")
Back to Journals » OncoTargets and Therapy » Volume 11
Identification of CD24 as a marker for tumorigenesis of melanoma
Authors Tang MR, Guo JY, Wang D, Xu N
Received 14 November 2017
Accepted for publication 9 April 2018
Published 12 June 2018 Volume 2018:11 Pages 3401—3406
DOI https://doi.org/10.2147/OTT.S157043
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Samir Farghaly
Ming-Rui Tang, Jia-Yan Guo, Di Wang, Nan Xu
Department of Plastic Surgery, The First Hospital of China Medical University, Shenyang 110001, People’s Republic of China
Objective: Cutaneous melanoma (CM) is a common skin cancer. Surgery is still the primary treatment for CM, as melanoma is resistant to chemotherapy. In the recent years, it has been found that cancer stem-like cells (CSCs) are responsible for this drug resistance. CD24 is a widely used marker to isolate CSCs. In this study, we aimed to analyze the properties of CD24+ and CD24− subpopulation of melanoma cells.
Materials and methods: We isolated CD24+ cells CSCs using magnetic-activated cell sorting system. We extracted total RNA and carried out reverse transcription polymerase chain reaction analysis. We counted the cell colonies using soft agar assay and assessed the cell invasion using cell migration assay. We implanted CD24+ or CD24− cells into the flank of non-obese diabetic severe combined immunodeficiency mice, and measured the tumor volumes every 5 days until the end of the experiment. We carried out immunohistochemical analysis to study the tissue sections.
Results: We demonstrated that the CD24+ subpopulation has self-renewal properties in vitro and in vivo by using soft agar assay and xenograft tumor model. Furthermore, we confirmed that CD24 expression is accompanied by activation of Notch1 signaling pathway.
Conclusion: This study provides new knowledge on the role of CD24 in the tumorigenic ability of melanoma.
Keywords: melanoma, CD24, apoptosis, migration, therapy
Introduction
Cutaneous melanoma (CM), a malignant tumor of the pigment-producing melanocytes, is the most lethal form of skin cancer.1 Surgery is still the primary treatment for CM.1 Several adjuvant therapies have been used for CM, such as radiation therapy and immunotherapy.2,3 However, the major obstacle to an effective treatment for melanoma is tumor heterogeneity.4
Melanoma contains multiple cell populations with various antigens.5 In order to elucidate the roles of the distinct cancer cell subpopulations in melanoma, we isolated two subpopulations based on CD24 expression. In our previous study, we have demonstrated that the levels of CD24 mRNA and protein were higher in cancer tissues than that in normal tissues of the melanoma patients.6 CD24, a 27-amino-acid single-chain protein that binds to the extracellular matrix, is a widely used cancer stem cell marker.7 CD24+ cancer cell colonies isolated from some cancers, such as nasopharyngeal carcinoma,8 ovarian cancer,9 and pancreatic cancer, showed increased self-renewal and enhanced chemoresistance.10 However, other studies showed that lack of CD24 expression appears to be critical for the identification of the breast cancer stem-like cells (CSCs).11,12
In this study, we demonstrated that CD24 was markedly associated with the properties of melanoma cells. CD24 expression in these cells is accompanied by activation of Notch1 signaling pathway. This study provides new knowledge on the role of CD24 in the tumorigenic ability of melanoma.
Materials and methods
Cell lines and culture
The melanoma cell lines, A375 and B16F10, were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). Cell lines were grown in Dulbecco’s Modified Eagle’s Medium (Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin) and maintained in a humidified incubator with 5% CO2 at 37°C.
Isolation of CD24+ cells from A375 and B16F10 cell lines
Magnetic-activated cell sorting (FACS Vantage SE; Becton Dickinson, Franklin Lakes, NJ, USA) was carried out using CD24 microbeads, according to the manufacturer’s protocol (STEMCELL Technologies, Vancouver, BC, Canada). Cells were labeled with anti-CD24 antibody cocktail and mixed with magnetic microbeads, and then separated by a magnet. Sorted CD24+ and CD24- cells were cultured on a six-well plate and used in the following studies.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was isolated from cells using the RNeasy Mini Kit (Biomed, Beijing, People’s Republic of China). RT-PCR was carried out as previously described.6
Soft agar assay
This was performed using the method of Lin et al.10 A six-well culture plate was coated with 2 mL of bottom agar mixture (0.6% agar). After the bottom layer solidified, 2 mL of top agar mixture (0.3% agar) containing 1 × 104 CD24+ or CD24- cells was added, and the plate was incubated for 4 weeks. The plate was stained with 0.5% crystal violet, and the colonies were counted using a light microscope (Olympus CX31; Olympus, Tokyo, Japan).
Cell migration assay
Cell invasion was assessed using 24-well transwell chambers with a pore size of 8 μm (Corning Incorporated, Corning, NY, USA). Cells (1 × 105) were seeded in the serum-free media in the upper well and incubated for 48 h. Cells in the lower chamber were stained with Giemsa staining solution and counted by a microscope.
Western blot analysis
Proteins from cells were resolved by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), transferred to a nitrocellulose membrane, and detected using the following antibodies: anti-Galectin-3 (1:200; sc-53127), anti-cleaved NICD1 (1:200; sc-32745), anti-JAG1 (1:100; sc-135955), anti-DLL4 (1:100; sc-18640), anti-Notch1 (1:200; sc-376403), anti-N-cadherin (1:200; sc-53488), and anti-E-cadherin (1:200; sc-71008) (Santa Cruz Biotechnology, Dallas, TX, USA). The bands were detected using an enhanced chemiluminescence system (Amersham Biosciences, Amersham, UK).
Xenograft tumor model
This study was approved by the Ethics Committee of China Medical University. The animal experiments were performed in accordance with the Care and Use of Laboratory Animals guideline of the Ethics Committee of China Medical University. CD24+ or CD24- cells (3 × 107) were subcutaneously implanted into the flank of non-obese diabetic severe combined immunodeficiency mice (4- to 6-weeks old; Charles River, Wilmington, MA, USA). Ten mice were in each group. Every five days until the end of the experiment, one mouse from each group was randomly selected and anesthetized, photographed, and sacrificed. Tumors were excised, and then prepared for immunohistochemical analyses. Tumors were measured using calipers, and tumor volumes were calculated (tumor volume = length × width2 × 0.52).
Immunohistochemistry
Tumor samples were formalin-fixed and embedded in paraffin. Paraffin sections were stained with the first antibody for CD24 (1:200; sc-7034) and Notch1 (1:200; sc-376403) (Santa Cruz Biotechnology) by incubating overnight at 4°C. Secondary staining with biotinylated secondary antibodies with horseradish peroxidase (HRP; Beyotime Biotechnology, Nanjing, People’s Republic of China) was performed for 30 min at room temperature. Then, the sections were counterstained with hematoxylin (Beyotime).
Statistical analysis
Each experiment was performed in triplicate. Statistical analysis was performed using one-tailed Student’s t-test (unilateral and unpaired). Survival rate was analyzed by the Kaplan–Meier method. Differences with a P-value less than 0.05 were considered statistically significant.
Results
Tumorigenic ability of CD24- and CD24+ subpopulation of A375 and B16F10 cells in vitro and in vivo
In this study, CD24+ cells were isolated from A375 and B16F10 cells by using a CSC-specific marker, CD24 (Figure 1). In addition, we found that CD24 mRNA was significantly higher in CD24+ cells than that in CD24- cells (Figure 1). The results of colony formation assays showed that CD24+ cells from both A375 and B16F10 cells could form more cell colonies than CD24- ones (P<0.05, Figure 2A). It indicated that the proliferation ratio of CD24+ cells is higher than CD24- ones. Migration of CD24- and CD24+ cells was detected using transwell assay. More CD24+ cells migrated to the lower membrane compared with CD24- ones (P<0.05, Figure 2B).
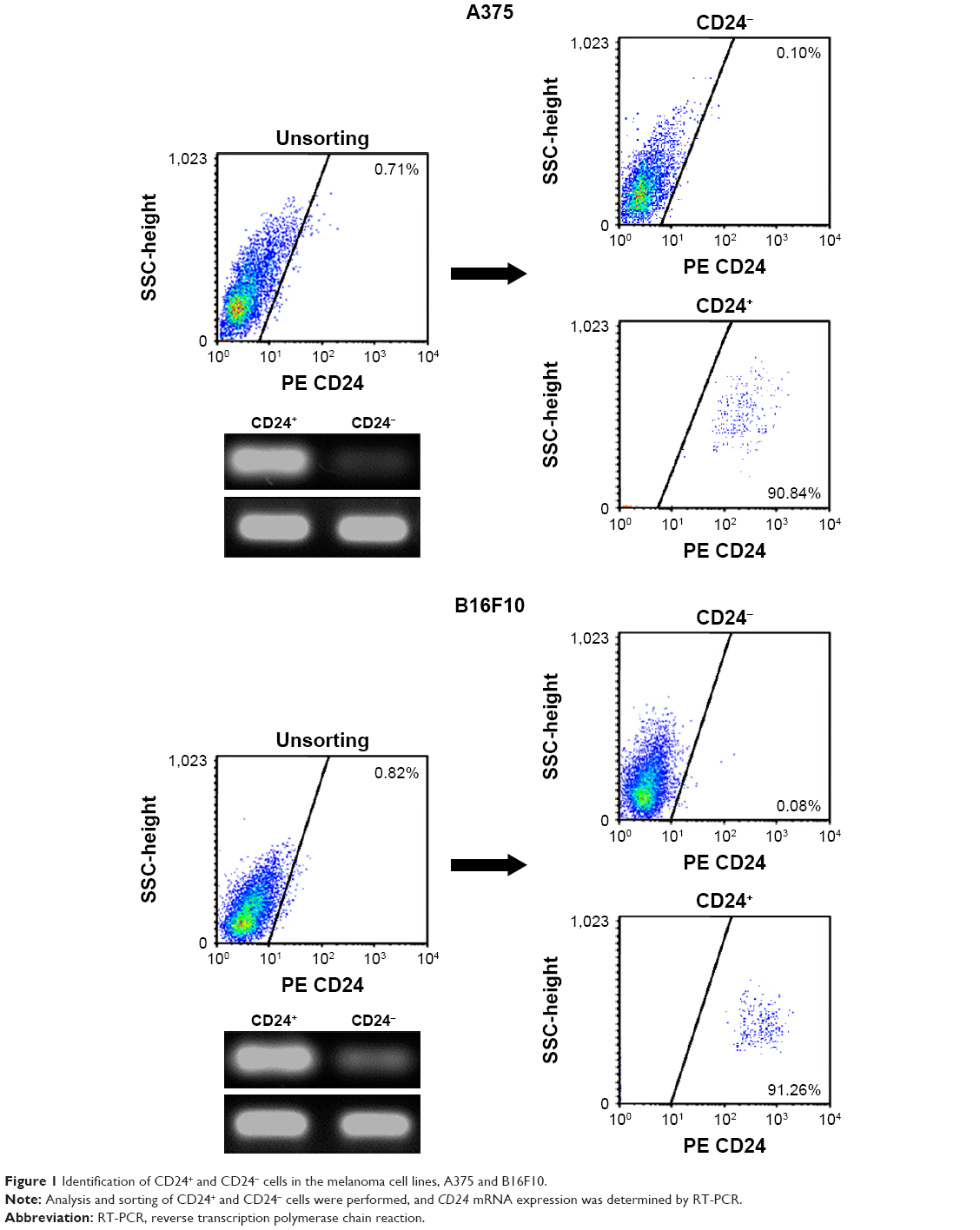 | Figure 1 Identification of CD24+ and CD24- cells in the melanoma cell lines, A375 and B16F10. |
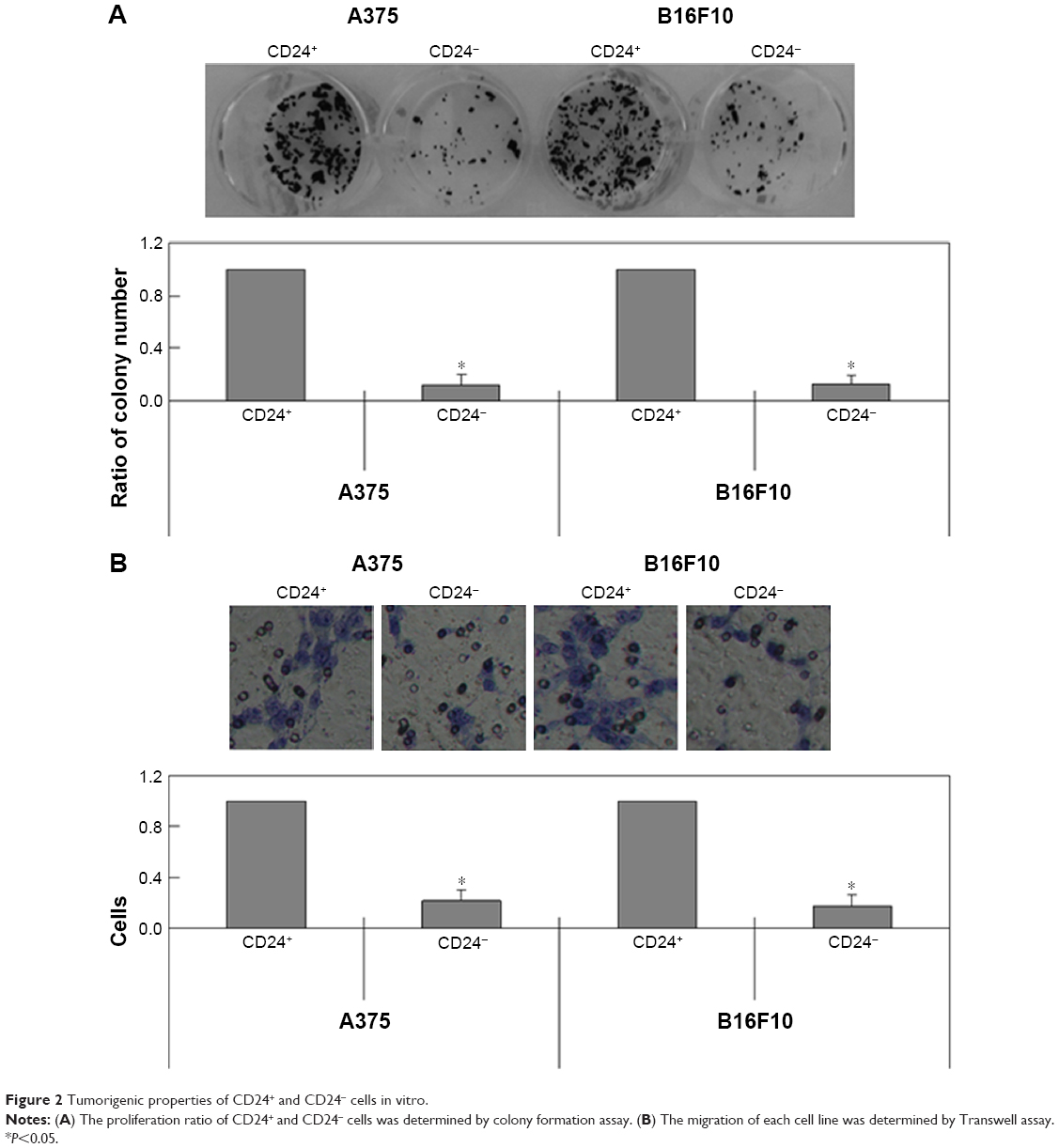 | Figure 2 Tumorigenic properties of CD24+ and CD24- cells in vitro. |
Next, we determined the tumorigenic ability of CD24- and CD24+ cells in mouse models. As in the in vitro experiments, the tumor size in mice carrying CD24- cells was smaller than that in mice with CD24+ cells (Figure 3A). We observed fewer metastases in liver tissues in CD24- groups compared with CD24+ groups (Figure 3B). The survival rate of the CD24- tumor-bearing mice was increased up to 60% compared with CD24+ tumor-bearing mice (P<0.05, Figure 3D).
 | Figure 3 Tumorigenic properties of CD24+ and CD24- cells in vivo. |
Changes of Notch1 signaling pathway in CD24- and CD24+ cells
The levels of Galectin-3, NICD1, JAG1, DLL4, Notch1, and N-cadherin were observed to be higher in CD24+ cells compared with CD24- cells using western blot (Figure 4). The expression level of E-cadherin was lower in CD24+ cells than that in CD24- ones (Figure 4). Furthermore, we found that increased CD24 was accompanied by high Notch1 expression in vivo (Figure 3C).
 | Figure 4 Western blot analysis of Notch1 signaling pathway. |
Discussion
CD24 is a widely used biomarker for isolating CSCs.13 Previous studies have shown that CD24+ cells have high capability to form tumors.8–10 However, other studies showed that CD24 is not a cancer stem cell marker.11,12,14 For example, Xu et al14 found that both A549 and H560 CD24+ cells did not show enhanced tumor-forming ability than CD24- cells. In this study, using the soft agar assay, we demonstrated that the CD24+ subpopulation has self-renewal properties. We also confirmed the tumorigenic potential of the distinct population in mouse models.
The main finding of this study was that CD24 expression is accompanied by activation of Notch1 signaling pathway. Notch1 was found increased in many kinds of malignant tumors, such as melanoma, prostate cancer, and hepatocellular carcinoma.15 Notch1 could promote the growth, survival, and metastasis of melanoma.16 Previous studies showed that Notch1 controls another cancer stem cell marker (CD133) expression at transcriptional and posttranscriptional levels and regulates the survival and proliferation of glioma cells17 and melanoma cells.18 So et al19 found that the CD44+/CD24-/low subpopulation of breast cancer cells showed elevated Notch1 signaling and increased cell proliferation compared to the CD44+/CD24high subpopulation. In contrast, we found the activation of the Notch1 signaling pathway in CD24+ cells. Many previous studies showed that Notch1 pathway plays a crucial role in CSCs maintaining stemness.20–22 Despite the opposite roles of Notch1 signaling in CD24 expression in breast cancer cells and melanoma cells, the results of both our study and So et al’s study confirmed that Notch1 signaling maintains the tumorigenic ability of CSCs.
Conclusion
In conclusion, in our study, we found CD24+ population in melanoma cells has the characteristics of CSCs. CD24 expression may be related with activation of Notch1 signaling pathway.
Acknowledgment
The authors thank Miss Zhang Yu for excellent technical assistance with the animal experiments.
Disclosure
The authors report no conflicts of interest in this work.
References
Eggermont AM, Spatz A, Robert C. Cutaneous melanoma. Lancet. 2014;383(9919):816–827. | ||
Cheraghi N, Cognetta A, Goldberg D. Radiation therapy in dermatology: non-melanoma skin cancer. J Drugs Dermatol. 2017;16(5):464–469. | ||
de Rosa F, Ridolfi L, Fiammenghi L, et al. Dendritic cell vaccination for metastatic melanoma: a 14-year monoinstitutional experience. Melanoma Res. 2017;27(4):351–357. | ||
Kunz M, Hölzel M. The impact of melanoma genetics on treatment response and resistance in clinical and experimental studies. Cancer Metastasis Rev. 2017;36(1):53–75. | ||
Crépin R, Gentien D, Duché A, et al. Nanobodies against surface biomarkers enable the analysis of tumor genetic heterogeneity in uveal melanoma patient-derived xenografts. Pigment Cell Melanoma Res. 2017;30(3):317–327. | ||
Tang MR, Wang YX, Guo S, Han SY, Li HH, Jin SF. CD24 expression predicts poor prognosis for patients with cutaneous malignant melanoma. Int J Clin Exp Med. 2014;7(11):4337–4341. | ||
Pirruccello SJ, LeBien TW. The human B cell-associated antigen CD24 is a single chain sialoglycoprotein. J Immunol. 1986;136(10):3779–3784. | ||
Shen YA, Wang CY, Chuang HY, et al. CD44 and CD24 coordinate the reprogramming of nasopharyngeal carcinoma cells towards a cancer stem cell phenotype through STAT3 activation. Oncotarget. 2016;7(36):58351–58366. | ||
Nakamura K, Terai Y, Tanabe A, et al. CD24 expression is a marker for predicting clinical outcome and regulates the epithelial-mesenchymal transition in ovarian cancer via both the Akt and ERK pathways. Oncol Rep. 2017;37(6):3189–3200. | ||
Lin L, Jou D, Wang Y, et al. STAT3 as a potential therapeutic target in ALDH+ and CD44+/CD24+ stem cell-like pancreatic cancer cells. Int J Oncol. 2016;49(6):2265–2274. | ||
Lü X, Xu K, Lü H, et al. CD44(+)/CD24(−) cells are transit progenitors and do not determine the molecular subtypes and clinical parameters in breast carcinomas. Ultrastruct Pathol. 2011;35(2):72–78. | ||
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A. 2003;100(7):3983–3988. | ||
Xia P. Surface markers of CSCs in solid tumors. Curr Stem Cell Res Ther. 2014;9(2):102–111. | ||
Xu H, Mu J, Xiao J, et al. CD24 negative lung cancer cells, possessing partial cancer stem cell properties, cannot be considered as CSCs. Am J Cancer Res. 2015;6(1):51–60. | ||
Gu Y, Masiero M, Banham AH. Notch signaling: its roles and therapeutic potential in hematological malignancies. Oncotarget. 2016;7(20):29804–29823. | ||
Shao H, Cai L, Moller M, et al. Notch1-WISP-1 axis determines the regulatory role of mesenchymal stem cell-derived stromal fibroblasts in melanoma metastasis. Oncotarget. 2016;7(48):79262–79273. | ||
Fan X, Khaki L, Zhu TS, et al. Notch pathway blockade depletes CD133-positive glioblastoma cells and inhibits growth of tumor neurospheres and xenografts. Stem Cells. 2010;28(1):5–16. | ||
Kumar D, Kumar S, Gorain M, et al. Notch1-MAPK signaling axis regulates CD133+ cancer stem cell-mediated melanoma growth and angiogenesis. J Invest Dermatol. 2016;136(12):2462–2474. | ||
So JY, Wahler J, Das Gupta S, et al. HES1-mediated inhibition of Notch1 signaling by a Gemini vitamin D analog leads to decreased CD44(+)/CD24(−/low) tumor-initiating subpopulation in basal-like breast cancer. J Steroid Biochem Mol Biol. 2015;148:111–121. | ||
Panaccione A, Zhang Y, Ryan M, et al. MYB fusions and CD markers as tools for authentication and purification of CSCs from salivary adenoid cystic carcinoma. Stem Cell Res. 2017;21:160–166. | ||
Zhang Y, Xu W, Guo H, et al. NOTCH1 signaling regulates self-renewal and platinum chemoresistance of cancer stem-like cells in human non-small cell lung cancer. Cancer Res. 2017;77(11):3082–3091. | ||
Feng HB, Wang J, Jiang HR, et al. β-Elemene selectively inhibits the proliferation of glioma stem-like cells through the downregulation of Notch1. Stem Cells Transl Med. 2017;6(3):830–839. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.