXCR4 (ab181020, abcam), anti-IL1B (ab156791, abcam), anti-CD4 (ab133616, abcam), anti-CD11c (ab52632, abcam). Secondary antibodies were goat anti-rabbit a conjugated antibody (abcam). Sections were stained with 3,3-N-Diaminobenzidine Tertrahydrochloride. Mean IOD values were counted using Image-Pro-Plus software (National Institutes of Health).
Immunofluorescence
Tissue sections were de-paraffined and then heated in citric acid buffer (pH 6.0) at 100°C for 15 minutes for antigen retrieval. The sections were blocked with 5% bovine serum albumin for 60 min at room temperature, and then incubated overnight at 4°C with primary antibody anti-CD68 antibody (Abcam, ab955), anti-CD206 (ab64693, abcam), anti-iNOS (ab178945, abcam). After incubation, the sections were incubated with Alexa Fluoro secondary antibodies for 1 hour and stained with 4’,6-diamidino-2-phenylindole (DAPI) (P36935, Invitrogen) to stain cellular nuclei. Positively staining cells were counted per high power fields or measured the intensity. For negative controls for the antibodies, IgG isotype controls, negative tissue controls and endogenous tissue background controls were used.
Toluidine Blue Staining
0.02% toluidine blue solution acidified with acetic acid with a final pH of 3.0. Tissue sections were de-paraffined and stained with Toluidine Blue. Excess Toluidine Blue was rinsed off with distilled water. Sections were dehydrated using a series of increasing concentrations of ethanol and then cleared with xylene. Finally, neutral mounting medium was used to cover the sections. Under the microscope, cells showing a purple color and appearing as large, round, or oval-shaped are identified as mast cells.
Statistical Analysis
Data are represented as mean value ± SEM. All data were analyzed using Prism 8 software (GraphPad Software, Inc, La Jolla, CA). Statistical significance was determined using Student t test or ANOVA. P value<0.05 is considered statistically significant.
Results
Identification and Significant Enrichment Analysis of DEGs
We found a total of 2172 DEGs (adjusted p-value < 0.05 and |log2 fold change| >1), including 1417 up-regulated DEGs and 755 down-regulated DEGs. Volcano plot was utilized to visualize DEGs (Figure 2A). The DAVID database was used for the GO and KEGG analysis. GO enrichment analysis was performed on up-regulated DEGs (Figure 2B) and down-regulated DEGs (Figure 2C). For all up-regulated DEGs, biological process mainly focuses on extracellular matrix organization, immune response and inflammatory response; cellular component mainly focuses on extracellular region, proteinaceous extracellular matrix and plasma membrane; molecular function mainly focuses on extracellular matrix structural constituent, metalloendopeptidase activity and growth factor activity. KEGG analysis of all DEGs showed that DEGs are closely related to Cytokine-cytokine receptor interaction, Malaria, Osteoclast differentiation, Rheumatoid arthritis and IL-17 signaling pathway (Figure 2D; Table S2).
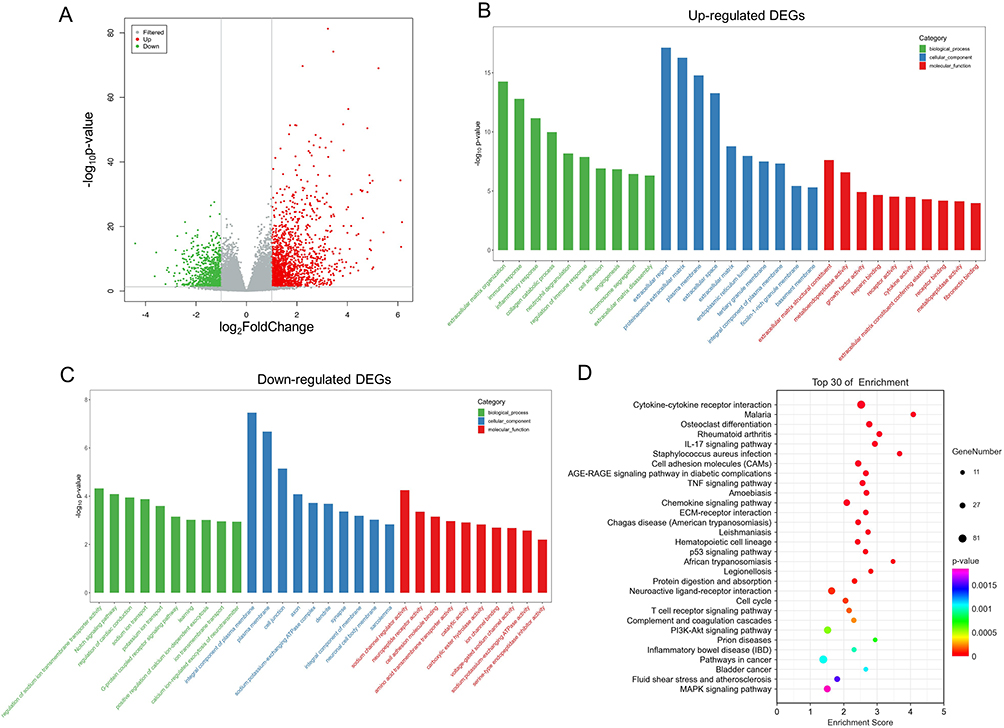 |
Figure 2 Identification and significant enrichment analysis of differentially expressed genes (DEGs). (A) Volcano map of DEGs between native veins and matured AVFs. The x-axis is the log2 scale of the fold change of gene expression in veins that matured and matured AVFs. Negative values indicate downregulation; positive values indicate upregulation. The y-axis is the minus log10 scale of the adjusted p values (–log10 (p-value)), which indicate the significant level of expression difference. The red dots represent significantly upregulated genes with at least twofold change, while the green dots represent significantly downregulated genes with at least twofold change. (B) The GO terms of up-regulated DEGs. (C) The GO terms of down-regulated DEGs. (D) The KEGG pathway terms of DEGs. Abbreviations: AVF, arteriovenous fistula; GO, gene ontology; KEGG, the Kyoto encyclopedia of genes and genomes. |
Gene Set Enrichment Analysis
For the GSEA of the pathways, MAPK signaling pathway, cytokine-cytokine receptor interaction, chemokine signaling pathway, JAK-STAT signaling pathway were significantly enriched in overflow AVFs (Figure 3). The complete significantly enriched KEGG pathway from GSEA are in the Table S3.
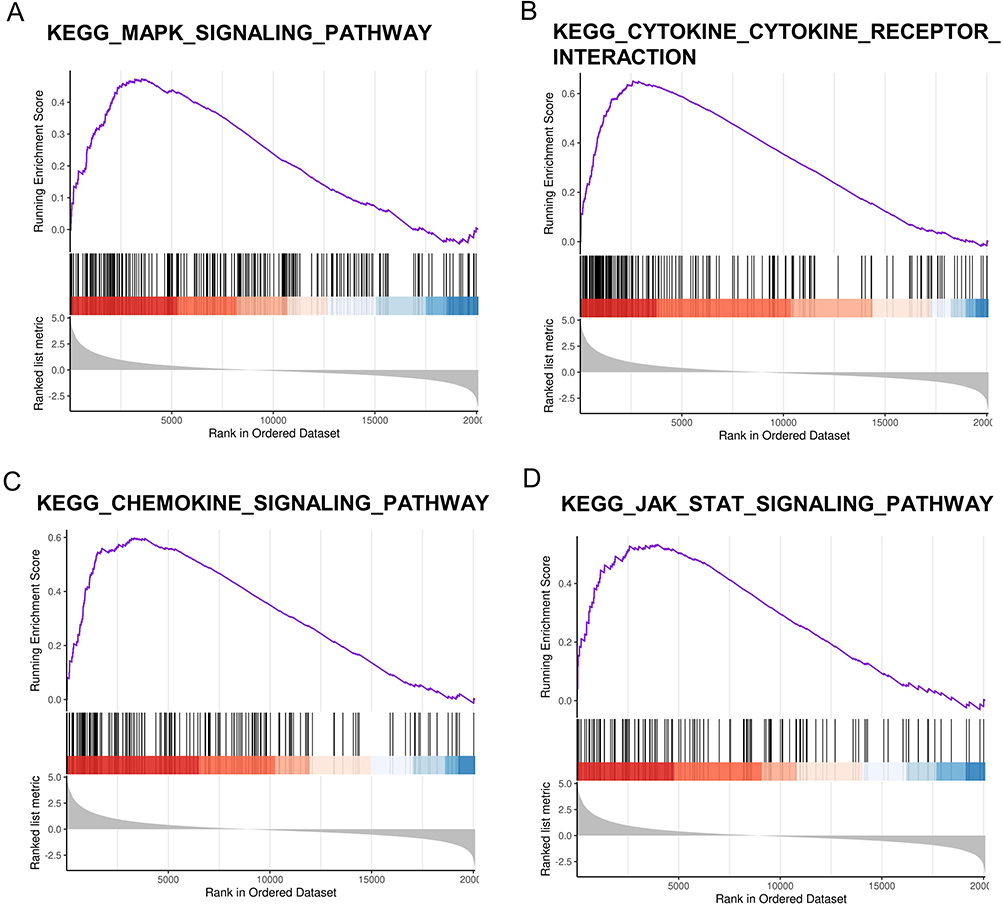 |
Figure 3 The enriched signaling pathway through the gene set enrichment analysis (GSEA). (A) MAPK signaling pathway. (B) cytokine-cytokine receptor interaction. (C) chemokine signaling pathway. (D) JAK-STAT signaling pathway. The gene set c2 (cp.kegg.v.6.2.symbols.gmt) database was used to analyze the whole gene expression value of the AVF and control samples. The cutoff criteria of the significant gene set were the absolute value of normalized enrichment score (NES) > 1, p-value < 0.05, and FDR < 0.05. The enrichment plots (A-D) represent four pathways that are significantly enriched. Abbreviation: AVF, arteriovenous fistula. |
Identification and Significant Enrichment Analysis of DEIRGs
The results of the enrichment analysis of DEGs were highly correlated with immune-related molecular functions and signaling pathways, as well as inflammation pathways. Since inflammation is often regulated by immune cells and immune molecules, we further identified 274 DEIRGs, including 225 up-regulated DEIRGs and 49 down-regulated DEIRGs. Volcano plot was utilized to visualize DEIRGs (Figure 4A). GO enrichment analysis (Figure 4B and C) and KEGG analysis (Figure 4D) were performed.
 |
Figure 4 Identification and significant enrichment analysis of DEIRGs. (A) Volcano map of the differentially expressed immune-related genes (DEIRGs) between matured AVFs and native veins. The x-axis is the log2 scale of the fold change of gene expression in matured AVFs and veins. Negative values indicate downregulation; positive values indicate upregulation. The y-axis is the minus log10 scale of the adjusted p values (–log10 (p-value)), which indicate the significant level of expression difference. The red dots represent significantly upregulated genes with at least twofold change, while the green dots represent significantly downregulated genes with at least twofold change. (B) The GO terms of up-regulated DEIRGs. (C) The GO terms of down-regulated DEIRGs. (D) The KEGG pathway terms of DEIRGs. Abbreviations: AVF, arteriovenous fistula; GO, gene ontology; KEGG, Kyoto encyclopedia of genes and genomes. |
PPI Network
We selected 40 up-regulated DEIRGs with the smallest adjusted p-value and submitted them into STRING database (the confidence score ≥0.7) to obtain a PPI network (Figure 5). There are 9 genes highly correlated with other genes, therefore we selected them as candidate hub geneses, including IL1B, IL6, IL10, CCL2, CXCR4, OSM, LIF, SOCS3, HBEGF.
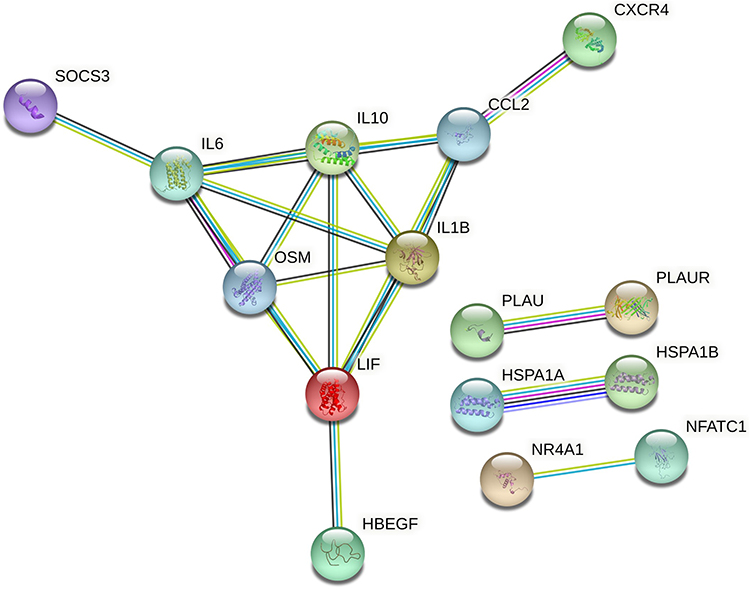 |
Figure 5 Protein–protein interaction (PPI) network of DEIRGs, threshold required confidence (combined score) > 0.7. |
Hub Gene Identification and Validation in Clinical Samples
We examined the expression of 9 candidate hub genes obtained from the PPI network in each group (native veins, failed AVFs, matured AVFs) in the GSE119296 dataset (Figure 6). The results showed that the gene expression increased sequentially in native vein, failed AVF, and matured AVF group. Then we identified the DEIRGs in GSE39488 dataset to verified 9 candidate hub genes (|log2FC|>2, adjusted p-value <0.1 (Table S4). After intersection, 4 candidate hub genes (CXCR4, IL1B, IL6, OSM) were identified. Then we used these 4 candidate hub genes for experimental verification.
 |
Figure 6 Candidate genes’ expression in veins, failed AVFs and matured AVFs. (A) Gene expression between veins that matured (n=10) and matured AVFs (n=10), (a-i) denote CCL2, CXCR4, OSM, LIF, SOCS3, HBEGF, IL1B, IL6, IL10. (B) Gene expression between veins (n=16) and AVFs (n=16), (a-i) denote CCL2, CXCR4, OSM, LIF, SOCS3, HBEGF, IL1B, IL6, IL10. (C) Gene expression between veins (n=16), failed AVFs (n=6) and matured AVFs (n=10), (a-i) denote CCL2, CXCR4, OSM, LIF, SOCS3, HBEGF, IL1B, IL6, IL10. AVF, arteriovenous fistula. Significant differences are indicated by *P<0.05, **P<0.01, ***P<0.001. |
To further validate the results of the bioinformatics analysis, we performed verification (qRT-PCR and IHC) using clinical samples. We collected AVF samples from uremic patients as the experimental group, native cephalic vein samples as the control group. Demographic data of patients showed no statistical differences in all clinical characteristics between the two groups. The results of qPCR proved that the expression of hub genes in the AVF group were significantly higher than that in the native vein group, and the Statistical significance existed in the three groups of IL1B, IL6, and CXCR4 (Figure 7). Compared to native veins, the clinical samples we used exhibit distinct AVF characteristics, indicating that the samples are indeed AVF tissues (Figure 8A and B). We subsequently performed IHC to assess the expression of IL1B, IL6, and CXCR4 in native vein and AVF (Figure 8C–G). We found that IL1B, IL6 and CXCR4 expression was significantly higher in AVF tissues than in native veins.
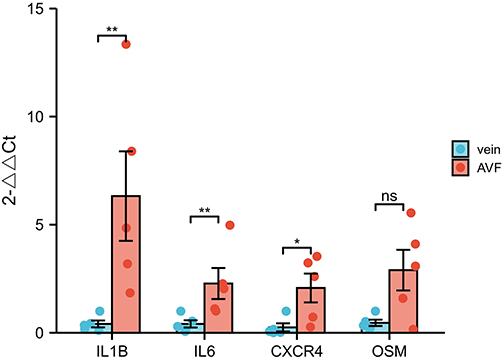 |
Figure 7 Gene expression of IL-1B, IL6, CXCR4, OSM by qRT-PCR in AVFs and veins. Gene expression of IL1B, IL6 and CXCR4 was significantly higher in the AVF group compared to the vein group, while OSM expression was not statistically different. Significant differences are indicated by *P<0.05, **P<0.01. |
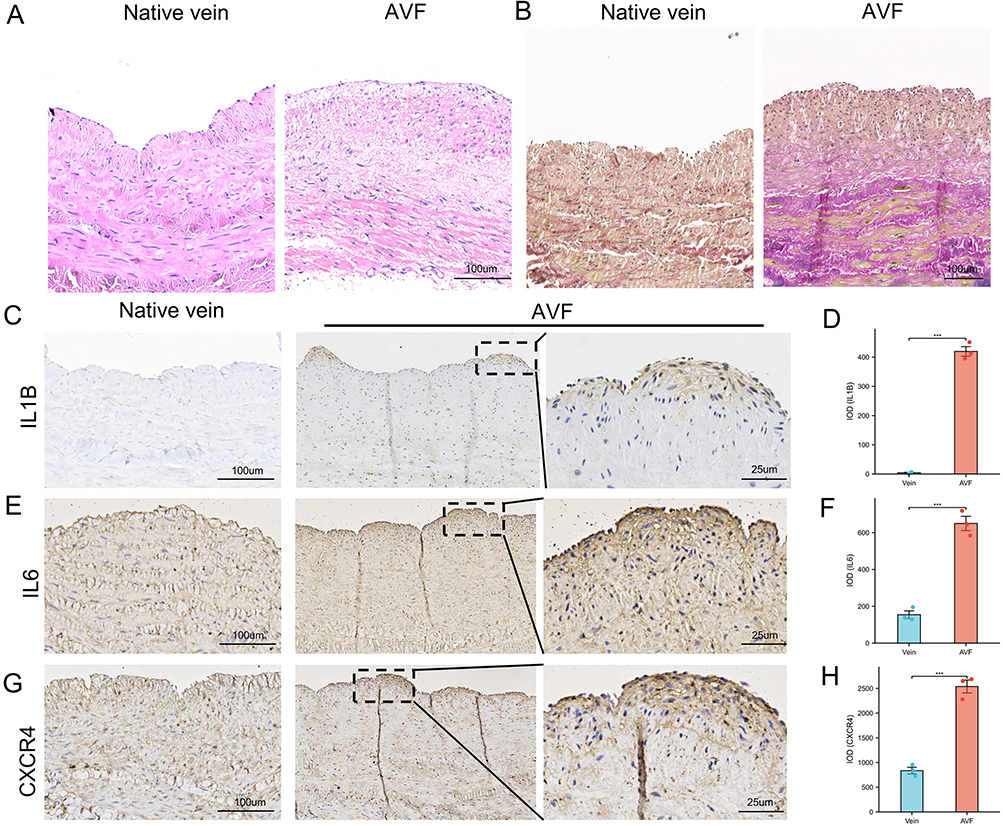 |
Figure 8 The result of immunohistochemistry staining showed that the expression of IL6, CXCR4 and IL1B were increased in AVFs. (A) The Hematoxylin and Eosin staining of native vein and AVF. (B) The Elastica van Gieson (EVG) staining of native vein and AVF. (C) The IHC result of IL1B in native veins and AVFs. (D) Box plot was used to show that the expression of IL1B significantly increased in AVF. (E) the IHC result of IL6 in native veins and AVFs. (F) Box plot was used to show that the expression of IL6 significantly increased in AVF. (G) the IHC result of CXCR4 in native veins and AVFs. (H) Box plot was used to show that the expression of CXCR4 significantly increased in AVF. Significant differences are indicated by ***P<0.001. Abbreviation: AVF, arteriovenous fistula. |
Immune Cell Infiltration and Validation in Clinical Samples
The infiltration of immune cells was analyzed between native veins and AVFs. According to the results, the bar chart and heatmap illustrate the proportions of infiltrated immune cells (Figure 9A and B). By comparing the differences in immune cell infiltration between matured veins and matured AVFs, we found that the proportions of four types of immune cells (T-cell CD4 memory resting, dendritic cells activated, mast cells activated, and Macrophages M2) were significantly elevated during the conversion of mature veins to matured AVFs (Figure 9C–F). Interestingly, we observed that compared to matured AVFs, the infiltration of T-cell CD4 memory resting and Macrophages M2 did not show a stable increasing trend during the conversion of failed veins to failed AVFs. Additionally, although the proportions of dendritic cells activated and mast cells activated showed an increasing trend during the conversion of failed veins to failed AVFs, the magnitude was lower than that observed during the conversion from matured veins to matured AVFs. Importantly, we found that mast cells activated were completely absent in native veins but present in a certain number in AVFs (Figure 9E). The proportions of T-cell CD4 memory resting, dendritic cells activated, and mast cells activated increased sequentially in native veins, failed AVFs, and matured AVFs (Figure 9G–I). Compared to native veins, the proportions of Macrophages M2 exhibited a disordered trend in failed AVFs, while a consistent increasing trend was observed in matured AVFs (Figure 9J). We partially validated these findings using clinical samples. We observed increased infiltration of CD4+ T cells, toluidine blue-stained cells (Mast cells), CD11c+ cells (dendritic cells), and CD68 and CD206 dual-positive cells (Macrophages M2) in AVFs tissue (Figure 10).
 |
Figure 9 Immune infiltration analysis of native veins, failed AVFs and matured AVFs. (A) Histograms show the relative proportion of 22 immune cell in native veins, failed AVFs and matured AVFs. Each column represents a sample, and each color represents a cell type. (B) Clustered heatmap on the expression of 22 immune cells in tissues of native veins, failed AVFs and matured AVFs, with red indicating a greater percentage. (C) Infiltration of T-cell CD4 memory resting was significantly elevated during conversion of mature veins to AVFs, whereas failed AVFs lacked infiltration of T-cell CD4 memory resting. (D) Infiltration of dendritic cells activated was significantly elevated during conversion of both mature veins and failed veins to AVFs. (E) Infiltration of Mast cells activated was significantly elevated during conversion of both mature veins and failed veins to AVFs, but the increase was more pronounced in the matured group. (F) Infiltration of Macrophages M2 was significantly elevated during conversion of mature veins to AVFs, whereas failed AVFs lacked infiltration of Macrophages M2. (G) Relative abundance of T cells CD4 memory resting between veins (n=16), failed AVFs (n=6) and matured AVFs (n=10). (H) Relative abundance of Dendritic cells activated between veins (n=16), failed AVFs (n=6) and matured AVFs (n=10). (I) Relative abundance of Mast cells activated between veins (n=16), failed AVFs (n=6) and matured AVFs (n=10). (J) Relative abundance of Macrophages M2 between veins (n=16), failed AVFs (n=6) and matured AVFs (n=10). Significant differences are indicated by *P<0.05, **P<0.01, ***P<0.001. Abbreviation: AVF, arteriovenous fistula. |
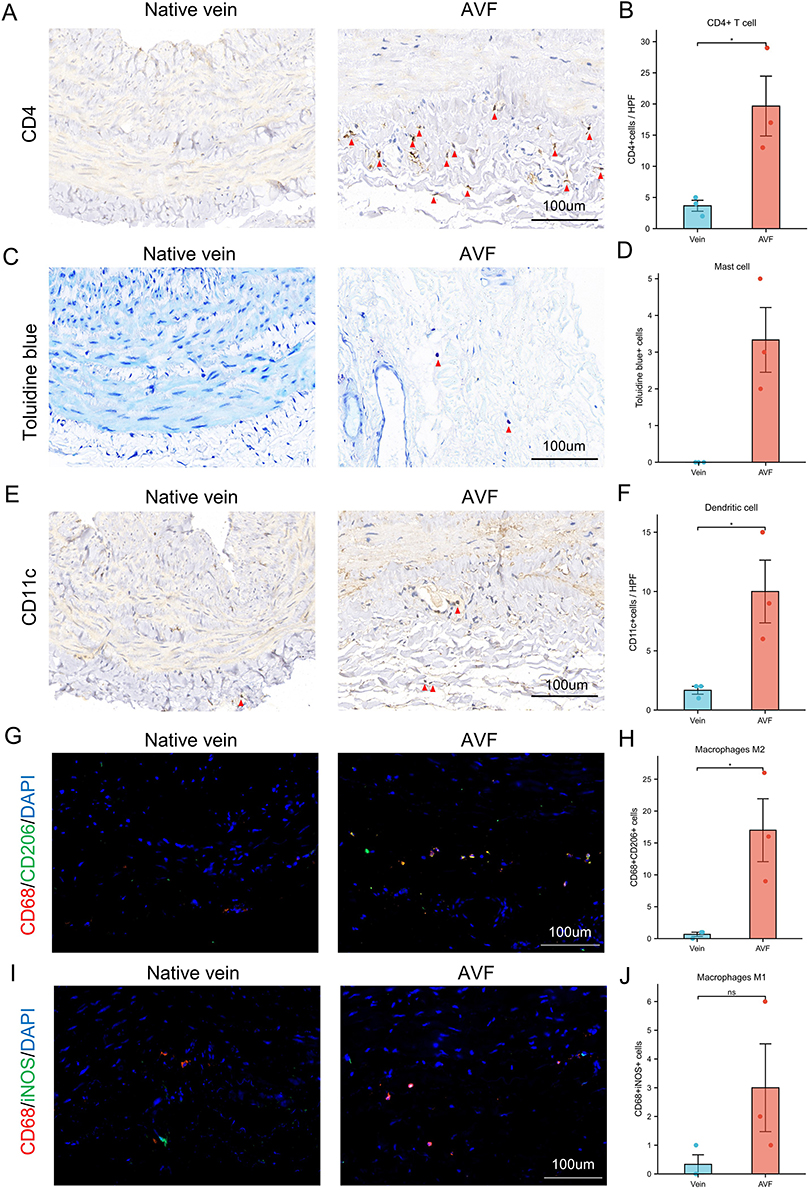 |
Figure 10 Validation of the expression of four immune cells in native veins and AVFs. (A) The immunohistochemistry (IHC) result of CD4 in native veins and AVFs. Red triangles indicate CD4+ cells. (B) Box plot indicates significant infiltration of CD4+ T cells in AVFs. (C) The toluidine blue staining in native veins and AVFs. Purple cells shown in red triangles are positive cells and can be identified as mast cells when combined with staining results and cell morphology. (D) Box plot indicates significant infiltration of Mast cells in AVF. (E) The IHC result of CD11c in native veins and AVFs. Red triangles indicate CD11c+ cells. (F) Box plot indicates significant infiltration of dendritic cell in AVF. (G) Immunofluorescence staining of CD68 and CD206 in native veins and AVFs. Yellow cells are double-positive for CD68 and CD206, indicating M2 macrophages. (H) Box plot indicates significant infiltration of M2 macrophages in AVFs. (I) Immunofluorescence staining of CD68 and iNOS in native veins and AVFs. Yellow cells are double-positive for CD68 and iNOS, indicating M1 macrophages. (J) Box plot indicates the infiltration of M1 macrophages in native veins and AVFs. Significant differences are indicated by *P<0.05. Abbreviation: AVF, arteriovenous fistula. |
Discussion
As the failed arteriovenous fistula (AVF) typically does not lead to acute patient mortality, and the failed AVF or those that fail after a period of use often require interventional treatment due to the demands of AVF surgical techniques, it becomes challenging to obtain the patient’s accurate tissue at the follow-up endpoint in clinical settings.2 Only the 2-stage AVF creation surgery approach can obtain patient tissues before and after AVF creation. Currently, a rare study recruited 38 patients over a four-year period (19 samples of mature AVFs and 19 samples of failed AVFs) and identified the Hallmarks of Maturation Failure through transcriptome analysis.17 However, it is also unclear which immune-related genes or immune cells may have potential predictive roles in the maturation of AVF.
In this study, we used the GSE119296 dataset (samples from two-stage AVF creation surgeries) to compare paired native veins and mature AVFs which originate from the same individual and same anatomical location to identify differentially expressed genes (DEGs). Then we performed GO and KEGG enrichment analysis on these differentially expressed genes, we found that immune-related signaling pathways were mainly enriched, as well as inflammation pathways. GSEA was used to explore the potentially activated signaling pathways with the advantage of avoiding undetectable, small changes in the expression of single genes. Due to the significant enrichment of immune and inflammation-related pathways, we selected all differentially expressed immune-related genes (DEIRGs) and performed enrichment analysis again. PPI network analysis was performed on DEIRGs to select candidate hub genes. Then we detected their expression in the GSE39488 dataset, and finally identified 4 hub genes. We did verification in Clinical Samples by qPCR and IHE. Finally, we depicted the immune cell infiltration in native veins, failed AVFs, and mature AVFs, and partially validated it in clinical samples. We found that 1) immune-related pathways and inflammation-related pathways were abundantly enriched, indicating that they may play an important role in the AVF maturation; 2) we identified 3 immune-related hub genes, which may be involved in the maturation of AVF, and the results of qRT-PCR and IHC by clinical samples validated our bioinformatic results; 3) we found that certain immune cells (T-cell CD4 memory resting, activated dendritic cells, activated mast cells, and Macrophages M2) have an impact on the maturation of AVF and may have potential predictive roles.
Enrichment analysis of DEGs indicated that activation of immune-related pathways and immune-related genes are indispensable in the maturation of AVFs. We found that immune response and inflammatory response are the main biological process of GO terms (Figure 2B and C). In addition, a number of immune-related signaling pathways were enriched (Figures 2D, 3), such as IL17 signaling pathway and T cell receptor signaling pathway. Some current studies about immunity and inflammation mediating AVF maturation consistent with our research results. The latest study found that circulating IL6 and CD16 positive monocytes increase following angioplasty of AVF.18 Interestingly, the inflammatory response does not seem to be more favorable for the maturation of AVF. Systemic inflammation correlates inversely with AVF maturation, for example, the plasma levels of C-reactive protein are inversely correlated with successful AVF maturation.19 In addition, glucocorticoids, a well-known anti-inflammatory drug, can enhance venous outward remodeling in a murine arteriovenous fistula model.15 However, local glucocorticoids did not seem to work in humans, although negative results may be due to lack of power or drug dosage/delivery.20 These studies are consistent with some of our results, and in addition, our results provide other potential directions for studying the relationship between immunity, inflammation, and AVF maturation.
Furthermore, the immune cell infiltration in AVFs shows that the proportion of T-cell CD4 memory resting, activated dendritic cells, activated mast cells, and Macrophages M2 increased significantly compared to native veins. The results of a previous study showed significant infiltration of mast cells, neutrophils, monocytes, and macrophages around the arteriovenous fistula at day 7.21 Previous studies have confirmed that AVF maturation requires inflammation that is characterized by accumulation of macrophages and T-cell.22–25 M2 macrophages are associated with vascular wall thickening, and promoting the accumulation of M2 macrophages may promote AVF maturation.22 Inhibition of T-cells by cyclosporine A reduces macrophage accumulation to regulate venous adaptive remodeling and increase arteriovenous maturation.24 Inhibition of PD-L1 reduced regulatory T cells to increase M1 macrophages and reduce M2 macrophages, reducing vascular wall thickening and AVF patency.26 The absence of mature CD4+ T-cells results in reduced macrophage infiltration resulting in failure of AVF maturation.27 These findings are consistent with our results. However, it is interesting to note that we observed that compared to matured AVFs, the infiltration of T-cell CD4 memory resting and Macrophages M2 did not show a stable increasing trend during the conversion of failed veins to failed AVFs (Figure 9). This suggests that inadequate infiltration of CD4+ T-cells and M2 macrophages could be a factor leading to the failure of AVF maturation. We validated the high infiltration of CD4+ T-cells and M2 macrophages in clinical samples (Figure 10). The impact of dendritic cells and mast cells on AVF maturation is currently not well understood. Our research found that during the transition from native vein to AVF, the infiltration of these two cell types increases (Figures 9 and 10). However, compared to failed AVF, both dendritic cells and mast cells show higher infiltration in matured AVF. This observation suggests that these two cell types may be involved in the process of AVF maturation. It is important to note that the infiltration of immune cells was achieved through algorithms and may not fully reflect the actual immune cell infiltration in the tissues.28 Further single-cell research is necessary to corroborate these findings and provide a more accurate understanding of the immune cell infiltration in AVF maturation.
Through bioinformatics analysis, we obtained 3 significantly up-regulated immune-related hub genes, namely IL1B, IL6 and CXCR4, which were validated by qRT-PCR and IHE using clinical samples (Figures 8 and 9). IL1B, a potent proinflammatory cytokine, initially discovered as the major endogenous pyrogen, induces neutrophil influx and activation, T-cell activation and cytokine production, B-cell activation and antibody production, as well as fibroblast proliferation and collagen production.29 IL1B also plays a role in angiogenesis by inducing VEGF production synergistically with TNF and IL6.30 IL1B promotes cell migration and angiogenesis in tumors,31 and induces chemokine expression.32 These studies allow to speculate that IL1B-mediated inflammatory cell chemotaxis and VEGF production, among others, might play a role in the maturation of the venous wall after AVF surgery. The results of our study likewise support this hypothesis. However, excessive expression of IL1B may lead to a more pronounced inflammatory response. It may be a viable treatment option to use PCSK9 inhibitors, which have a cardiovascular protective effect, to promote VEGFA expression while mitigating the pro-inflammatory effects of high IL1B.33,34 Further studies are needed to confirm the role of IL1B in the maturation of arteriovenous fistula.
IL6 is generally considered a pro-inflammatory factor and is involved in many immune regulations. In the innate immune response, IL6 is synthesized by myeloid cells, such as macrophages.35 IL6 plays a major role in the differentiation of CD4+ T cell subsets and is also an essential factor for the T follicular helper cells.35 The Accumulation of macrophages and CD4+ T-cells has an important impact on AVF maturation.23,24 IL6 also induces the production of VEGF synergistically with IL1B and TNF through “trans-signaling”.30 The role of IL6 in the maturation of AVF remains controversial. Circulating IL-6 was significantly higher in patients with AVF maturation failure than with successful maturation.36 Studies have found that circulating IL6 increases following angioplasty of AVF (1–2 days).18 However, other studies hold the opposite conclusion. They found that IL6 in the adipose tissue around the fistula vein were associated with a reduction in the fractional vein diameter during the early time frame of 1 day to 2 weeks,37 which shows that IL6 appears to have an early negative effect after AVF creation. In conclusion, the effect of IL6 on AVF maturation has been demonstrated, which is consistent with our experimental results.
CXCR4 is one of the C-X-C chemokine receptors which involved in various immunity and inflammatory responses. IL6 is best known as a coreceptor (CD4 being the primary receptor) for human immunodeficiency virus-1/HIV-1 X4 isolates and as a primary receptor for some HIV-2.38 CXCR4 is a receptor for the C-X-C chemokine CXCL12 /SDF-1, and is involved in the enhancing of MAPK1/MAPK3 activation and the AKT signaling cascade.39 These signaling pathways have been shown to play an important role in AVF maturation. The role of chemokines in AVF maturation has been demonstrated. For instance, CX3CR1 blockade has been found to reduce VNH/VS formation by decreasing proinflammatory cues.40 Previous bioinformatics analysis also identified CXCR4 as hub gene for AVF maturation41 This is in agreement with our finding. In summary, the role of CXCR4 in AVF maturation deserves to be further investigated.
Conclusion
Due to the unique Two-stage AVF creation surgeries, which allow for the acquisition of preoperative native vein and postoperative 30-day AVF samples from the same individual and anatomical location, clinical physicians can obtain native veins, failed AVFs, and matured AVFs. These samples are highly valuable for investigating factors influencing AVF maturation. We collected the high-throughput sequencing results from these samples and conducted a series of experiments using bioinformatics methods. Additionally, we partially validated our findings using clinical samples. In conclusion, our findings indicate significant differences in immune-related genes and immune cell infiltration between failing and mature AVFs. We have identified three immune-related genes and several immune cell types that potentially predict AVF maturation outcomes. Our study also suggests that immunity and inflammation may play a more important role in AVF maturation than is currently recognized.
Abbreviations
AVF, Arteriovenous fistula; GO, gene ontology; KEGG, Kyoto encyclopedia of genes and genomes pathway enrichment analysis; GSEA, gene set enrichment analysis; DEGs, differentially expressed genes; DEIRGs, differentially expressed immune-related genes; CKD, chronic kidney disease; ESRD, end-stage renal disease; PPI, protein-protein Interaction.
Acknowledgments
We thank Professor Chengyuan, Tang for his help in the design of this experiment.
Funding
This research was funded by International (Regional) Cooperation and Exchange (ICE) Projects of the National Natural Science Foundation of China (NSFC) (grant number 82120108005, National Natural Science Foundation of China (grant number 81900695) and Natural Science Foundation of Hunan Province of China (grant number 2021JJ40874).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Chen TK, Knicely DH, Grams ME. Chronic Kidney Disease Diagnosis and Management: a Review. JAMA. 2019;322(13):1294–1304. doi:10.1001/jama.2019.14745
2. Daugirdas JT, Depner TA, Inrig J. KDOQI Clinical Practice Guideline for Hemodialysis Adequacy: 2015 update. Am J Kidney Dis. 2015;66(5):884–930. doi:10.1053/j.ajkd.2015.07.015
3. Hu H, Patel S, Hanisch JJ, et al. Future research directions to improve fistula maturation and reduce access failure. Semin Vasc Surg. 2016;29(4):153–171. doi:10.1053/j.semvascsurg.2016.08.005
4. Robbin ML, Greene T, Allon M, et al. Prediction of Arteriovenous Fistula Clinical Maturation from Postoperative Ultrasound Measurements: findings from the Hemodialysis Fistula Maturation Study. J Am Soc Nephrol. 2018;29(11):2735–2744. doi:10.1681/asn.2017111225
5. Bashar K, Zafar A, Elsheikh S, et al. Predictive parameters of arteriovenous fistula functional maturation in a population of patients with end-stage renal disease. PLoS One. 2015;10(3):e0119958. doi:10.1371/journal.pone.0119958
6. Libetta C, De Nicola L, Rampino T, De Simone W, Memoli B. Inflammatory effects of peritoneal dialysis: evidence of systemic monocyte activation. Kidney Int. 1996;49(2):506–511. doi:10.1038/ki.1996.72
7. Cavaillon JM, Poignet JL, Fitting C, Delons S. Serum interleukin-6 in long-term hemodialyzed patients. Nephron. 1992;60:307–313. doi:10.1159/000186770
8. Donati D, Degiannis D, Combates N, Raskova J, Raska K Jr. Effects of hemodialysis on activation of lymphocytes: analysis by an in vitro dialysis model. J Am Soc Nephrol. 1992;2(10):1490–1497. doi:10.1681/ASN.V2101490
9. Memoli B, Libetta C, Rampino T, et al. Hemodialysis related induction of interleukin-6 production by peripheral blood mononuclear cells. Kidney Int. 1992;42(2):320–326. doi:10.1038/ki.1992.292
10. Honkanen E, Grönhagen-Riska C, Teppo AM, Maury CP, Meri S. Acute-phase proteins during hemodialysis: correlations with serum interleukin-1 beta levels and different dialysis membranes. Nephron. 1991;57:283–287. doi:10.1159/000186276
11. Cooke JP. Inflammation and Its Role in Regeneration and Repair. Circ Res. 2019;124(8):1166–1168. doi:10.1161/circresaha.118.314669
12. Wynn TA, Vannella KM. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity. 2016;44(3):450–462. doi:10.1016/j.immuni.2016.02.015
13. Martínez-Martínez E, Miana M, Jurado-López R, et al. A role for soluble ST2 in vascular remodeling associated with obesity in rats. PLoS One. 2013;8(11):e79176. doi:10.1371/journal.pone.0079176
14. Bezhaeva T, Wong C, de Vries MR, et al. Deficiency of TLR4 homologue RP105 aggravates outward remodeling in a murine model of arteriovenous fistula failure. Sci Rep. 2017;7(1):10269. doi:10.1038/s41598-017-10108-4
15. Wong C, Bezhaeva T, Rothuizen TC, et al. Liposomal prednisolone inhibits vascular inflammation and enhances venous outward remodeling in a murine arteriovenous fistula model. Sci Rep. 2016;6(1):30439. doi:10.1038/srep30439
16. Martinez L, Tabbara M, Duque JC, et al. Transcriptomics of Human Arteriovenous Fistula Failure: genes Associated with Nonmaturation. Am J Kidney Dis. 2019;74(1):73–81. doi:10.1053/j.ajkd.2018.12.035
17. Martinez L, Rojas MG, Tabbara M, et al. The Transcriptomics of the Human Vein Transformation After Arteriovenous Fistula Anastomosis Uncovers Layer-Specific Remodeling and Hallmarks of Maturation Failure. Kidney Int Rep. 2023;8(4):837–850. doi:10.1016/j.ekir.2023.01.008
18. Hakki S, Robinson EJ, Robson MG. Circulating Interleukin-6 and CD16 positive monocytes increase following angioplasty of an arteriovenous fistula. Sci Rep. 2022;12(1):1427. doi:10.1038/s41598-022-05062-9
19. Kaygin MA, Halici U, Aydin A, et al. The relationship between arteriovenous fistula success and inflammation. Ren Fail. 2013;35(8):1085–1088. doi:10.3109/0886022x.2013.815100
20. Voorzaat BM, van der Bogt KEA, Bezhaeva T, et al. A Randomized Trial of Liposomal Prednisolone (LIPMAT) to Enhance Radiocephalic Fistula Maturation: a Pilot Study. Kidney Int Rep. 2020;5(8):1327–1332. doi:10.1016/j.ekir.2020.05.030
21. Shehadeh SA, Tabbara M, Martinez L, Vazquez-Padron RI. A snapshot of early venous remodeling in a 7-day-old arteriovenous fistula. J Vasc Access. 2022;112972982210917. doi:10.1177/11297298221091757
22. Kuwahara G, Hashimoto T, Tsuneki M, et al. CD44 Promotes Inflammation and Extracellular Matrix Production During Arteriovenous Fistula Maturation. Arterioscler Thromb Vasc Biol. 2017;37(6):1147–1156. doi:10.1161/atvbaha.117.309385
23. Roy-Chaudhury P, Khan R, Campos B, et al. Pathogenetic role for early focal macrophage infiltration in a pig model of arteriovenous fistula (AVF) stenosis. J Vasc Access. 2014;15(1):25–28. doi:10.5301/jva.5000151
24. Matsubara Y, Kiwan G, Liu J, et al. Inhibition of T-Cells by Cyclosporine A Reduces Macrophage Accumulation to Regulate Venous Adaptive Remodeling and Increase Arteriovenous Fistula Maturation. Arterioscler Thromb Vasc Biol. 2021;41(3):e160–e174. doi:10.1161/atvbaha.120.315875
25. Matsubara Y, Kiwan G, Fereydooni A, Langford J, Dardik A. Distinct subsets of T cells and macrophages impact venous remodeling during arteriovenous fistula maturation. JVS Vasc Sci. 2020;1:207–218. doi:10.1016/j.jvssci.2020.07.005
26. Matsubara Y, Gonzalez L, Kiwan G, et al. PD-L1 (Programmed Death Ligand 1) Regulates T-Cell Differentiation to Control Adaptive Venous Remodeling. Arterioscler Thromb Vasc Biol. 2021;41(12):2909–2922. doi:10.1161/atvbaha.121.316380
27. Duque JC, Martinez L, Mesa A, et al. CD4(+) lymphocytes improve venous blood flow in experimental arteriovenous fistulae. Surgery. 2015;158(2):529–536. doi:10.1016/j.surg.2015.02.018
28. Rusk N. Expanded CIBERSORTx. Nat Methods. 2019;16:577. doi:10.1038/s41592-019-0486-8
29. Van Damme J, De Ley M, Opdenakker G, et al. Homogeneous interferon-inducing 22K factor is related to endogenous pyrogen and interleukin-1. Nature. 1985;314(6008):266–268. doi:10.1038/314266a0
30. Nakahara H, Song J, Sugimoto M, et al. Anti-interleukin-6 receptor antibody therapy reduces vascular endothelial growth factor production in rheumatoid arthritis. Arthritis Rheum. 2003;48(6):1521–1529. doi:10.1002/art.11143
31. Lee C-H, Chang JS-M, Syu S-H, et al. IL-1β promotes malignant transformation and tumor aggressiveness in oral cancer. J Cell Physiol. 2015;230(4):875–884. doi:10.1002/jcp.24816
32. Song H, Chan J, Rovin BH. Induction of chemokine expression by adiponectin in vitro is isoform dependent. Transl Res. 2009;154(1):18–26. doi:10.1016/j.trsl.2009.04.003
33. Hrovat K, Rehberger Likozar A, Zupan J, Šebeštjen M. Gene Expression Profiling of Markers of Inflammation, Angiogenesis, Coagulation and Fibrinolysis in Patients with Coronary Artery Disease with Very High Lipoprotein(a) Levels Treated with PCSK9 Inhibitors. J Cardiovasc Dev Dis. 2022;9. doi:10.3390/jcdd9070211
34. Kheirkhah A, Lamina C, Kollerits B, et al. PCSK9 and Cardiovascular Disease in Individuals with Moderately Decreased Kidney Function. Clin J Am Soc Nephrol. 2022;17(6):809–818. doi:10.2215/cjn.01230122
35. Kang S, Tanaka T, Narazaki M, Kishimoto T. Targeting Interleukin-6 Signaling in Clinic. Immunity. 2019;50(4):1007–1023. doi:10.1016/j.immuni.2019.03.026
36. Martinez L, Perla M, Tabbara M, et al. Systemic Profile of Cytokines in Arteriovenous Fistula Patients and Their Associations with Maturation Failure. Kidney. 2022;360(3):677–686. doi:10.34067/kid.0006022021
37. Sharma G, Kuppler C, He Y, et al. Local Adipose-Associated Mediators and Adaptations Following Arteriovenous Fistula Creation. Kidney Int Rep. 2018;3(4):970–978. doi:10.1016/j.ekir.2018.02.008
38. Doranz BJ, Orsini MJ, Turner JD, et al. Identification of CXCR4 domains that support coreceptor and chemokine receptor functions. J Virol. 1999;73(4):2752–2761. doi:10.1128/jvi.73.4.2752-2761.1999
39. Cao Y, Hunter ZR, Liu X, et al. The WHIM-like CXCR4(S338X) somatic mutation activates AKT and ERK, and promotes resistance to ibrutinib and other agents used in the treatment of Waldenstrom’s Macroglobulinemia. Leukemia. 2015;29(1):169–176. doi:10.1038/leu.2014.187
40. Misra S, Kilari S, Yang B, et al. Anti Human CX3CR1 VHH Molecule Attenuates Venous Neointimal Hyperplasia of Arteriovenous Fistula in Mouse Model. J Am Soc Nephrol. 2021;32(7):1630–1648. doi:10.1681/asn.2020101458
41. Zhao Z, Fu Q, Hu L, Liu Y. Identification of the Crucial Gene in Overflow Arteriovenous Fistula by Bioinformatics Analysis. Front Physiol. 2021;12:621830. doi:10.3389/fphys.2021.621830
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Elucidating the Gene Signatures and Immune Cell Types in HIV-Infected Immunological Non-Responders by Bioinformatics Analyses
Xiao Q, Han J, Yu F, Yan L, Li Q, Lao X, Zhao H, Zhang F
International Journal of General Medicine 2022, 15:8491-8507
Published Date: 7 December 2022
Fusing WGCNA and Machine Learning for Immune-Related Gene Prognostic Index in Lung Adenocarcinoma: Precision Prognosis, Tumor Microenvironment Profiling, and Biomarker Discovery

He J, Luan T, Zhao G, Yang Y
Journal of Inflammation Research 2023, 16:5309-5326
Published Date: 16 November 2023
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024
Single-Cell Sequencing Combined with Transcriptome Sequencing to Explore the Molecular Mechanisms Related to Psoriasis
E C, Wang R, Meng Z, Zou Y
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2197-2213
Published Date: 3 October 2024
Transcriptomic Changes and Pathological Mechanisms in Familial and Sporadic Idiopathic Restless Legs Syndrome: Implications for Inflammation and Cell Adhesion Molecules
He XR, Song JM, Zhao JP, Zhang J, Feng JT, Chen SQ, Zhou ZY, Wang HM, Zhang Y, Feng Y, Wu YC, Zhu XY
Nature and Science of Sleep 2025, 17:1231-1247
Published Date: 11 June 2025