")
Back to Journals » International Journal of General Medicine » Volume 16
Identification of MiR-223 Associated with Diagnosis in Ectopic Pregnancy
Authors Qiu J , Chen J, Deng G, Yuan S
Received 31 March 2023
Accepted for publication 20 June 2023
Published 27 June 2023 Volume 2023:16 Pages 2693—2705
DOI https://doi.org/10.2147/IJGM.S412439
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Woon-Man Kung
Jiahan Qiu,1,2 Jiaxun Chen,2 Gaopi Deng,1,* Shuo Yuan1,*
1Department of Gynaecology, the First Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 2First Clinical Medical School, Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shuo Yuan, Department of Gynaecology, the First Affiliated Hospital of Guangzhou University of Chinese Medicine, Jichang Road 16#, District Baiyun, Guangzhou, 510405, People’s Republic of China, Tel +86-13632466654, Email [email protected]
Background: In this study, we conducted an integrated study of the diagnostic value of MiR-223 in ectopic pregnancy (EP).
Methods: We used GSE44731 downloaded from GEO and GEO2R to identify differentially expressed miRNA. The hub genes corresponding to the differential miRNA were then identified by using the Xiantao academic tool, GO (Gene Ontology), and KEGG (Kyoto Encyclopedia of Genes and Genomes). Afterward, we used the miEAA database to perform gene set enrichment analysis (GSEA) of differential miRNA, and used Xiantao academic tools again to conduct the ceRNA network based on the target genes. Protein–protein interaction (PPI) network construction and lncRNA of hub miRNA target genes were then predicted by the starbase database. For validation, the villus tissue from intrauterine pregnancy and tubal pregnancy was collected and assayed by qPCR.
Results: In total 19 differentially expressed miRNAs were screened out, of which MiR-223 had a relatively clear diagnostic significance. Hub genes were enriched and analyzed by GO, KEGG, and GSEA, and the results showed that regulation of NF-κB and other signaling pathways are primarily enriched in ectopic pregnancy. We also obtained 215 key genes from PPI analysis. Our ceRNA analysis indicated that LRRC75A-AS1 and PITPNA-AS1 were associated with MiR-223, and the expression of MiR-223 in qPCR was significantly high in tubal pregnancy group.
Conclusion: We found that MiR-223 can be used in the diagnosis of EP. Our findings provide valuable information and direction for future research into novel targets for EP diagnosis.
Keywords: MiRNA-223, tubal pregnancy, disease marker, biomarker, GEO
Introduction
As the leading cause of maternal mortality in early pregnancy, ectopic pregnancy (EP) is responsible for 4–10% of all pregnancy-related deaths,1 but drugs can only be used to treat early EP. However, significant differences in the effects on fertility between surgical treatment and medical treatment have been reported in the literature, suggesting that more diagnosis markers are needed for the early stages of EP in order to reduce the risk of disease.
Early diagnosis and treatment of EP are very important. However, early detection of EP is not easy since the initial signs and symptoms, such as lack of a menstrual period, low abdominal pain, and vaginal bleeding, are often ambiguous and usually seen in a normal pregnancy as well.
MicroRNAs (miRNAs) are a class of short (21–25 nt), on-coding RNAs, that control their target genes’ expression at the post-transcriptional level.2 MiRNAs have been implicated in the regulation of various biological functions, including cell proliferation, apoptosis, and cell differentiation.3 Thus, miRNAs can be used as biomarkers in disease diagnoses. However, there are only a few studies on EP and miRNAs, and none of them are related to the diagnosis of the disease.
Recently, MiR-223 has become one of a major focus of study in many different areas.4 As a key component of innate immunity and dysregulation, its expression is involved in the pathogenesis of multiple inflammatory diseases and cancers.5 However, the mechanism of action of MiR-223 in EP remains poorly understood, and there is only a small number of basic research studies on the subject. In this study, we adopt data from the gene expression omnibus (GEO) to identify differentially expressed genes (DEGs). DEGs were performed pathway analyses by Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis, combining with Gene Set Enrichment Analysis (GSEA). Then, we predicted the target genes of DEmiRNAs and the lncRNA and constructing of the molecule–molecule network and determine their diagnostic value for EP.
Materials and Methods
Data
The chip dataset was downloaded and retrieved from the GEO database GSE 44731,6 which provided us with Homo sapiens gene samples. The platform for this data was built based on GPL14767 Agilent-021827 Human miRNA Microarray G4470C. The data set included eight patients with tubal EP were selected as the experimental group for this study and eight additional patients with normal pregnancies planned for voluntary termination of pregnancy were used as the control group.
Identification of Differentially Expressed miRNAs
The GEO2R7 (https://www.ncbi.nlm.nih.gov/geo) website is an interactive web tool for the differential analysis of expression spectrum chips based on the GEO database and allows users to get results by comparing two or more datasets in the GEO series. This time we use the GEOquery8 and limma9 R packages in the Bioconductor project8–10 to acquire comparisons of those processed data tables provided by the original author.
In this study, we carried out differentially expressed analysis of our dataset using GEO2R for dataset GSE44731 with default values for the options parameters. The differential expression miRNA screening restriction was P < 0.05 and |logFC|> 1, where only the matched probe was retained in the expression profile during re-annotation, as well as the probe with the largest differential expression multiple when the same miRNA matched multiple probes. Subsequently, we used the Xiantao academic tools (https://www.xiantao.love/) to create volcano and heat maps of differentially expressed miRNAs (DEmiRNAs) in order to analyze their differential expression visually. In addition, we evaluated the diagnostic efficacy of DEmiRNAs. To do this we drew the ROC curve again using the Xiantao academic tool and calculated the area below the ROC curve (AUC). We used the AUC to evaluate the credibility of the diagnostic curve, where AUC > 0.9 for hub DEmiRNAs.
The Target Genes of DEmiRNAs and lncRNA Prediction
To find microRNA targets we searched the Starbase database (http://starbase.sysu.edu.cn/) using high-throughput CLIP-Seq experimental data and degradation group experimental data, and we used a wide variety of visual interfaces that were provided by the database publisher to explore microRNA targets. The database contains rich data of miRNA–ncRNA, miRNA–mRNA, RBP–RNA, and RNA–RNA, with multiple forecasting tools (PITA) available such as RNA22, miRmap, DIANA-microT, miRanda, PicTar, and TargetScan that can be used to search for miRNA–mRNA interactions.11 Using this data and our above analysis, we predicted the target genes of hub DEmiRNAs and the lncRNA in combination. The parameters for our search were set to Clade (mammal), Genome (human), Assembly (hg19), CLIP-Data (≥1), Degradome-Data (≥3), pan-Cancer (≥0), program Num (≥3), program (None), and separately Clade (mammal), Genome (human), Assembly (hg19), CLIP-Data (≥1), Degradome-Data (≥1), and pan-Cancer (≥0).
GO and KEGG Enrichment Analysis of Target Genes in DEmiRNAs
Gene Ontology (GO) is a major bioinformatics tool for annotating genes and studying the biological processes of these genes, including molecular function (MF), biological processes (BPs), and cellular composition (CC).12 The Kyoto Encyclopedia of Genes and Genomes (KEGG) is a database resource for understanding advanced functions and biological systems by generating large-scale molecular datasets through high-throughput experimental technologies.13 In order to defining target genes, we subjected them for hub miRNA predicted by the StarBbase database to GO annotation and KEGG pathway enrichment analysis using the Xiantao academic tool. We considered a p-value <0.05 to indicate a statistically significant test result.
GSEA
The miEAA (https://ccb-compute2.cs.uni-saarland.de/mieaa2/) is an online tool for miRNA enrichment analysis with annotations that contribute to the functional analysis of miRNA sets. This tool is tailored for miRNA precursors and mature miRNAs in multiple species, including humans, mice, and rats.14 We took the gene set enrichment analysis (GSEA) of differential miRNA from the miEAA database, and selected “Pathway (KEGG)” and “Gene Ontology (miRWalk)” as the reference gene sets and considered adjusted p-values <0.05 and p-values <0.05 to indicate statistically significant enrichment.
Construction of a Molecule–Molecule Network
The STRING (https://string-db.org/) is a database for searching for known interactions between predicted proteins that integrates other databases data and also house predictive results from bioinformatics methods.15 This tool also contains experimental data and text mining results from summaries of articles on PubMed (https://pubmed.ncbi.nlm.nih.gov) summaries. STRING was used to construct the protein–protein interaction (PPI) network of hub miRNA target genes. The ceRNA networks of miRNA–mRNA, 1ncRNA–miRNA, and 1ncRNA–miRNA were constructed by Xiantao academic tools based on the target genes and lncRNA predicted by the Starbase database, and we found a regulatory relationship between the nodes of the ceRNA networks.
Specimen Collection
In this research, the protocol was approved by ethics review boards at the First Affiliated Hospital of Guangzhou University of Chinese Medicine. The intrauterine pregnancy group consisted of patients undergoing termination of pregnancy entirely voluntarily in the day operating room in the First Affiliated Hospital of Guangzhou University of Chinese Medicine from January to March 2022, and the tubal pregnancy group consisted of patients undergoing laparoscopic surgery to treat the ectopic pregnancy from January to March 2022 in the inpatient department of Guangzhou First Affiliated Hospital. Part of the villous tissue in both groups (washed 3 times in PBS) from the above two groups were collected and stored in a 4°C refrigerator in a centrifugal tube equipped with RNA preservation solution and then transferred to a −18°C refrigerator for 1 month.
The Expressions of MiR-223 as Determined by qPCR
In order to validation data-set, we extracted total trophoblast RNA by TRIzol, reverse transcribed cDNA according to the measured RNA concentration, and amplified cDNA by qPCR using cDNA as the template. The primers were synthesized by Guangzhou Xinli, He Biotechnology Co., Ltd., using the following reaction conditions: 95°C pre-denatured DNA for 10 min; 95°C denatured DNA for 10s, and 55°C annealing for 20s, for a total of 40 cycles. The internal reference used was U6, and the results were calculated using the 2−ΔΔCT method. Primer sequences are shown in Table 1.
 |
Table 1 The Sequence of PCR Primers |
Statistical Methods
The data obtained in 2.8 were analyzed using the R language (version 3.6.3) for statistical analysis and data visualization using the ggplot2 package (version 3.3.3) with the R language (version 3.6.3), respectively.
Results
Differentially Expressed miRNAs
The workflow of the study is displayed in Figure 1.
 |
Figure 1 Study workflow. |
We obtained 19 DEmiRNAs molecules were obtained from the differential analysis of the GSE44731 dataset, including 7 upregulated miRNAs and 12 downregulated miRNAs. Figure 2A is a volcano plot of DEmiRNAs marking the Top 3 upregulated and downregulated miRNAs. This figure also contains the heat map (Figure 2) drawn with DEmiRNAs.
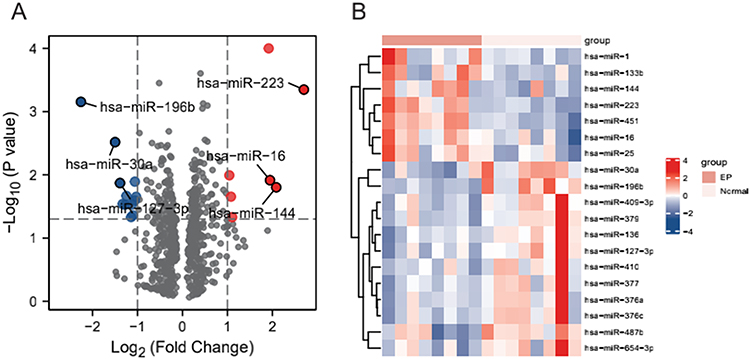 |
Figure 2 Differentially expressed miRNAs. (A) Volcano map showing the miRNAs molecules obtained from the differential analysis of the GSE44731 dataset, red represents upregulated genes and blue represents downregulated genes. (B) Heat map showing the miRNA molecules obtained from the differential analysis of the GSE44731 dataset. |
To screen hub miRNA further, we plotted the diagnostic ROC curves against the 19 miRNAs molecules obtained from our differential analysis. As shown in Figure 3, we found that four miRNAs had relatively high diagnostic accuracy for EP (AUC above 0.9): miRNA-30a, miRNA-196b, MiR-223, and miRNA-451.
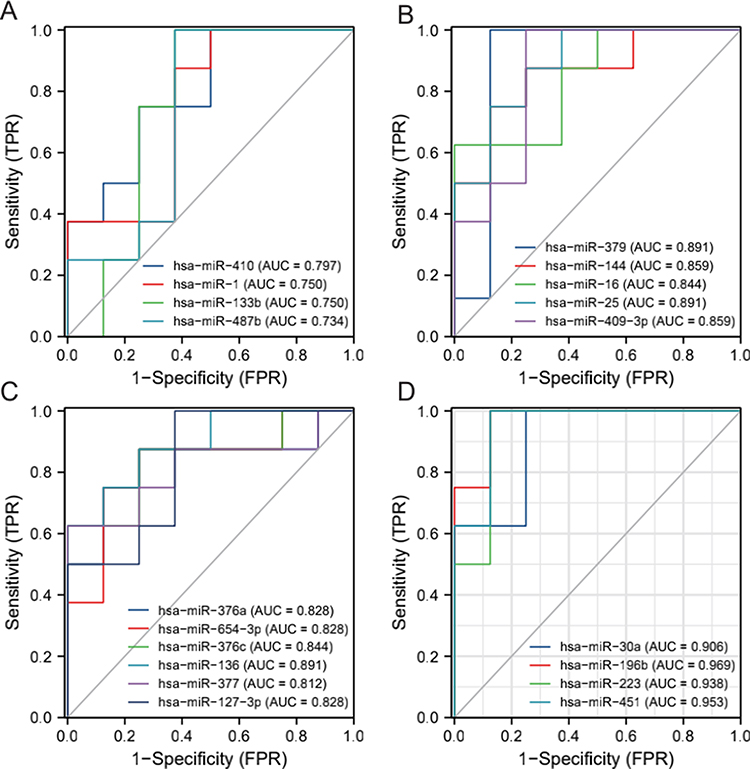 |
Figure 3 ROC for Differentially expressed miRNA molecules. (A) miRNA-410/miRNA-1/miRNA-133b/miRNA-487b; (B) miRNA-379/miRNA-144/miRNA-16/miRNA-25/miRNA-409-3p; (C) miRNA-376a/miRNA-645-3p/miRNA-376c/miRNA-136/miRNA-377/miRNA-127-3p; (D) miRNA-30a/miRNA-196b/MiR-223/. |
GO and KEGG Analysis
The results from our GO and KEGG enrichment analysis of the target genes predicted by hub miRNAs are shown in (Tables 2–5 and Figure 4). The results of GO Analysis in Biological Process implied that the target genes of hub miRNA in the GSE44731 dataset were primarily involved in the regulation of protein polymerization, the positive regulation of viral genome replication, clathrin coat assembly, the positive regulation of viral life cycle, the regulation of actin filament polymerization, clathrin-dependent endocytosis, antigen processing and presentation of exogenous peptide antigen via the MHC class II low-density lipoprotein receptor particle metabolic process, antigen processing, and presentation of peptide antigen via MHC class II, and antigen processing and presentation of peptide or polysaccharide antigen via MHC class II (Figure 4A). The results of GO Analysis in Cellular Component are related to focal adhesion, neuronal cell body, clathrin-coated vesicle, cell–substrate junction and cell–substrate adherens junction (Figure 4B). Finally, the results of GO Analysis in Molecular Function implied that adhesion molecule binding, cadherin binding, small GTPase binding, actin binding, Ras GTPase binding, and phosphatidylinositol phosphate binding (Figure 4C). However, our KEGG results showed that the target genes of hub miRNA in the GSE44731 dataset were primarily enriched in bacterial invasion of epithelial cells, Yersinia infection, tight junction, regulation of actin cytoskeleton, Salmonella infection, leukocyte transendothelial migration, focal adhesion, shigellosis, 2-oxocarboxylic acid metabolism, and endocytosis (Figure 4D).
 |
Table 2 BP (Biological Process) of GSE44731 Dataset |
 |
Table 3 CC (Cellular Component) of GSE44731 Dataset |
 |
Table 4 MF (Molecular Function) of GSE44731 Dataset |
 |
Table 5 KEGG (Kyoto Encyclopedia of Genes and Genomes) of GSE44731 Dataset |
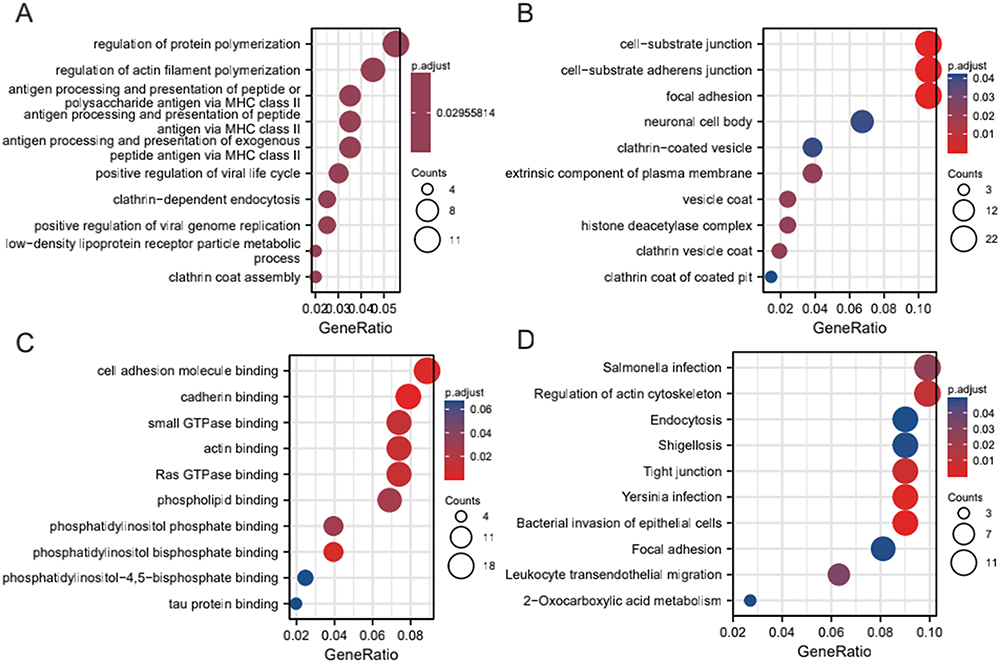 |
Figure 4 GO and KEGG enrichment analysis of DEM target genes. (A) Biological processes enriched (BPs) in the GSE44731 dataset; (B) Cell compositions enriched in GSE44731 dataset (CCs); (C) Molecular functions enriched in the GSE44731 dataset (MFs); (D) Signaling pathways enriched in the GSE44731 dataset (KEGG). |
GSEA
For GSEA, we selected pathway (KEGG) and GO (miRWalk) as reference gene sets and regarded p-adjusted <0.05 and p-value <0.05 as indications of statistically significant essential enrichment. We performed the analysis on the GSE44731 dataset, and the results are displayed in Table 6 and Figure 5. The results show that hub miRNAs were mostly enriched in signaling pathways such as Th1 and Th2 cell differentiation, Prion diseases, the NF-κB signaling pathway, the chemokine signaling pathway, DNA binding, ATP binding, nucleolus, the transcription-DNA-dependent, and other biological processes.
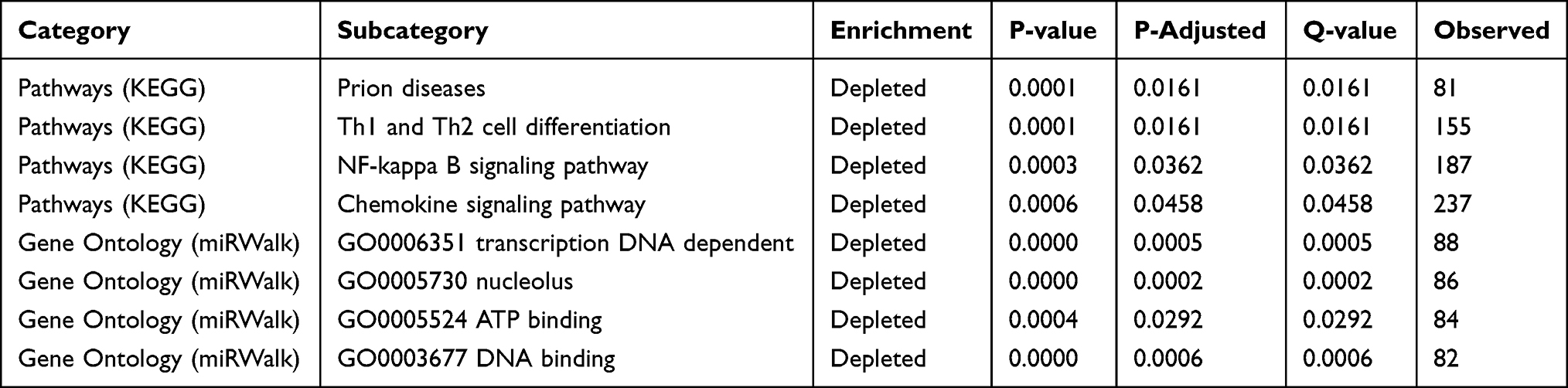 |
Table 6 GSEA of miRNA |
 |
Figure 5 GSEA of miRNAs: (A) Th1 and Th2 cell differentiation; (B) Prion diseases; (C) NF-κB signaling pathway; (D) Chemokine signaling pathway; (E) DNA binding; (F) ATP binding; (G) nucleolus; (H) transcription DNA dependent. |
Construction of the Molecule–Molecule Network
To identify potential molecular interaction relationships among target genes, the protein–protein interaction network based on the STRING database was constructed, as shown in Figure 6A. The PPI network map contains a total of 215 nodes and 458 pairs of interaction relationships. In addition, we predicted the hub miRNAs, mRNA–mRNA interaction network using the Starbase database and the ceRNA network for miRNA (Figure 6B), lncRNA-miRNA (Figure 6C), and lncRNA–miRNA–mRNA interaction networks, respectively (Figure 6D).
 |
Figure 6 Network construction. (A) PPI network of target genes; (B) miRNA–mRNA interaction network; (C) miRNA–lncRNA interaction network; (D) ceRNA network. |
Comparison of the Expression Levels of MiR-223 in the Two Groups of Cells
To study differential expression in diseases further, we performed qPCR experiments on villous tissue in utero and in ectopic pregnancy site. Compared to the intrauterine pregnancy group, the expression of MiR-223 was higher in the control group, as shown in Figure 7.
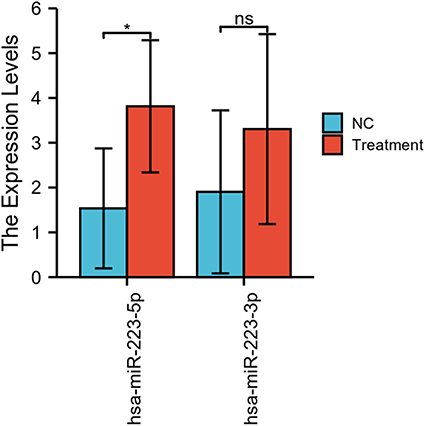 |
Figure 7 Expression of miRNA-223 in both groups of cells (ns, p≥0.05; *p <0.05). |
Discussion
Early diagnosis of EP can better preserve patients’ fertility. However, the current methods used to diagnose early EP are primarily dynamic observation of the change in HCG and the location of the amniotic sac through ultrasound, both of which require an extended period of observation.16 High-throughput sequencing to detect EP is extremely rare, as is the clinical application of its results. This study aimed to describe the miRNA of EP and intrauterine pregnancy and to provide a research basis for its use in the early clinical diagnosis of EP in the future.
The current miRNA research focuses almost exclusively on the field of oncology and immune diseases. Trophoblasts have proliferative properties similar to tumor cells, and as with alloantigen, avoid immune rejection and help maintain pregnancy continuation.17 The process of trophoblast implantation, with its proper depth, and the establishment of immune balance at the maternal and fetal interface, all involve the precise regulation of genes, and the MiR-223 gene specifically, which is located within the q12 locus of the X chromosome and has been highly conserved during human evolution.5
The long primary transcript of MiR-223 (pri-MiR-223) contains a hairpin structure in exon 3 of the non-coding transcript that principally causes production of the MiR-223-3p strand. A minor product, MiR-223-5p, is also generated that is prone to degradation yet has been shown to be involved in several diseases.5 It generates a minor product MiR-223-5p as well, which is inclined to degradation yet has been proved to be involved in different roles of various diseases. Apoptosis,18 proliferation,19 and metastasis20 have also been shown to be regulated by MiR-223, and a growing number of evidence shows that MiR-223 may work as a biomarker for carcinogenesis and cancer progression in cancers such as breast cancer,21 oral cancer,22 liver cancer,23 colon cancer,24 and lung cancer.25 By sponging MiR-223, LRRC75A-AS1 and PITPNA-AS1 can also worsen glioblastoma.26
More recently, the role of MiR-223 has also been described for numerous immunological and inflammatory disorders. Researchers have found that MiR-223 can adjust immune function27 and increase the success rate of heart transplantation,28 and that is also closely related to immune diseases such as asthma29 and multiple sclerosis.30 Currently, the research in an integrated analysis of GSE44731 dataset shows that MiR-223 has a relatively high diagnostic performance for EP, with an AUC of 0.938.
In this study, we used GSEA to research the potential signaling pathways of hub genes in EP. According to the results, EP patients were shown to be enriched in Th1 and Th2 cell differentiation, the NF-κB signaling pathway, and the chemokine signaling pathway, which are closely related to cell proliferation and immune regulation. Regulation of Th1/Th2 response also plays an important role in autoimmune diseases, and numerous miRNAs including MiR-223 are known to affect Th1 and Th2 cytokines and alter Th1/Th2 balance.8,31,32 The chemokine signaling pathway is composed of a variable set of chemokines. Abnormal regulation of chemokines can give rise to the excess inflammation characteristic of many autoimmune diseases, such as rheumatoid arthritis, psoriasis, Crohn’s disease, type-1 diabetes, and many others.33–35 Additionally, protection of chondrocyte extracellular matrix degradation by MiR-223 occurs via SDF1/CXCR4 signaling, a branched pathway of the chemokine signaling pathway.36,37 In particular, miR-223 has been unexplored in the literature until now. This study will provide new insights into the biomarker for early detection of EP.
Furthermore, the NF-κB signaling pathway has been shown to be inhibited by MiR-223, and this can delay cancer cell proliferation24 and is closely related to immune regulation as well.35 However, results acquired from computational analysis typically require further validation through wet-lab testing, such as qPCR experiments, which we also performed. In our qPCR results, we found that MiR-223 showed a high expression status in the villous tissues of EP and significant differences between the experiment and control groups. So far, there have been very few studies characterized the expression of miRNAs in EP, and none have applied it diagnostically. We believe this approach is an important direction for future research that could lead to new biomarkers for early diagnosis of EP.
The current study is not without its limitations of course. Firstly, the lack of complete clinical data in GEO results in a relatively small sample size. Hence, the statistical power is also low. Therefore, our research can benefit from larger sample sizes whose larger statistical power can be used to obtain more significant results. Secondly, despite the fact that microarray-based bioinformatics analysis is a powerful tool to effectively understand molecular mechanisms and identify potential biomarkers underlying EP, further experimental validations of MiR-223 at the molecular, cellular, and organism levels are needed.
Conclusion
In conclusion, our integrated bioinformatics analysis of EP datasets from GEO databases revealed 19 DEGs, many potential molecular mechanisms, and several key pathways involved in EP. Results of our study imply that MiR-223 may be a potential diagnostic biomarker for the very early stages of EP. Furthermore, the biological role of MiR-223 in EP has yet to be validated experimentally.
Data Availability
The data sets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Acknowledgments
This work was supported by Traditional Chinese Medicine Bureau of Guangdong Province (Grant No. 20231172) and National Natural Science Foundation of China (NSFC) (Grant No. 82174417). I also appreciate AiMi Academic Services (www.aimieditor.com) for the English language editing and review services. The study complies with the Declaration of Helsinki and was approved by the ethics committee of First Affiliated Hospital of Guangzhou University of Chinese Medicine (No. K-2021-093), and informed consent was obtained from all patients.
Disclosure
The authors declare no conflict of interest.
References
1. Chen C-Y, Hwu Y-M, Chen C-P, Chang -C-C. Quantitative analysis of total β-subunit of human chorionic gonadotropin concentration in urine by immunomagnetic reduction to assist in the diagnosis of ectopic pregnancy. Int J Nanomedicine. 2015;10:2475. doi:10.2147/IJN.S81201
2. Zhang C-M, Zhao J, Deng H-Y. MiR-155 promotes proliferation of human breast cancer MCF-7 cells through targeting tumor protein 53-induced nuclear protein 1. J Biomed Sci. 2013;20(1):1–10. doi:10.1186/1423-0127-20-79
3. Aziz F. The emerging role of miR-223 as novel potential diagnostic and therapeutic target for inflammatory disorders. Cell Immunol. 2016;303:1–6. doi:10.1016/j.cellimm.2016.04.003
4. Aziz F, Chakraborty A, Khan I, Monts J. Relevance of miR-223 as potential diagnostic and prognostic markers in cancer. Biology. 2022;11(2):249. doi:10.3390/biology11020249
5. Gu J, Xu H, Chen Y, Li N, Hou X. miR-223 as a regulator and therapeutic target in liver diseases. Front Immunol. 2022;13.
6. Dominguez F, Moreno-Moya JM, Lozoya T, et al. Embryonic miRNA profiles of normal and ectopic pregnancies. PLoS One. 2014;9(7):e102185. doi:10.1371/journal.pone.0102185
7. Clough E, Barrett T. The gene expression omnibus database. other. 2016;1:65.
8. Davis S, Meltzer PS. GEOquery: a bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics. 2007;23(14):1846–1847. doi:10.1093/bioinformatics/btm254
9. Gentleman R, Carey VJ, Huber W, Irizarry RA, Dudoit S. Bioinformatics and Computational Biology Solutions Using R and Bioconductor. Vol. 1. Springer; 2005.
10. Smyth GK. Linear models and empirical Bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol. 2004;3(1):1–25. doi:10.2202/1544-6115.1027
11. Li J-H, Liu S, Zhou H, L-H Q, Yang J-H. starBase v2. 0: decoding miRNA-ceRNA, miRNA-ncRNA and protein–RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(D1):D92–D97. doi:10.1093/nar/gkt1248
12. Ashburner M, Ball CA, Blake JA, et al. Gene ontology: tool for the unification of biology. Nat Genet. 2000;25(1):25–29. doi:10.1038/75556
13. Kanehisa M, Sato Y, Kawashima M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022;31(1):47–53. doi:10.1002/pro.4172
14. Kern F, Fehlmann T, Solomon J, et al. miEAA 2.0: integrating multi-species microRNA enrichment analysis and workflow management systems. Nucleic Acids Res. 2020;48(W1):W521–W528. doi:10.1093/nar/gkaa309
15. Szklarczyk D, Gable AL, Nastou KC, et al. The STRING database in 2021: customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021;49(D1):D605–D612. doi:10.1093/nar/gkaa1074
16. Perkins KM, Boulet SL, Kissin DM, Jamieson DJ, Group NAS. Risk of ectopic pregnancy associated with assisted reproductive technology in the United States, 2001–2011. Obstet Gynecol. 2015;125(1):70. doi:10.1097/AOG.0000000000000584
17. Yijie G, Mengdan S, Min L, et al. MiR-95-3p/EPM2A/MMP2 contributes to the pathogenesis of severe preeclampsia through the regulation of trophoblast biological behaviour. Arch Biochem Biophys. 2023;741:109596.
18. Michela C, Francesca P, Patrizia G, et al. MiR-223 Exclusively Impairs In Vitro Tumor Growth through IGF1R Modulation in Rhabdomyosarcoma of Adolescents and Young Adults. Int J Mol Sci. 2022;23(22):46.
19. Zhijuan X, Jun T, Zhong C, Lanji W, Jianying C, Qin L. Human bone marrow mesenchymal stem cell-derived extracellular vesicles reduce inflammation and pyroptosis in acute kidney injury via miR-223-3p/HDAC2/SNRK. Inflammation Res. 2023;72(3):847.
20. Lin L, Pengwei L, Chiyi H, Chunfang X. miRNA-223-3p regulates ECT2 to promote proliferation, invasion, and metastasis of gastric cancer through the Wnt/β-catenin signaling pathway. J Cancer Res Clin Oncol. 2022;149(1):5986.
21. Favero A, Segatto I, Perin T, Belletti B. The many facets of miR‐223 in cancer: oncosuppressor, oncogenic driver, therapeutic target, and biomarker of response. Wiley Interdiscip Rev RNA. 2021;12(6):e1659. doi:10.1002/wrna.1659
22. Wei Z, Wang Y, Jiang L, et al. miR-223 regulates oral squamous cell carcinoma metastasis through the Wnt/β-catenin signaling pathway. Oral Oncol. 2020;109:104941. doi:10.1016/j.oraloncology.2020.104941
23. Dong YW, Wang R, Cai QQ, et al. Sulfatide epigenetically regulates miR-223 and promotes the migration of human hepatocellular carcinoma cells. J Hepatol. 2014;60(4):792–801. doi:10.1016/j.jhep.2013.12.004
24. Jeffries J, Zhou W, Hsu AY, Deng Q. miRNA-223 at the crossroads of inflammation and cancer. Cancer Lett. 2019;451:136–141. doi:10.1016/j.canlet.2019.02.051
25. Li Q, Jiang B, Qi Y, Zhang H, Ma H. Long non-coding RNA SLCO4A1-AS1 drives the progression of non-small-cell lung cancer by modulating miR-223-3p/IKKα/NF-κB signaling. Cancer Biol Ther. 2020;21(9):806–814. doi:10.1080/15384047.2020.1787757
26. Geng S, Tu S, Fu W, Wang J, Bai Z. LncRNA PITPNA-AS1 stimulates cell proliferation and suppresses cell apoptosis in glioblastoma via targeting miR-223-3p/EGFR axis and activating PI3K/AKT signaling pathway. Cell Cycle. 2021;20(19):1988–1998. doi:10.1080/15384101.2021.1958503
27. Rose SA, Wroblewska A, Dhainaut M, et al. A microRNA expression and regulatory element activity atlas of the mouse immune system. Nat Immunol. 2021;22(7):914–927. doi:10.1038/s41590-021-00944-y
28. Yuan S, Chen Y, Zhang M, et al. Overexpression of miR-223 Promotes Tolerogenic Properties of Dendritic Cells Involved in Heart Transplantation Tolerance by Targeting Irak1. Front Immunol. 2021;12:676337.
29. Roffel MP, Boudewijn IM, van Nijnatten JL, et al. Identification of asthma-associated microRNAs in bronchial biopsies. Eur Respir J. 2022;59(3):2101294. doi:10.1183/13993003.01294-2021
30. Haacke EM, Ge Y, Sethi SK, Buch S, Zamboni P. An overview of venous abnormalities related to the development of lesions in multiple sclerosis. Front Neurol. 2021;589.
31. Morquette B, Juźwik CA, Drake SS, et al. MicroRNA-223 protects neurons from degeneration in experimental autoimmune encephalomyelitis. Brain. 2019;142(10):2979–2995. doi:10.1093/brain/awz245
32. Yang J, Kuang H, Li N, Hamdy AM, Song J. The modulation and mechanism of probiotic-derived polysaccharide capsules on the immune response in allergic diseases. Crit Rev Food Sci Nutr. 2022;1–13.
33. Mázló A, Jenei V, Burai S, Molnár T, Bácsi A, Koncz G. Types of necroinflammation, the effect of cell death modalities on sterile inflammation. Cell Death Dis. 2022;13(5):1–12. doi:10.1038/s41419-022-04883-w
34. Specjalski K, Jassem E. MicroRNAs: potential biomarkers and targets of therapy in allergic diseases? Arch Immunol Ther Exp (Warsz). 2019;67(4):213–223. doi:10.1007/s00005-019-00547-4
35. Snyder BL, Blackshear PJ. Clinical implications of tristetraprolin (TTP) modulation in the treatment of inflammatory diseases. Pharmacol Ther. 2022;239:108198. doi:10.1016/j.pharmthera.2022.108198
36. Chen Q, Li H, Liu Y, Zhao M. Epigenetic regulation of immune and inflammatory responses in rheumatoid arthritis. Front Immunol. 2022;2:1700.
37. Zheng X, Zhao F-C, Pang Y, et al. Downregulation of miR-221-3p contributes to IL-1β-induced cartilage degradation by directly targeting the SDF1/CXCR4 signaling pathway. J Mol Med. 2017;95(6):615–627. doi:10.1007/s00109-017-1516-6
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.