Back to Journals » Breast Cancer: Targets and Therapy » Volume 15
Identification of GZMA as a Potential Therapeutic Target Involved in Immune Infiltration in Breast Cancer by Integrated Bioinformatical Analysis
Authors Huo Q ![]() , Ning L
, Ning L ![]() , Xie N
, Xie N ![]()
Received 8 December 2022
Accepted for publication 18 February 2023
Published 10 March 2023 Volume 2023:15 Pages 213—226
DOI https://doi.org/10.2147/BCTT.S400808
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Pranela Rameshwar
Qin Huo, Lvwen Ning, Ni Xie
Biobank, Shenzhen Institute of Translational Medicine, Shenzhen Second People’s Hospital, First Affiliated Hospital of Shenzhen University, Health Science Center, Shenzhen University, Shenzhen, People’s Republic of China
Correspondence: Ni Xie, Biobank, Shenzhen Institute of Translational Medicine, Shenzhen Second People’s Hospital, First Affiliated Hospital of Shenzhen University, Health Science Center, Shenzhen University, No. 3002 Sungang West Road, Futian District, Shenzhen, 518035, People’s Republic of China, Tel +86 1 350 158 0802, Fax +86 7 558 300 3435, Email [email protected]
Purpose: Granzyme A (GZMA) is a potential prognostic target for various cancer types. However, its therapeutic significance in breast cancer with immune infiltration remains controversial. We analyzed GZMA expression and its prognostic value in breast cancer with immune cell infiltration.
Patients and methods: Data was obtained from patients with breast cancer registered at The Cancer Genome Atlas. A correlation was performed between GZMA expression and patient’s clinicopathological features such as age, pathologic stage, metastasis stage, overall survival (OS), disease-specific survival (DSS), and progress free interval (PFI). Kaplan-Meier analyses and Cox proportional hazard regression model were used to examine the predictive significance of GZMA expression for breast cancer. The co-expression pattern of GZMA was assessed by the LinkedOmics web portal. The relationship between GZMA expression and immune cells was analyzed using the TIMER database. The correlation between GZMA and lymphocytes and immunomodulators was established with the TISIDB database.
Results: There was a lower GZMA expression in breast cancer tissue than in normal tissue. Interestingly, GZMA expression was associated with age, pathologic stage, and the Tumour, Node, and Metastasis stage. Overexpression of GZMA was also associated with better OS, DSS, and PFI. Based on the Cox regression analysis, GZMA was identified as an independent favorable prognostic factor for breast cancer. Our findings demonstrated a strong association between GZMA and T-cell checkpoints (PD-1, PD-L1, and cytotoxic T lymphocyte-associated antigen (CTLA-4)) in breast cancer. Moreover, we evaluated the interactions between GZMA expression and markers of dendritic and CD8+ T cells using quantitative immunofluorescence. We discovered that increased infiltration of dendritic and CD8+ T cells was associated with GZMA expression in breast cancer.
Conclusion: GZMA expression is associated with a favorable prognosis in breast cancer and is significantly correlated with immune cell infiltration. GZMA may be considered a promising therapeutic target for patients with breast cancer.
Keywords: GZMA, expression, prognostic factor, immune infiltration, breast cancer
Graphical Abstract:

Introduction
Breast cancer is considered one of the most prevalent malignant tumors.1,2 It has surpassed lung cancer as the most prevalent cancer type worldwide in 2020, with an estimated 2.26 million new cases. Despite advances in research and treatment methods, the incidence of breast cancer continues to rise.3,4 As a result, finding new prognostic and diagnostic biomarkers for breast cancer is critical.
Breast cancer immunotherapy has made significant progress in recent years. Despite the fact that monoclonal antibodies against programmed cell death protein 1 (PD-1) and programmed cell death-ligand 1 (PD-L1) had achieved a breakthrough in immunotherapy, only 20% of patients benefited from the treatment.5 It is known that carcinogenesis is significantly influenced by the degree of immune cell infiltration in the tumor microenvironment (TME).6–8 Consistently, a significant predictive factor for patient survival in breast cancer is the number of tumor-infiltrating lymphocytes (TILs).9–11 T-lymphocytes constitute 75% of TME immune cells.12 CD8+ T cells are the main cell group with specific killing function, and their number determines the prognosis of patients with cancer. Activated CD8+ T cells can rapidly release high concentrations of cytokines such as interferon-gamma (IFN-γ) and interleukin (IL)-2, which play an essential function in anti-tumor immunity.13,14 Thus far, no effective markers have been found for breast cancer that can effectively stimulate antigen-specific CD8+ T-cell responses and be used in combination with PD-1/PD-L1 immunotherapy.15 Hence, there is a pressing need to find markers that can induce CD8+ T-cell response to stimulate an autoimmune response.
Granzyme A (GZMA) is a member of the granzyme family of serine proteases, and is exclusively present in the granules of cytotoxic T lymphocytes (CTL) and natural killer (NK) cells.16 Recent studies have shown that GZMA acts as a key effector molecule of regulatory T-cell function in cancer. It is also an important molecular target for inhibiting tumor growth, inducing apoptosis and antigen-specific cytotoxic CD8+ T-lymphocytes, and protecting mice from tumor challenges.17 Santiago L. reported that GZMA promoted colorectal cancer by enhancing intestinal inflammation.18 Recently, a study identified that GZMA protein derived from cytotoxic lymphocytes can specifically activate a gasdermin-B protein in the process of killing target cells by cytotoxic lymphocytes. This molecular mechanism promotes an anti-tumor immune response.19 Therefore, GZMA performs an essential position in the anti-tumor immune method and affords a new goal for most cancer immunotherapy.
This study aimed to analyze the expression of GZMA and further study its prognostic value in breast cancer with immune cell infiltration. We evaluated whether GZMA can be used as a therapeutic target for breast cancer prognosis assessment, and explored the possibility of using GZMA as a target to trigger anti-tumor immunity in patients with breast cancer.
Materials and Methods
GZMA Expression Level and Its Correlation with Clinicopathological Features
Data from 1222 patients from The Cancer Genome Atlas (TCGA; https://portal.gdc.cancer.gov/) were downloaded for the clinical trial and the RNA-seq study. Correlations between GZMA expression levels and various classification parameters were assessed. The parameters included age (≤60; >60), pathologic stage (stages I, II, III, and IV), radiation therapy, tumor, node, metastasis (TNM) stage (T: T1, 2, 3, 4; N: N0, 1, 2, 3; M: M0, 1), overall survival (OS), disease-specific survival (DSS), and progress free interval (PFI) of patients with breast cancer who were investigated. A value of p < 0.05 was considered statistically significant.
Kaplan–Meier Survival Curve Analysis
We assessed the prognostic significance of GZMA mRNA expression in breast cancer using the Kaplan-Meier survival curve. Using this data, we calculated hazard ratios (HR), 95% confidence intervals (CI), and log-rank p values. Analyses of univariate and multivariate Cox regression identified independent prognostic factors.
Linked Omics Database Analysis
The LinkedOmics database (http://www.linkedomics.org/login.php) is an online website for exploring the gene expression profile.20 We identified the differentially expressed genes related to GZMA through the LinkFinder module. Next, the Gene Ontology biological process (GO_BP) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway were evaluated using the gene set enrichment analysis.
GZMA and Immune Cell Infiltration
The Tumour Immune Estimation Resource (TIMER) database (https://cistrome.shinyapps.io/timer/) is a comprehensive resource for the systematic analysis of immune infiltrates across diverse cancer types.21 First, the correlation between GZMA expression and the level of different immune cells was evaluated by the Spearman correlation test. Next, the association between GZMA and lymphocytes, immunomodulators, and chemokines was assessed using the Tumour and Immune System Interaction Database (TISIDB) (http://cis.hku.hk/TISIDB).22
Immunohistochemistry and Multiplex Quantitative Immunofluorescence
The immunohistochemical assay was conducted on human breast cancer tissue and adjacent tissue. Tissue samples were collected from Shenzhen Second People’s Hospital, China. Ethics Committee approval of the study was obtained from Shenzhen Second People’s Hospital. All the above tissue samples were cryopreserved and routinely processed into frozen sections. Briefly, after deparaffinization and rehydration, Antigens were extracted from sections by heating them at 95°C for 15 min in 10 mM citrate buffer (pH 6.0). After treatment with 3% H2O2 for 10 min. The samples were incubated overnight at 4°C with primary antibodies targeting GZMA at a 1:250 dilution, followed by a horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G at room temperature for 30 minutes. Next, diaminobenzidine (DAB) (Sigma, USA) was used to stain the slides for 10 min to label GZMA proteins. The slides were then counterstained with hematoxylin to label nuclei. Using visual grading, the intensity of staining was used to score GZMA expression levels. The 10 tissue samples of patients with breast cancer were dyed using a protein multiplex immunohistochemistry test kit, and the images were obtained by Vectra 3 pathological imaging system microscope (Perkin Elmer). Antibodies used in this study were anti-GZMA (1:100, Affinity Biosciences), anti-CD8A (1:100, Affinity Biosciences), and HLA-DPB1 (1:100, Affinity Biosciences). Nuclear counterstaining was performed with DAPI (4, “prime”, 6,6-diamidino-2-phenylindole).
Statistical Analysis
Most analyses were conducted by using R statistical software. Clinical characteristics and GZMA expression were analyzed with the Wilcoxon rank sum test. For assessing the association between GZMA and clinicopathological variables, both univariate and multivariate Cox regression analyses were performed. A p-value of <0.05 was considered statistically significant.
Results
GZMA and Clinical Parameters in patients with Breast Cancer
GZMA protein expression was analyzed between the tumor and adjacent normal tissue using immunohistochemistry. Compared to adjacent normal breast tissue, we observed lower expression of GZMA in breast cancer tissue (Figure 1). To further evaluate the function of GZMA in breast cancer, a correlation was performed between GZMA expression and clinicopathological features such as age, pathologic stage, radiation therapy, TNM stage, OS, DSS, and PFI (Figure 2). For age, patients younger than 60 years displayed upregulation of GZMA expression compared with older than 60 years. Specifically, GZMA showed a higher expression in stages II and III than in normal breast tissues (p < 0.05). In addition, the expression of GZMA in the radiation therapy group was higher than in normal breast tissues (p < 0.001). The expression of GZMA in tumor classified as T2, N0, N2, and M0 was higher than in normal tissue (p < 0.05). Interestingly, it was observed that deceased patients who had breast cancer had lower GZMA expression than living patients (OS: p < 0.01; DSS: p < 0.05; DFI: p < 0.01). Moreover, the correlation between GZMA expression and clinicopathological characteristics was established with logistical analysis (Table 1). We found a strong correlation between GZMA expression and both estrogen receptor (ER) status and OS.
 |
Table 1 Clinicopathological Characteristics of Breast Cancer Patients with Differential GZMA Expression |
 |
Figure 1 Analysis of GZMA protein expression in tumor and adjacent normal tissue using immunohistochemistry. |
 |
Figure 2 Box plots evaluating GZMA expression among different groups of patients based on clinicopathologic parameters. (A) Age (≤ 60; > 60); (B) Pathologic stage (stage I, stage II, stage III, and stage IV); (C) Radiation therapy (Yes; No); (D–F) Tumor, node, metastasis (TNM) stage (T: T1, T2, T3, T4; N: N0, N1, N2, N3; M: M0, M1); (G) Overall survival (OS); (H) Disease-specific survival (DSS); and (I) Progress free interval (PFI). *p < 0.05; **p < 0.01; ***p < 0.001. Abbreviation: ns, no significance. |
Prognostic Potential of GZMA in patients with Breast Cancer
Next, we used Kaplan-Meier analyses to examine the predictive significance of GZMA expression in breast cancer. High expression of GZMA exhibited good OS (HR= 0.61, p=0.003), DSS (HR= 0.62, p = 0.033), and PFI (HR=0.69, p=0.026) (Figure 3A–C). The link between GZMA expression and clinicopathological characteristics was also examined using the Cox proportional hazard regression model. In patients with breast cancer, we discovered a strong correlation between high GZMA expression and the following factors: age (p < 0.001), N1, N2 stage (p < 0.001), M1 stage (p < 0.001), pathologic stage III and IV (p < 0.001), and radiation therapy (p = 0.004) (Figure 3D). To further assess the GZMA’s ability to predict clinical outcomes, we also ran a multivariate Cox regression analysis. GZMA expression in age, M1, stage III, and radiation therapy was independently associated with OS in patients with breast cancer (all p<0.05). As presented in Figure 3E, we show a correlation between GZMA expression and clinicopathological parameters with DSS in these patients. We found a strong correlation between high GZMA expression and T2&T3&T4 (p = 0.038), N1&N2&N3 (p < 0.001), M1 stage (p < 0.001), and pathologic stage II & stage III& stage IV (p = 0.004). A multivariate Cox regression analysis showed GZMA was a reliable prognostic factor for DSS (p = 0.046). Next, the analysis of univariate and multivariate Cox regression was performed to determine whether GZMA expression was an independent predictor for PFI (Figure 3F). The clinical N and M stages exhibited predictive advantages for clinical outcomes. GZMA was a significant good prognostic factor for DFI (p = 0.037).
 |
Figure 3 Correlation between GZMA expression and prognosis in breast cancer. (A) Overall survival (OS). (B) Disease-specific survival (DSS). (C) Progress free interval (PFI). Forest plot of univariate and multivariate Cox regression analysis for OS (D), DSS (E), and PFI (F). |
The calibration curve constructed the prognosis nomogram using these important prognostic parameters (Figure 4). The nomogram to predict OS, with a C-index of 0.731, includes age, M stage, pathologic stage, radiation_therapy as well as GZMA. A nomogram was created to predict DSS (C-index = 0.683) and PFI (C-index = 0.598) using the clinical N and M stages, and GZMA expression. The three nomograms of 1, 3, and 5-year clinical findings were all perfectly predicted by the calibration curves. However, OS was slightly overestimated in the one-year prognosis. The above results suggest the prognostic potential of GZMA for patients with breast cancer.
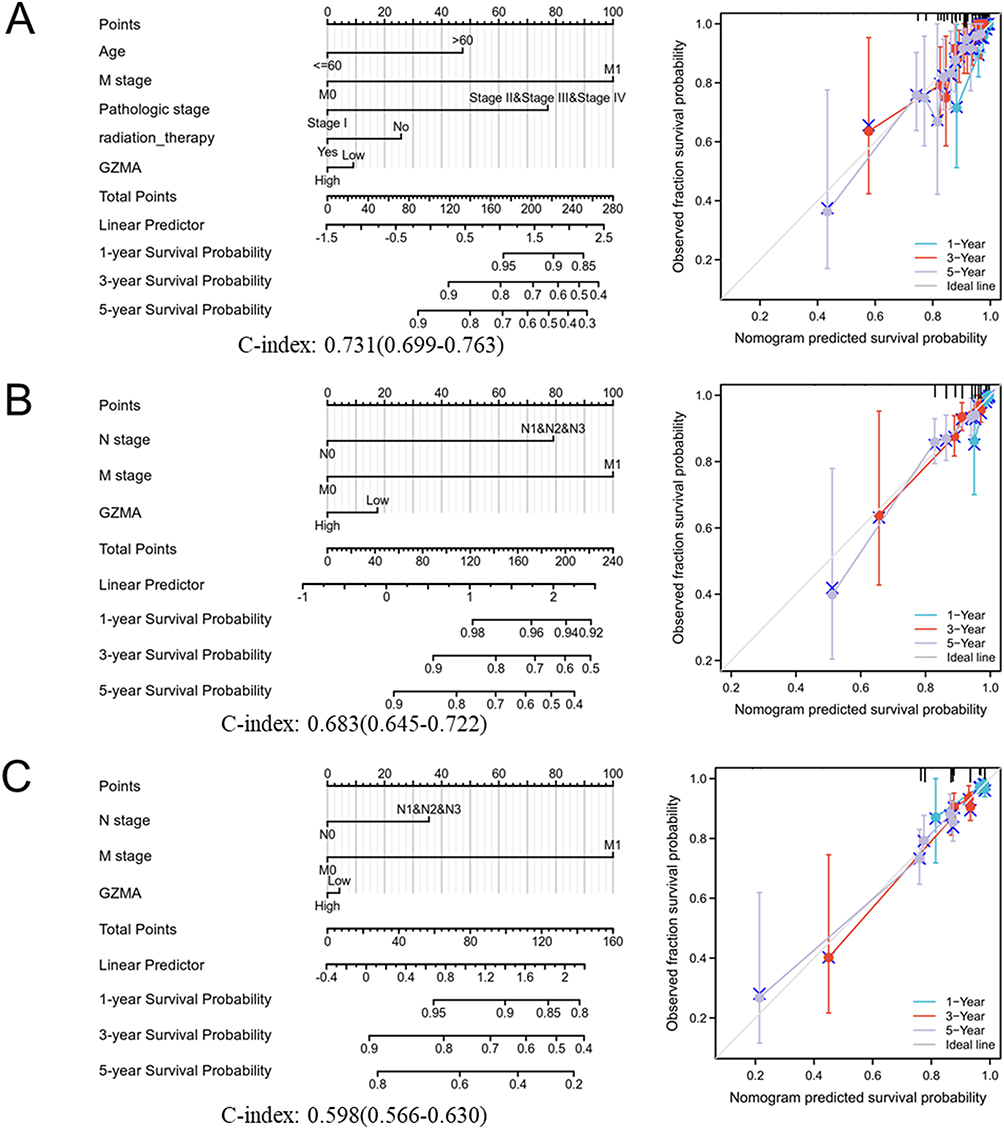 |
Figure 4 Construction and validation of nomograms based on GZMA expression. (A) Overall survival (OS). (B) Disease-specific survival (DSS). (C) Progress free interval (PFI). |
Analysis of Genes Co-Expressed with GZMA in Breast Cancer
To investigate the functions of GZMA, the co-expression pattern of GZMA was assessed with the LinkedOmics web portal. The top 50 genes related to GZMA, both positively and negatively, were examined (Figure 5). Six genes significantly correlated with GZMA, including the zeta chain of T-cell receptor-associated protein kinase 70 (ZAP70) (r = 0.728, p = 8.895e-18), myosin IG (MYO1G) (r = 0.718, p = 4.04e-17), apolipoprotein B mRNA editing enzyme, catalytic subunit 3G (APOBEC3G) (r =0.717, p = 4.63e-17), septin 1 (SEPT1) (r = 0.716, p = 5.87e-17), dedicator of cytokinesis 2 (DOCK2) (r = 0.712, p = 9.97e-17), and pleckstrin (PLEK) (r = 0.712, p =1.07e-16). The genes co-expressed with GZMA, including ZAP70, MYO1G, APOBEC3G, SEPT1, DOCK2, and PLEK had markedly better OS in patients with breast cancer (Supplementary Figure 1). Therefore, we speculate that GZMA and its co-expressed genes contribute to the higher OS in patients with breast cancer.
 |
Figure 5 Analysis of genes co-expressed with GZMA in breast cancer. (A) Co-expression genes with GZMA in breast cancer. Top 50 genes positively and negatively associated with GZMA in breast cancer are shown. (B) The six most significant genes correlated with GZMA. |
The annotation of GO_BP terms revealed that the co-expressed genes of GZMA had primary roles in phagocytosis, response to interferon-gamma, adaptive immune response, immune response-regulating signaling pathways, leukocyte cell-cell adhesion, positive regulation of cell activation, leukocyte proliferation, T cell activation, lymphocyte-mediated immunity, and cell killing (Figure 6A). Analysis of KEGG pathway revealed enrichment in NK cell-mediated cytotoxicity, Fc gamma R-mediated phagocytosis, chemokine signaling pathway, tuberculosis, antigen processing and presentation, leishmaniasis, staphylococcus aureus infection, osteoclast differentiation, B-cell receptor signaling pathway, and Epstein-Barr virus infection (Figure 6B).
 |
Figure 6 The Gene Ontology biological process (GO_BP) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway of GZMA in breast cancer. (A) The annotation of GO_BP terms revealed the co-expressed genes of GZMA. (B) Analysis of KEGG pathway of GZMA in breast cancer. |
Correlation Between GZMA Expression and Infiltrating Immune Cells
Using the TIMER database, we analyzed the relationship between GZMA expression and immune cells. Figure 7A demonstrates a positive correlation between GZMA expression and B-cell levels (r = 0.492, p = 7.99e-61), CD4+ T cells (r = 0.593, p = 2.59e-92), and neutrophils (r = 0.591, p = 1.56e-90). Significantly positive correlations between GZMA expression and dendritic cells (r = 0.675, p = 3.17e-127) and CD8+ T cells (r = 0.634, p = 5.55e-111) were found. GZMA was positively correlated with dendritic cell markers (HLA-DPB1: r = 0.780, p <0.001); HLA-DQB1 (r = 0.680, p < 0.001) and CD8+ T-cell markers (CD8B: r =0.830, p < 0.001; CD8A: r = 0.910, p < 0.001). More specifically, our findings demonstrated a strong association between GZMA and T-cell checkpoints (PD-1, PD-L1, and cytotoxic T lymphocyte-associated antigen (CTLA-4)) in breast cancer (Figure 7B and C). Further, we assessed the association between genes co-expressed with GZMA and infiltrating immune cells. Significant correlations between immune cells and the GZMA-related genes ZAP70, MYO1G, APOBEC3G, SEPT1, DOCK2, and PLEK have been found (Supplementary Figure 2). ZAP70 expression was positively correlated with dendritic cells (r = 0.596, p = 1.78e-92), CD4+ T cells (r =0.692, p = 3.30e-138), and neutrophils (r = 0.504, p = 2.30e-62). MYO1G expression was associated with dendritic cells (r = 0.621, p = 1.37e-102), CD4+ T cells (r = 0.562, p = 3.33e-81), and neutrophils (r = 0.535, p =1.22e-71). APOBEC3G expression were substantially correlated with dendritic cells (r = 0.618, p = 2.60e-101), neutrophils (r = 0.598 p =2.22e-93), CD8+ T cells (r =0.56, p = 9.88e-82). SEPT1 expression showed a strong correlation with CD4+ T cells (r =0.337, p = 5.24e-27). Furthermore, DOCK2 was associated with dendritic cells (r = 0.872, p = 1.02e-296), neutrophils (r = 0.838, p=1.63e-251), CD4+ T cells (r = 0.717, p = 1.46e-152), and CD8+ T cells (r = 0.602, p = 1.89e-97). Also, we observed that PLEK was positively correlated with dendritic cells (r = 0.887, p =6.32e-321), neutrophils (r = 0.856, p = 2.94e-274), CD4+ T cells (r = 0.669, p = 4.47e-126), and CD8+ T cells (r = 0.617, p =2.74e-103).
 |
Figure 7 Correlation of GZMA expression and immune infiltration levels in breast cancer. (A) Correlation analysis of GZMA expression and infiltration levels of immune cells (B-cell abundance, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells) using the TIMER database. (B) GZMA expression was significantly correlated with dendritic and CD8+ T cells using TCGA data. (C) The expression of GZMA was positively correlated with dendritic cells markers (HLA-DPB1 and HLA-DQB1) and CD8+ T-cell markers (CD8A and CD8B). PDL1 (CD274); PD1(PDCD1). |
Correlation Between GZMA and Immune Molecules
The correlation between GZMA and lymphocytes and immunomodulators was established with the TISIDB database. The strongest associations were found between GZMA expression and Act_CD8 (r = 0.865, p<2.2e-16), Tem_CD8 (r = 0.825, p<2.2e-16), Act_B (r = 0.797, p<2.2e-16), and Imm_B (r = 0.796, p<2.2e-16) (Figure 8A). Immunomodulators included immune inhibitors, immune stimulators, and major histocompatibility complex (MHC) molecules.23 As shown in Figure 8B, among these immune inhibitors, we found that GZMA was positively associated with CD96 (r = 0.904, p<2.2e-16), T-cell immunoreceptor with Ig and ITIM domains (TIGIT) (r =0.863, p<2.2e-16), PDCD1 (r = 0.853, p<2.2e-16), and B- and T-lymphocyte attenuator (BTLA) (r = 0.844, p<2.2e-16). The immune stimulators displaying the greatest correlation included killer cell lectin-like receptor K1 (KLRK1) (r =0.904, p<2.2e-16), CD48 (r =0.894, p<2.2e-16), CD27 (r =0.878, p<2.2e-16), and CD40 ligand (CD40LG) (r =0.839, p<2.2e-16) (Figure 8C). In addition, MHC molecules and GZMA expression were discovered to be positively linked. (Figure 8D), including HLA-DRA (r =0.775, p<2.2e-16), HLA-DPB1 (r = 0.768, p<2.2e-16), HLA-DPA1 (r =0.754, p<2.2e-16), and HLA-DMB (r = 0.752, p<2.2e-16).
 |
Figure 8 Correlation of GZMA expression and lymphocytes, immunomodulators and chemokines in breast cancer. (A) Correlation between abundance of TILs and GZMA (plus the four TILs with the highest correlation, respectively). (B) Correlation between the level of immune inhibitors and GZMA expression (plus the four immune inhibitors with the highest correlation, respectively). (C) Association between immune stimulators and GZMA expression (plus the four immune stimulators with the highest correlation, respectively). (D) Correlation between the abundance of major histocompatibility complex (MHC) molecules and GZMA expression (plus the four MHC molecules with the highest correlation, respectively). |
Immune Infiltration Levels of Dendritic and CD8+ T Cells in Breast Cancer
Based on the aforementioned findings, we made the assumption that GZMA expression was positively correlated with the level of infiltration of dendritic and CD8+ T cells. To test this hypothesis, we evaluated the interactions between GZMA expression and markers of dendritic and CD8+ T cells using quantitative immunofluorescence. We discovered that breast cancer tissue with high GZMA expression had a high level of dendritic and CD8+T cells (Figure 9A). Among breast cancer tissue expressing relatively little GZMA, there are few dendritic and CD8+T cells (Figure 9B).
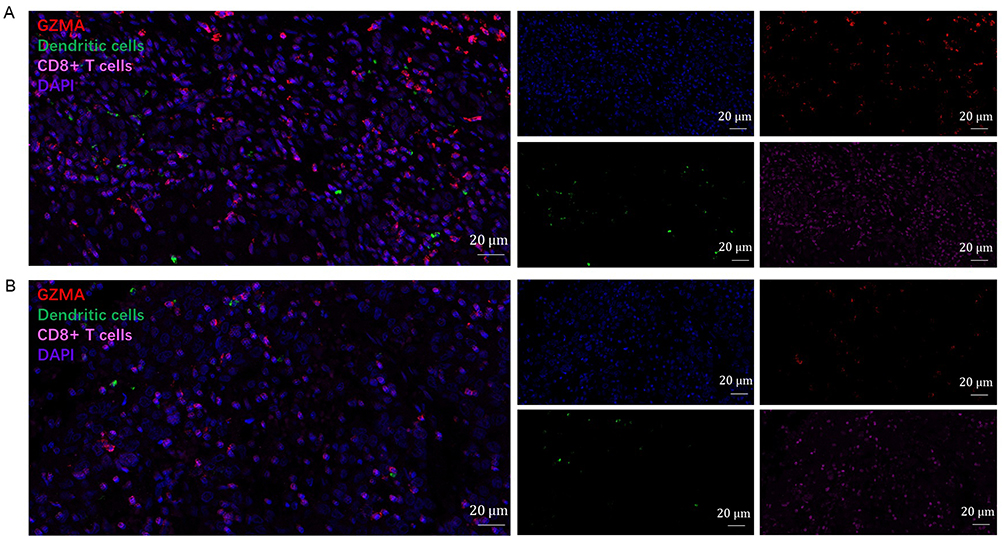 |
Figure 9 Detection of immune infiltration levels of dendritic and CD8+ T cells in breast cancer using multiplex quantitative immunofluorescence. (A) Relatively high GZMA expression in breast cancer tissue. (B) Relatively low GZMA expression in breast cancer tissue. Representative fluorescence images showing the detection of immune cell samples by simultaneous staining of DAPI, GZMA (red channel), CD8A (pink channel), and HLA-DPB1 (green channel) in breast cancer. CD8+ T-cell marker: CD8A; Dendritic cells marker: HLA-DPB1. |
Discussion
The goal of this study was to use the TCGA and a comprehensive bioinformatics data mining technique to assess the predictive significance of the degree of immune infiltration of GZMA in breast cancer. The present investigation is the first to fully elucidate the prognostic significance and expose the relationship between GZMA and immune infiltration in breast cancer.
GZMA is produced by a variety of cytotoxic lymphocytes, such as NK cells,24 CD8 cytotoxic T-lymphocytes,16 and CD4 CTL.25 Circulating GZMA protein levels are elevated in various infectious diseases.26,27 It has been demonstrated that only GZMA with its proteolytic activity is sufficient to induce inflammation.28 To verify the role of GZMA in cells, the NK cell-killing experimental system and a CTL cell-killing experimental system were used. Knocking down GZMA through siRNA revealed the important role of the GZMA-GSDMB pathway in the killing of target cells by cytotoxic lymphocytes.19
This study focused on the expression and clinical characteristics of GZMA to determine its potential prognostic value in breast cancer. We observed a marked association of GZMA expression with patients’ age, pathologic stage, radiation therapy, TNM stage, OS, DSS, and PFI. Interestingly, the expression of GZMA was higher in younger patients and significantly more associated with the early stage of TNM. By comparing patients with breast cancer who survived and died, we found that GZMA was expressed at a higher level in living patients with breast cancer than in deceased ones. To determine the predictive value of GZMA, we used Kaplan-Meier analysis and discovered that increased GZMA expression was related to a better prognosis in terms of OS, DSS, and PFI. According to univariate and multivariate Cox regression analysis, GZMA may be an independent prognostic marker for patients with breast cancer. Taken together with the results from the study, GZMA was strongly confirmed as a predictive biomarker in breast cancer.
We also investigated GZMA co-expression patterns utilizing the LinkedOmics online platform. The results indicated that GZMA was strongly correlated with ZAP70, MYO1G, APOBEC3G, SEPT1, DOCK2, and PLEK. ZAP70 is necessary for T-cell activation.29 Deletion of MYO1G leads to a reduction in cell rigidity and affects cell adhesion, diffusion, phagocytosis, and endocytosis of B-lymphocytes.30 Studies have shown that APOBEC3G regulates signal transducer and activator of transcription 3 (STAT3) and nuclear factor kappa B (NF-κB), chemokine ligand 5 (CCL5), IL-6, IL-4, IFN-γ, the intrinsic ability of IL-10, and IL-17 genes. APOBEC3G can also induce T-cell plasticity and regulate the immune response.31 Consistently our analysis further pointed out that these proteins were primarily engaged in immune response or tumorigenesis. GO and KEGG pathway analysis were conducted to determine the protein functions. Interestingly, we found that GZMA is primarily expressed in phagocytosis, response to IFN-γ, adaptive immune response, and immune response-regulating signaling pathways. Therefore, GZMA promotes breast cancer development by triggering an immune response.
An earlier investigation showed that TME immune infiltration is related to survival in many patients with cancer.9,32,33 Additionally, the degree of immune infiltration was directly associated with the prognosis of breast cancer.34,35 According to our research, there is a strong correlation between the level of immune infiltration and GZMA expression in breast cancer. Dendritic and CD8+ T cells were positively associated with GZMA expression and in turn with improved patient prognosis. Moreover, GZMA expression was also substantially correlated with T-cell checkpoints (PD-1, PD-L1, and CTLA-4) in breast cancer. Furthermore, the relationship between GZMA expression and immune inhibitors, immune stimulators, and MHC molecules was assessed. Notably, GZMA expression was discovered to be tightly connected to lymphocytes (such as Act_CD8, Tem_CD8, Act_B, and Imm_B), immune inhibitors (such as CD96, TIGIT, PDCD1, and BTLA), immune stimulators (such as KLRK1, CD48, CD27, and CD40LG), and MHC molecules (such as HLA-DRA, HLA-DPB1, HLA-DPA1, and HLA-DMB). TIGIT is an inhibitory receptor expressed on lymphocytes.36 Recently, TIGIT has drawn interest as a significant new target for cancer immunotherapy. According to earlier research, CD27 is a member of the tumor necrosis factor receptor superfamily and is essential for the activation of T cells via costimulatory signals.37 A prospective therapeutic target for invasive breast cancer is the PD-1/PD-L1 axis.38 Furthermore, significant positive relevance between GZMA expression and T-cell exhaustion marker (PDCD1) was discovered in breast cancer. On the other hand, we demonstrated GZMA had the strongest correlation with levels of CD8+ T cells (marker: CD8A) and dendritic cells (marker: HLA-DPB1) infiltration. The findings support our hypothesis, which reveal the role of GZMA in regulating tumor immunology. However, the connection between GZMA and immune infiltration in breast cancer has yet to be confirmed by additional research and clinical studies.
Since the results on bioinformatic analyses are limited and and require a large number of samples to reliably interpret the data, we aim to further verify the estimated results in cell and animal experiments to confirm the prognostic potential of GZMA for patients with breast cancer, together with the relationship between GZMA in breast cancer and immune cell infiltration.
Conclusions
In summary, this study shows that higher GZMA expression is independently associated with a better prognosis for breast cancer and correlates with it. Moreover, the underlying evidence indicates that GZMA is closely associated with immune cells. Our research indicates that GZMA may be crucial for immune cell infiltration and may function as a potential prognostic indicator for breast cancer.
Data Sharing Statement
We kindly thank the TCGA database (https://portal.gdc.cancer.gov/), TIMER (https://cistrome.shinyapps.io/timer/), the LinkedOmics database (http://www.linkedomics.org/login.php) and the TISIDB database (http://cis.hku.hk/TISIDB).
Ethics Approval and Informed Consent
This study was approved by the Ethics Committee of Shenzhen Second People’s Hospital in accordance with the principles of the Declaration of Helsinki. Informed consent from patients for their medical data to be used in the study was obtained.
Consent for Publication
All authors have seen and agreed to publish.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This project was supported by the National Natural Science Foundation of China, China (No. 82172356, No. 81972003), the Natural Science Foundation of Guangdong, China (No. 2021A1515012144, No 2020A1515111165).
Disclosure
The authors declare no competing interests in this work.
References
1. Chichiabellu TY, Mekonnen B, Astawesegn FH, Demissie BW, Anjulo AA. Essential newborn care practices and associated factors among home delivered mothers in Damot pulasa Woreda, southern Ethiopia. Reprod Health. 2018;15(1):162. doi:10.1186/s12978-018-0609-1
2. Huo Q, Li Z, Cheng L, Yang F, Xie N. SIRT7 is a prognostic biomarker associated with immune infiltration in luminal breast cancer. Front Oncol. 2020;10:621. doi:10.3389/fonc.2020.00621
3. Siegel RL, Miller KD, Jemal A. Breast cancer statistics, 2019. CA Cancer J Clin. 2019;69(1). doi:10.3322/caac.21583
4. Huo Q, Chen S, Li Z, Wang J, Li J, Xie N. Inhibiting of TACC3 promotes cell proliferation, cell invasion and the EMT pathway in breast cancer. Front Genet. 2021;12:640078. doi:10.3389/fgene.2021.640078
5. Brahmer JR, Tykodi SS, Chow LQ, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012;366(26):2455–2465. doi:10.1056/NEJMoa1200694
6. Kalinski P, Talmadge JE. Tumor immuno-environment in cancer progression and therapy. Adv Exp Med Biol. 2017;1036:1–18. doi:10.1007/978-3-319-67577-0_1
7. Wang M, Zhao J, Zhang L, et al. Role of tumor microenvironment in tumorigenesis. J Cancer. 2017;8(5):761–773. doi:10.7150/jca.17648
8. Buoncervello M, Gabriele L, Toschi E. The janus face of tumor microenvironment targeted by immunotherapy. Int J Mol Sci. 2019;20:17. doi:10.3390/ijms20174320
9. Baxevanis CN, Sofopoulos M, Fortis SP, Perez SA. The role of immune infiltrates as prognostic biomarkers in patients with breast cancer. Cancer Immunol Immunother. 2019;68(10):1671–1680. doi:10.1007/s00262-019-02327-7
10. Savas P, Virassamy B, Ye C, et al. Single-cell profiling of breast cancer T cells reveals a tissue-resident memory subset associated with improved prognosis. Nat Med. 2018;24(7):986–993. doi:10.1038/s41591-018-0078-7
11. Stanton SE, Disis ML. Clinical significance of tumor-infiltrating lymphocytes in breast cancer. J Immunother Cancer. 2016;4(1):59. doi:10.1186/s40425-016-0165-6
12. Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature. 2014;513(7517):202–209. doi:10.1038/nature13480
13. Mahmoud SM, Paish EC, Powe DG, et al. Tumor-infiltrating CD8+ lymphocytes predict clinical outcome in breast cancer. J Clin Oncol. 2011;29(15):1949–1955. doi:10.1200/JCO.2010.30.5037
14. Matsumoto H, Thike AA, Li H, et al. Increased CD4 and CD8-positive T cell infiltrate signifies good prognosis in a subset of triple-negative breast cancer. Breast Cancer Res Treat. 2016;156(2):237–247. doi:10.1007/s10549-016-3743-x
15. Li XD, Wu J, Gao D, et al. Pivotal roles of cGAS-cGAMP signaling in antiviral defense and immune adjuvant effects. Science. 2013;341(6152):1390–1394. doi:10.1126/science.1244040
16. Schanoski AS, Le TT, Kaiserman D, et al. Granzyme A in Chikungunya and other arboviral infections. Front Immunol. 2020;10:3083. doi:10.3389/fimmu.2019.03083
17. Shimizu K, Yamasaki S, Sakurai M, et al. Granzyme A stimulates pDCs to promote adaptive immunity via induction of type I IFN. Front Immunol. 2019;10:1450. doi:10.3389/fimmu.2019.01450
18. Santiago L, Castro M, Sanz-Pamplona R, et al. Extracellular granzyme A promotes colorectal cancer development by enhancing gut inflammation. Cell Rep. 2020;32(1):107847. doi:10.1016/j.celrep.2020.107847
19. Zhou Z, He H, Wang K, et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science. 2020;368(6494):eaaz7548. doi:10.1126/science.aaz7548
20. Vasaikar SV, Straub P, Wang J, Zhang B. LinkedOmics: analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018;46(D1):D956–D963. doi:10.1093/nar/gkx1090
21. Li B, Severson E, Pignon JC, et al. Comprehensive analyses of tumor immunity: implications for cancer immunotherapy. Genome Biol. 2016;17(1):174. doi:10.1186/s13059-016-1028-7
22. Ru B, Wong CN, Tong Y, et al. TISIDB: an integrated repository portal for tumor-immune system interactions. Bioinformatics. 2019;35(20):4200–4202. doi:10.1093/bioinformatics/btz210
23. Gou R, Zhu L, Zheng M, et al. Annexin A8 can serve as potential prognostic biomarker and therapeutic target for ovarian cancer: based on the comprehensive analysis of annexins. J Transl Med. 2019;17(1):275. doi:10.1186/s12967-019-2023-z
24. Wu CH, Li J, Li L, et al. Extracellular vesicles derived from natural killer cells use multiple cytotoxic proteins and killing mechanisms to target cancer cells. J Extracell Vesicles. 2019;8(1):1588538. doi:10.1080/20013078.2019.1588538
25. Della-Torre E, Bozzalla-Cassione E, Sciorati C, et al. A CD8α- subset of CD4+SLAMF7+ cytotoxic T cells is expanded in patients with IgG4-related disease and decreases following glucocorticoid treatment. Arthritis Rheumatol. 2018;70(7):1133–1143. doi:10.1002/art.40469
26. García-Laorden MI, Stroo I, Terpstra S, et al. Expression and function of Granzymes A and B in Escherichia coli peritonitis and sepsis. Mediators Inflamm. 2017;2017:4137563. doi:10.1155/2017/4137563
27. de Jong HK, Garcia-Laorden MI, Hoogendijk AJ, et al. Expression of intra- and extracellular granzymes in patients with typhoid fever. PLoS Negl Trop Dis. 2017;11(7):e0005823. doi:10.1371/journal.pntd.0005823
28. Stewart SE, D’Angelo ME, Bird PI. Intercellular communication via the endo-lysosomal system: translocation of granzymes through membrane barriers. Biochim Biophys Acta. 2012;1824(1):59–67. doi:10.1016/j.bbapap.2011.05.020
29. Moore JC, Mulligan TS, Yordán NT, et al. T cell immune deficiency in zap70 Mutant Zebrafish. Mol Cell Biol. 2016;36(23):2868–2876. doi:10.1128/MCB.00281-16
30. López-Ortega O, Ovalle-García E, Ortega-Blake I, et al. Myo1g is an active player in maintaining cell stiffness in B-lymphocytes. Cytoskeleton. 2016;73(5):258–268. doi:10.1002/cm.21299
31. Garg A, Kaul D. APOBEC3G has the ability to programme T cell plasticity. Blood Cells Mol Dis. 2016;59:108–112. doi:10.1016/j.bcmd.2016.04.013
32. Huo Q, Li Z, Chen S, Wang J, Li J, Xie N. VWCE as a potential biomarker associated with immune infiltrates in breast cancer. Cancer Cell Int. 2021;21(1):272. doi:10.1186/s12935-021-01955-3
33. Fridman WH, Pagès F, Sautès-Fridman C, Galon J. The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer. 2012;12(4):298–306. doi:10.1038/nrc3245
34. Oda K, Kato K, Nakamura M, et al. Surface marker profiles on lung lymphocytes may predict the mechanism of immune-mediated pneumonitis triggered by tumor infiltrating lymphocytes in lung cancer patients treated with pembroli zumab. Lung Cancer. 2018;118:171–172. doi:10.1016/j.lungcan.2018.02.012
35. Shimizu S, Hiratsuka H, Koike K, et al. Tumor-infiltrating CD8+ T cell density is an independent prognostic marker for oral squamous cell carcinoma. Cancer Med. 2019;8(1):80–93. doi:10.1002/cam4.1889
36. Harjunpää H, Guillerey C. TIGIT as an emerging immune checkpoint. Clin Exp Immunol. 2020;200(2):108–119. doi:10.1111/cei.13407
37. Starzer AM, Berghoff AS. New emerging targets in cancer immunotherapy: CD27 (TNFRSF7). ESMO Open. 2020;4(Suppl3):e000629. doi:10.1136/esmoopen-2019-000629
38. Fang J, Chen F, Liu D, Gu F, Chen Z, Wang Y. Prognostic value of immune checkpoint molecules in breast cancer. Biosci Rep. 2020;40(7):BSR20201054. doi:10.1042/BSR20201054
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.