Back to Journals » Infection and Drug Resistance » Volume 16
Humoral Immune Responses to P. falciparum Circumsporozoite Protein (Pfcsp) Induced by the RTS, S Vaccine – Current Update
Authors Ngulube P ![]()
Received 13 December 2022
Accepted for publication 23 March 2023
Published 12 April 2023 Volume 2023:16 Pages 2147—2157
DOI https://doi.org/10.2147/IDR.S401247
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Peter Ngulube
Department of Biological Sciences, Academy of Medical Sciences, Malawi University of Science and Technology, Thyolo, Malawi
Correspondence: Peter Ngulube, Email [email protected]
Abstract: Malaria vaccines targeting the circumsporozoite protein (CSP) of the P. falciparum parasite have been overall relatively promising. RTS, S is a pre-erythrocytic recombinant protein-based malaria vaccine that targets CSP. RTS, S effectiveness shows some limited success regardless of its 58% efficacy for severe disease. P. falciparum circumsporozoite protein (Pfcsp) has stood to be the main candidate protein for most pre-erythrocytic stage vaccines. Studies on the structural and biophysical characteristics of antibodies specific to CSP (anti-CSP) are underway to achieve fine specificity with the CSP polymorphic regions. More recent studies have proposed the use of different kinds of monoclonal antibodies, the use of appropriate adjuvants, ideal vaccination dose and frequency, and improved targeting of particular epitopes for the robust production of functional antibodies and high complement-fixing activity as other potential methods for achieving long-lasting RTS, S. This review highlights recent findings regarding humoral immune responses to CSP elicited by RTS, S vaccine.
Keywords: Malaria, RTS, S, Circumsporozoite protein, RTS, S/AS01, Antibody response
Introduction
Malaria poses a huge burden with estimated cases of 245 million in 2020 and 247 million in 2021 globally. 1,2 New tools, techniques, and interventions to combat malaria are needed, especially in highly endemic areas such as sub-Saharan Africa. It is noted that infants, children under the age of five, pregnant women, and HIV/AIDS patients are at considerably higher risk of contracting malaria and developing severe disease because their immunity weakens during the infection.3 Research and innovation including drug efficacy monitoring programs, the development of new anti-malarial drugs, and novel insecticide-based vector control tools have further paved a step ahead to malaria eradication.4
RTS, S is a Plasmodium falciparum (Pf) pre-erythrocytic malaria vaccine that has undergone Phase III clinical trials in children from Africa.3,5 The generalisability of much-published clinical work overlooks the incredible complexity of Pf,6 such as expressing various kinds of proteins, hence invading multiple kinds of host cells.7 Notably, Pf is genetically diverse with approximately 5000 genes, hence prone to antigenic variation.8 This has been observed in a series of Phase I, III, and III clinical trials performed in many different populations including children, and infants, as young as 6 weeks of age.9–11
The Strategic Advisory Group of Experts on Immunization (SAGE) and the Malaria Policy Advisory Committee (MPAC) recommendations on pilot implementations of RTS, S was officially adopted by the World Health Organization (WHO) in January 2016. 12,13 Over the five years (2009–2014), the Phase III study for RTS, S/AS01 (RTS, S) was carried out in seven sub-Saharan African nations namely Burkina Faso, Gabon, Ghana, Kenya, Malawi, Mozambique, and the United Republic of Tanzania. The trial settings in these nations were representative of various malaria transmission environments (low, medium, and high).14 The results indicated that the vaccine was safe and 58% efficacious against severe malaria.14
The CSP is present on the surface of Pf sporozoites and contains highly polymorphic regions that interact with antibodies.15 The mechanism of Pfcsp C-terminal antibodies has been described with various characteristics such as the structural and biophysical correlation of the C-terminal-specific antibodies that have been widely discussed in several studies.15 However, anti-Pfcsp antibodies that are highly specific to merozoite CSP polymorphic regions elicit different immune responses, which has further affected the development of the broadly effective vaccine.11,15 Thus, this article presents recent findings on the antibody-mediated responses elicited by the RTS, S malaria vaccine.
The P. falciparum – Life Cycle and Vaccine Targets
There are essentially three phases in the malaria life cycle: Sporogony, exoerythrocytic schizogony, and erythrocytic schizogony.16 The mosquito phase involves the development of sporozoites that are infectious to humans from gametocytes through a process called sporogony (Figure 1). The process of transforming gametocytes into sporozoites takes place within the oocysts attached to the stomach wall of the mosquito.17 Sporozoites are injected into circulation after one to two hours following a mosquito bite and transmitted to the liver through the bloodstream,18 where they produce merozoites that are released into the bloodstream to invade red blood cells. The liver phase process where merozoites are formed from sporozoites is called exoerythrocytic schizogony, which takes seven to ten days.19
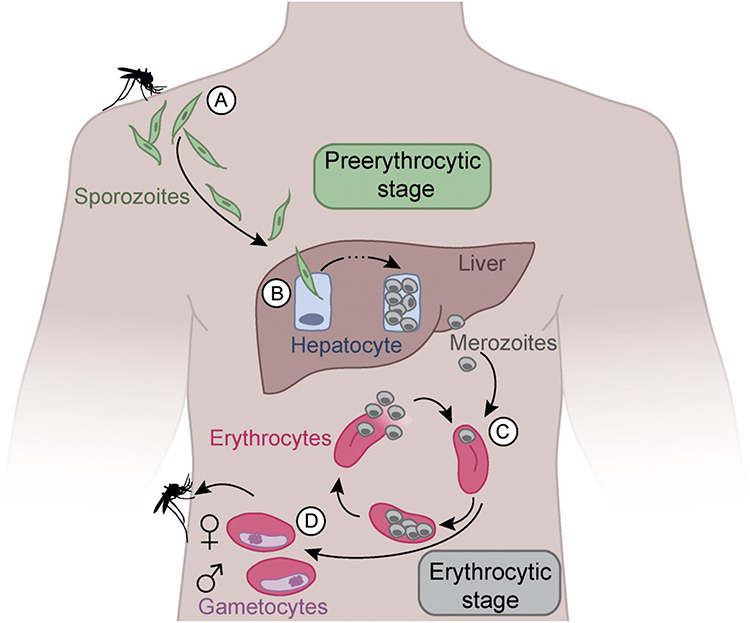 |
Figure 1 The P. falciparum life cycle and vaccine specific targets. Mosquito bite injects sporozoites into the circulation (A). Sporozoites travel to the liver and invade hepatocytes (B). Schizonts raptures sporozoites into the circulation hence invading erythrocytes (C). Merozoites enter the asexual stage forming gametocytes hence another cycle (D). Notes: Used with permission of Rockefeller University Press, from How to induce protective humoral immunity against Plasmodium falciparum circumsporozoite protein, Wahl I, Wardemann H, volume 219/ edition 2, 2022; permission conveyed through Copyright Clearance Center, Inc.19 |
The blood phase involves merozoites from the liver that enter circulation and transform into trophozoites and then schizonts that are composed of many merozoites.20 The schizonts burst, releasing merozoites into the bloodstream that subsequently invade more red blood cells.21 The blood phase of the malaria life cycle is called erythrocytic schizogony.21 Almost all malaria drugs treat the blood phase of malaria. Malaria vaccines target specific regions on the surface of the parasite to elicit an immune response such as CSP and thrombospondin-related adhesion protein.22,23 CSP has stood to be the main candidate protein for all pre-erythrocytic stage vaccines.
The P. falciparum Circumsporozoite Protein
Pfcsp (Figure 2) presents an important aspect of the new malaria vaccine era. Researchers and vaccine developers have spent decades studying its structural and biophysical characteristics and hence found that it is the only promising protein that can lead to an effective malaria vaccine.24 It is important to note that, CSP plays a huge role during the invasion of liver cells, and it is the largest and most prominent immunogenic region on the surface of Pf. CSP has three distinct regions: N-terminal, C-terminal, and central repeat.25 Nevertheless, these regions are characterized by highly polymorphic motifs that inhibit effective immune responses. Additionally, CSP contains 420 amino acid residues and is approximately 42kD.26
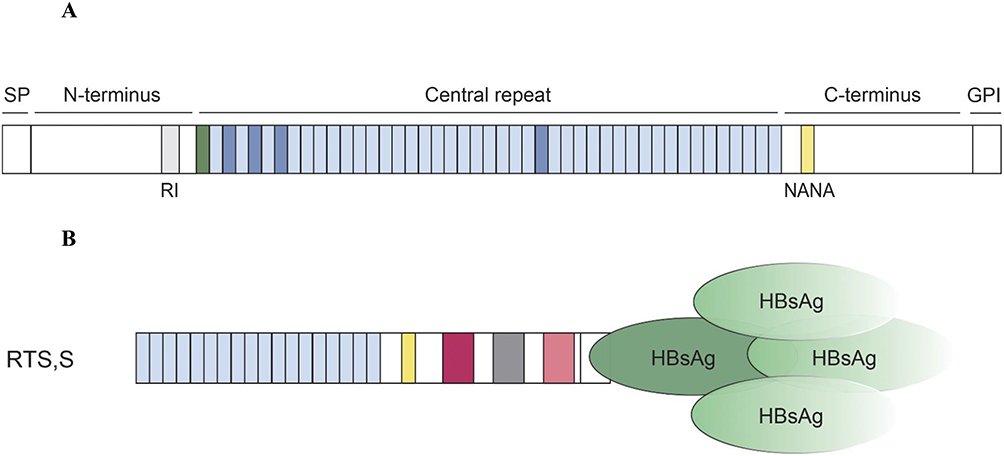 |
Figure 2 (A). The structure of the P. falciparum CSP; contains three main regions which are the N-terminal that consists of signal peptide (SP) and the region 1 (RI), the C-terminal, and the central repeat. CSP is strongly attached to the membrane of sporozoites with a GP1 anchor. Repeating NANP motifs (light blue), are present in the central repeat region that joins the N-terminus via a junction. NPDP (green), NANP, and NVDP (dark blue) are the main motifs targeted by the antibodies. Used with permission of Rockefeller University Press, from How to induce protective humoral immunity against Plasmodium falciparum circumsporozoite protein, Wahl I, Wardemann H, volume 219/ edition 2, 2022; permission conveyed through Copyright Clearance Center, Inc.19 (B). The RTS, S vaccine; makes use of complete Pfcsp 3D7 C-terminal domain and 18 NANP repeats.26 It is also combined with HBs Ag. |
The RTS, S Malaria Vaccine
The RTS, S vaccine contains epitopes designed to stimulate both the cell-mediated and humoral adaptive immune systems to produce an optimal immune response against Pf.19 The “R” stands for the central repeat region of Plasmodium (P.) falciparum circumsporozoite protein (CSP); the “T” for the T-cell epitopes of the CSP; and the ‘S’ for hepatitis B surface antigen (HBsAg).27 The molecular structure of the vaccine antigen is made up of 19 NANP central tandem repeats and the N-terminus of the amino acids in the hepatitis B surface antigen attached to the C-terminus of the CSP.28 The RTS, S/As01 vaccine is made of a combination of the RTS, S antigen coupled with the AS01 adjuvant,29 and the RTS, S/AS02 vaccine is made of a combination of the RTS, S antigen coupled with the adjuvant composed of monophosphoryl lipid A and Quillaja Saponaria Molina in an oil-in-water emulsion.4
RTS, S vaccine targets CSP of the Pf trying to block its development,30 during the pre-erythrocytic stage before the parasite invades the liver cells.31,32 However, some recent clinical trials have highlighted much on the efficacy of pre-erythrocytic stage vaccines since blocking the parasite at this stage would probably stop the asymptomatic phase of the disease.33,34 Meanwhile, only a small tilter of malaria parasites can cause disease once they manage to escape the blood-stage immune response, hence creating another challenge.34
Cell-Mediated Immune Responses by RTS, S Vaccine-Induced Antibodies
RTS, S Vaccine-induced antibodies elicit significant immune cellular responses including phagocytosis by monocytes and neutrophils, as well as activation of natural killer (NK) cells for opsonization and antibody-dependent cellular cytotoxicity (ADCC). However, the magnitude of neutrophil phagocytosis by antibodies is only moderate, and responses varied widely amongst children, which may have contributed to the vaccine’s underwhelming effectiveness.35 Interestingly, within a year of immunization, functional antibodies began to drop in substantial part, showing the waning vaccine effectiveness throughout that period. However, neutrophil phagocytosis activity exhibited longer-lasting persistence. Immunity to malaria appears to be significantly influenced by the interaction of IgG with Fc Gamma Receptors (FcRs), which supports phagocytosis, notably by neutrophils, and other cellular activities against sporozoites,35–37 and other factors shown in Table 1, which due to space I will comment on only a few.
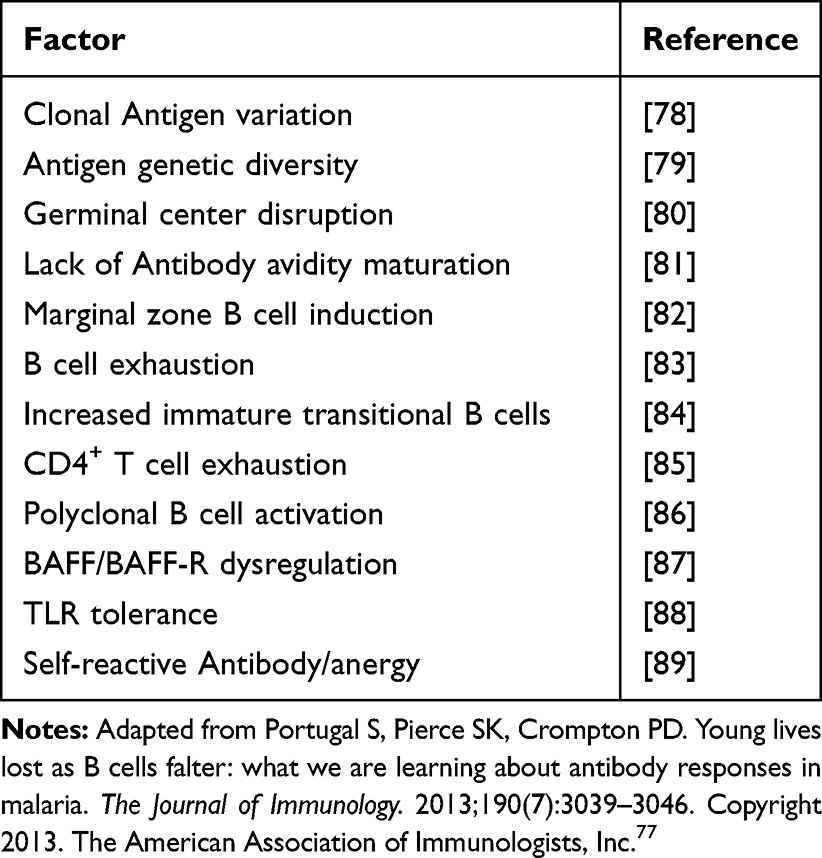 |
Table 1 Potential Factors That Affect the Development of Broadly Effective Antibodies Against P. falciparum by RTS, S Vaccine |
In addition, IgG antibodies also elicit activation of NK cells through signaling of FcγRIIIa that also supports immune response against sporozoites.35,36 Recent data36,38 suggest that monocytes are far less active in phagocytosis than neutrophils, which are also much less abundant in blood, hence hypothesizing that neutrophils could be more important in the elimination of sporozoites in the blood. There is still much to learn about the function of NK cells in RTS, S mediated immunity, including how they may be stimulated to perhaps eliminate sporozoites. Understanding the functions and behavior of liver macrophages (Kupffer cells) as well as neutrophils, monocytes, and NK cells in other organs including the skin may aid in the effective elimination of sporozoites. In mouse models, sporozoites can be immobilized in the skin by antibodies to CSP, which may render them more amenable to removal by dermal phagocytes. Complement fixation, sporozoite traversal inhibition, and hepatocyte invasion inhibition are further possible antibody defense mechanisms.39
Monoclonal Antibodies (moAbs) and the Pfcsp Complexity
A recent study conducted in Kenya investigated the long-term activity of RTS, S/AS01 anti-CSP antibodies,40 where 447 children who were previously immunized during the 2007 phase IIb clinical trial of RTS, S/AS01, were followed up for 7 years. The kinetic activity and levels of IgM, IgG1, IgG2, IgG3, and IgG4 were measured.40 According to the study, RTS, S/AS01 induces high levels of anti-CSP IgG Abs, which decline relatively quickly 6.5 months after primary vaccination, followed by a gradual decay, which is consistent with previous research.41,42 Intriguingly, there were differences in the anti-CSP IgG and IgM kinetics across and within the groups receiving the vaccine and the control group. RTS, S/AS01 vaccination efficiently produced IgM Abs against the NANP region of the CSP. Interestingly, IgM Abs persisted over the course of the seven-year follow-up, showing that natural exposure may induce and then increase the IgM memory response. This view is supported by the fact that the IgM levels of the control group gradually grew from baseline due to natural exposure boosting and caught up with the vaccine group 21 months later.40
The exact mechanism to elicit effective anti-CSP antibody responses as well as the complexity of the CSP region and introduction of effective specific adjuvants remain unclear. A related study provided the first evidence that investigated the possibility of Bacillus subtilis spores to act as possible vaccine carriers and adjuvants, hence eliciting an anti-CSP nasal humoral immune response. The spores were characterized as possessing the possibility of increasing immunogenicity of the Pfcsp, hence inducing high levels of IgG.28 These proofs of concept results provide novel facts to be applied in the design of effective adjuvants of ongoing vaccines.
Monoclonal antibody IgA has also been proven to play an important role in Pfcsp, due to its ability to recognize N-terminus polymorphic region.43 IgA antibodies have a basic role in protecting mucosal areas against pathogens. Meanwhile, sporozoites injected into the skin after a mosquito bite can induce IgA responses at dermal inoculation sites.43 However, the Pf parasite inhibition mechanism caused by antibodies in the skin, blood, and liver remains not understood. Monoclonal antibody CIS43L5 induced by Pfcsp can prevent malaria infection in individuals who had never had malaria infection or were vaccinated.44 A clear understanding of the interaction between antibodies and the CSP of live sporozoites would help to come up with effective moAbs that can elicit an effective immune response.45
Antibody Fine-Specificity with CSP Regions
The C-terminus region acts as a primary determinant of phagocytic activities during the elimination of sporozoites in humans. Serum samples from RTS, S recipients exhibited robust phagocytic activity by terminal specific antibody titres than repeat region antibody titres, hence, marks a significant observation that necessitates additional research into the root cause.46 Several significant functional components, including adhesion motifs for the compliment, thrombospondin, and properdin, are found in the C-terminus of CSP. All Plasmodium species include the properdin-binding sequence, which may affect an infection’s susceptibility.47,48
Specific antibodies against the CSP region are highly protective since the C-terminus has also been linked to the inaugural entry of the sporozoites into hepatocytes.46,49 Studies have demonstrated that the CSP region is essential to malaria parasites that enhance immune escape.46 Before making any generalizations about the significance of C-terminal antibodies in vaccine-induced protection, it is necessary to thoroughly examine their biological activity.
Biophysical and Structural Impact of the Anti-Pfcsp Antibodies
Kinetic and biophysical characteristics of the anti-NANP antibodies show high affinity in an in-vivo mouse model. Anti-NANP responses to other polymorphic secondary motifs such as type 1B and Asn Pseudo 310 uncovers an important tool for the development of an effective vaccine design.50 In contrast, high-affinity antibodies are also characterized by low binding and inhibitory activity to live Pf sporozoites. A monoclonal antibody 5d5 specific to the N-terminus of the Pfcsp was detected with high-affinity binding to an α-helical epitope in N-terminal and hence does not support the use of the epitope in the development of CSP-based vaccines.51
A related study suggested that an antibody that can target all three epitopes in the polymorphic region of the Pfcsp, such as NPDP, NVDP, and NAP, can effectively prevent malaria infection.52 The breadth of RTS, S-induced antibodies against CSP polymorphic regions is relatively associated with fine specificity and hence should be incorporated into the design of current malaria vaccines.53 Anti-Pfcsp antibodies contain an L9 kappa chain that has been suggested to have important structural characteristics that enable its binding ability to NVDP on the Pfcsp, hence eliciting neutralization of the sporozoites. Hence, the presence of relatively greater than two NVDPs would promote rapid cross-linkage and elicit a high antibody response.29
The Role of Complement Fixing Antibodies in RTS, S
The presence of complement factors involved in the final phase of complement activation that form the membrane attack complex indicates that RTS and S-induced antibodies increase activation of the complement cascade (C5b-C9). Antibodies that fix complement specifically target the C-terminal and central repeat regions of CSP. The highest complement-fixing ability is observed in the IgG1 and IgG3 subtypes, whilst IgG2 and IgG4 show minimal or no ability.54 In children aged one to four years, RTS, S/AS02A substantially elicited anti-CSP IgG1 followed by IgG3 and IgG2. An analysis of a cohort (n = 195) of participants from the phase III RTS, S/AS01B vaccine study in young children (6–12 weeks and 5–17 months) revealed similar results, with a predominance of IgG1 and IgG3 responses. It was shown that vaccination-induced immunity was correlated with a larger ratio of cytophilic antibodies (IgG1 and IgG3) to non-cytophilic antibodies (IgG2 and IgG4).55 Notably, human complement interactions, a mechanism specifically related to protection in investigations of naturally acquired immunity to sporozoites in children, maybe one mechanism in which RTS, S-induced cytophilic antibodies may offer protection.56
The Decay of Complement-Fixing Antibodies
IgM antibodies can potentially function as a mediator for complement action. Intriguingly, functional C1q-fixation reactions were only noticeable after RTS, and S vaccination, when IgM and IgG levels had greatly increased, although anti-CSP IgM seropositivity was very mild at baseline.57 IgM is not a prominent mediator of complement-fixing activity, according to a model of antibody degradation over time; IgG1 and IgG3 are instead the main mediators. Although the function of IgM in immunological memory is not well understood, a recent study employing a mouse malaria model showed that IgM memory B cells were long-lived and engaged in secondary responses, and IgM responses persisted even in those with severe malaria exposure.57
Epitope-specificity also affects complement activity.58–60 RTS, S-induced antibodies were shown to be particularly effective in fixing complement to the central repeat region of CSP in animal models; however, antibodies to the C-terminal region may also fix complement. It is interesting to note that individuals with greater complement-fixing antibodies typically had comparable levels of IgG in both CSP areas, whereas individuals with lower functional antibodies were more often epitope-skewed. As a result, antibodies to both regions may more effectively promote the formation of immunological complexes that result in complement fixation and activation. Children’s IgG epitope profiles were often observed to be very varied. Only IgG to the central repeat motif has generally been determined and reported in field studies of RTS, S. This is most likely because RTS, S-induced antibodies to the central repeat region, rather than the C-terminal area, show some correlation with protection in studies of healthy people who have never had malaria.61–63
However, field assessments of children and infants exposed to malaria have inconsistently revealed a correlation between immunity and repeat-specific antibodies.41,64,65 According to a recent study, protection in phase III clinical trial of RTS, S/AS01B was related to antibodies to both regions of CSP that belonged to a certain IgG subclass.55 The central repeat region is also thought to be immunodominant since it is a primary target of antibodies produced by whole, irradiation, attenuated sporozoite vaccines, and naturally occurring malaria exposure, and repeat-specific antibodies can inhibit sporozoite infectivity in vitro.66 However, non-repeat area antibodies can also be developed spontaneously and have been shown to have inhibitory action in vitro.66–69 When considered as a whole, the evidence points to the significance of antibodies to the central repeat region of CSP but also encourages further research into antibody responses to the C-terminal region, especially given that antibodies specific to the repeats alone are a poor correlate of protection.
Functional Antibodies are Poorly Sustained Overtime After RTS, S Vaccination
Most malaria vaccines under development face difficulties due to the gradual loss of vaccine effectiveness.70 Nevertheless, knowledge of the dynamic nature of functional specific antibodies by RTS, S, or any other malaria vaccine is currently inadequate. One major drawback of RTS, S is that it loses most of its efficacy after a year of initial vaccination completion.11 Additionally, this is supported by recent research that discovered that complement-fixing activity by antibodies rapidly decreased following vaccination in young children.35,36,71
Related studies suggest that improving the induction of neutrophil phagocytosis function of antibodies may be a potential future approach for increasing vaccine effectiveness given the evidence of a low decay rate of neutrophil phagocytosis during infection. Maximizing the induction of IgG3 or the ratio of cytophilic to non-cytophilic antibodies, for example through the use of adjuvants; increasing induction of antibodies to key epitopes that mediate phagocytosis; and adding additional epitopes or antigens that promote phagocytosis (for instance, targets of antibodies that promote phagocytosis were identified in the N-terminal sequence of CSP that is not in the RTS, S vaccine construct).36,71
Poor Induction of Functional Antibodies in Areas with Higher Malaria Exposure and Blood-Stage Immunity
In the phase III trial, the high malaria transmission area had substantially greater post-vaccination levels of IgG2 and IgG4 to CSP than the low malaria transmission site, but not IgG1 and IgG3.55 Notably, RTS, S was shown to mostly generate IgG1 and IgG2 in studies of individuals who had never had malaria,62,72,73 hence supporting the idea that immunogenicity can significantly vary between those that have been exposed to the disease and those who have never had malaria. Even though IgG3 is a powerful modulator of complement fixation and activation, these distinctions are poorly understood and call for more research. Interestingly, a recent related study found that children (ages one to five) responded in a highly diverse manner, and it was essential to note that children who had higher malaria exposure also had much significantly poorer antibody activity. In some circumstances, addressing this could be essential to achieving better effectiveness of the RTS, S malaria vaccine. Implications that recurrent malaria exposure can have on B cells, CD4+ T cells, and innate immune cell phenotypes and activities that drive antibody production may lead to reduced vaccination responses.74
On the contrary, higher malaria exposure may be anticipated to result in CSP antibodies, enhance responses, and accelerate antibody affinity maturation, which may affect functional activity. Vaccine responses may potentially be impacted by aging-related immune system changes. To maximize vaccination responses in young infants, further research is required to understand these processes and how they might be handled. The effectiveness and responsiveness to vaccines may be increased by using malaria preventive medication concurrently with vaccine delivery.75 Additionally, host genetics may affect vaccination responses and human leukocyte antigen genotypes were related to RTS and S vaccine effectiveness against experimental infection in people without malaria exposure.76 High complement-fixing activity and possibly greater effectiveness are anticipated to result from the use of adjuvants or immunization regimens that decrease IgG2 induction and elevate IgG3.
Conclusions and Future Recommendations
RTS, S malaria vaccine effectiveness shows some limited success and promising results regardless of its 58% efficacy for severe disease. Antibody specificity has been a well-recognized problem in the malaria vaccine research community. The induction of long-lived and high antibody responses against the CSP of sporozoites stands as a major landmark towards the development of a broadly effective vaccine. However, RTS, S has been characterized by high but short-lived anti-CSP antibodies. A large and growing body of literature still expresses concerns over the complexity of CSP that highly impacts the specificity and affinity in many good studies of monoclonal antibodies, thereby increasing the difficulty of effective malaria vaccine design. More recent data has proposed the use of different kinds of monoclonal antibodies, incorporating biophysical mechanisms of the target antibody, considering the anti-repeat and minor repeat regions, and complement-fixing antibodies on the CSP.
Alternatively, changing adjuvants has also been proposed as it can elicit high immunogenicity of the CSP, hence inducing rapid antibody response. Ideally, understanding the functional mechanisms as well as the relationship between CSP and the anti-CSP antibody specificity will then put effort into the identification of alternative structural mechanisms of the CSP, development of effective adjuvants through combination with immunogens and putting into consideration the frequency of dose administration, hence achieving effective response. Related studies have demonstrated that RTS, S generates complement-fixing antibodies in both children and adults, which has marked an important landmark in the development of efficacious vaccines. Monoclonal antibodies obtained from individuals who had received the RTS, S vaccine did exhibit invasion inhibitory action, indicating another important mechanism toward control of malaria parasites in humans. Hence, this review supports future investigation of large phase III trials to determine whether novel techniques including complement fixation are valuable correlates of protective immunity and advance the development of more efficacious and long-lasting vaccines.
Author Contributions
The author made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The author has declared that no competing interests exist.
References
1. World Health Organization. World malaria report 2022. Available from: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2022.
2. Manescu P, Shaw MJ, Elmi M, et al. Expert-level automated malaria diagnosis on routine blood films with deep neural networks. Am J Hematol. 2020;95(8):883–891. doi:10.1002/AJH.25827
3. Laurens MB. RTS,S/AS01 vaccine (MosquirixTM): an overview. Hum Vaccin Immunother. 2020;16(3):480–489. doi:10.1080/21645515.2019.1669415
4. Elimination T malERA RCP on T for M. malERA: an updated research agenda for diagnostics, drugs, vaccines, and vector control in malaria elimination and eradication. PLOS Med. 2017;14(11):e1002455. doi:10.1371/JOURNAL.PMED.1002455
5. Peterson I, Kapito-Tembo A, Bauleni A, et al. Overdiagnosis of malaria illness in an endemic setting: a facility-based surveillance study in Malawi. Am J Trop Med Hyg. 2021;104(6):2123–2130. doi:10.4269/ajtmh.20-1209
6. Scally SW, Murugan R, Bosch A, et al. Rare PfCSP C-terminal antibodies induced by live sporozoite vaccination are ineffective against malaria infection. J Exp Med. 2018;215(1):63–75. doi:10.1084/JEM.20170869
7. Cha S-J, Kim M-S, Na CH, Jacobs-Lorena M. Plasmodium sporozoite phospholipid scramblase interacts with mammalian carbamoyl-phosphate synthetase 1 to infect hepatocytes. Nat Commun. 2021;12(1). doi:10.1038/S41467-021-27109-7
8. Langowski MD, Khan FA, Savransky S, et al. Restricted valency (NPNA)n repeats and junctional epitope-based circumsporozoite protein vaccines against Plasmodium falciparum. Npj Vaccines. 2022;7(1):13. doi:10.1038/S41541-022-00430-Y
9. Uyar Y, Akşit A, Karaca S, Ceylan ŞS, Yürük M. 37 NANP ve 4 NVDP Epitoplu Plasmodium falciparum Rekombinant Circumsporozoite Proteinin Ekspresyonu ve Moleküler Karakterizasyonu [Production and molecular characterization of Plasmodium falciparum recombinant circumsporozoite protein with 37 NANP and 4 NVDP epitopes]. Mikrobiyol Bul. 2017;51(1):41–51. Turkish. doi:10.5578/mb.46549
10. RTS, S Clinical Trials Partnership. A Phase 3 trial of RTS,S/AS01 malaria vaccine in African Infants. N Engl J Med. 2012;367(24):2284–2295. doi:10.1056/NEJMOA1208394
11. First Results of Phase. A Phase 3 trial of RTS,S/AS01 malaria vaccine in African Children. N Engl J Med. 2011;365(20):1863–1875. doi:10.1056/nejmoa1102287
12. Malaria: phase 3 trial results for vaccine RTS,S/AS01. Available from: https://www.who.int/news-room/questions-and-answers/item/phase-3-trial-results-for-malaria-vaccine-rtss-as01.
13. Zhang YL, Pan WQ. The world’s first malaria vaccine: hope and challenge. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2022;33(6):560–562. doi:10.16250/j.32.1374.2021283
14. Arora N, Anbalagan L, Pannu AK. Towards eradication of malaria: is the WHO’s RTS,S/AS01 vaccination effective enough? Risk Manag Healthc Policy. 2021;14:1033–1039. doi:10.2147/RMHP.S219294
15. Kurtovic L, Wetzel D, Reiling L, et al. Novel virus-like particle vaccine encoding the circumsporozoite protein of plasmodium falciparum is immunogenic and induces functional antibody responses in mice. Front Immunol. 2021;12. doi:10.3389/FIMMU.2021.641421
16. McCann RS, Kabaghe AN, Moraga P, et al. The effect of community-driven larval source management and house improvement on malaria transmission when added to the standard malaria control strategies in Malawi: a cluster-randomized controlled trial. Malar J. 2021;20(1). doi:10.1186/s12936-021-03769-0
17. Vareta J, Buchwald AG, Barrall A, et al. Submicroscopic malaria infection is not associated with fever in cross-sectional studies in Malawi. Malar J. 2020;19(1). doi:10.1186/s12936-020-03296-4
18. Oseno B, Marura F, Ogwang R, et al. Characterization of Anopheles gambiae D7 salivary proteins as markers of human–mosquito bite contact. Parasit Vectors. 2022;15(1). doi:10.1186/s13071-021-05130-5
19. Wahl I, Wardemann H. How to induce protective humoral immunity against Plasmodium falciparum circumsporozoite protein. J Exp Med. 2022;219(2). doi:10.1084/jem.20201313
20. Reiling L, Boyle MJ, White MT, et al. Targets of complement-fixing antibodies in protective immunity against malaria in children. Nat Commun. 2019;10(1). doi:10.1038/s41467-019-08528-z
21. Leitner WW, Haraway M, Pierson T, Bergmann-Leitner ES. Role of opsonophagocytosis in immune protection against malaria. Vaccines. 2020;8(2):264. doi:10.3390/vaccines8020264
22. Langowski MD, Khan FA, Bitzer AA, et al. Optimization of a Plasmodium falciparum circumsporozoite protein repeat vaccine using the tobacco mosaic virus platform. Proc Natl Acad Sci U S A. 2020;117(6):3114–3122. doi:10.1073/pnas.1911792117
23. Kisalu NK, Idris AH, Weidle C, et al. A human monoclonal antibody prevents malaria infection by targeting a new site of vulnerability on the parasite. Nat Med. 2018;24(4):408–416. doi:10.1038/nm.4512
24. Jelínková L, Jhun H, Eaton A, Petrovsky N, Zavala F, Chackerian B. An epitope-based malaria vaccine targeting the junctional region of circumsporozoite protein. Npj Vaccines. 2021;6(1). doi:10.1038/S41541-020-00274-4
25. Francica JR, Shi W, Chuang GY, et al. Design of alphavirus virus-like particles presenting circumsporozoite junctional epitopes that elicit protection against malaria. Vaccines. 2021;9(3):272. doi:10.3390/VACCINES9030272
26. Herrera R, Anderson C, Kumar K, et al. Reversible conformational change in the plasmodium falciparum circumsporozoite protein masks Its adhesion domains. Infect Immun. 2015;83(10):3771–3780. doi:10.1128/IAI.02676-14
27. PATH’s malaria vaccine initiative. Available from: https://www.malariavaccine.org/files/content/page/files/RTSS.FAQs_FINAL.pdf.
28. de Almeida MEM, Alves KCS, de Vasconcelos MGS, et al. Bacillus subtilis spores as delivery system for nasal Plasmodium falciparum circumsporozoite surface protein immunization in a murine model. Sci Rep. 2022;12(1):1–8. doi:10.1038/s41598-022-05344-2
29. Wang LT, Hurlburt NK, Schön A, et al. The light chain of the L9 antibody is critical for binding circumsporozoite protein minor repeats and preventing malaria. Cell Rep. 2022;38(7):110367. doi:10.1016/j.celrep.2022.110367
30. White NJ. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J. 2011;10(1):297. doi:10.1186/1475-2875-10-297
31. Calvo-Calle JM, Mitchell R, Altszuler R, Othoro C, Nardin E. Identification of a neutralizing epitope within minor repeat region of Plasmodium falciparum CS protein. Npj Vaccines. 2021;6(1):1–8. doi:10.1038/s41541-020-00272-6
32. McNamara HA, Idris AH, Sutton HJ, et al. Antibody feedback limits the expansion of B cell responses to malaria vaccination but drives diversification of the humoral response. Cell Host Microbe. 2020;28(4):572–585.e7. doi:10.1016/j.chom.2020.07.001
33. McCall MBB, Kremsner PG, Mordmüller B. Correlating efficacy and immunogenicity in malaria vaccine trials. Semin Immunol. 2018;39:52–64. doi:10.1016/j.smim.2018.08.002
34. Jensen AR, Adams Y, Hviid L. Cerebral Plasmodium falciparum malaria: the role of PfEMP1 in its pathogenesis and immunity, and PfEMP1-based vaccines to prevent it. Immunol Rev. 2020;293(1):230–252. doi:10.1111/imr.12807
35. Suscovich TJ, Fallon JK, Das J, et al. Mapping functional humoral correlates of protection against malaria challenge following RTS,S/AS01 vaccination. Sci Transl Med. 2020;12(553). doi:10.1126/SCITRANSLMED.ABB4757
36. Feng G, Wines BD, Kurtovic L, et al. Mechanisms and targets of Fcγ-receptor mediated immunity to malaria sporozoites. Nat Commun. 2021;12(1). doi:10.1038/S41467-021-21998-4
37. Kurtovic L, Atre T, Feng G, et al. Multifunctional antibodies are induced by the RTS,S malaria vaccine and associated with protection in a phase 1/2a trial. J Infect Dis. 2021;224(7):1128–1138. doi:10.1093/INFDIS/JIAA144
38. Garcia-Senosiain A, Kana IH, Singh S, et al. Neutrophils dominate in opsonic phagocytosis of P. falciparum blood-stage merozoites and protect against febrile malaria. Commun Biol. 2021;4(1). doi:10.1038/S42003-021-02511-5
39. Opi DH, Kurtovic L, Chan JA, Horton JL, Feng G, Beeson JG. Multi-functional antibody profiling for malaria vaccine development and evaluation. Expert Rev Vaccines. 2021;20(10):1257–1272. doi:10.1080/14760584.2021.1981864
40. Mugo RM, Mwai K, Mwacharo J, et al. Seven-year kinetics of RTS, S/AS01-induced anti-CSP antibodies in young Kenyan children. Malar J. 2021;20(1):1–8. doi:10.1186/s12936-021-03961-2
41. Bejon P, Lusingu J, Olotu A, et al. Efficacy of RTS,S/AS01E vaccine against malaria in children 5 to 17 months of age. N Engl J Med. 2008;359(24):2521. doi:10.1056/NEJMOA0807381
42. Olotu A, Fegan G, Wambua J, et al. Four-year efficacy of RTS,S/AS01E and its interaction with malaria exposure. N Engl J Med. 2013;368(12):1111–1120. doi:10.1056/NEJMOA1207564/SUPPL_FILE/NEJMOA1207564_DISCLOSURES.PDF
43. Tan J, Cho H, Pholcharee T, et al. Functional human IgA targets a conserved site on malaria sporozoites. Sci Transl Med. 2021;13(599). doi:10.1126/scitranslmed.abg2344
44. Gaudinski MR, Berkowitz NM, Idris AH, et al. A monoclonal antibody for malaria prevention. N Engl J Med. 2021;385(9):803–814. doi:10.1056/NEJMOA2034031
45. Kurtovic L, Drew DR, Dent AE, Kazura JW, Beeson JG. Antibody targets and properties for complement-fixation against the circumsporozoite protein in malaria immunity. Front Immunol. 2021;12:1–12. doi:10.3389/fimmu.2021.775659
46. Bergmann-Leitner ES, Scheiblhofer S, Weiss R, et al. C3d binding to the circumsporozoite protein carboxy-terminus deviates immunity against malaria. Int Immunol. 2005;17(3):245–255. doi:10.1093/INTIMM/DXH205
47. Schofield L, Uadia P. Lack of Ir gene control in the immune response to malaria. I. A thymus-independent antibody response to the repetitive surface protein of sporozoites. J Immunol. 1990;144(7):2781–2788. doi:10.4049/jimmunol.144.7.2781
48. Goundis D, Reid KBM. Properdin, the terminal complement components, thrombospondin and the circumsporozoite protein of malaria parasites contain similar sequence motifs. Nature. 1988;335(6185):82–85. doi:10.1038/335082A0
49. Meraldi V, Audran R, Romero JF, et al. OM-174, a new adjuvant with a potential for human use, induces a protective response when administered with the synthetic C-terminal fragment 242-310 from the circumsporozoite protein of Plasmodium berghei. Vaccine. 2003;21(19–20):2485–2491. doi:10.1016/S0264-410X(03)00093-8
50. Pholcharee T, Oyen D, Flores-Garcia Y, et al. Structural and biophysical correlation of anti-NANP antibodies with in vivo protection against P. falciparum. Nat Commun. 2021;12(1). doi:10.1038/s41467-021-21221-4
51. Thai E, Costa G, Weyrich A, et al. A high-affinity antibody against the CSP N-terminal domain lacks Plasmodium falciparum inhibitory activity. J Exp Med. 2020;217(11). doi:10.1084/JEM.20200061
52. Flores-Garcia Y, Wang LT, Park M, et al. The P. falciparum CSP repeat region contains three distinct epitopes required for protection by antibodies in vivo. PLoS Pathog. 2021;17(11):1–15. doi:10.1371/journal.ppat.1010042
53. Chaudhury S, MacGill RS, Early AM, et al. Breadth of humoral immune responses to the C-terminus of the circumsporozoite protein is associated with protective efficacy induced by the RTS,S malaria vaccine. Vaccine. 2021;39(6):968–975. doi:10.1016/J.VACCINE.2020.12.055
54. Irani V, Guy AJ, Andrew D, Beeson JG, Ramsland PA, Richards JS. Molecular properties of human IgG subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases. Mol Immunol. 2015;67(2Pt A):171–182. doi:10.1016/J.MOLIMM.2015.03.255
55. Ubillos I, Ayestaran A, Nhabomba AJ, et al. Baseline exposure, antibody subclass, and hepatitis B response differentially affect malaria protective immunity following RTS,S/AS01E vaccination in African children. BMC Med. 2018;16(1). doi:10.1186/S12916-018-1186-4
56. Kurtovic L, Behet MC, Feng G, et al. Human antibodies activate complement against Plasmodium falciparum sporozoites, and are associated with protection against malaria in children. BMC Med. 2018;16(1). doi:10.1186/S12916-018-1054-2
57. Stanisic DI, Fowkes FJI, Koinari M, et al. Acquisition of antibodies against Plasmodium falciparum merozoites and malaria immunity in young children and the influence of age, force of infection, and magnitude of response. Infect Immun. 2015;83(2):646–660. doi:10.1128/IAI.02398-14
58. Diebolder CA, Beurskens FJ, De Jong RN, et al. Complement is activated by IgG hexamers assembled at the cell surface. Science. 2014;343(6176):1260–1263. doi:10.1126/SCIENCE.1248943
59. Cragg MS, Morgan SM, Chan HTC, et al. Complement-mediated lysis by anti-CD20 mAb correlates with segregation into lipid rafts. Blood. 2003;101(3):1045–1052. doi:10.1182/BLOOD-2002-06-1761
60. Teeling JL, Mackus WJM, Wiegman LJJM, et al. The biological activity of human CD20 monoclonal antibodies is linked to unique epitopes on CD20. J Immunol. 2006;177(1):362–371. doi:10.4049/JIMMUNOL.177.1.362
61. Chaudhury S, Ockenhouse CF, Regules JA, et al. The biological function of antibodies induced by the RTS,S/AS01 malaria vaccine candidate is determined by their fine specificity. Malar J. 2016;15(1):1–12. doi:10.1186/S12936-016-1348-9/FIGURES/8
62. Kester KE, McKinney DA, Tornieporth N, et al. A phase I/IIa safety, immunogenicity, and efficacy bridging randomized study of a two-dose regimen of liquid and lyophilized formulations of the candidate malaria vaccine RTS,S/AS02A in malaria-naïve adults. Vaccine. 2007;25(29):5359–5366. doi:10.1016/J.VACCINE.2007.05.005
63. Kester KE, Cummings JF, Ofori-Anyinam O, et al. Randomized, double-blind, phase 2a trial of falciparum malaria vaccines RTS,S/AS01B and RTS,S/AS02A in malaria-naive adults: safety, efficacy, and immunologic associates of protection. J Infect Dis. 2009;200(3):337–346. doi:10.1086/600120
64. Alonso PL, Sacarlal J, Aponte JJ, et al. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomised controlled trial. Lancet. 2004;364(9443):1411–1420. doi:10.1016/S0140-6736(04)17223-1
65. Agnandji ST, Lell B, Fernandes JF, et al. Efficacy and safety of the RTS,S/AS01 malaria vaccine during 18 months after vaccination: a phase 3 randomized, controlled trial in children and young infants at 11 African sites. PLOS Med. 2014;11(7):e1001685. doi:10.1371/JOURNAL.PMED.1001685
66. Hollingdale MR, Nardin EH, Tharavanij S, Schwartz AL, Nussenzweig RS. Inhibition of entry of Plasmodium falciparum and P. vivax sporozoites into cultured cells; an in vitro assay of protective antibodies. J Immunol. 1984;132(2):909–913. doi:10.4049/jimmunol.132.2.909
67. Chatterjee S, Wery M, Sharma P. A conserved peptide sequence of the Plasmodium falciparum circumsporozoite protein and antipeptide antibodies inhibit Plasmodium berghei sporozoite invasion of Hep-G2 cells and protect immunized mice against P. berghei sporozoite challenge. Infect Immun. 1995;63(11). doi:10.1128/IAI.63.11.4375-4381.1995
68. Rathore D, Nagarkatti R, Jani D, et al. An immunologically cryptic epitope of Plasmodium falciparum circumsporozoite protein facilitates liver cell recognition and induces protective antibodies that block liver cell invasion. J Biol Chem. 2005;280(21):20524–20529. doi:10.1074/JBC.M414254200
69. Oyen D, Torres JL, Wille-Reece U, et al. Structural basis for antibody recognition of the NANP repeats in Plasmodium falciparum circumsporozoite protein. Proc Natl Acad Sci U S A. 2017;114(48):E10438–E10445. doi:10.1073/PNAS.1715812114
70. Beeson JG, Kurtovic L, Dobaño C, et al. Challenges and strategies for developing efficacious and long-lasting malaria vaccines. Sci Transl Med. 2019;11(474). doi:10.1126/SCITRANSLMED.AAU1458
71. Kurtovic L, Agius PA, Feng G, et al. Induction and decay of functional complement-fixing antibodies by the RTS,S malaria vaccine in children, and a negative impact of malaria exposure. BMC Med. 2019;17(1). doi:10.1186/S12916-019-1277-X
72. Kester KE, McKinney DA, Tornieporth N, et al. Efficacy of recombinant circumsporozoite protein vaccine regimens against experimental Plasmodium falciparum malaria. J Infect Dis. 2001;183(4):640–647. doi:10.1086/318534
73. Stoute JA, Slaoui M, Heppner DG, et al.; RTS,S Malaria Vaccine Evaluation Group. A preliminary evaluation of a recombinant circumsporozoite protein vaccine against Plasmodium falciparum malaria. N Engl J Med. 1997;336(2):86–91. doi:10.1056/NEJM199701093360202
74. Illingworth J, Butler NS, Roetynck S, et al. Chronic exposure to Plasmodium falciparum is associated with phenotypic evidence of B and T cell exhaustion. J Immunol. 2013;190(3):1038–1047. doi:10.4049/JIMMUNOL.1202438
75. Chandramohan D, Zongo I, Sagara I, et al. Seasonal malaria vaccination with or without seasonal malaria chemoprevention. N Engl J Med. 2021;385(11):1005–1017. doi:10.1056/NEJMOA2026330
76. Nielsen CM, Vekemans J, Lievens M, Kester KE, Regules JA, Ockenhouse CF. RTS,S malaria vaccine efficacy and immunogenicity during Plasmodium falciparum challenge is associated with HLA genotype. Vaccine. 2018;36(12):1637–1642. doi:10.1016/J.VACCINE.2018.01.069
77. Portugal S, Pierce SK, Crompton PD. Young lives lost as B cells falter: what we are learning about antibody responses in malaria. J Immunol. 2013;190(7):3039–3046. doi:10.4049/JIMMUNOL.1203067
78. Scherf A, Lopez-Rubio JJ, Riviere L. Antigenic variation in Plasmodium falciparum. Annu Rev Microbiol. 2008;62:445–470. doi:10.1146/ANNUREV.MICRO.61.080706.093134
79. Takala SL, Plowe CV. Genetic diversity and malaria vaccine design, testing and efficacy: preventing and overcoming “vaccine resistant malaria”. Parasite Immunol. 2009;31(9):560–573. doi:10.1111/J.1365-3024.2009.01138.X
80. Urban BC, Hien TT, Day NP, et al. Fatal Plasmodium falciparum malaria causes specific patterns of splenic architectural disorganization. Infect Immun. 2005;73(4):1986–1994. doi:10.1128/IAI.73.4.1986-1994.2005
81. Ibison F, Olotu A, Muema DM, et al. Lack of avidity maturation of merozoite antigen-specific antibodies with increasing exposure to Plasmodium falciparum amongst children and adults exposed to endemic malaria in Kenya. PLoS One. 2012;7(12):e52939. doi:10.1371/JOURNAL.PONE.0052939
82. Weill JC, Weller S, Reynaud CA. Human marginal zone B cells. Annu Rev Immunol. 2009;27:267–285. doi:10.1146/ANNUREV.IMMUNOL.021908.132607
83. Weiss GE, Crompton PD, Li S, et al. Atypical memory B cells are greatly expanded in individuals living in a malaria-endemic area. J Immunol. 2009;183(3):2176–2182. doi:10.4049/JIMMUNOL.0901297
84. Asito AS, Moormann AM, Kiprotich C, Ng’ang’A ZW, Ploutz-Snyder R, Rochford R. Alterations on peripheral B cell subsets following an acute uncomplicated clinical malaria infection in children. Malar J. 2008;7. doi:10.1186/1475-2875-7-238
85. Butler NS, Moebius J, Pewe LL, et al. Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection. Nat Immunol. 2011;13(2):188–195. doi:10.1038/ni.2180
86. Donati D, Mok B, Chêne A, et al. Increased B cell survival and preferential activation of the memory compartment by a malaria polyclonal B cell activator. J Immunol. 2006;177(5):3035–3044. doi:10.4049/JIMMUNOL.177.5.3035
87. Nduati E, Gwela A, Karanja H, et al. The plasma concentration of the B cell activating factor is increased in children with acute malaria. J Infect Dis. 2011;204(6):962–970. doi:10.1093/INFDIS/JIR438
88. Traore B, Koné Y, Doumbo S, et al. The TLR9 agonist CpG fails to enhance the acquisition of Plasmodium falciparum-specific memory B cells in semi-immune adults in Mali. Vaccine. 2009;27(52):7299–7303. doi:10.1016/J.VACCINE.2009.08.023
89. Daniel-Ribeiro CT, Zanini G. Autoimmunity and malaria: what are they doing together? Acta Trop. 2000;76(3):205–221. doi:10.1016/S0001-706X(00)00099-1
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.