Back to Archived Journals » Journal of Neurorestoratology » Volume 4
Hypoxic and ischemic effects on gene and protein expression levels of paracrine factors by human olfactory mucosa mesenchymal-like stem cells
Authors Yuan T, Zhuo Y, Su C, Li X, Duan D, Ge L, Wu P, Wang H, Deng Y, Lu M
Received 30 July 2016
Accepted for publication 4 October 2016
Published 7 December 2016 Volume 2016:4 Pages 85—94
DOI https://doi.org/10.2147/JN.S118538
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Hongyun Huang
Ting Yuan,1* Yi Zhuo,1* Chaying Su,2 Xuan Li,1 Da Duan,1 Lite Ge,1 Pei Wu,1 Hao Wang,1 Yujia Deng,1 Ming Lu1
1Department of Neurosurgery, Second Affiliated Hospital of Hunan Normal University (163 Hospital of the People’s Liberation Army), Changsha, 2Department of Pediatrics, 169 Hospital of the People’s Liberation Army, Hengyang, Peoples’ Republic of China
* These authors contributed equally to this work
Abstract: Human olfactory mucosa mesenchymal-like stem cells (hOM-MSCs) secrete paracrine factors that may exert a protective effect on the cerebral ischemia. This study was done to determine the hypoxic and ischemic effects on the mRNA and protein expression level of paracrine factors by hOM-MSCs. The hOM-MSCs were cultured with 5% or 20% serum and under either normoxic (21% O2) or hypoxic (3% O2) conditions. Expression levels of mRNA and protein for vascular endothelial growth factor (VEGF), nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF) and matrix metalloproteinase-2 (MMP-2) were determined by quantitative reverse transcription polymerase chain reaction and Western blot, respectively. Hypoxia reduced gene expression for VEGF (5% serum), GDNF, BDNF (5% serum) and NGF (5% serum) and increased it for BDNF (20% serum), MMP-2 (5% serum) and NGF (20% serum). Ischemia lowered gene expression for VEGF (hypoxia), GDNF, BDNF (hypoxia), MMP-2 (normoxia) and NGF (hypoxia) and increased it for VEGF (normoxia), BDNF (normoxia), MMP-2 (hypoxia) and NGF (normoxia).The protein level of these factors was almost in line with the gene level. These data demonstrate that serum and oxygen levels have a significant effect on the gene and protein expression levels of paracrine factors by hOM-MSCs, which will affect how hOM-MSCs interact in vivo during cerebral ischemia.
Keywords: human olfactory mucosa mesenchymal-like stem cells, paracrine factors, hypoxia, ischemia, cerebral ischemia
Introduction
Work done in numerous laboratories has demonstrated that stem cell administration is able to reduce the loss of function in the brain after cerebral ischemia. There is a study that has demonstrated that intranasal treatment with mesenchymal stem cells (MSCs) reduces lesion volume and improves motor and cognitive behaviors in the neonatal hypoxic-ischemic mouse model.1 These MSCs secrete the paracrine factors, vascular endothelial growth factor (VEGF), nerve growth factor (NGF), brain derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF) and matrix metalloproteinase-2 (MMP-2), which play an important role in the biological functions of MSCs.2–7 It is important to understand the way in which the synthesis and secretion of these factors are controlled. This will help us better utilize MSCs in clinical situations.
The oxygen concentration present in the cultured medium has a significant effect on the behavior of MSCs. The atmospheric oxygen concentration routinely used for culture is significantly higher than that of physiological conditions in the body. A oxygen concentration is 10%–13% in arterial blood.8 A 1%–7% oxygen concentration is found in bone marrow microenvironment.9–11 There is only 3%–14% oxygen in brain.12 Hypoxia13 can affect the secretion of paracrine factors,14 alter gene expression of MSCs15,16 and enhance proliferation of MSCs.17–21
The amount of serum present in the culture media also affects stem cell behavior. When MSCs are cultured in 10% serum, some factors may not be present. Moreover, the ischemic conditions present after cerebral ischemia include both hypoxia and serum deprivation.22 Reduction in serum levels reduces cell proliferation and upregulates genes for angiogenic factors and endothelial differentiation in MSCs.23 Serum deprivation changes the expression of endothelial markers and the secretion of paracrine factors in MSCs.24,25
As a population of the MSCs, olfactory mucosa mesenchymal-like stem cells (OM-MSCs) may work effectively in the treatment of cerebral ischemia. Olfactory mucosa is a self-renewal tissue, containing olfactory ensheathing cells (OECs) and OM-MSCs. OECs are able to promote axonal outgrowth by secreting growth factors such as NGF.26,27 OM-MSCs can proliferate rapidly in vitro. Furthermore, OM-MSCs have the multiple potential to differentiate into cartilage, bone, smooth muscle and nerve cells.28 Our laboratory has analyzed the proteins that the human olfactory mucosa mesenchymal-like stem cells (hOM-MSCs) secrete. These hOM-MSCs also secrete the paracrine factors VEGF, NGF, BDNF, GDNF and MMP-2. Hence, the purpose of the present study was to mimic the conditions of cerebral ischemia by lowering oxygen and serum concentrations during culture. In order to determine the effects of these conditions on the gene and protein expression levels of hOM-MSCs, we cultured hOM-MSCs in 5% or 20% serum in either normoxic (21% O2) or hypoxic (3% O2) environment.
Methods
hOM-MSCs culture
The olfactory mucosa was obtained from the volunteers whose olfactory mucosa tissue was normal. Each volunteer signed a written informed consent to participate in this study. The ethics committee of Hunan Normal University approved this study. hOM-MSCs were isolated from olfactory mucosa and cultured in Dulbecco’s Modified Eagle Medium (DMEM)/F-12 medium with 20% fetal bovine serum (FBS) supplement as described previously.29 hOM-MSCs from passage 5 were cultured at 37°C in a flask in 2 mL/flask DMEM/F-12 with 5% or 20% FBS supplement. Normoxic cells were cultured for 48 h under normal atmospheric oxygen (21%) plus 5% CO2. Hypoxic cells were cultured for 48 h in a reduced oxygen atmosphere (3% O2, 5% CO2).
Phenotypic characterization of hOM-MSCs
Flow cytometric analysis of hOM-MSCs was performed as previously described.29 Briefly, cell suspensions were washed twice with phosphate-buffered saline (PBS) containing 0.1% FBS. One hundred thousand cells were incubated with CD34-fluorescein isothiocyanate (FITC), CD45-FITC, CD73–FITC and CD90-FITC (Abcam, Cambridge, UK) at 4°C for 30 min and then washed twice with PBS containing 0.1% FBS. As a negative control, the cells were incubated with the corresponding irrelevant immunoglobulin G (IgG)-FITC antibodies. The cells were analyzed by cytometric analysis using an EPICS XL flow cytometer (Becton-Dickinson, San Diego, CA, USA) with EXPO32 software (Beckman Coulter Inc., Brea, CA, USA).
Western blot analysis
After 48 h of culture, the cells were washed once with ice precooled PBS. Then media were removed and centrifuged at 3,000 rpm for 2 min. After supernatant removal, 50 mL of radioimmunoprecipitation assay lysis buffer was added and cleavaged on the ice for 30 min. The homogenate was centrifuged at 12,000 rpm for 15 min at 4°C. The supernatant was separated into a 0.5 mL centrifuge tube and stored at -20°C. In brief, lysates containing 50 μg proteins were subjected to gel electrophoresis. Proteins were then transferred to polyvinylidene difluoride membranes. The blots were blocked in 4% bovine serum albumin in Tris-buffered saline with Tween 20 solution for 30 min at room temperature and then incubated at 4°C overnight with the primary antibody: VEGF (1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), NGF (1:1,000; Proteintech, Rosemont, IL, USA), GDNF (1:1,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), BDNF (1:1,000; Santa Cruz Biotechnology, Inc.), MMP-2 (1:1,000; Abcam) and β-actin (1:4,000; Proteintech). After incubation with secondary antibodies at room temperature for 1 h, the blot was visualized using ChemiDoc XRS imaging system (Bio-Rad Laboratories Inc., Hercules, CA, USA).
mRNA quantification using quantitative reverse transcription polymerase chain reaction
Total RNA was isolated after 48 h of culture using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. RNA was treated with DNAse I and then quantified and assessed for quality by measuring absorbance at 260 and 280 nm. Reverse transcription was performed using PrimeScript RT Master Mix (Genstar, Beijing, People’s Republic of China). The cDNA amplification was performed by real-time polymerase chain reaction (PCR; LightCycler 480; Hoffman-La Roche Ltd., Roche, Switzerland) using GoTaq qPCR Master Mix (Promega Corporation, Fitchburg, WI, USA). Reverse transcription PCR primers were designed by Beacon 7.0 Primer Express software (Thermo Fisher Scientific). Specific primers of VEGF, NGF, GDNF, BDNF and MMP-2 in this study are listed in Table 1. Real-time PCR reactions were incubated at 95°C for 10 min and then for 40 cycles at 95°C for 5 s followed by at 60°C for 30 s. The measurements were performed in triplicate.
 | Table 1 PCR primers Abbreviations: PCR, polymerase chain reaction; VEGF, vascular endothelial growth factor; NGF, nerve growth factor; GDNF, glial cell line-derived neurotrophic factor; BDNF, brain-derived neurotrophic factor; MMP-2, matrix metalloproteinase-2. |
Statistical analysis
Mean and standard deviation were calculated for each treatment group, and significance was determined using Student’s t-test. IBM SPSS Statistics 18.0 (IBM Corporation, Armonk, NY, USA) was used for data analysis. A P-value <0.05 was considered significant. The following biologically relevant comparisons were made: 1) normoxic cells grown in 5% and 20% serum; 2) hypoxic cells grown in 5% and 20% serum; 3) normoxic cells with hypoxic cells grown in 5% serum; and 4) hypoxic cells with normoxic cells grown in 20% serum.
Results
Surface antigen characteristics of hOM-MSCs
As shown in Figure 1, we analyzed a panel of five membrane markers by flow cytometry. A characteristic feature of hOM-MSCs is a CD34-,CD45-, CD73+, CD90+ and CD105+ cell-surface phenotype.30 The CD analysis was performed in parallel on cells that were either adherent or clustered in spheres before being trypsinized.
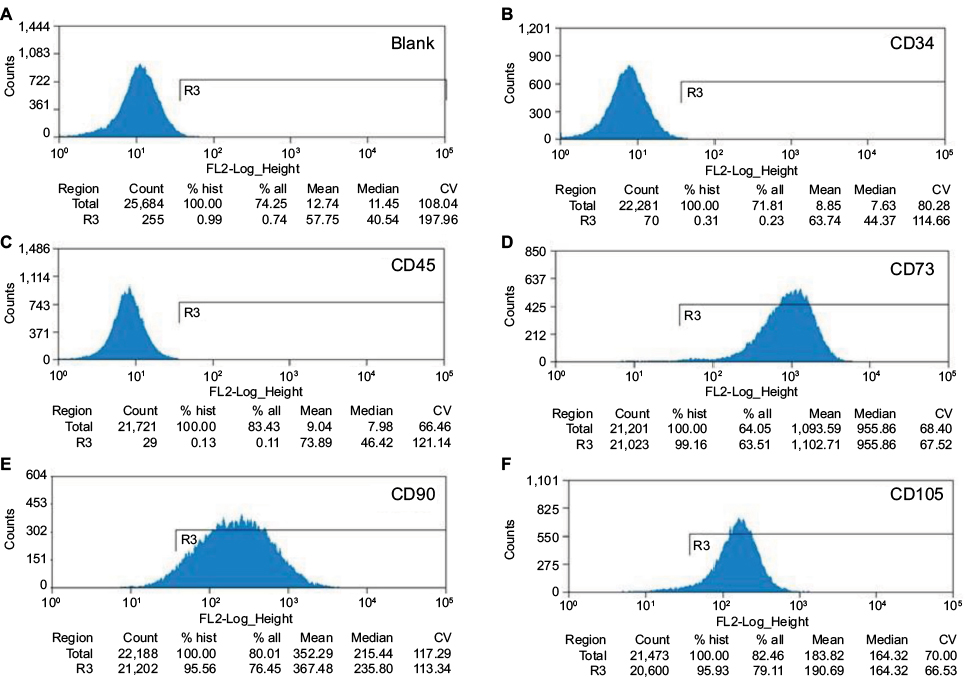 | Figure 1 Flow cytometric analysis of the expression of surface antigens on human OM-MSCs. Notes: (A) Isotype control, (B–C) there is no expression of CD34 and CD45. The cells were immunolabeled with the FITC conjugated monoclonal antibody specific for the indicated surface antigen, (D–F) there is notable expression of CD73, CD90 and CD105. % all indicates the percentage of target cells in the defined counting cells, % hist indicates the percentage of target cells in the actually measured cells. Abbreviations: OM-MSCs, olfactory mucosa mesenchymal-like stem cells; FITC, fluorescein isothiocyanate; CV, coefficient of variation |
Paracrine factor secretion
The secretion of paracrine factors by hOM-MSCs into the culture media was measured using Western blot (Figure 2). Serum reduction from 20% to 5% and changing from a normoxic to a hypoxic environment had remarkable effects on the protein levels detected in the media.
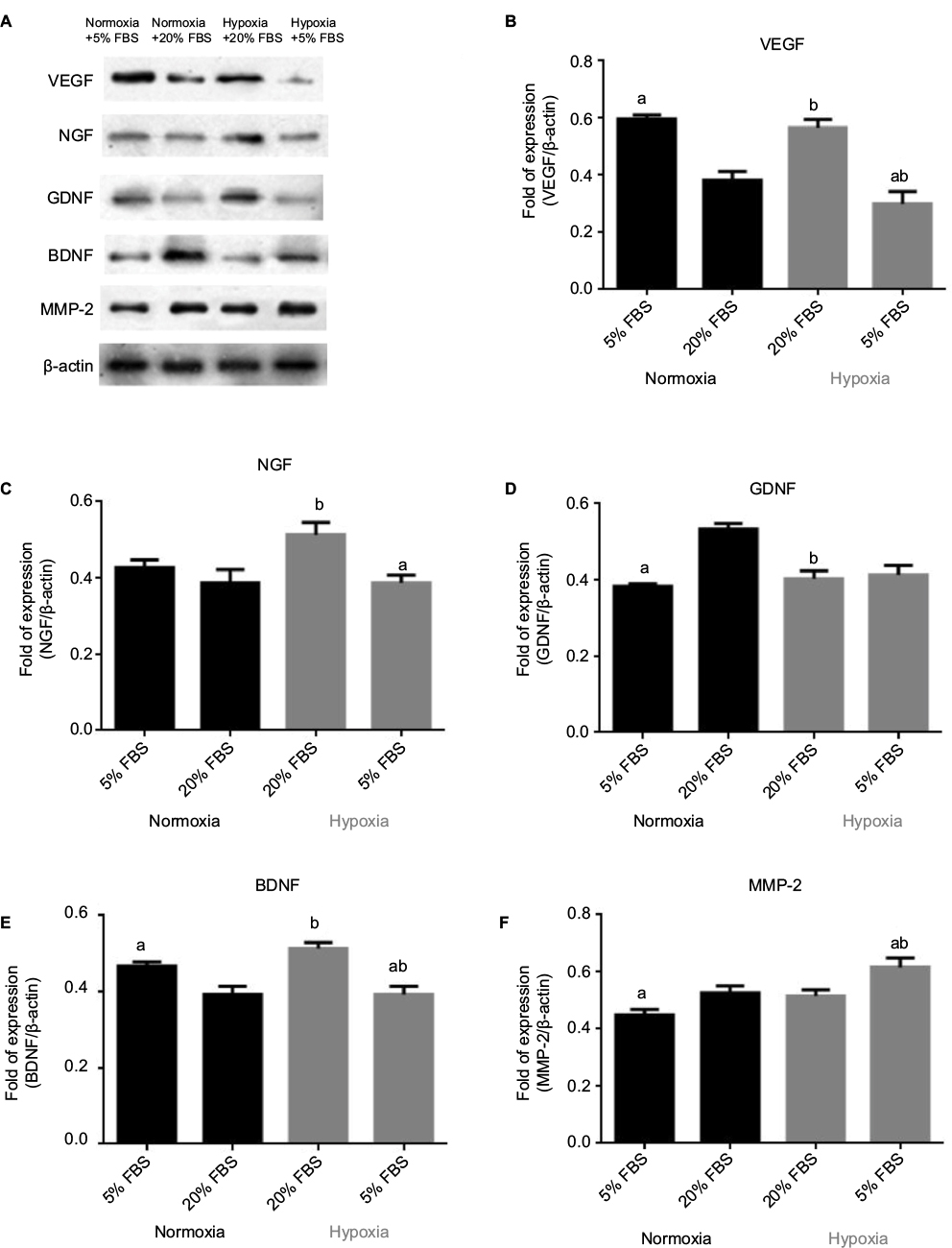 | Figure 2 Secretion of paracrine factors under different oxygen and serum conditions. Notes: Representative gel electrophoresis images (A) and analyzed data of the protein levels of (B) VEGF, (C) NGF, (D) GDNF, (E) BDNF and (F) MMP-2. Bar graph from densitometry analysis shows the ratio. Data reported as mean ± SEM. 5% and 20% refer to the amount of serum supplement present in the media. n=3 independent assays. (a) P<0.05 versus 20%. (b) P<0.05 versus normoxia. Abbreviations: VEGF, vascular endothelial growth factor; NGF, nerve growth factor; GDNF, glial cell line-derived neurotrophic factor; BDNF, brain-derived neurotrophic factor; MMP-2, matrix metalloproteinase-2; SEM, standard error of the mean; FBS, fetal bovine serum. |
VEGF levels were significantly elevated (P<0.05) in media from cells cultured in 5% serum supplement compared to those in 20% supplement in normoxic conditions, but in hypoxic conditions, the results were the opposite. Hypoxia had some effects on the secretion of VEGF.
As seen with VEGF, hypoxic conditions significantly increased (P<0.05) NGF levels in media compared to those in normoxic conditions in 20% serum supplement. Moreover, 5% serum supplement significantly reduced (P<0.05) NGF levels in the hypoxic environment.
GDNF levels were obviously reduced (P<0.05) in 5% serum compared to 20% serum in normoxic conditions, an effect not seen in hypoxic conditions. Also, hypoxia significantly reduced (P<0.05) GDNF secretion in 20% serum supplement.
In addition, serum reduction to 5% resulted in a meaningful elevation (P<0.05) of BDNF levels compared to 20% in normoxic conditions. Hypoxic conditions increased (P<0.05) BDNF levels in media compared to those in normoxic conditions in 20% serum supplement.
Changes in MMP-2 secretion followed the general trends as seen for GDNF. MMP-2 levels were obviously reduced (P<0.05) in 5% serum compared to 20% serum in normoxic conditions. However, MMP-2 levels were unaffected by oxygen levels in 20% serum supplement.
mRNA levels for paracrine factors
The normalized relative expression of mRNA in hOM-MSCs for the various paracrine factors was determined using quantitative reverse transcription PCR (Figure 3). Changes in mRNA paralleled changes in secretion even though a few differences were observed.
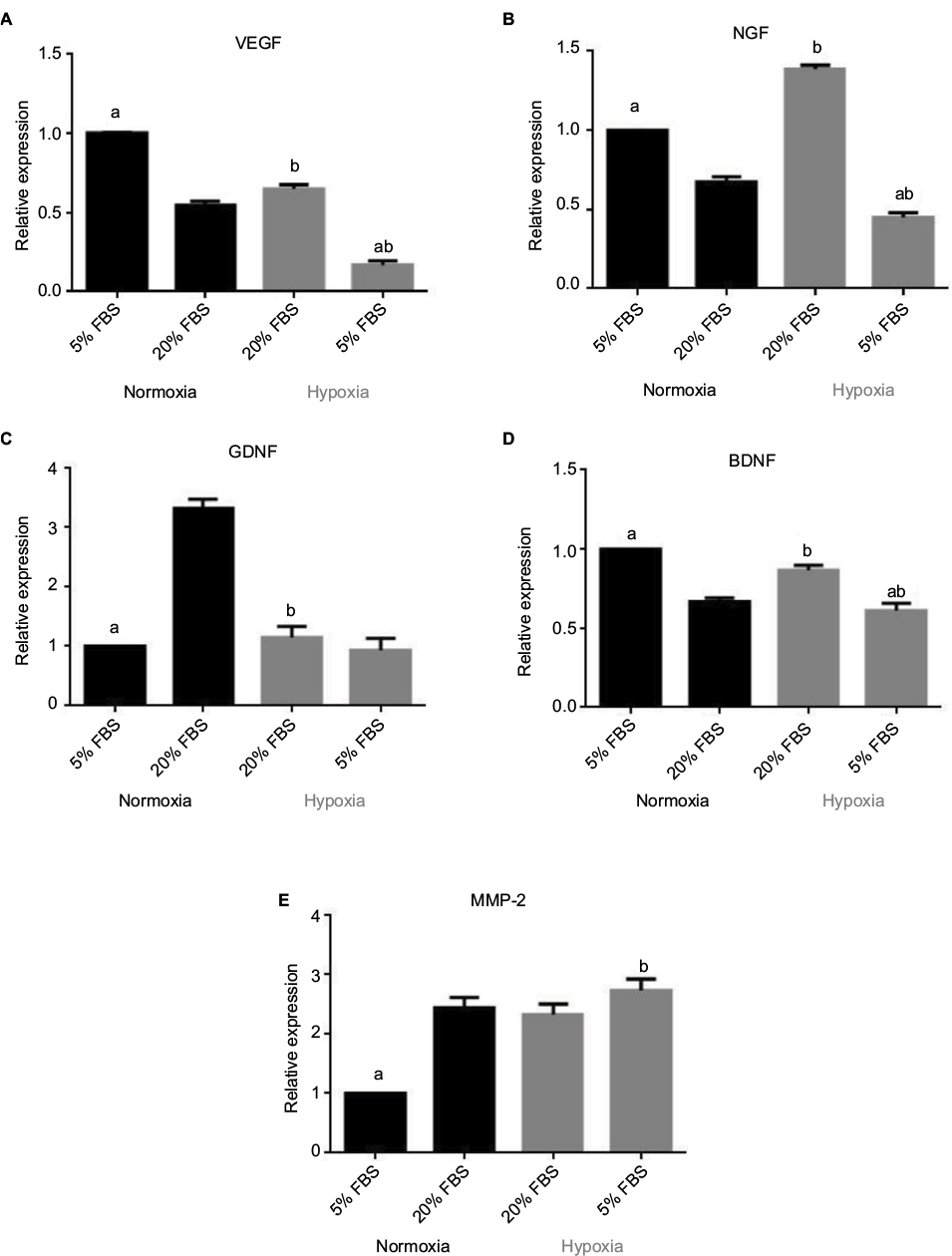 | Figure 3 Normalized relative expression of (A) VEGF, (B) NGF, (C) GDNF, (D) BDNF and (E) MMP-2 under different oxygen and serum conditions. Notes: Data reported as mean ± SEM. 5% and 20% refer to the amount of serum supplement present in the media. n=3 independent assays. (a) P<0.05 versus 20%. (b) P<0.05 versus normoxia. Abbreviations: VEGF, vascular endothelial growth factor; NGF, nerve growth factor; GDNF, glial cell line-derived neurotrophic factor; BDNF, brain-derived neurotrophic factor; MMP-2, matrix metalloproteinase-2; SEM, standard error of the mean; FBS, fetal bovine serum. |
Serum reduction to 5% caused a significant upgrade in the relative expression of VEGF mRNA compared to 20% in normoxic conditions (P<0.05), but in hypoxic conditions, the results were the opposite. Hypoxia resulted in an elevation of VEGF expression levels in 20% serum (P<0.05) but not in 5% serum. This is similar to the response observed with VEGF secretion into the surrounding media.
Serum reduction to 5% significantly increased (P<0.05) the relative expression of NGF mRNA compared to 20% serum supplement in normoxic conditions, but in hypoxic conditions, serum reduction to 5% decreased (P<0.05) the relative expression of NGF mRNA. Hypoxia reduced (P<0.05) the relative expression of NGF mRNA in 5% serum supplement, and in 20% serum supplement, the results were the opposite. This was similar to what was observed in NGF secretion except for the expression levels in normoxic conditions.
The relative expression of GDNF mRNA was significantly reduced (P<0.05) in 5% serum compared to 20% serum in normoxic conditions. No effect was seen in hypoxic conditions. Hypoxia obviously reduced mRNA expression in 20% (P<0.05) supplements. This effect was similar to what was observed for GDNF secretion.
For BDNF mRNA, reducing the serum from 20% to 5% led to a meaningful elevation (P<0.05) of the relative expression of BNDF mRNA in normoxic conditions. However, in hypoxic conditions, the relative expression of BDNF mRNA was reduced (P<0.05) in 5% serum compared to 20% serum. Hypoxic conditions increased (P<0.05) the relative expression of BDNF mRNA compared to that in normoxic conditions in 20% serum supplement. These effects were similar to secretion of BDNF into the culture media where hypoxia had no effect on BDNF secretion in 5% serum supplement.
Reducing the serum from 20% to 5% significantly decreased (P<0.05) the relative expression of MMP-2 mRNA in normoxic conditions. Nevertheless, reducing the oxygen concentration had no significant effect in 20% serum supplement. Hypoxia increased (P<0.05) the relative expression of MMP-2 mRNA in 5% serum supplement. These findings were similar to the results of the protein secretion experiments.
Discussion
The research distinctly demonstrates that alteration in oxygen concentrations and serum levels brings about significant changes in the mRNA expression and secretion of paracrine factors by hOM-MSCs. It is the first time (to the best of our knowledge) that these two parameters have been studied simultaneously. The data suggest that the low serum and oxygen conditions present during the ischemia found after brain ischemia may have significant effects on the secretory function of MSCs and the role they play in preventing the loss of cerebral function after cerebral infarction.31 The particular changes observed depend on the specific paracrine factors that were studied. Changes in mRNA often were in line with changes in secretion, and there were few exceptions.
Our research indicated that in 20% serum supplement, hypoxia significantly reduced the gene expression for GDNF but significantly increased the expression of genes for VEGF, NGF and BDNF, while MMP-2 expression was unaffected. Secretions of VEGF, NGF and BDNF were all significantly elevated by hypoxia in 20% serum supplement, while GDNF was decreased and MMP-2 was unaffected. Changes in secretions of VEGF, NGF, BDNF, GDNF and MMP-2 paralleled changes in expression of genes for VEGF, NGF, BDNF, GDNF and MMP-2 in 20% serum supplement. Meanwhile, the gene expression for NGF was reduced by hypoxia in 5% serum supplement, while the secretion of NGF was unaffected. The reasons for the differences between mRNA expression and secretion for NGF are not immediately apparent but may be due to differences in the control of gene expression versus control of translational and posttranslational events that lead from mRNA production to secretion. Other investigators have demonstrated an increase in VEGF gene expression and secretion by MSCs in hypoxia.14,15 Our results generally did demonstrate an increase in VEGF after hypoxia in 20% serum supplement, while the results are the opposite in 5% serum supplement. The serum levels may influence the gene expression for VEGF, so the secretion of VEGF in hypoxia was different between the 20% serum supplement and the 5% serum supplement. In addition, hypoxia increased gene expression for trophic/growth factors in MSCs, including BDNF, GDNF and VEGF and its receptor FIK-1; erythropoietin (EPO) and its receptor EPOR; stromal derived factor-1 and its CXC chemokine receptor 4; placental growth factor; heparin-binding epidermal growth factor; MMP-9, and basic fibroblast growth factor (bFGF).14,15,32 Hypoxia caused an increase in the secretion of transforming growth factor-β2; insulin-like growth factor (IGF) binding proteins 2, 3, 4 and 6; IGF-II and interleukin-7,33,34 and reduced the secretion of stromal cell derived factor-1, macrophage colony stimulating factor, interleukin-1 receptor antagonist, RANTES, chemokine (C-X-C motif) ligand 1, lactate dehydrogenase and chemokine (C-X-C motif) ligand 10 by MSCs.16,35 In short, hypoxia changes the paracrine secretions of MSCs, which would have implications for the role they play in reducing the loss of function in the brain after cerebral ischemia.
Exposure to a hypoxic environment leads to obvious changes in cellular physiology. Hypoxia-inducible factor (HIF)-1 is the key mediator of the physiological responses associated with reduction in tissue oxygen tension (hypoxia) and plays an important role in VEGF transcriptional activation.36,37 Transcription factor HIF-1 consists of two subunits: HIF-1α, the hypoxia-induced component, and HIF-1β, which is expressed constitutively. HIF-1α is rapidly degraded in normoxic cells. However, under conditions of low oxygen tension, HIF-1α stabilizes due to hypoxia-prevented degradation. One of the reasons is that the stabilization of HIF-1α in an oxygen-dependent manner is between 0.5% and 5%, which may be due to the release of reactive oxygen species by the mitochondria that prevents its degradation.38,39 This enhances the concentration of constitutively expressed HIF-1α, which forms an active transcription factor with HIF-1β to promote cell survival.40 A variety of genes are affected by this cascade. HIF-1α mRNA expression was upgraded in MSCs after 48 h of culture in hypoxic conditions.15 On the basis of this, it is not surprising that we found that hypoxia alters the gene expression and secretion of paracrine factors, and HIF-1α may play an important role in the response.
Our study demonstrated that reducing the serum concentration from 20% to 5% caused an increase in the mRNA expression for MMP-2 (hypoxia only) but a significant decrease in the expression for VEGF (hypoxia only), NGF (hypoxia only), GDNF (normoxia only) and BDNF (hypoxia only). Secretion of MMP-2 was elevated by serum reduction in hypoxia, while VEGF, NGF and BDNF were significantly reduced in hypoxia. Serum deprivation could promote the apoptosis of MSCs; the effect was stronger when hypoxia and serum deprivation were combined together. Moreover, the effect of serum deprivation on apoptosis was stronger than that of hypoxia.41 Alterations in gene expression and secretion after serum reduction are not surprising because the media contain less protein and fewer signaling growth factors. Some studies have proved that serum reduction or deprivation causes an increased expression and secretion of VEGF by MSCs.24,42 But in our research, only in normoxic conditions, serum reduction caused an increase in the mRNA expression for VEGF. It is unclear why lowering serum reduced expression and secretion of VEGF in hypoxic conditions. The reason may be that the effect of promoting the apoptosis of hOM-MSCs is stronger when hypoxia and serum deprivation are combined together. When hOM-MSCs were in a state of apoptosis, the expression and secretion of VEGF were decreased. Reports from other laboratories have indicated that reducing serum from 20% to 2% in MSC cultures resulted in a significant decline in proliferation42 and an increased expression of genes for bFGF and angiogenin-1.23 Serum deprivation led to epigenetic changes in MSCs, which upregulated the expression of the IGF-1 and leptin.43 Hence, the results have significant therapeutic implications for the paracrine actions of hOM-MSCs, because cerebral ischemia results in a serum-deprived state in the brain.
Hypoxia and serum reduction also have an effect on gene expression associated with differentiation in MSCs. Hypoxic MSCs maintained significantly higher colony-forming unit capabilities and expressed higher levels of stem cell genes than MSCs cultured at normoxic conditions.44 Hypoxia promoted osteogenic differentiation of MSCs, and the mechanism of hypoxia-mediated signaling in osteogenic differentiation in MSCs is direct regulation of RUNX2 by TWIST.45 Besides, hypoxia and transforming growth factor-beta drive differentiation of MSCs toward a phenotype consistent with that of the nucleus pulposus.46 More importantly, MSCs expanded under hypoxic conditions exhibited telomerase activity with maintained telomere length, normal karyotyping and intact genetic integrity and did not form tumors.47 These results supported that hypoxic culture as a method for efficiently expanding MSCs is safe. MSCs maintained most of their stemness and angiogenic genes’ expression in 10% FBS, but this increased significantly in 2% FBS. Reduction in the serum levels to 2% caused an increased expression of endogenic genes such as platelet endothelial cell adhesion molecule-1, vascular endothelial cadherin and VEGF receptor-2.23 Serum reduction resulted in an increased endothelial differentiation by MSCs.24 Reduction in the serum levels to 2% caused a lower growth rate in MSCs.23 The MSCs’ death rates were not affected by the shift to 48h hypoxia, but increased with FBS starvation, demonstrating that serum deprivation is the stronger factor. Long-term serum deprivation combined with hypoxia led to the complete death of MSCs.48 Hence, the hypoxic/low serum conditions present after cerebral ischemia provide conditions that maintain the pluripotency of MSCs while also promoting their ability to differentiate. The changes in the differentiation state might also affect the expression and secretion of paracrine factors in MSCs.
The paracrine factors secreted by MSCs have significant biologic activities that are believed to play a role in their brain effects.49,50 MSCs are a potent source of VEGF, which is a promoter and regulator of angiogenesis.2,51 Neurons exposed to hypoxic conditions exhibited neuronal apoptosis, but the combined hypoxia-inducible VEGF and MSCs treatment indicated a significant increase in VEGF expression and decreased neuronal apoptosis.52 Also, grafting MSCs after a spinal cord compression injury revealed a significant increase in NGF expression in the injured tissue, which promoted the recovery of spinal cord function.3 Another study indicated that grafted MSCs survived after transplantation into intact spinal cord and produced cells that expressed NGF and BDNF.53 GDNF has been shown to be capable of protecting motor neurons in amyotrophic lateral sclerosis, and MSCs improved the motor tests performed, which coincided with a higher GDNF immunoreactivity in the grafted spinal cord.5 MMP-2 is a gelatinase that is secreted by MSCs.54 Hypoxia preconditioning-treated MSCs showed a higher level of expression of MMP-2 and exhibited enhanced homing efficiency to the infarct cortex.55 All these data suggest that MSCs could be a potential source of cells to promote cell survival and axonal growth of host tissue. Differential regulation of these paracrine factors by hypoxic and low serum conditions as demonstrated in this research may play an important role in the beneficial effects of MSCs injection during cerebral ischemia.
Acknowledgments
This work was supported by National Natural Science Foundation of China (No. 81371358). The authors would like to thank Yangyun Liu and Xiurong Feng from the Department of Otolaryngology in the Second Affiliated Hospital of Hunan Normal University for their assistance.
Disclosure
The authors report no conflicts of interest in this work.
References
Donega V, Nijboer CH, Braccioli L, et al. Intranasal administration of human MSC for ischemic brain injury in the mouse: in vitro and in vivo neuroregenerative functions. PLoS One. 2014;9(11):e112339. | ||
Wang M, Zhang W, Crisostomo P, et al. STAT3 mediates bone marrow mesenchymal stem cell VEGF production. J Mol Cell Cardiol. 2007;42(6):1009–1015. | ||
Quertainmont R, Cantinieaux D, Botman O, Sid S, Schoenen J, Franzen R. Mesenchymal stem cell graft improves recovery after spinal cord injury in adult rats through neurotrophic and pro-angiogenic actions. PLoS One. 2012;7(6):e39500. | ||
Somoza R, Juri C, Baes M, Wyneken U, Rubio FJ. Intranigral transplantation of epigenetically induced BDNF-secreting human mesenchymal stem cells: implications for cell-based therapies in Parkinson’s disease. Biol Blood Marrow Transplant. 2010;16(11):1530–1540. | ||
Pastor D, Viso-Leon MC, Jones J, et al. Comparative effects between bone marrow and mesenchymal stem cell transplantation in GDNF expression and motor function recovery in a motorneuron degenerative mouse model. Stem Cell Rev. 2012;8(2):445–458. | ||
Yu Q, Chen L, You Y, et al. Erythropoietin combined with granulocyte colony stimulating factor enhances MMP-2 expression in mesenchymal stem cells and promotes cell migration. Mol Med Rep. 2011;4(1):31–36. | ||
Bhoopathi P, Chetty C, Gogineni VR, et al. MMP-2 mediates mesenchymal stem cell tropism towards medulloblastoma tumors. Gene Ther. 2011;18(7):692–701. | ||
Sullivan M, Galea P, Latif S. What is the appropriate oxygen tension for in vitro culture. Mol Hum Reprod. 2006;12(11):653. | ||
Fehrer C, Brunauer R, Laschober G, et al. Reduced oxygen tension attenuates differentiation capacity of human mesenchymal stem cells and prolongs their lifespan. Aging Cell. 2007;6(6):745–757. | ||
Guitart AV, Debeissat C, Hermitte F, et al. Very low oxygen concentration (0.1%) reveals two FDCP-Mix cell subpopulations that differ by their cell cycling, differentiation and p27KIP1 expression. Cell Death Differ. 2011;18(1):174–182. | ||
Lennon DP, Edmison JM, Caplan AI. Cultivation of rat marrow-derived mesenchymal stem cells in reduced oxygen tension: effects on in vitro and in vivo osteochondrogenesis. J Cell Physiol. 2001;187(3):345–355. | ||
Lee JH, Kemp DM. Human adipose-derived stem cells display myogenic potential and perturbed function in hypoxic conditions. Biochem Biophys Res Commun. 2006;341(3):882–888. | ||
Ma T, Grayson WL, Frohlich M, Vunjak-Novakovic G. Hypoxia and stem cell-based engineering of mesenchymal tissues. Biotechnol Prog. 2009;25(1):32–42. | ||
Ohnishi S, Yasuda T, Kitamura S, Nagaya N. Effect of hypoxia on gene expression of bone marrow-derived mesenchymal stem cells and mononuclear cells. Stem Cells. 2007;25(5):1166–1177. | ||
Liu L, Gao J, Yuan Y, Chang Q, Liao Y, Lu F. Hypoxia preconditioned human adipose derived mesenchymal stem cells enhance angiogenic potential via secretion of increased VEGF and bFGF. Cell Biol Int. 2013;37(6):551–560. | ||
Burlacu A, Grigorescu G, Rosca AM, Preda MB, Simionescu M. Factors secreted by mesenchymal stem cells and endothelial progenitor cells have complementary effects on angiogenesis in vitro. Stem Cells Dev. 2013;22(4):643–653. | ||
Lavrentieva A, Majore I, Kasper C, Hass R. Effects of hypoxic culture conditions on umbilical cord-derived human mesenchymal stem cells. Cell Commun Signal. 2010;8:18. | ||
Dos SF, Andrade PZ, Boura JS, Abecasis MM, da SCL, Cabral JM. Ex vivo expansion of human mesenchymal stem cells: a more effective cell proliferation kinetics and metabolism under hypoxia. J Cell Physiol. 2010;223(1):27–35. | ||
Moussavi-Harami F, Duwayri Y, Martin JA, Moussavi-Harami F, Buckwalter JA. Oxygen effects on senescence in chondrocytes and mesenchymal stem cells: consequences for tissue engineering. Iowa Orthop J. 2004;24:15–20. | ||
Grayson WL, Zhao F, Bunnell B, Ma T. Hypoxia enhances proliferation and tissue formation of human mesenchymal stem cells. Biochem Biophys Res Commun. 2007;358(3):948–953. | ||
Carrancio S, Lopez-Holgado N, Sanchez-Guijo FM, et al. Optimization of mesenchymal stem cell expansion procedures by cell separation and culture conditions modification. Exp Hematol. 2008;36(8):1014–1021. | ||
Horita Y, Honmou O, Harada K, Houkin K, Hamada H, Kocsis JD. Intravenous administration of glial cell line-derived neurotrophic factor gene-modified human mesenchymal stem cells protects against injury in a cerebral ischemia model in the adult rat. J Neurosci Res. 2006;84(7):1495–1504. | ||
Chua KH, Raduan F, Wan SWK, Manzor NF, Pingguan-Murphy B, Sathapan S. Effects of serum reduction and VEGF supplementation on angiogenic potential of human adipose stromal cells in vitro. Cell Prolif. 2013;46(3):300–311. | ||
Oskowitz A, McFerrin H, Gutschow M, Carter ML, Pochampally R. Serum-deprived human multipotent mesenchymal stromal cells (MSCs) are highly angiogenic. Stem Cell Res. 2011;6(3):215–225. | ||
Follin B, Tratwal J, Haack-Sorensen M, Elberg JJ, Kastrup J, Ekblond A. Identical effects of VEGF and serum-deprivation on phenotype and function of adipose-derived stromal cells from healthy donors and patients with ischemic heart disease. J Transl Med. 2013;11:219. | ||
Rao Y, Zhu W, Liu H, Jia C, Zhao Q, Wang Y. Clinical application of olfactory ensheathing cells in the treatment of spinal cord injury. J Int Med Res. 2013;41(2):473–481. | ||
Tabakow P, Jarmundowicz W, Czapiga B, et al. Transplantation of autologous olfactory ensheathing cells in complete human spinal cord injury. Cell Transplant. 2013;22(9):1591–1612. | ||
Shafiee A, Kabiri M, Ahmadbeigi N, et al. Nasal septum-derived multipotent progenitors: a potent source for stem cell-based regenerative medicine. Stem Cells Dev. 2011;20(12):2077–2091. | ||
Ge L, Zhuo Y, Duan D, et al. [Biological characteristics of human olfactory mucosa mesenchymal stem cells]. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2015;40(1):53–58. | ||
Lindsay SL, Johnstone SA, Mountford JC, et al. Human mesenchymal stem cells isolated from olfactory biopsies but not bone enhance CNS myelination in vitro. Glia. 2013;61(3):368–382. | ||
van Velthoven CT, Sheldon RA, Kavelaars A, et al. Mesenchymal stem cell transplantation attenuates brain injury after neonatal stroke. Stroke. 2013;44(5):1426–1432. | ||
Wei L, Fraser JL, Lu ZY, Hu X, Yu SP. Transplantation of hypoxia preconditioned bone marrow mesenchymal stem cells enhances angiogenesis and neurogenesis after cerebral ischemia in rats. Neurobiol Dis. 2012;46(3):635–645. | ||
Hung SC, Pochampally RR, Chen SC, Hsu SC, Prockop DJ. Angiogenic effects of human multipotent stromal cell conditioned medium activate the PI3K-Akt pathway in hypoxic endothelial cells to inhibit apoptosis, increase survival, and stimulate angiogenesis. Stem Cells. 2007;25(9):2363–2370. | ||
Hung SP, Ho JH, Shih YR, Lo T, Lee OK. Hypoxia promotes proliferation and osteogenic differentiation potentials of human mesenchymal stem cells. J Orthop Res. 2012;30(2):260–266. | ||
Bader AM, Klose K, Bieback K, et al. Hypoxic preconditioning increases survival and pro-angiogenic capacity of human cord blood mesenchymal stromal cells in vitro. PLoS One. 2015;10(9):e0138477. | ||
Forsythe JA, Jiang BH, Iyer NV, et al. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol Cell Biol. 1996;16(9):4604–4613. | ||
Jiang BH, Agani F, Passaniti A, Semenza GL. V-SRC induces expression of hypoxia-inducible factor 1 (HIF-1) and transcription of genes encoding vascular endothelial growth factor and enolase 1: involvement of HIF-1 in tumor progression. Cancer Res. 1997;57(23):5328–5335. | ||
Guzy RD, Schumacker PT. Oxygen sensing by mitochondria at complex III: the paradox of increased reactive oxygen species during hypoxia. Exp Physiol. 2006;91(5):807–819. | ||
Jiang BH, Semenza GL, Bauer C, Marti HH. Hypoxia-inducible factor 1 levels vary exponentially over a physiologically relevant range of O2 tension. Am J Physiol. 1996;271(4 pt 1):C1172–C1180. | ||
Haque N, Rahman MT, Abu KNH, Alabsi AM. Hypoxic culture conditions as a solution for mesenchymal stem cell based regenerative therapy. ScientificWorldJournal. 2013;2013:632972. | ||
Fu WL, Jia ZQ, Wang WP, et al. Proliferation and apoptosis property of mesenchymal stem cells derived from peripheral blood under the culture conditions of hypoxia and serum deprivation. Chin Med J (Engl). 2011;124(23):3959–3967. | ||
Katsuno T, Ozaki T, Saka Y, et al. Low serum cultured adipose tissue-derived stromal cells ameliorate acute kidney injury in rats. Cell Transplant. 2013;22(2):287–297. | ||
Sanchez C, Oskowitz A, Pochampally RR. Epigenetic reprogramming of IGF1 and leptin genes by serum deprivation in multipotential mesenchymal stromal cells. Stem Cells. 2009;27(2):375–382. | ||
Grayson WL, Zhao F, Izadpanah R, Bunnell B, Ma T. Effects of hypoxia on human mesenchymal stem cell expansion and plasticity in 3D constructs. J Cell Physiol. 2006;207(2):331–339. | ||
Yang DC, Yang MH, Tsai CC, Huang TF, Chen YH, Hung SC. Hypoxia inhibits osteogenesis in human mesenchymal stem cells through direct regulation of RUNX2 by TWIST. PLoS One. 2011;6(9):e23965. | ||
Risbud MV, Albert TJ, Guttapalli A, et al. Differentiation of mesenchymal stem cells towards a nucleus pulposus-like phenotype in vitro: implications for cell-based transplantation therapy. Spine (Phila Pa 1976). 2004;29(23):2627–2632. | ||
Tsai CC, Chen YJ, Yew TL, et al. Hypoxia inhibits senescence and maintains mesenchymal stem cell properties through down-regulation of E2A-p21 by HIF-TWIST. Blood. 2011;117(2):459–469. | ||
Potier E, Ferreira E, Meunier A, Sedel L, Logeart-Avramoglou D, Petite H. Prolonged hypoxia concomitant with serum deprivation induces massive human mesenchymal stem cell death. Tissue Eng. 2007;13(6):1325–1331. | ||
Sart S, Liu Y, Ma T, Li Y. Microenvironment regulation of pluripotent stem cell-derived neural progenitor aggregates by human mesenchymal stem cell secretome. Tissue Eng Part A. 2014;20(19–20):2666–2679. | ||
Segal-Gavish H, Karvat G, Barak N, et al. Mesenchymal stem cell transplantation promotes neurogenesis and ameliorates autism related behaviors in BTBR mice. Autism Res. 2016;9(1):17–32. | ||
Shibuya M. Vascular endothelial growth factor and its receptor system: physiological functions in angiogenesis and pathological roles in various diseases. J Biochem. 2013;153(1):13–19. | ||
An SS, Jin HL, Kim KN, et al. Neuroprotective effect of combined hypoxia-induced VEGF and bone marrow-derived mesenchymal stem cell treatment. Childs Nerv Syst. 2010;26(3):323–331. | ||
Alexanian AR, Maiman DJ, Kurpad SN, Gennarelli TA. In vitro and in vivo characterization of neurally modified mesenchymal stem cells induced by epigenetic modifiers and neural stem cell environment. Stem Cells Dev. 2008;17(6):1123–1130. | ||
Lozito TP, Jackson WM, Nesti LJ, Tuan RS. Human mesenchymal stem cells generate a distinct pericellular zone of MMP activities via binding of MMPs and secretion of high levels of TIMPs. Matrix Biol. 2014;34:132–143. | ||
Wei N, Yu SP, Gu X, et al. Delayed intranasal delivery of hypoxic-preconditioned bone marrow mesenchymal stem cells enhanced cell homing and therapeutic benefits after ischemic stroke in mice. Cell Transplant. 2013;22(6):977–991. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.