Back to Journals » Journal of Inflammation Research » Volume 14
IL-33/ST2 Axis Deficiency Exacerbates Hepatic Pathology by Regulating Treg and Th17 Cells in Murine Schistosomiasis Japonica
Authors Bai Y, Guan F, Zhu F, Jiang C, Xu X, Zheng F, Liu W, Lei J
Received 27 August 2021
Accepted for publication 27 October 2021
Published 15 November 2021 Volume 2021:14 Pages 5981—5998
DOI https://doi.org/10.2147/JIR.S336404
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Monika Sharma
Yang Bai,1,* Fei Guan,1,* Feifan Zhu,1 Chunjie Jiang,1 XiaoXiao Xu,1 Fang Zheng,2 Wenqi Liu,1 Jiahui Lei1
1Department of Pathogen Biology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430030, People’s Republic of China; 2Department of Immunology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430030, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jiahui Lei
Department of Pathogen Biology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430030, People’s Republic of China
Tel +86-13329722645
Fax +86-27-83691246
Email [email protected]
Purpose: Schistosoma japonicum-infected IL-33 and ST2 gene deficiency (IL-33−/− and ST2−/−, respectively) mice were used to explore the role of the IL-33/ST2 axis in liver pathology targeting regulatory T cells (Treg)/T helper 17 cells (Th17).
Materials and Methods: Each mouse was infected percutaneously with 20 S. japonicum cercariae. Hepatic mass index (HMI), liver egg granulomas, hepatic fibrosis biomarkers and serum levels of alanine aminotransferase (ALT) were investigated. Treg and Th17 frequency was determined by flow cytometry. Expressions of Foxp3, ST2, TGF-β 1, IL-10, RORγt, and IL-17A were measured via quantitative real-time polymerase chain reaction (qRT-PCR). Concentrations of TGF-β 1, IL-10 and IL-17A were tested with ELISA. In vitro experiments, mRNA expressions of Foxp3, TGF-β 1, IL-10, Atg5, Beclin-1 and p62 associated with polarization of Treg by recombinant mouse IL-33 (rmIL-33) were detected by qRT-PCR.
Results: An increased expression of IL-33/ST2 was shown in S. japonicum-infected mice. Deficiency of IL-33 or ST2 gene led to an aggravated liver pathology, which was evidenced by elevated hepatic granuloma volume, HMI and ALT levels and fibrosis, which was demonstrated by increased hepatic collagen deposition in the infected mice. Injection of rmIL-33 into the infected IL-33−/− mice strongly abrogated the liver pathology and fibrosis, whereas no detectable effect with injecting rmIL-33 into the infected ST2−/− mice. Furthermore, depletion of the IL-33/ST2 axis inhibited Treg, accompanied by increased Th17. rmIL-33 treatment upregulated Treg and downregulated Th17 in the infected IL-33−/− mice, while no effect in the infected ST2−/− mice. rmIL-33 led to elevated expressions of Atg5, Beclin-1 and inhibited expression of p62 in expansion of Treg.
Conclusion: The IL-33/ST2 axis plays a protective role in S. japonicum infected mice, which is closely related to increasing Treg responses as well as suppressing Th17 responses. Expansion of Treg by IL-33 may be associated with its regulation of autophagy.
Keywords: IL-33, ST2, liver pathology, fibrosis, Treg, Th17, autophagy
Introduction
Schistosomiasis is one of the most devastating tropical diseases, caused by blood flukes (trematode worms) of the genus Schistosoma. According to the WHO report 2020, at least 290.8 million people required preventive treatment for schistosomiasis, out of which more than 97.2 million people were reported to have been treated in 2018.1,2 Schistosomiasis is characteristic of egg granulomas and fibrosis due to egg encapsulation in the liver. Chronic hepatic egg granulomas can result in severe significant complications including liver cirrhosis, portal hypertension and even liver failure.3,4 Host granulomatous response to Schistosoma eggs can be both protective and pathological, dependent on degrees of the corresponding immune responses.5 It is estimated that only 5–10% of individuals with schistosomiasis develop severe forms of the disease in endemic areas.6 It is therefore suggested that Schistosoma-infected hosts have developed a variety of strategies in their immunomodulatory armamentarium to initiate immune responses to clear the pathogen as well as to reduce the damage caused by excessive immune responses.7 However, cellular mechanism regarding immune modulation in schistosomiasis is mostly unknown.
The occurrence and progression of immune responses related with granuloma formation and fibrosis in schistosomiasis are primarily mediated by T helper cells.8 Th17 and regulatory T cells (Treg) are two of the main Th phenotypes involved in schistosomiasis. Th17 is closely related to the occurrence and development of egg granuloma and fibrosis, and suppression of excessive Th17 immune responses can reduce the liver immunopathological damage.3,9 Evidence is emerging that Treg plays an essential role in regulation of immune response and immune pathological reaction in schistosomiasis, thus determines the magnitude and phenotype of Th effector population and intensity of the immune response as well as the outcome of the disease.8,10–13 In some conditions, Treg and Th17 can also trans-differentiate from each other. Activation of Th cells is known to be regulated by signal proteins, cytokines, transcription factors and gene elements, and it is still elusive how Schistosoma infection induces the activation of Treg in hosts.3
The pathology of schistosomiasis results from host immune responses against the pathogen.1,3 It has been indicated that many inflammatory mediators are associated with immune alterations.3,14,15 Recent studies have found that interleukin-33 (IL-33) participates in various inflammatory diseases.16,17 IL-33 is a member of the IL-1 family and generally resides as a nuclear factor in the nucleus of the cells, whereas it can be quickly released into extracellular space upon tissue injury or necrosis. Thus, it is regarded as a typical alarmin due to its indication of cellular damage.16 To exert its biological functions, IL-33 binds to the specific receptor known as ST2 (suppress of tumorigenicity 2), which is called as IL-33/ST2 axis.18 The membrane-bound ST2 functions as the only known receptor for IL-33. The ST2 can be expressed by various immune cells participating in innate immunity and adaptive immunity, including innate lymphoid cell type 2 (ILC2), dendritic cells (DCs), mast cells, macrophages, Th2 cells, and Tregs.16,18 The IL-33/ST2 axis plays diverse but crucial roles in the homeostasis of immune responses and the pathogenesis of immune-mediated diseases.18,19 The exacerbated or protective role of the IL-33/ST2 axis during different diseases is dependent on diseases models, involved organs, stages of disease, host immune compartments, and cellular and cytokine microenvironments.
Both S. japonicum and S. mansoni infection resulted in upregulated IL-33 in experimental mice.20–22 Moreover, serum IL-33 levels are increased in patients with schistosomiasis japonica, mansoni and haematobium.21,23,24 Although IL-33 promotes type 2 immune responses in infected mice with S. japonicum while not in infected mice with S. mansoni, it has been associated with liver pathology in both experimental models.20–22 The previously reported kinetics of IL-33 levels and the proportions of Treg and Th17 peak at the 8th week post infection are consistent with the period of immunopathological damage of schistosomiasis japonica.20,25 However, a link between IL-33, an endogenous mediator of tissue damage and its immunomodulatory armamentarium of Th cells, especially Treg and Th17, has not been further elucidated.
Based on the literature research summarized above, in this study, we explored the effects of the IL-33/ST2 axis in liver egg granuloma and fibrosis and the potential mechanism focusing on its regulation of Treg and Th17 responses in murine schistosomiasis japonica.
Materials and Methods
Ethics Statement
All animal experiments were performed in strict accordance with the Institutional Animal Care and Use Committee at Tongji Medical College, Huazhong University of Science and Technology (SCXK2016-0011). All infection and sacrifice were performed under anesthesia.
Parasites and Animals
IL-33 knockout (IL-33−/−) and ST2 knockout (ST2−/−) C57BL/6 mice were kind gifts from Department of Immunology, Tongji Medical College.26,27 Wild-type C57BL/6 mice (WT) were obtained from Hubei Province Center for Disease Control and Prevention (Wuhan, China). All mice were maintained in a standard specific pathogen-free research animal facility for further use. S. japonicum-infected Oncomelania hupensis snails were provided by Jiangsu Province Institute of Parasitosis Control and Prevention (Wuxi, China).
Animal Challenge Infection and Treatment with Recombinant IL-33
Eight-week-old female WT, IL-33−/− and ST2−/− mice were divided into uninfected and infected groups. For the infected groups, mice were anesthetized with 0.67% pentobarbital sodium solution (10 μL per 1 g of body weight) and infected percutaneously with 20 S. japonicum cercariae per mouse through the shaved abdominal skin. Half of infected mice were treated with intraperitoneal injection of 0.5 µg of recombinant mouse IL-33 (rmIL-33, Biolegend, San Diego, CA) in phosphate-buffered saline (PBS) per mouse, twice a week, from the 4th week to their sacrifice at the 8th week post infection. Non-treated infected mice had intraperitoneal injections of the same volume of PBS and were used as the infected controls.
Assessment of Activities, Appetite, Body Weight and Hepatic Mass Index
Activities and appetite of all mice were monitored during the experiment. All experimental animals were sacrificed at the 8th week post infection. The liver and body weight of each mouse were weighed and the proportion of the liver weight to its body weight was calculated and recorded as hepatic mass index (HMI).
Determination of Hepatic Number and Size of Egg Granulomas
Liver sections were embedded in paraffin and cut into 5 μm section and stained with haematoxylin and eosin (H&E). The number and size of liver egg granulomas were calculated according to our previous publication.28
Briefly, egg granulomas in 0.5 cm2 of each of the two sections from each liver were counted. The diameter of single egg granuloma per liver was measured for 10 granulomas per liver (five granulomas from each section, two sections per liver) with a video micrometre in accordance with the manufacturer’s instructions (Olympus, Tokyo, Japan). Means of two perpendicular diameters (d) were evaluated in micrometre (μm). Size of each granuloma was assessed according to the following formula: area = πd2/4.
Masson Trichrome Staining
To observe collagen fibre deposition, liver sections were dehydrated in a graded sucrose series and followed by Masson trichrome staining performed with the standard methods (BASO, Zhuhai, China). The optical density of collagen fibres was quantified and analysed using the Image-Pro Plus 6.0 software (Media Cybernetics Inc, Rockville, MD, USA).
Immunohistochemical Assay
Immunohistochemical staining for collagen 1a1 (Col 1a1) and α-smooth muscle actin (α-SMA) were performed according to our previous publication.29 Liver sections were incubated with rabbit anti-mouse Col 1a1 (dilution 1:200, ab34710; Abcam, Cambridge, MA, USA) or rabbit anti-mouse α-SMA (1:500 dilution, ab124964; Abcam, Cambridge, MA, USA) at 4°C overnight. Horseradish peroxidase (HRP)-conjugated anti-rabbit IgG antibody was used as the secondary antibody (1:1000 dilution, Cell Signaling Technology). Diaminobenzidine solution (DAB) was applied as the chromogen. Optical density (OD) of the target protein was measured with the Image-Pro Plus 6.0 software. The OD value, which represents the quantity of the targeted protein, was expressed as ratio of positive optical density (yellow) and total pixels.
Flow Cytometric Analysis
Flow cytometry analyses were performed to assess the proportion of Th17 and Treg cells in livers and spleens. Hepatic and Spleen lymphocytes were prepared as previously described.30 Lymphocytes were purified using lympholyte M (Cedarlane, Ontario, Canada) and washed with PBS twice before staining. Next, surface staining of anti-CD4-FITC and anti-ST2-APC were performed at 4°C for 30 minutes. Followed by fixation and permeabilization, anti-Foxp3-PE or anti-IL-17A-PE fluorescent antibodies were added and incubated at 4°C for 30 minutes. Cell sorting was performed using a FACS cytometer (BD Biosciences). All antibodies were purchased from eBioscience, San Diego, CA. Gating strategies for identifying Treg and Th17 subsets from lymphocyte populations were shown in Supplementary Figures 1 and 2, respectively. CD4 + IL-17 + cells were recognized as the Th17 cells and CD4 + Foxp3 + cells as Treg cells. Data were analysed with FlowJo (Tree Star, version 10.0.7).
Polarization and Stimulation of CD4+ T Cells
Liver naïve CD4+ T cells (CD4+ CD25− CD44− CD62L+) were sorted from the infected WT mice as previously described.31 Naïve CD4+ T cells were then cultured (3×105 cells/well) in 96-well plates coated with anti-CD3 mAb (5 μg/mL, eBioscience) and soluble anti-CD28 mAb (3 μg/mL, eBioscience) and kept in the presence of IL-2 (100 U/mL, eBioscience). Exogenous rmIL-33 (10 ng/mL) was added to investigate its ability to polarize Treg and PBS used as the control. Cells were cultured in RPMI-1640 Medium, 10% fetal calf serum, 2 mM L-glutamine, 100 U/mL of Penicillin/Streptomycin for 5 days.
Quantitative Real-Time PCR
Total RNA was extracted with a TRIzol® Reagent kit (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instruction. For tissues, 1 mL of Reagent per 100 mg of tissue was added to the sample and homogenized using a homogenizer. For cells, 1 mL of Reagent per 1×106 cells was added directly to the culture dish to lyse the cells, and then pipetted the lysate up and down several times to homogenize. The quantity and purity of the extracted RNA were assessed using a spectrophotometer Nanodrop‐2000 (Thermo Scientific, USA). RNA (1 μg) with OD 260/280 ratio in the range of 1.8–2.0 was reverse-transcribed into complementary DNA (cDNA) by High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, Thermo Fisher Scientific). Then, real-time PCR was performed using Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) with gene-specific primers as listed in Table 1. The amplification reactions were carried out with an initial hold step (95°C for 5 minutes), followed by 40 cycles of a three-step PCR (95°C for 30 seconds, 60°C for 20 seconds, and 72°C for 20 seconds), and a final extension at 72°C for 5 minutes. Step One Plus real-time PCR System was used to detect the fluorescence signal intensity (Ct value) of cyclic amplification of target and reference genes. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an endogenous normalization control, and relative quantity values of each sample were calculated with the 2−ΔΔCt method.
 |
Table 1 Primer Sequences of Target mRNA |
Determination of Serum Levels of Alanine Aminotransferase (ALT)
Serum levels of ALT were detected by kits according to the manufacturer’s instruction (Jiancheng Bioengineering Institute of Nanjing, Nanjing, China).
Sample Collection for Cytokine Analysis and ELISA
Serum was collected from blood and prepared for ELISA analysis according to the previous publication.32 Cytokine concentrations in mouse serum or splenocyte supernatant were measured with ELISA kits (R&D Systems) according to the manufacturer’s instructions.
Statistical Analysis
All experimental data were expressed as means ± SEMs and analysed with SPSS v19.0 (SPSS, Chicago, IL, USA). All in vivo studies were repeated two independent times and the in vitro experiments were repeated three times. Comparisons of two groups were analyzed by Student’s unpaired t-test. Comparison among multiple groups was performed by multivariate analysis of variance (ANOVA). A P value of <0.05 was considered statistically significant (*P < 0.05, **P < 0.01, ***P < 0.001). Graphs were generated using the software GraphPad Prism v.7.0.
Results
S. japonicum Infection Results in Elevated Expression of IL-33 and ST2 in Mice
In comparison to the uninfected controls, the infected mice had much higher concentration of serum IL-33 at the 8th week post infection (Figure 1A). mRNA levels of IL-33 and ST2 in the liver from the infected mice also increased significantly relative to the uninfected controls (Figure 1B and C). Together, S. japonicum infection results in elevated expression of IL-33 and ST2 at the peak of pathological damage in mice, suggesting that the IL-33/ST2 axis might play an important role in the pathogenesis of schistosomiasis.
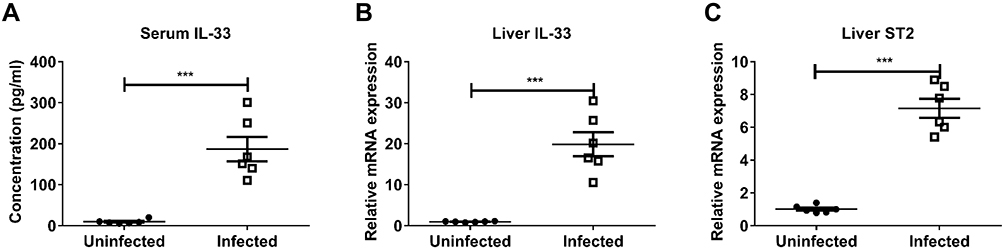 |
Figure 1 Schistosoma japonicum infection upregulated the expression of IL-33/ST2 axis in mice. WT mice were divided into the infected and uninfected group. Each mouse in the infected group was infected with 20 cercariae through shaved abdominal skin. At the 8th week post infection, all mice were sacrificed and the peripheral blood and liver were collected. (A) The protein expression level of IL-33 in peripheral blood was measured by ELISA. The mRNA expression levels of IL-33 (B) and ST2 (C) in liver of mice was measured by real-time PCR. Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences between two groups (***P < 0.001). |
IL-33 Deficiency Aggravates Liver Pathological Lesions in Murine Schistosomiasis Japonica
At the 8th week post S. japonicum infection, reduced activities and appetite were observed in all infected groups with weight loss and listlessness. To determine the effects of IL-33 on liver pathology associated with the infection, we compared parameters of hepatic pathological lesions and fibrosis between the infected WT and the infected IL-33−/− mice. The results of the histopathological examinations of liver tissues using H&E staining are shown in Figure 2A. Livers of the both infected groups had interstitial oedema and vacuolization of hepatocytes with more severity shown in the infected IL-33−/− group. Moreover, IL-33 deficiency led to increased size of liver egg granulomas in the infected mice (Figure 2A and B). As shown in Figure 2C and D, hepatic fibrosis was aggravated in the infected IL-33−/− mice, evidenced by increased blue fibre spots encircling egg granuloma in Masson staining, accompanied by elevated deposits of α-SMA (Figure 2E and F) and Col 1a1 (Figure 2G and H). In addition, the infected IL-33−/− mice had increased HMI and a higher level of serum ALT relative to the infected WT mice, respectively, indicating a more severely damaged liver function (Figure 2I and J). Collectively, these results demonstrated that IL-33 deficiency could exacerbate parasitological lesions and liver fibrosis in the infected mice with S. japonicum.
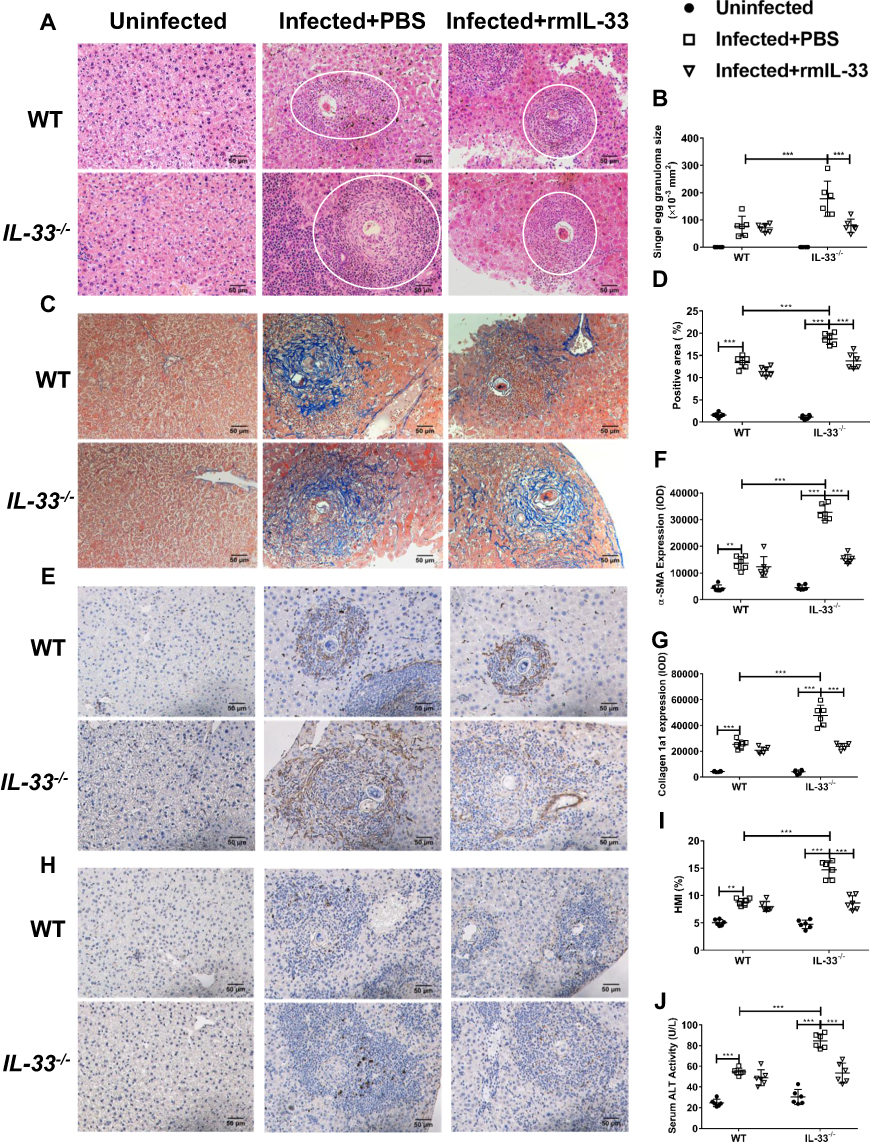 |
Figure 2 IL-33 deficiency aggravated liver pathological lesions in murine schistosomiasis japonica. WT and IL-33−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. The liver weight and body weight of all mice were recorded. At the 8th week post infection, all mice were sacrificed and the liver and peripheral blood were collected. (A) Representative images of HE staining of liver sections (single egg granuloma indicated by white circles) and (B) the statistical graphs of single granuloma area. (C) Representative images of Masson’s staining of liver sections and (D) the statistical graphs of the blue collagen area proportion. The hepatic expression of two main extracellular matrix components, α-SMA (E) and Collagen 1a1 (H). The statistics of IOD of α-SMA (F) and Collagen 1a1 (G). The statistical graphs of HMI (Hepatic Mass Index, (I)) and ALT (Alanine aminotransferase, (J)) levels in peripheral blood from mice. Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (**P < 0.01, ***P < 0.001). |
To further verify the role of IL-33 during S. japonicum infection, exogenous rmIL-33 was injected into the infected IL-33−/− mice from 4 weeks (the beginning of egg deposition) to 8 weeks post infection. Interestingly, administration of rmIL-33 significantly rescued both the liver pathology and fibrosis in the infected mice caused by IL-33 deficiency (Figure 2A–J). Altogether, these results indicated that IL-33 has a protective effect on the hepatic pathology in murine schistosomiasis japonica, given the striking protection promoted by exogenous rmIL-33 treatment even in gene deficient with IL-33.
ST2 is Indispensable for the Protective Effect of IL-33 on Liver Pathology in Murine Schistosomiasis Japonica
It has been reported that IL-33 exerts its biological effects through binding to its receptor ST2. To verify the role of ST2 among the protective impact of IL-33 in the infected mice, we investigated the liver pathology and fibrosis in the infected ST2−/− mice. As Figure 3A and B shows, ST2 deficiency led to increased size of liver egg granulomas in the infected mice. In addition, hepatic fibrosis was also aggravated in the infected ST2−/− mice, evidenced by increased blue fibre spots encircling egg granuloma in Masson staining, accompanied by elevated deposits of α-SMA (Figure 3E and F) and Col 1a1 (Figure 3G and H). Moreover, the infected ST2−/− mice had increased HMI and a higher level of serum ALT relative to the infected WT control, indicating a more severely damaged liver function (Figure 3I and J). Collectively, these results demonstrated that ST2 deficiency resulted in deteriorated parasitological lesions and liver fibrosis in the infected mice. Furthermore, administration of rmIL-33 had no effect on the liver pathology and fibrosis in the infected mice caused by ST2 deficiency (Figure 3A–J). Taken together, ST2 is indispensable for the protective effect of IL-33 on liver pathology in murine schistosomiasis japonica.
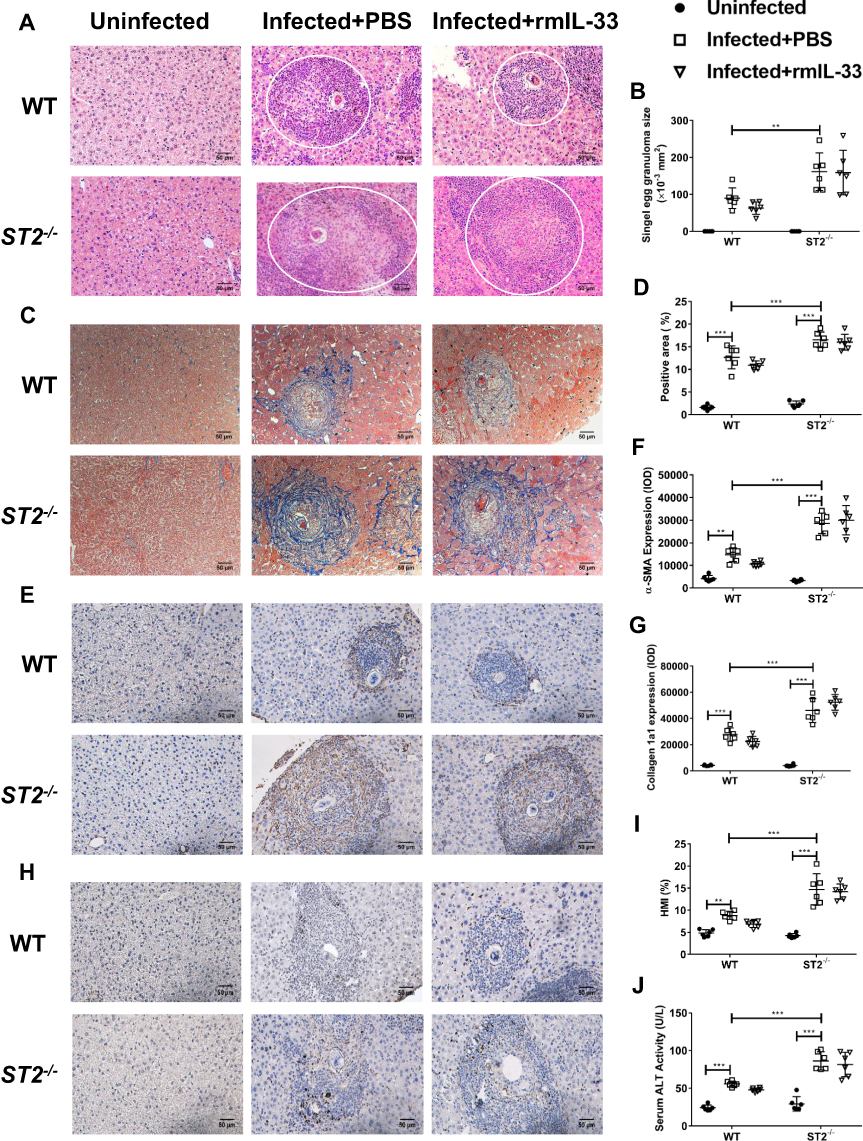 |
Figure 3 ST2 is indispensable for the protective effect of IL-33 on liver pathology in murine schistosomiasis japonica. WT and ST2−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to the 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. The liver weight and body weight of all mice were recorded. At the 8th week post infection, all mice were sacrificed and the liver and peripheral blood were collected. (A) Representative images of HE staining of liver sections (single egg granuloma indicated by white circles) and (B) the statistical graphs of single granuloma area. (C) Representative images of Masson’s staining of liver sections and (D) the statistical graphs of the blue collagen area proportion. The hepatic expression of two main extracellular matrix components, α-SMA (E) and Collagen 1a1 (H). The statistics of IOD of α-SMA (F) and Collagen 1a1 (G). The statistical graphs of HMI (Hepatic Mass Index, (I)) and ALT (Alanine aminotransferase, (J)) levels in peripheral blood from mice. Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (**P < 0.01, ***P < 0.001). |
IL-33 Deficiency Leads to Decreased Proportion and Function of Treg in Murine Schistosomiasis Japonica
Previous studies show that IL-33 expands Treg population and enhances Treg function in some immune-mediated diseases.33–35 To confirm whether IL-33 could amplify Treg population in murine schistosomiasis japonica, we compared the frequencies of Treg between different groups. Consistent with the previous studies, S. japonicum infection led to increased percentages of Treg cells in all infected mice (Figure 4A–D), accompanied by enhanced mRNA expression of liver Foxp3 (Figure 4I). Moreover, percentages of ST2+Treg cells in liver and spleen were upregulated in the infected mice (Figure 4E–H), along with increased mRNA expression of liver ST2 (Figure 4J). However, IL-33 deficiency resulted in the diminished percentages of Treg and ST2+Treg cells in the spleen and liver from the infected mice, along with decreased mRNA expressions of liver Foxp3 and ST2 (Figure 4A–J). Notably, administration of rmIL-33 reversed all the above parameters to varying degrees in the infected IL-33−/− mice (Figure 4A–J).
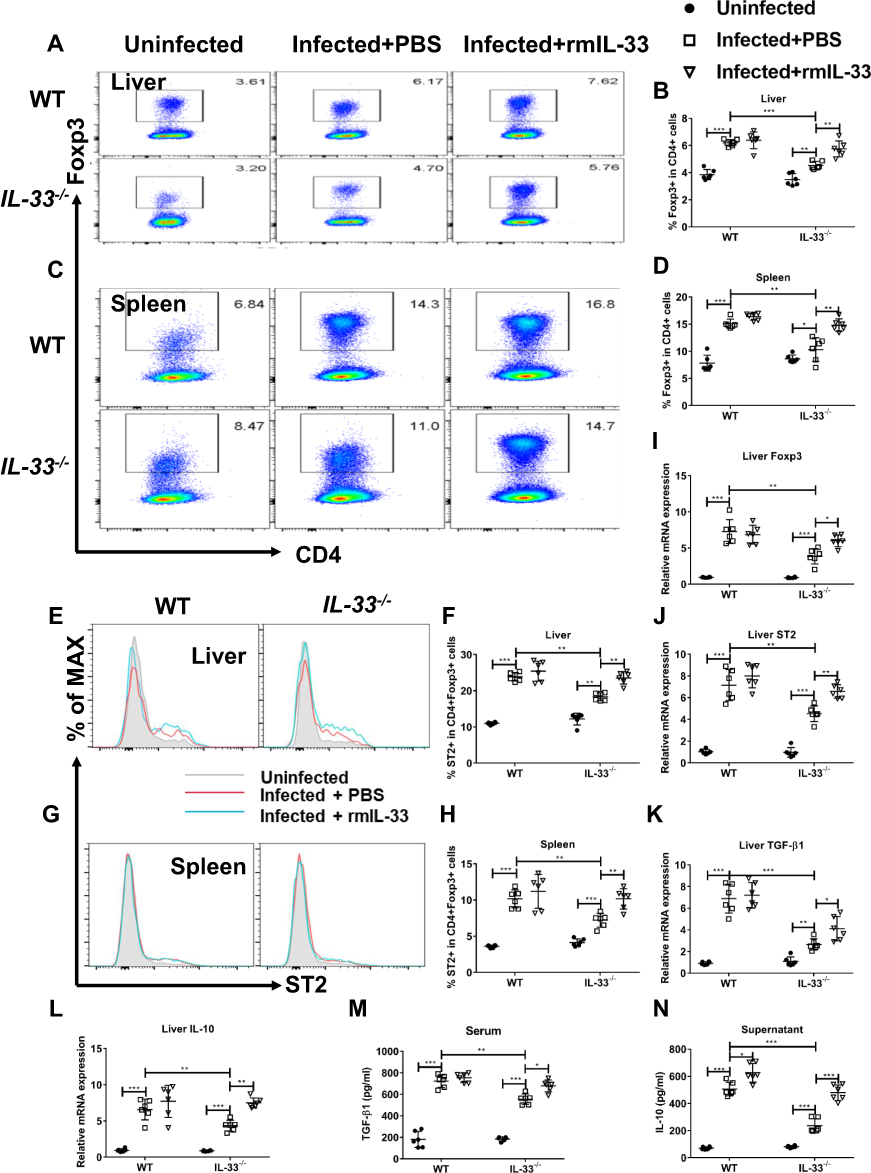 |
Figure 4 IL-33 deficiency leads to decreased proportion and function of Treg in murine schistosomiasis japonica. WT and IL-33−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. At the 8th week post infection, all mice were sacrificed. The liver, spleen and peripheral blood were collected. Splenic lymphocytes were stimulated with soluble egg antigen (SEA, 10 μg/mL) in vitro for 72 h and then the supernatant was collected. (A and B) The proportion of Foxp3+Treg in CD4+ T cells in liver. (C and D) The proportion of Foxp3+Treg in CD4+ T cells in spleen. The proportion of ST2+Treg in total Treg in liver (E and F) and in spleen (G and H). The mRNA expression levels of Foxp3 (I), ST2 (J), TGF-β1 (K) and IL-10 (L) in liver. The concentrations of TGF-β1 in serum (M) and IL-10 in splenic supernatants (N). Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (*P < 0.05, **P < 0.01, ***P < 0.001). |
To verify the effects of IL-33 on Treg function in the infected mice, we next detected expression levels of the functional cytokines TGF-β1 and IL-10. The results showed that S. japonicum infection upregulated mRNA expression levels of hepatic TGF-β1 and IL-10, accompanied with elevated concentrations of TGF-β1 in serum and IL-10 in splenocyte supernatant in mice (Figure 4K–N). However, IL-33 deficiency attenuated all these aforementioned expression levels in the infected mice, while rmIL-33 treatment could significantly invert the above parameters of Treg in the infected IL-33−/− mice (Figure 4K–N). In Figure 4N, compared with that of the infected plus PBS WT, the higher splenic IL-10 concentration in the infected plus rmIL-33 WT might be associated with the effect of the exogenous rmIL-33 injection, although the amount of exogenous IL-33 is much lower than that of endogenous IL-33. Collectively, our results show that IL-33 expands both Treg and ST2+Treg populations and enhances Treg function in murine schistosomiasis japonica.
ST2 is Indispensable for Activation of Treg by IL-33 in Murine Schistosomiasis Japonica
To confirm whether IL-33 could amplify the Treg population and function through binding to ST2 in the infected mice, we compared the Treg percentages and functions between the infected WT and ST2−/− mice. The results showed that ST2 deficiency resulted in the diminished percentage of Treg and ST2+Treg cells in the spleen and liver from the infected mice (Figure 5A–H), along with decreased mRNA expression of liver Foxp3 and ST2 (Figure 5I and J). Furthermore, compared with the infected WT mice, both mRNA expression and concentrations of TGF-β and IL-10 were decreased in the infected ST2−/− mice (Figure 4K–N). Administration of rmIL-33 had no effect on all the above parameters in the infected mice without ST2 (Figure 5A–N). Taken together, ST2 is indispensable for activation of Treg by IL-33 in murine schistosomiasis japonica.
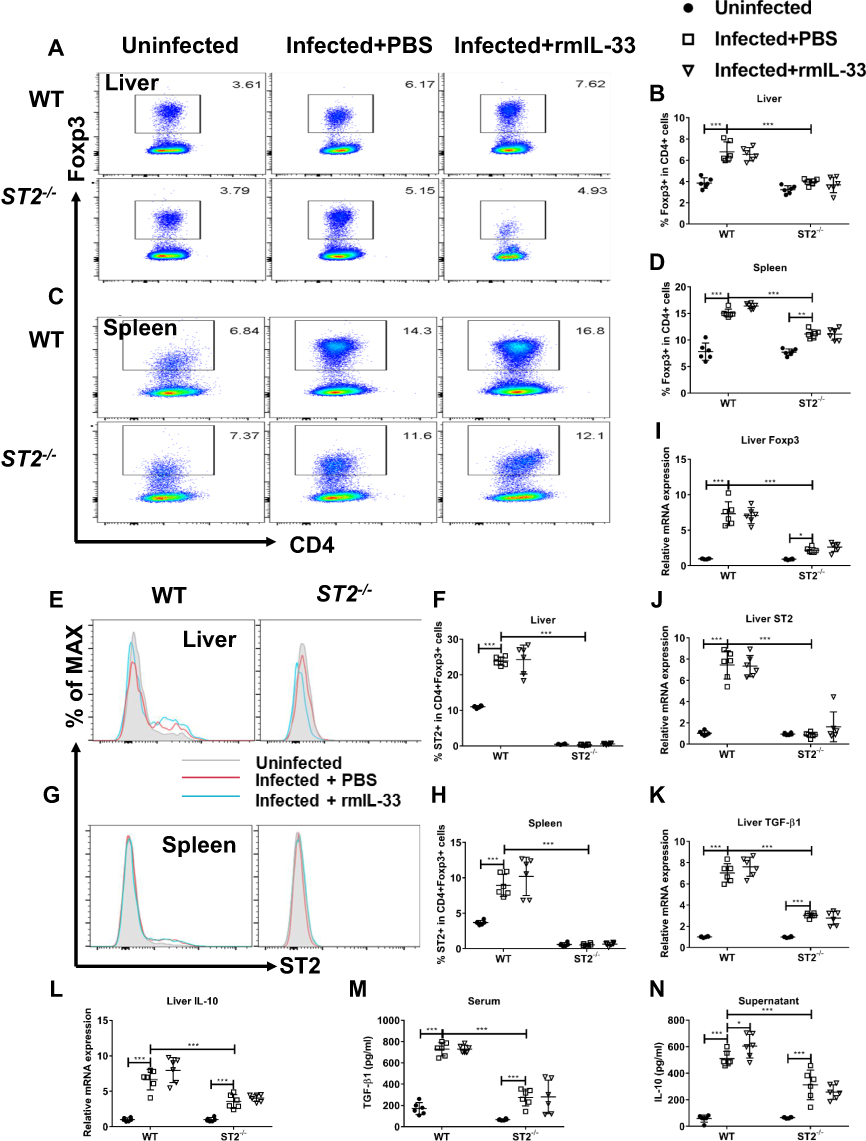 |
Figure 5 ST2 is indispensable for activation of Treg by IL-33 in murine schistosomiasis japonica. WT and ST2−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. At the 8th week post infection, all mice were sacrificed. The liver, spleen and peripheral blood were collected. Splenic lymphocytes were stimulated with soluble egg antigen (SEA, 10 μg/mL) in vitro for 72 h and then the supernatant was collected. (A and B) The proportion of Foxp3+Treg in CD4+ T cells in liver. (C and D) The proportion of Foxp3+Treg in CD4+ T cells in spleen. The proportion of ST2+Treg in total Treg in liver (E and F) and in spleen (G and H). The mRNA expression levels of Foxp3 (I), ST2 (J), TGF-β1 (K) and IL-10 (L) in liver. The concentrations of TGF-β1 in serum (M) and IL-10 in splenic supernatants (N). Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (*P < 0.05, ***P < 0.001). |
IL-33 Deficiency Leads to Increased Proportion and Function of Th17 in Murine Schistosomiasis Japonica
IL-33 is reported to affect Th17 population and function in some immune-mediated diseases, although its effects on Th17 vary in different disease models.37,38 To explore role of IL-33 on Th17 immune responses in murine schistosomiasis japonica, we compared the frequencies of Th17 between different groups. Consistent with the previous studies, S. japonicum infection led to increased percentages of Th17 in all infected mice (Figure 6A–D), accompanied with enhanced mRNA expression of liver RORγt (Figure 6E). Compared with the infected WT control, the infected IL-33−/− mice had higher percentage of Th17 and increased mRNA expression of liver RORγt. Administration of rmIL-33 inhibited all the above parameters to varying degrees in the infected IL-33−/− mice (Figure 6A–E).
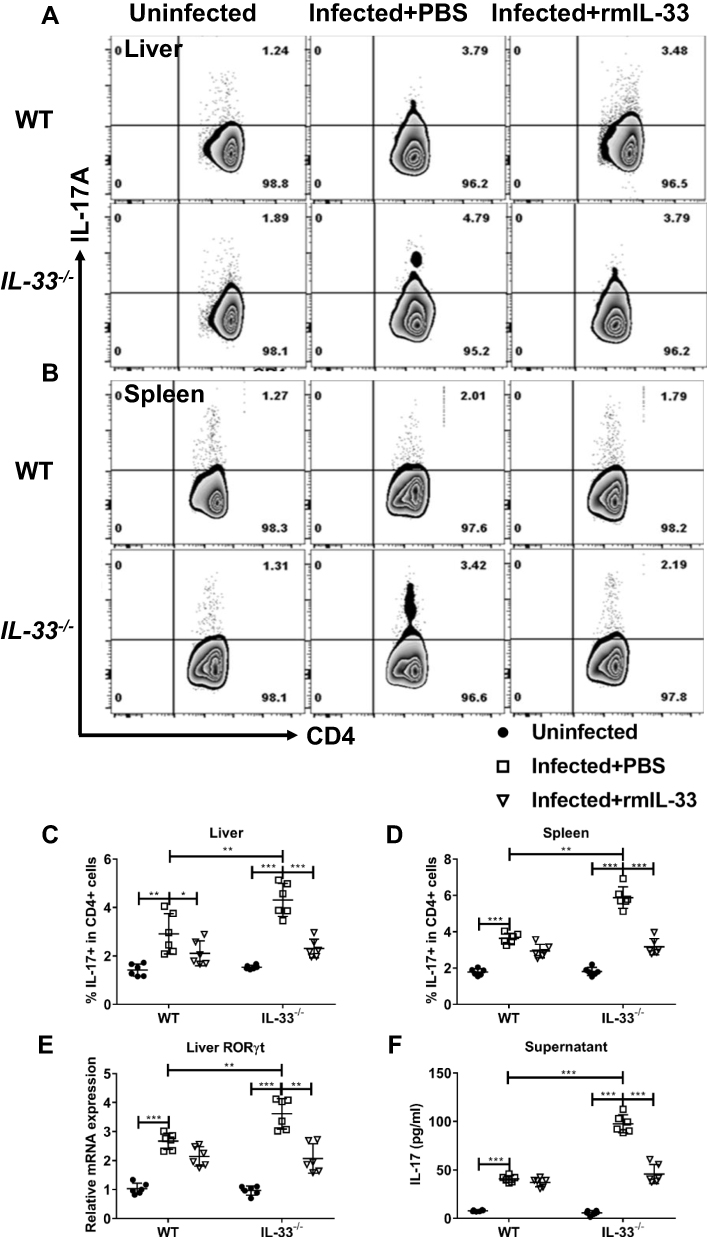 |
Figure 6 IL-33 deficiency leads to increased proportion and function of Th17 in murine schistosomiasis japonica. WT and IL-33−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. At the 8th week post infection, all mice were sacrificed. The liver, spleen and peripheral blood were collected. The proportion of IL-17A+T cells in CD4+ T cells in liver (A and C) and spleen (B and D) of mice. (E) The mRNA expression level of Th17 specific transcription factor RORγt in liver and (F) The concentration of IL-17 in splenic supernatant. Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (*P < 0.05, **P < 0.01, ***P < 0.001). |
To verify the effect of IL-33 on Th17 function in the infected mice, next we detected IL-17 concentration in splenocyte supernatant from all groups. Figure 6F shows that S. japonicum infection upregulated concentration of IL-17 in all infected mice. However, IL-33 deficiency augmented IL-17 concentration post infection. Administration of rmIL-33 restored elevated IL-17 levels caused by IL-33 deficiency in the infected WT mice (Figure 6F). Collectively, our results show that IL-33 inhibits Th17 population and function in murine schistosomiasis japonica.
ST2 is Indispensable for IL-33 Inhibition of Th17 in Murine Schistosomiasis Japonica
To verify whether IL-33 could depress Th17 cell population and function through binding to ST2 in the infected mice, we compared the Th17 percentages and functions between the infected WT and ST2−/− mice. The results showed that ST2 deficiency diminished percentage of Th17 cells in the spleen and liver from the infected mice (Figure 7A–D), along with decreased mRNA expression of liver RORγt (Figure 7E). Furthermore, compared with the infected WT mice, IL-17 concentration in splenocyte supernatant attenuated in the infected ST2−/− mice (Figure 7F). Administration of rmIL-33 had no effect on all the above parameters in the infected mice without ST2 (Figure 7A–F). As such, ST2 is indispensable for IL-33 inhibition of Th17 in murine schistosomiasis japonica.
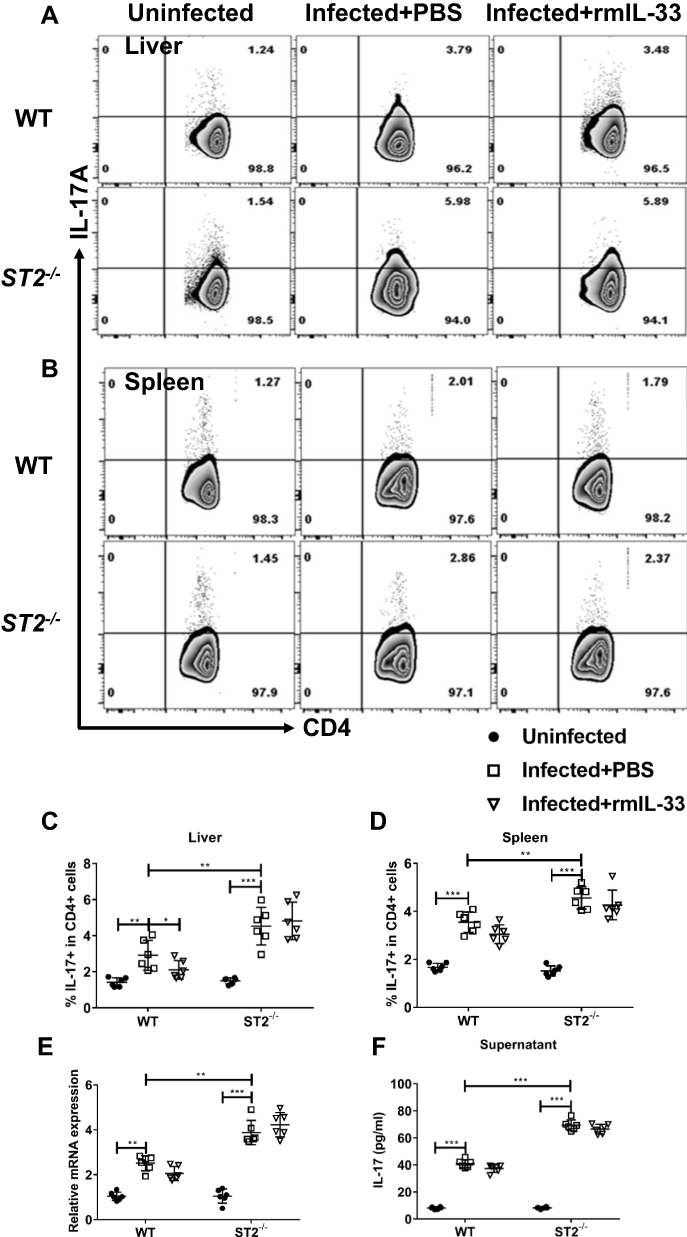 |
Figure 7 ST2 is indispensable for IL-33 inhibition of Th17 in murine schistosomiasis japonica. WT and ST2−/− mice were divided into uninfected group, infected plus rmIL-33 group and infected plus PBS group. Each mouse in the infected groups was infected with 20 cercariae through shaved abdominal skin. Mice in the infected plus rmIL-33 group were intraperitoneally injected with exogenous rmIL-33 (dissolved in sterile PBS solution) from the 4th week to 8th week post infection, with the total 5 μg of rmIL-33 per mouse. The mice in the infection plus PBS group were simultaneously given the equal volume of PBS. At the 8th week post infection, all mice were sacrificed. The liver, spleen and peripheral blood were collected. The proportion of IL-17A+T cells in CD4+ T cells in liver (A and C) and spleen (B and D) of mice. (E) The mRNA expression level of Th17 specific transcription factor RORγt in liver and (F) The concentration of IL-17 in splenic supernatant. Data are expressed as means ± SEMs based on 6 mice in each group and from 2 independent experiments. Asterisks mark significant differences among different groups (*P < 0.05, **P < 0.01, ***P < 0.001). |
Expansion of Treg by IL-33 via Modifying Autophagy in vitro
It has demonstrated that IL-33 promotes autophagy in several cell types, although the target cells and underlying mechanism vary in different immune disorders.39–41 Given autophagy plays an important role in CD4+ cells differentiation and is indispensable for Treg cell maintenance and function in peripheral tissues,31,42,43 we hypothesized that IL-33 might expand Treg through upregulating autophagy in the infected mice. To test our hypothesis, we sorted naïve CD4+ T cells from the infected WT mice and then co-cultured them with rmIL-33 in vitro. As shown in Figure 8, IL-33 could promote Treg polarization, evidenced by increased mRNA expressions of Foxp3, TGF-β1 and IL-10, as compared with the PBS control. IL-33 was associated with elevated mRNA levels of Atg5, Beclin-1 and inhibited mRNA levels of p62 in Treg. Both Atg5 and Beclin-1 are indicators of autophagy.42 p62, indicator of autophagy flux, can be reduced intracellularly when autophagy is activated and vice versa.44 These results provide evidence that IL-33 promotes naïve CD4+ T cells to differentiate into Treg through upregulating autophagy in vitro.
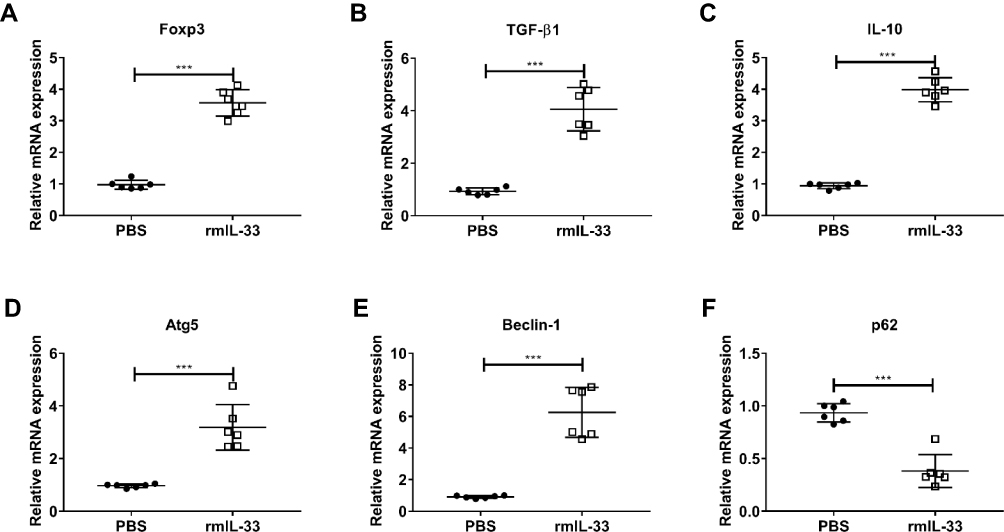 |
Figure 8 Expansion of Treg by IL-33 via modifying autophagy in vitro. Liver naïve CD4+ T cells (CD4+ CD25− CD44− CD62L+) were sorted from the infected WT mice at the 8th week post infection. T cells were cultured (3×105 cells/well) in 96-well plates coated with anti-CD3 mAb (5 μg/mL, eBioscience) and soluble anti-CD28 mAb (3 μg/mL, eBioscience) and kept in presence of IL-2 (100 U/mL, eBioscience). Exogenous rmIL-33 (10 ng/mL) was added to investigate its ability to polarize Treg and PBS used as the control. Cells were cultured in RPMI-1640 Medium, 10% fetal calf serum, 2 mM L-glutamine, 100 U/mL of Penicillin/Streptomycin for 5 days. The mRNA expression level of Treg-related parameters, Foxp3 (A) and TGF-β1 (B) and IL-10 (C) in cultured cells. The mRNA expression level of autophagy-related parameters, Atg5 (D) and Beclin-1 (E) and p62 (F) in cultured cells. Data are expressed as means ± SEMs based on 6 samples in each group and from 3 independent experiments. Asterisks mark significant differences among different groups (***P < 0.001). |
Discussion
Schistosomiasis is the most important helminth disease in the world that causes millions of morbidities and mortalities in endemic areas. A plethora of cytokines produced by diverse cell types, including local tissue and immune cells, are involved in Schistosoma-induced liver immunopathology.3,8 The alarmin IL-33 has been reported to function as a pleiotropic cytokine involved in immune regulation, host defense, tissue repair and metabolic homeostasis.45 However, the role of IL-33 remains controversial in different schistosomiasis settings.20–22 Therefore, more comprehensive exploration and deeper understanding of the IL-33/ST2 pathway especially in the peak of immunopathology during Schistosoma infection are required. In this work, we compiled evidence that the absence of the IL-33/ST2 axis aggravates the liver immunopathology and fibrosis at the 8th week post S. japonicum infection and the lack of this pathway decreases the percentage and function of Treg, along with upregulating the percentage and function of Th17. The modulation of IL-33 in Treg is associated with its regulation of autophagy.
IL-33 is mainly expressed by epithelial cells, endothelial cells, and fibroblasts of barrier tissues, indicating a central role for this cytokine in barrier tissue defense.33,46 Emerging studies have identified that expression of IL-33 can be induced by parasite infections in humans and mice.47–49 Our previous work shows that Schistosoma infection results in increased levels of serum IL-33 in mice due to epithelial cells and endothelial cells damaged caused by the parasite migration, with a peak plateau from 7 to 9 weeks post infection.20 As observed in humans, the serum IL-33 level is significantly elevated in acute patients of schistosomiasis japonica (7–9 weeks post infection).50 These are the experimental basis for choosing 8 weeks post infection as our assessment time point in this experiment. We found that the S. japonicum infected mice not only had higher serum levels of IL-33, but also had elevated hepatic mRNA expression of IL-33 and ST2, compared to the uninfected control. It indicated that there was activation of the IL-33/ST2 axis in schistosome infection, which was consistent with the previous findings.51
The IL-33/ST2 axis may play pleiotropic roles during parasite infections, depending on parasite species, organs involved and related immune mechanism underlying the pathogenesis of disease conditions.52 The current work showed that S. japonicum infection resulted in much severe hepatic pathology in the gene depletion mice with either IL-33 or ST2 relative to the WT mice, although there was no difference in egg granuloma number among these groups (data not shown). Notably, administration of exogenous rmIL-33 could reverse the exaggerated trend of the liver immunopathology in the infected IL-33−/− mice whereas had no effect on the liver pathology in the infected ST2−/− mice. These results indicate that the IL-33/ST2 axis plays a crucial protective role in schistosomiasis japonica via suppressing granuloma formation and associated pathology. The IL-33/ST2 axis is actively involved in various infections in humans and animals, among them IL-33 is shown to be a Janus cytokine in the development of infectious disease mechanisms.35 IL-33 can be protective during infections with Trichuris muris,47 hookworm,53 and Toxoplasma gondii,54 whereas it acts as a new deleterious regulator during visceral leishmaniasis49 and Helicobacter hepaticus infection.55 The IL-33/ST2 axis is necessary to prevent host mortality by modulating granuloma-mediated pathology in schistosomiasis mansoni.56 In experimental schistosomiasis, the IL-33/ST2 axis has been associated with increased Th2 response in infections with S. japonicum,20 but not with S. mansoni.56 IL-33 has been reported as one of keystones in liver inflammatory diseases, with protective or pro-inflammatory effects, depending on assessment timing and disease models.57 Reports from different groups have suggested that variable functions of IL-33 in Schistosoma infected mice, which might duo to different species involved, different infection stages and assessment time points.21,22,56 Given the diverse function of IL-33 during inflammation disorders, further investigation of detailed molecular mechanisms by which the IL-33/ST2 axis protects hepatic pathology associated with schistosomiasis may be favorable for defining novel targets to prevent its onset.
With the possibility of being a double-edged sword, in-depth studies are required to explore how IL-33 at the peak of its expression and its receptor ST2 are involved in immune responses that S. japonicum elicits. Since the infected host is unable to clear the parasite, the body, especially the liver, is confronted with ongoing cycles of inflammation and healing, leading to immunopathology and tissue fibrosis.58 Therefore, hosts infected with Schistosoma have developed a variety of strategies in its regulatory mechanisms to inflammatory responses to newly deposited eggs, so as to mitigate damages caused by excessive immune responses.7 Numerous studies have proved that Treg is the most important negative immune regulator and plays a pivotal role in limiting hepatic immunopathological damage during the development of schistosomiasis.7,29,59,60 Treg is one of the main immune cells with high expression of ST2 on its surface. Recent studies have found that activation of Treg is associated with the IL-33/ST2 axis in many inflammatory diseases, such as inflammatory bowel disease, transplantation, immune-mediated hepatitis and experimental arthritis.33,34,61,62 Since S. japonicum infection induced elevated expression of the IL-33/ST2 axis, which could reduce the hepatic immunopathology in mice, we were to verify whether the protective role of the IL-33/ST2 axis was associated with its regulation of Treg. The data presented indicated that frequencies of both hepatic and splenic Treg and ST2+Treg were downregulated in the infected gene depletion mice with either IL-33 or ST2 relative to the infected WT mice, which were consistent with increased mRNA expression levels of their corresponding biomarkers Foxp3 and ST2. In addition, depletion of the IL-33/ST2 axis inhibited expressions of Treg functional cytokines TGF-β1 and IL-10, in both protein and mRNA levels. Notably, exogenous rmIL-33 treatment could reverse the inhibition trend of Treg in the infected IL-33−/− mice but not in the infected ST2−/− mice. These results indicate that IL-33, with the presence of its receptor ST2, could activate Treg in S. japonicum infected mice. More importantly, IL-33 could directly induce the up-regulation of ST2 expression on the surface of Treg.63 It has been demonstrated that Treg plays a pivotal role in limiting hepatic immunopathological damage during the development of schistosomiasis.59,64 Therefore, the elevated IL-33/ST2 axis might mitigate the liver immunopathology by activating Treg in the infected mice. Additional studies may provide insights for more experimental evidence to verify and clarify the comprehensive mechanisms involved in the regulation of the axis.
IL-33 has the capacity to target several immune cells under a variety of immunological responses.52 The IL-33/ST2 axis has been found to modulate some disease progress through targeting Th17 and exert ambivalent effects on Th17 response in different diseases.37,38,64,66 The role of Th17 has attracted attention in liver pathology in murine schistosomiasis.8,67 There is significant increased IL-17 levels in the ST2−/− infected mice with S. mansoni at 14 weeks post infection.56 Our results showed that frequencies of both hepatic and splenic Th17 were upregulated in the infected IL-33−/− or ST2−/− relative to the infected WT mice, which were consistent with increased mRNA expression levels of its corresponding biomarker RORγt. In addition, analysis of the cytokine production profile revealed that depletion of the IL-33/ST2 axis resulted in the increased IL-17 level. Notably, exogenous rmIL-33 treatment could reverse the activation effect of Th17 in the infected IL-33−/− mice but not in the infected ST2−/− mice. These results suggest that the increased IL-33 could suppress Th17, and ST2 is indispensable for this inhibition effect in S. japonicum infected mice. The immune response is skewed to Th17 type in the infected ST2−/− mice with Toxocara canis.68 Our data, consistent with the previous reports, support that decreased Th17 immune response is associated with the protective role of the IL-33/ST2 axis in murine schistosomiasis.55 Further studies will be required to explore the detailed molecular mechanism of how IL-33 regulate Th17 responses in murine schistosomiasis japonica. Collectively, activation of Treg and inhibition of Th17 by IL-33 is beneficial in murine schistosomiasis. The exacerbated liver immunopathology caused by the absence of the IL-33/ST2 axis is most likely due to downregulation of Treg and upregulation of Th17 differentiation during schistosomiasis.
Increasing evidence suggests that Th17/Treg balance is critical to maintaining the immune homeostasis of bodies, and the imbalance caused by alteration of the quantity or function of Th17 or Treg plays a pathogenic role in inflammatory disorders.3,69,70 Th17 cells are actively involved in hepatic immunopathogenesis in schistosomiasis, whereas Treg cells play a pivotal role in limiting hepatic granuloma and fibrosis in schistosomiasis.8,9 Our results indicated that S. japonicum infection up-regulated the frequencies and functions of both Th17 and Treg in all mice. However, there was more dramatical increase of Treg in the infected WT mice, while much higher level of Th17 in the infected mice lacking the IL-33/ST2 axis. The result indicated that the IL-33/ST2 axis deficiency aggravated the imbalance of Th17/Treg in murine schistosomiasis. Therefore, the exacerbated liver immunopathology may be related to the functional imbalance of Th17/Treg in the infected IL-33−/− and ST2−/− mice.29 A better understanding of the detailed cellular and molecular mechanisms holds the key to development new therapeutic strategies against schistosomiasis.
Autophagy has emerged as a critical regulator in T cell differentiation and development.71 Abnormal autophagy has been characterized in immune disorders as well as infectious diseases.42 There is compelling evidence that autophagy serves as a central signal-dependent controller for Treg in different disease settings.72,73 IL-33 is enriched in chronic hepatitis B virus (HBV) infection and has a context-dependent role in autophagy.74 In our study, we demonstrated that IL-33 could promote naïve CD4+ T cells to differentiate into Treg in vitro by upregulating autophagy. Induction of Treg by IL-33 through activating autophagy may be due to the fact that Treg constitutively expresses the receptor ST2 and thus are able to act immediately upon IL-33 treatment.63 Autophagy has been reported to alleviate liver inflammation in S. japonicum-infected mice.75 Therefore, the protective role of the IL-33/ST2 axis in hepatic immunopathology might be associated with the increase of Treg, resulted from upregulation of autophagy by IL-33 in the infected mice. This hypothesis was further strengthened by the observation that the IL‐33/ST2 deficiency led to decreased Treg and thus aggravated hepatic pathology in the infected mice.
Conclusion
In summary, we demonstrate that the IL‐33/ST2 axis plays a protective function through induction of Treg and inhibition of Th17 in S. japonicum infected mice. IL-33 promotes naïve CD4+ T cells to differentiate into Treg by upregulating autophagy in vitro, although the underlying mechanism is needed for further exploration. Our findings may open new perspectives for better understanding immune mechanisms in schistosomiasis.
Acknowledgments
We thank Dr. Yan Gan from Barrow Neurological Institute, St. Joseph’s Hospital and Medical Center (Phoenix, USA) for her critical readings of this manuscript. This work was funded by grants from the National Nature Science Foundation of China (grant 81772220 to Jiahui Lei) and the Fundamental Research Funds for the Central Universities (grant HUST 2016YXMS199 to Jiahui Lei).
Disclosure
The authors state no conflicts of interest in this work.
References
1. LoVerde PT. Schistosomiasis. Adv Exp Med Biol. 2019;1154:45–70.
2. WHO, schistosomiasis fact sheet; 2020. Available from: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis.
3. Chuah C, Jones MK, Burke ML, McManus DP, Gobert GN. Cellular and chemokine-mediated regulation in schistosome-induced hepatic pathology. Trends Parasitol. 2014;30(3):141–150. doi:10.1016/j.pt.2013.12.009
4. McManus DP, Bergquist R, Cai P, Ranasinghe S, Tebeje BM, You H. Schistosomiasis-from immunopathology to vaccines. Semin Immunopathol. 2020;42(3):355–371. doi:10.1007/s00281-020-00789-x
5. Takaki KK, Rinaldi G, Berriman M, Pagán AJ, Ramakrishnan L. Schistosoma mansoni eggs modulate the timing of granuloma formation to promote transmission. Cell Host Microbe. 2021;29(1):58–67.e5. doi:10.1016/j.chom.2020.10.002
6. Santana JB, de Almeida TVVS, Lopes DM, et al. Phenotypic characterization of CD4+ T lymphocytes in periportal fibrosis secondary to schistosomiasis. Front Immunol. 2021;12:605235. doi:10.3389/fimmu.2021.605235
7. Zhang Y, Xiong DH, Li Y, et al. Schistosoma japonicum infection in treg-specific USP21 knockout mice. J Immunol Res. 2021;2021:6613162. doi:10.1155/2021/6613162
8. Zheng B, Zhang J, Chen H, et al. T lymphocyte-mediated liver immunopathology of schistosomiasis. Front Immunol. 2020;11:61. doi:10.3389/fimmu.2020.00061
9. Angeles JMM, Mercado VJP, Rivera PT. Behind enemy lines: immunomodulatory armamentarium of the schistosome parasite. Front Immunol. 2020;11:1018. doi:10.3389/fimmu.2020.01018
10. Tang CL, Lei JH, Wang T, et al. Effect of CD4+ CD25+ regulatory T cells on the immune evasion of Schistosoma japonicum. Parasitol Res. 2011;108(2):477–480. doi:10.1007/s00436-010-2089-2
11. Herbert DR, Orekov T, Perkins C, Finkelman FD. IL-10 and TGF-beta redundantly protect against severe liver injury and mortality during acute schistosomiasis. J Immunol. 2008;181(10):7214–7220. doi:10.4049/jimmunol.181.10.7214
12. Shrestha S, Yang K, Guy C, Vogel P, Neale G, Chi H. Treg cells require the phosphatase PTEN to restrain TH1 and TFH cell responses. Nat Immunol. 2015;16(2):178–187. doi:10.1038/ni.3076
13. Turner JD, Jenkins GR, Hogg KG, et al. CD4+CD25+ regulatory cells contribute to the regulation of colonic Th2 granulomatous pathology caused by schistosome infection. PLoS Negl Trop Dis. 2011;5(8):e1269. doi:10.1371/journal.pntd.0001269
14. Ahmad SF, Ansari MA, Nadeem A, et al. Dysregulation of the expression of HLA-DR, costimulatory molecule, and chemokine receptors on immune cells in children with autism. Int Immunopharmacol. 2018;65:360–365. doi:10.1016/j.intimp.2018.10.027
15. Ahmad SF, Ansari MA, Nadeem A, et al. Dysregulation of T cell immunoglobulin and mucin domain 3 (TIM-3) signaling in peripheral immune cells is associated with immune dysfunction in autistic children. Mol Immunol. 2019;106:77–86. doi:10.1016/j.molimm.2018.12.020
16. Molofsky AB, Savage AK, Locksley RM. Interleukin-33 in tissue homeostasis, injury, and inflammation. Immunity. 2015;42(6):1005–1019. doi:10.1016/j.immuni.2015.06.006
17. Stanczak MA, Sanin DE, Apostolova P, et al. IL-33 expression in response to SARS-CoV-2 correlates with seropositivity in COVID-19 convalescent individuals. Nat Commun. 2021;12(1):2133. doi:10.1038/s41467-021-22449-w
18. Rostan O, Arshad MI, Piquet-Pellorce C, Robert-Gangneux F, Gangneux JP, Samson M. Crucial and diverse role of the interleukin-33/ST2 axis in infectious diseases. Infect Immun. 2015;83(5):1738–1748. doi:10.1128/IAI.02908-14
19. Biton J, Khaleghparast Athari S, Thiolat A, et al. In vivo expansion of activated Foxp3+ Regulatory T cells and establishment of a type 2 immune response upon IL-33 treatment protect against experimental arthritis. J Immunol. 2016;197(5):1708–1719. doi:10.4049/jimmunol.1502124
20. Yu Y, Deng W, Lei J. Interleukin-33 promotes Th2 immune responses in infected mice with Schistosoma japonicum. Parasitol Res. 2015;114(8):2911–2918. doi:10.1007/s00436-015-4492-1
21. Peng H, Zhang Q, Li X, et al. IL-33 contributes to Schistosoma japonicum-induced hepatic pathology through induction of M2 macrophages. Sci Rep. 2016;6:29844. doi:10.1038/srep29844
22. Mukendi JPK, Nakamura R, Uematsu S, Hamano S. Interleukin (IL)-33 is dispensable for Schistosoma mansoni worm maturation and the maintenance of egg-induced pathology in intestines of infected mice. Parasit Vectors. 2021;14(1):70. doi:10.1186/s13071-020-04561-w
23. Kamdem SD, Konhawa F, Kuemkon EM, et al. Negative association of interleukin-33 plasma levels and schistosomiasis infection in a site of polyparasitism in rural Cameroon. Front Immunol. 2019;10:2827. doi:10.3389/fimmu.2019.02827
24. Wilson S, Jones FM, Fofana HK, et al. A late IL-33 response after exposure to Schistosoma haematobium antigen is associated with an up-regulation of IL-13 in human eosinophils. Parasite Immunol. 2013;35(7–8):224–228. doi:10.1111/pim.12035
25. Wen X, He L, Chi Y, et al. Dynamics of Th17 cells and their role in Schistosoma japonicum infection in C57BL/6 mice. PLoS Negl Trop Dis. 2011;5(11):e1399. doi:10.1371/journal.pntd.0001399
26. Xiao Y, Lai L, Chen H, et al. Interleukin-33 deficiency exacerbated experimental autoimmune encephalomyelitis with an influence on immune cells and glia cells. Mol Immunol. 2018;101:550–563. doi:10.1016/j.molimm.2018.08.026
27. Zhang Z, Zhang N, Shi J, et al. Allograft or recipient ST2 deficiency oppositely affected cardiac allograft vasculopathy via differentially altering immune cells infiltration. Front Immunol. 2021;12:657803. doi:10.3389/fimmu.2021.657803
28. Zhao F, Huang X, Hou X, et al. Schistosoma japonicum: susceptibility of neonate mice born to infected and noninfected mothers following subsequent challenge. Parasite Immunol. 2013;35(5–6):157–163. doi:10.1111/pim.12027
29. Guan F, Zhang C, Jiang C, et al. ApoE deficiency promotes hepatic pathology by aggravating Th17/Treg imbalance in murine schistosomiasis japonica. Parasite Immunol. 2020;42(12):e12785. doi:10.1111/pim.12785
30. Xu L, Li W, Wang X, et al. The IL-33-ST2-MyD88 axis promotes regulatory T cell proliferation in the murine liver. Eur J Immunol. 2018;48(8):1302–1307. doi:10.1002/eji.201747402
31. Kabat AM, Harrison OJ, Riffelmacher T, et al. The autophagy gene Atg16l1 differentially regulates Treg and TH2 cells to control intestinal inflammation. Elife. 2016;5:e12444. doi:10.7554/eLife.12444
32. Guan F, Jiang W, Bai Y, et al. Purinergic P2X7 receptor mediates the elimination of Trichinella spiralis by activating NF-κB/NLRP3/IL-1β pathway in macrophages. Infect Immun. 2021;89(5):e00683–20. doi:10.1128/IAI.00683-20
33. Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol. 2016;16(11):676–689. doi:10.1038/nri.2016.95
34. Schiering C, Krausgruber T, Chomka A, et al. The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature. 2014;513(7519):564–568. doi:10.1038/nature13577
35. Hung LY, Tanaka Y, Herbine K, et al. Cellular context of IL-33 expression dictates impact on anti-helminth immunity. Sci Immunol. 2020;5(53):eabc6259. doi:10.1126/sciimmunol.abc6259
36. Pastille E, Wasmer MH, Adamczyk A, et al. The IL-33/ST2 pathway shapes the regulatory T cell phenotype to promote intestinal cancer. Mucosal Immunol. 2019;12(4):990–1003. doi:10.1038/s41385-019-0176-y
37. Qiu X, Qi C, Li X, Fang D, Fang M. IL-33 deficiency protects mice from DSS-induced experimental colitis by suppressing ILC2 and Th17 cell responses. Inflamm Res. 2020;69(11):1111–1122. doi:10.1007/s00011-020-01384-4
38. Zhu JF, Xu Y, Zhao J, et al. IL-33 protects mice against DSS-induced chronic colitis by increasing both regulatory B cell and regulatory T cell responses as well as decreasing Th17 cell response. J Immunol Res. 2018;2018:1827901. doi:10.1155/2018/1827901
39. Wu MX, Wang SH, Xie Y, et al. Interleukin-33 alleviates diabetic cardiomyopathy through regulation of endoplasmic reticulum stress and autophagy via insulin-like growth factor-binding protein 3. J Cell Physiol. 2021;236(6):4403–4419. doi:10.1002/jcp.30158
40. Wang Z, Wu L, Pan B, Chen Y, Zhang T, Tang N. Interleukin 33 mediates hepatocyte autophagy and innate immune response in the early phase of Acetaminophen-induced acute liver injury. Toxicology. 2021;456:152788. doi:10.1016/j.tox.2021.152788
41. Wang Z, Shi L, Hua S, Qi C, Fang M. IL-33 ameliorates experimental colitis involving regulation of autophagy of macrophages in mice. Cell Biosci. 2019;9:10. doi:10.1186/s13578-019-0271-5
42. Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation and immunity. Nat Rev Immunol. 2013;13(10):722–737. doi:10.1038/nri3532
43. Jacquin E, Apetoh L. Cell-intrinsic roles for autophagy in modulating CD4 T cell functions. Front Immunol. 2018;9:1023. doi:10.3389/fimmu.2018.01023
44. Bjørkøy G, Lamark T, Pankiv S, Øvervatn A, Brech A, Johansen T. Monitoring autophagic degradation of p62/SQSTM1. Methods Enzymol. 2009;452:181–197.
45. Zhou Z, Yan F, Liu O. Interleukin (IL)-33: an orchestrator of immunity from host defence to tissue homeostasis. Clin Transl Immunol. 2020;9(6):e1146. doi:10.1002/cti2.1146
46. Liew FY, Pitman NI, McInnes IB. Disease-associated functions of IL-33: the new kid in the IL-1 family. Nat Rev Immunol. 2010;10(2):103–110. doi:10.1038/nri2692
47. Humphreys NE, Xu D, Hepworth MR, Liew FY, Grencis RK. IL-33, a potent inducer of adaptive immunity to intestinal nematodes. J Immunol. 2008;180(4):2443–2449. doi:10.4049/jimmunol.180.4.2443
48. Peng H, Sun R, Zhang Q, et al. Interleukin 33 mediates type 2 immunity and inflammation in the central nervous system of mice infected with Angiostrongylus cantonensis. J Infect Dis. 2013;207(5):860–869. doi:10.1093/infdis/jis682
49. Rostan O, Gangneux JP, Piquet-Pellorce C, et al. The IL-33/ST2 axis is associated with human visceral leishmaniasis and suppresses Th1 responses in the livers of BALB/c mice infected with Leishmania donovani. mBio. 2013;4(5):e00383–13. doi:10.1128/mBio.00383-13
50. Chen RQ, Zhou YB, Jiang LF, et al. Correlation between acute schistosomiasis japonica and serum interleukin-33 levels. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2012;24(1):32–34, 39.
51. Li ZY, Xiao L, Lin G, et al. Contribution of tissue transglutaminase to the severity of hepatic fibrosis resulting from Schistosoma japonicum infection through the regulation of IL-33/ST2 expression. Parasit Vectors. 2019;12(1):302. doi:10.1186/s13071-019-3542-4
52. Alvarez F, Fritz JH, Piccirillo CA. Pleiotropic effects of IL-33 on CD4(+) T cell differentiation and effector functions. Front Immunol. 2019;10:522. doi:10.3389/fimmu.2019.00522
53. Hung LY, Lewkowich IP, Dawson LA, et al. IL-33 drives biphasic IL-13 production for noncanonical Type 2 immunity against hookworms. Proc Natl Acad Sci U S A. 2013;110(1):282–287. doi:10.1073/pnas.1206587110
54. Clark JT, Christian DA, Gullicksrud JA, et al. IL-33 promotes innate lymphoid cell-dependent IFN-γ production required for innate immunity to Toxoplasma gondii. Elife. 2021;10:e65614. doi:10.7554/eLife.65614
55. Cao S, Zhu L, Zhu C, et al. Helicobacter hepaticus infection-induced IL-33 promotes hepatic inflammation and fibrosis through ST2 signaling pathways in BALB/c mice. Biochem Biophys Res Commun. 2020;525(3):654–661. doi:10.1016/j.bbrc.2020.02.139
56. Maggi L, Rocha IC, Camelo GMA, Fernandes VR, Negrão-Corrêa D. The IL-33/ST2 pathway is not essential to Th2 stimulation but is key for modulation and survival during chronic infection with Schistosoma mansoni in mice. Cytokine. 2021;138:155390. doi:10.1016/j.cyto.2020.155390
57. Barbier L, Ferhat M, Salamé E, et al. Interleukin-1 family cytokines: keystones in liver inflammatory diseases. Front Immunol. 2019;10:2014. doi:10.3389/fimmu.2019.02014
58. Fairfax KC, Amiel E, King IL, Freitas TC, Mohrs M, Pearce EJ. IL-10R blockade during chronic schistosomiasis mansoni results in the loss of B cells from the liver and the development of severe pulmonary disease. PLoS Pathog. 2012;8(1):e1002490. doi:10.1371/journal.ppat.1002490
59. Ondigo BN, Ndombi EM, Nicholson SC, et al. Functional studies of T regulatory lymphocytes in human schistosomiasis in Western Kenya. Am J Trop Med Hyg. 2018;98(6):1770–1781. doi:10.4269/ajtmh.17-0966
60. Kong H, He J, Guo S, et al. Endothelin receptors promote schistosomiasis-induced hepatic fibrosis via splenic B cells. PLoS Pathog. 2020;16(10):e1008947. doi:10.1371/journal.ppat.1008947
61. Faustino LD, Griffith JW, Rahimi RA, et al. Interleukin-33 activates regulatory T cells to suppress innate gammadelta T cell responses in the lung. Nat Immunol. 2020;21(11):1371–1383. doi:10.1038/s41590-020-0785-3
62. Kawai K, Uchiyama M, Hester J, Issa F. IL-33 drives the production of mouse regulatory T cells with enhanced in vivo suppressive activity in skin transplantation. Am J Transplant. 2021;21(3):978–992. doi:10.1111/ajt.16266
63. Liu Q, Turnquist HR. Controlling the burn and fueling the fire: defining the role for the alarmin interleukin-33 in alloimmunity. Curr Opin Organ Transplant. 2016;21(1):45–52. doi:10.1097/MOT.0000000000000265
64. Zhou S, Qi Q, Wang X, et al. SjHSP60 induces CD4+ CD25+ Foxp3+ Tregs via TLR4-Mal-drived production of TGF-β in macrophages. Immunol Cell Biol. 2018;96(9):958–968. doi:10.1111/imcb.12160
65. Terraza C, Fuentes R, Pino-Lagos K. IFN-gamma and IL-33 modulate mesenchymal stem cells function targeting Th1/Th17 axis in a murine skin transplantation model. Cytokine. 2018;111:317–324. doi:10.1016/j.cyto.2018.09.013
66. Jiang HR, Milovanović M, Allan D, et al. IL-33 attenuates EAE by suppressing IL-17 and IFN-gamma production and inducing alternatively activated macrophages. Eur J Immunol. 2012;42(7):1804–1814. doi:10.1002/eji.201141947
67. Zhou W, Yang Y, Mei C, et al. Inhibition of rho-kinase downregulates Th17 cells and ameliorates hepatic fibrosis by Schistosoma japonicum infection. Cells. 2019;8(10):1262. doi:10.3390/cells8101262
68. Leal-Silva T, Vieira-Santos F, Oliveira FMS, et al. Detrimental role of IL-33/ST2 pathway sustaining a chronic eosinophil-dependent Th2 inflammatory response, tissue damage and parasite burden during Toxocara canis infection in mice. PLoS Negl Trop Dis. 2021;15(7):e0009639. doi:10.1371/journal.pntd.0009639
69. Bakheet SA, Ansari MA, Nadeem A, et al. CXCR3 antagonist AMG487 suppresses rheumatoid arthritis pathogenesis and progression by shifting the Th17/Treg cell balance. Cell Signal. 2019;64:109395. doi:10.1016/j.cellsig.2019.109395
70. Zhang W, Liu X, Zhu Y, et al. Transcriptional and posttranslational regulation of Th17/Treg balance in health and disease. Eur J Immunol. 2021;51(9):2137–2150. doi:10.1002/eji.202048794
71. Wang L, Das JK, Kumar A, et al. Autophagy in T-cell differentiation, survival and memory. Immunol Cell Biol. 2021;99(4):351–360. doi:10.1111/imcb.12422
72. Zhang J, Chen L, Xiong F, et al. Autophagy in regulatory T cells: a double-edged sword in disease settings. Mol Immunol. 2019;109:43–50. doi:10.1016/j.molimm.2019.02.004
73. An N, Chen Y, Wang C, et al. Chloroquine autophagic inhibition rebalances Th17/Treg-mediated immunity and ameliorates systemic lupus erythematosus. Cell Physiol Biochem. 2017;44(1):412–422. doi:10.1159/000484955
74. Cheng LS, Li J, Liu Y, et al. HMGB1-induced autophagy: a new pathway to maintain Treg function during chronic hepatitis B virus infection. Clin Sci (Lond). 2017;131(5):381–394. doi:10.1042/CS20160704
75. Zhu J, Zhang W, Zhang L, et al. IL-7 suppresses macrophage autophagy and promotes liver pathology in Schistosoma japonicum-infected mice. J Cell Mol Med. 2018;22(7):3353–3363. doi:10.1111/jcmm.13610
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.