")
Back to Journals » OncoTargets and Therapy » Volume 13
HNF1A-Induced lncRNA HCG18 Facilitates Gastric Cancer Progression by Upregulating DNAJB12 via miR-152-3p
Authors Ma P, Li L, Liu F, Zhao Q
Received 11 March 2020
Accepted for publication 11 June 2020
Published 3 August 2020 Volume 2020:13 Pages 7641—7652
DOI https://doi.org/10.2147/OTT.S253391
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
Pei Ma,1 Lianhai Li,1 Fu Liu,1 Qi Zhao2
1Department of General Surgery, Nanyang First People’s Hospital, Nanyang City, Henan Province, People’s Republic of China; 2Department of Urological Surgery, Nanyang First People’s Hospital, Nanyang City, Henan Province, People’s Republic of China
Correspondence: Qi Zhao
Department of Urological Surgery, Nanyang First People’s Hospital, No. 12, Renmin Road, Nanyang City, Henan Province 473010, People’s Republic of China
Email [email protected]
Background: The aberrant expression of long non-coding RNAs (lncRNAs) plays a pivotal role in the development and progression of multiple cancers, including gastric cancer (GC). However, the underlying molecular mechanisms of lncRNA HCG18 in GC remain unknown.
Materials and Methods: The expression levels of HCG18, HNF1A, microRNA-152-3p (miR-152-3p), and DNAJB12 were determined by RT-qPCR. Cell viability, migration, and invasion were assessed by CCK-8, wound healing, and transwell assays, respectively. The interaction between miR-152-3p and HCG18 or DNAJB12 was predicted by bioinformatics analysis and verified by dual-luciferase reporter assay. The correlation between the gene expression levels was analyzed using Pearson’s correlation coefficient. Western blot was used to measure the levels of HNF1A, DNAJB12, epithelial-mesenchymal transition (EMT) proteins (E-cadherin and Vimentin), and proliferation-related protein (PCNA).
Results: It was found that HCG18 was upregulated in GC tissues and cell lines, and knockdown of HCG18 inhibited the proliferation, migration, and invasion of GC cells. Patients with high HCG18 expression had a shorter overall survival time compared with those with low HCG18 expression. In addition, transcription factor HNF1A could bind to the HCG18 promoter to facilitate its transcription. The upregulation of HCG18 could abolish the inhibitory effect of miR-152-3p overexpression on GC cell progression. Furthermore, DNAJB12 was demonstrated to be a target gene of miR-152-3p in GC cells, and HCG18 enhanced DNAJB12 expression by competitively binding with miR-152-3p. Finally, rescue assays proved that overexpression of DNAJB12 partially restored HCG18 knockdown-attenuated progression of GC cells.
Conclusion: Our results demonstrated that HNF1A-induced HCG18 overexpression promoted GC progression by competitively binding with miR-152-3p and upregulating DNAJB12 expression. These findings might provide potential treatment strategies for patients with GC.
Keywords: HNF1A, HCG18, miR-152-3p, DNAJB12, gastric cancer
Introduction
Gastric cancer (GC) is one of the most common malignancies, which ranks the second leading cause of cancer-related death worldwide.1,2 Several factors, such as smoking and atrophic gastritis, are related to the incidence of GC.3 Despite great advances have been made in the treatment of GC, the 5-year survival rate for GC patients is still low due to distant metastasis and high recurrence rate.4,5 Therefore, it is urgent to improve the understanding of GC pathogenesis and develop novel therapeutics for the treatment of GC.
Long non-coding RNAs (lncRNAs) are a class of RNA transcripts longer than 200 nucleotides in lengths, which have no protein-coding ability.6,7 Accumulating evidence indicated that the dysregulation of lncRNAs was involved in the occurrence and development of various types of cancers. For example, Zheng et al indicated that the upregulation of lncRNA HULC predicted a poor prognosis and promoted prostate cancer progression.8 Zhang et al demonstrated that lncRNA PICART1 inhibited the progression of non-small cell lung cancer cells through the AKT1 signaling pathway.9 HCG18 was reported to be associated with the tumorigenesis of glioma and bladder cancer.10,11 However, the specific mechanisms of HCG18 in GC remain unclear.
MicroRNAs (miRNAs) are another type of endogenous non-coding RNAs with a length of 22–25 nucleotides, which regulate gene expression by complementary binding or complex mechanisms.12 miRNAs have been reported to play vital roles in cell proliferation, apoptosis and metastasis in human cancers. For example, miR-338-3p suppressed prostate cancer cell proliferation, migration, and invasion via targeting RAB23.13 miR-203a-3p facilitated the proliferation and migration of colorectal cancer cells by regulating PDE4D.14 You et al found that miR-152-3p/miR-152-5p was lowly expressed in GC tissues and cell lines, and miR-152-5p inhibited cell proliferation and promoted apoptosis of GC cells by downregulating PIK3CA.15 Nevertheless, whether miR-152-3p is involved in GC remains to be further elucidated.
The present study aimed to determine the potential mechanisms of HCG18 in GC. The data of the present study demonstrated for the first time that HNF1A-induced HCG18 overexpression facilitated GC progression by sponging miR-152-3p to upregulate DNAJB12 expression. These findings may provide novel insights into the progression of GC and help to develop novel therapeutics for the treatment of GC.
Materials and Methods
Tissue Collection
A total of 26 pairs of GC tissues and adjacent normal tissues were collected from patients in Nanyang First People’s Hospital. The present study was approved by the Ethics Committee of Nanyang First People’s Hospital and written informed consent was obtained from all patients. All samples were frozen in liquid nitrogen and stored at −80 °C for further analysis. The clinicopathologic features of patients were presented in Table 1.
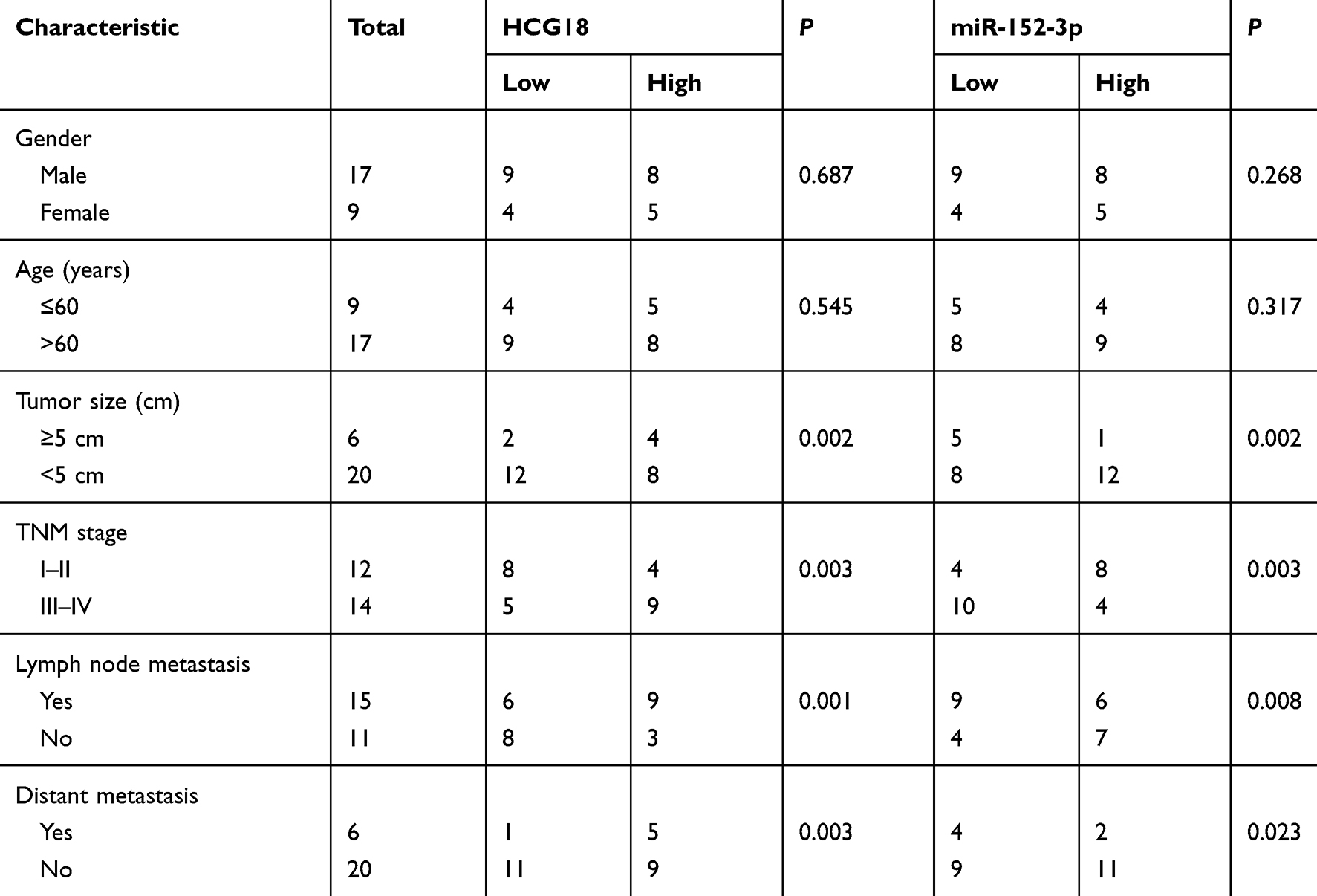 |
Table 1 Correlation Between HCG18 or miR-152-3p Expression and Clinicopathologic Features of GC Patients |
Cell Culture
Gastric cancer cell lines (MKN45, AGS, SCH and SNU638) and human gastric mucosa cell (GES-1) were purchased from Cell Bank of the Chinese Academy of Medical Science (Shanghai, China). The cell lines were cultured in RPMI-1640 medium and supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific) and 1% penicillin-streptomycin (Thermo Fisher Scientific). All cells were maintained at 37 °C in a humidified atmosphere with 5% CO2.
Cell Transfection
The short hairpin RNA (shRNAs) targeting HCG18 (sh-HCG18; 5′-UUGGCUUCAGUCCUGUUCAUCAG-3′) and HNF1A (sh-HNF1A; 5′-AGACUGCAGAAGUACCCUCAA-3′) with negative control (sh-NC; 5′- AAUUCUCCGAACGUGUCACGU-3′), miR-152-3p mimics (5ʹ-UCAGUGCAUGACAGAACUUGG-3ʹ) with negative control (NC mimics; 5ʹ-GGAACUUAGCCACUGUGAAUU-3ʹ) and miR-152-3p inhibitor (5ʹ-UCGCUUGGUGCAGGUCGGGAA-3ʹ) with negative control (NC inhibitor; 5ʹ-UCGCUUGGUGCAGGUCGGGAA-3ʹ) were synthesized by GenePharma (Shanghai, China). The full length of HCG18 or DNAJB12 was subcloned into pcDNA3.1 (GenePharma, Shanghai) to overexpress HCG18 or DNAJB12 levels with empty pcDNA3.1 serving as the control. The transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, USA).
RT-qPCR
Total RNA was extracted from GC tissues and cells using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer’s instruction. RNA concentrations were measured using a NanoDrop™ 2000 spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The RNA was reverse transcribed into cDNA using the Reverse Transcription kit (Takara Biotechnology Ltd., Dalian, China). Subsequently, PCR amplifications were performed using SYBR® Green PCR Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) The primer sequences were as follows: HCG18 forward, 5′-ATCCTGCCAATAGATGCTGCTCAC-3ʹ and reverse, 5′-AGCCACCTTGGTCTCCAGTCTC-3′; HNF1A forward, 5ʹ-CCCAGCA GATCCTGTTGTTCCAG-3ʹ and reverse, 5ʹ-GTGCCGG AAGGCTT CTTCTT-3ʹ; miR-152-3p forward, 5ʹ-TCGGCAGGTCAGTGCATGACAGAA-3ʹ and reverse, 5ʹ-CTCAACTGGTGTCGTGGA-3ʹ; DNAJB12 forward, 5′-TTCCCTTCTAGTAACGTCCACG-3′ and reverse, 5′-GCTGAGAGCTGACACGAGAAT-3′); GAPDH forward, 5′-TGACGTGCCGCCTGGAGAAAC-3ʹ and reverse, 5′-CCGGCATCGAAGGTGGAAGAG-3′; U6 forward, 5ʹ-GAGGGCCTATTTCCCATGATT-3ʹ and reverse, 5ʹ-TAATTAGAATTAATTTGACT-3ʹ. The relative expression of genes was computed using the 2−∆∆Cq method and normalized to GAPDH and U6, the endogenous controls.
CCK-8 Assay
Cell proliferation was determined by Cell Counting Kit-8 (CCK-8, Beyotime Biotechnology, Shanghai, China) assay. Briefly, GC cells (1×104 cells/well) were seeded in 96-well plates. After being incubated for 0, 24, 48, and 72 h, 10 μL of CCK-8 solution (Dojindo, Tokyo, Japan) was added to each well and incubated at 37°C for 2 h. The absorbance at 450 nm was determined through a microplate reader (Bio-Tek, Winooski, VT).
Wound Healing Assay
The transfected GC cells were plated into 6-well plates and cultured until 80% confluence for the wound healing assay. Subsequently, a 200-μL pipette tip was used to generate a single wound in the cell monolayer. Images of the migrating cells were acquired at 0 and 24 h using a microscope (DFC500; Leica, Wetzlar, Germany).
Transwell Assay
The invasive ability of the GC cells was analyzed using Transwell chambers (8.0-μm pore size; EMD Millipore) and Matrigel (Corning Inc.). Cells (8×104 cells/well) were plated in the upper chambers of the Transwell plates in serum-free medium. Transwell membranes were pre-coated with Matrigel. A volume of 600 µL DMEM, supplemented with 10% FBS was plated in the lower chambers. Following incubation for 48 h, the non-invasive cells in the upper chamber were removed and invasive cells in the lower chamber were fixed in 4% paraformaldehyde and stained with 0.1% crystal violet (Beyotime Institute of Biotechnology). Finally, invaded cells were counted under an inverted microscope (ZEISS, Jena, Germany) and photographed.
Luciferase Reporter Assay
Luciferase reporter assay was performed to verify the binding ability between genes. For the binding ability between HNF1A and HCG18 promoter region, wild-type (5ʹ-ATATTAAC-3ʹ) and mutant (5ʹ-TATAATTG-3ʹ) sequences of HCG18 promoter regions were inserted into pGL3 vectors. The pGL3 vectors were then transfected into MKN45 cells with sh-HNF1A or sh-NC. For the binding ability between HCG18 (or DNAJB12) and miR-152-3p, wild-type and mutant sequences of HCG18 (or DNAJB12) were cloned into pmirGLO vectors. These constructed pmirGLO vectors were then transfected into GC cells with miR-152-3p mimics or NC mimics. 48 hours after transfection, luciferase activities were evaluated by the Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA).
Western Blot
Proteins were extracted using RIPA buffer (Beyotime Institute of Biotechnology). Protein concentration was measured with the bicinchoninic acid assay (Beyotime Institute of Biotechnology). Following denaturation, 10 µg protein/lane was separated by 10% SDS-PAGE. Proteins were transferred onto polyvinylidene difluoride (PVDF) membranes and blocked in 5% non-fat milk for 2 h at room temperature. The membranes were incubated with primary antibodies against HNF1A (1:1000; ab96777; Abcam), DNAJB12 (1:1000; ab154410; Abcam), PCNA (1:1000; ab92552; Abcam), E-cadherin (1:1000; ab231303; Abcam), vimentin (1:1000; ab16700; Abcam) and GAPDH (1:1000; sc-47724; Santa Cruz Biotechnology) overnight at 4°C. Following primary incubation, membranes were incubated with secondary antibodies for 2 h at room temperature. Protein bands were visualized using the Pierce ECL Western Blotting kit (Pierce; Thermo Fisher Scientific, Inc.). Protein expression was quantified using Image-Pro® Plus software (version 6.0; Media Cybernetics, Inc.). GAPDH was used as an endogenous control for data normalization.
Statistical Analysis
Data are presented as the mean ± standard deviation (SD). Each experiment was performed at least 3 times. Statistical analysis was carried out with SPSS 18.0 software (SPSS, Chicago, IL, USA). Comparisons between two groups were performed by a Student’s t-test. Comparisons among three groups were analyzed using one-way ANOVA followed by Tukey’s test. The correlation between gene expression levels was analyzed by Pearson’s correlation coefficient. P < 0.05 was defined as statistically significant.
Results
HCG18 Is Upregulated in GC and Associated with a Poor Prognosis
According to the TCGA database, HCG18 expression levels were upregulated in stomach adenocarcinoma (STAD) tissues compared with normal tissues (Figure 1A). In addition, RT-qPCR revealed that HCG18 was highly expressed in GC tissues (n=26) or cell lines (MKN45, AGS, SCH and SNU638) compared with that in adjacent normal tissues (n=26) or human gastric mucosa cell line (GES-1) (Figure 1B and C). Moreover, GC patients with high levels of HCG18 possessed a lower survival rate compared with patients with low expression levels of HCG18 (Figure 1D). Furthermore, it was found that HCG18 expression was positively correlated with tumor size, TNM stage, lymph node metastasis and distant metastasis, while no correlation was found between the HCG18 expression and gender or age (Table 1).
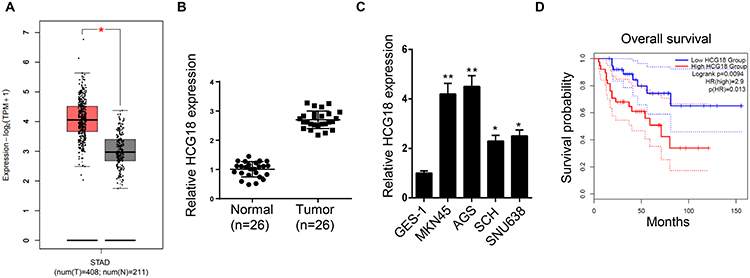 |
Figure 1 HCG18 is upregulated in GC and associated with a poor prognosis. (A) TCGA analysis showed the levels of HCG18 expression in stomach adenocarcinoma (STAD) tissues and normal tissues. (B) RT-qPCR analysis showed the relative expression of HCG18 in GC tissues (n=26) and adjacent normal tissues (n=26). (C) RT-qPCR showed the relative expression of HCG18 in human gastric mucosa cell (GES-1) and GC cells (MKN45, AGS, SCH and SNU638). (D) Kaplan-Meier survival analysis showed the correlation between HCG18 expression and prognosis of GC patients. *p<0.05, **p<0.01. |
Knockdown of HCG18 Inhibits the Tumorigenesis of GC
To investigate the biological role of HCG18 in GC progression, loss-of-function assays were performed. RT-qPCR confirmed that HCG18 was effectively silenced by transfecting sh-HCG18#1 and sh-HCG18#2 into GC cells (Figure 2A). CCK-8 assay revealed that HCG18 knockdown decreased the viability of MKN45 and AGS cells (Figure 2B). Wound healing and transwell assays indicated that knockdown of HCG18 inhibited the migration and invasion of GC cells (Figure 2C and D). These results suggested that HCG18 promoted the proliferation, migration and invasion of GC cells.
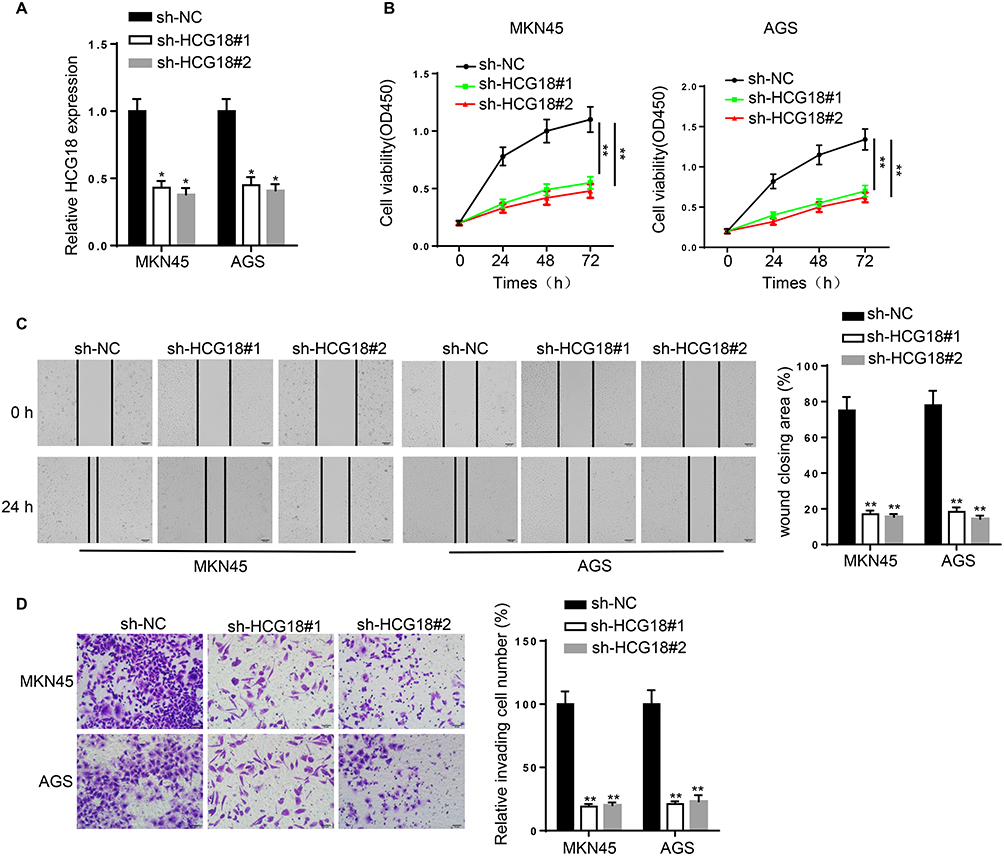 |
Figure 2 Knockdown of HCG18 inhibits the tumorigenesis of GC. (A) The knockdown efficiency of sh-HCG18#1/2 was assessed by RT-qPCR. (B) CCK-8 assay showed the proliferation of MKN45 and AGS cells transfected with sh-NC, sh-HCG18#1 and sh-HCG18#2. (C and D) Cell migration and invasion were detected by wound healing and transwell assays. *p<0.05, **p<0.01. |
HNF1A Activates the Transcription of HCG18
To further explore the upstream factors of HCG18 in GC, we used PROMO (http://alggen.lsi.upc.es/) and JASPAR (http://jaspar.genereg.net/) databases to determine the transcription factors that may regulate HCG18. The DNA motif of HNF1A and predicted five binding sites of HNF1A in HCG18 promoter were illustrated in Figure 3A and B. RT-qPCR and Western blot showed that the shRNAs targeting HNF1A (sh-HNF1A#1/2/3) caused an apparent decrease of HNF1A expression in MKN45 cells (Figure 3C and D). As shown in Figure 3E, the luciferase activity of vectors constructed with HCG18 wide-type promoter region was decreased by silencing of HNF1A, confirming the binding ability between HNF1A and HCG18 promoter region. Besides, knockdown of HNF1A also reduced HCG18 expression (Figure 3F). Among the five binding sites, we hypothesized the first binding site (Region-1) might be responsible for the binding between HNF1A and HCG18 promoter. The detailed binding sequences and designed mutant sequences were showed in Figure 3G. Luciferase reporter assay revealed that silencing HNF1A weakened the luciferase activity of vectors with HCG18 wild-type promoter region-1 whilst no such alteration was observed in vectors with HCG18 mutant promoter region-1, implying that HNF1A could bind with HCG18 promoter at the first binding site (Figure 3H). Moreover, we found that knockdown of HNF1A inhibited the proliferation, migration and invasion of GC cells (Figure 3I–K). Taken together, HNF1A activated the transcription of HCG18 by binding with the promoter region of HCG18.
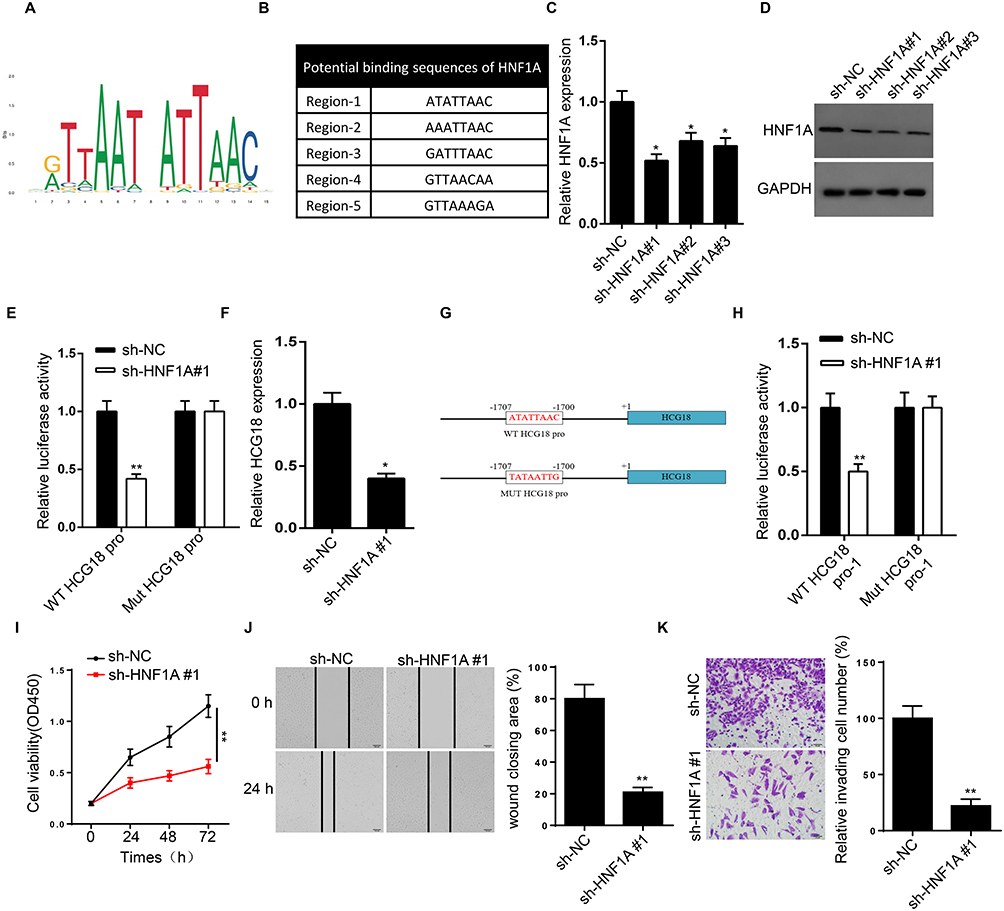 |
Figure 3 HNF1A activates transcription of HCG18. (A and B) DNA motif of HNF1A and predicted five binding sites of HNF1A in HCG18 promoter were provided by PROMO and JASPAR databases. (C and D) The knockdown efficiency of sh-HNF1A#1/2/3 in MKN45 cells was confirmed by RT-qPCR and Western blot. (E) Luciferase reporter assay was adopted to verify the binding ability between HNF1A and the HCG18 promoter in MKN45 cells. (F) RT-qPCR showed the relative expression of HCG18 in MKN45 cells transfected with sh-NC and sh-HNF1A#1. (G) The wild-type and mutant sequences of HCG18 promoter region-1 were presented. (H) Luciferase reporter assay verified that HNF1A bound with HCG18 at the first predicted promoter region. (I–K) CCK-8, wound healing and transwell assays showed the viability, migration and invasion of MKN45 cells transfected with sh-NC and sh-HNF1A. *p<0.05, **p<0.01. |
HCG18 Promotes GC Progression via miR-152-3p
By performing bioinformatics analysis on starBase, 5 potential targeting miRNAs (miR-34a-5p, miR-34c-5p, miR-152-3p, miR-148a-3p and miR-148b-3p) of HCG18 were screened out. RT-qPCR revealed that miR-152-3p was significantly downregulated in GC tissues (Figure 4A). Moreover, the expression of miR-152-3p was downregulated in GC cell lines (Figure 4B). Kaplan-Meier survival analysis showed that GC patients with low miR-152-3p expression had a shorter overall survival time compared with those patients with high miR-152-3p expression (Figure 4C). In addition, miR-152-3p expression was strikingly associated with tumor size, TNM stage, distant metastasis and lymph node metastasis (Table 1).
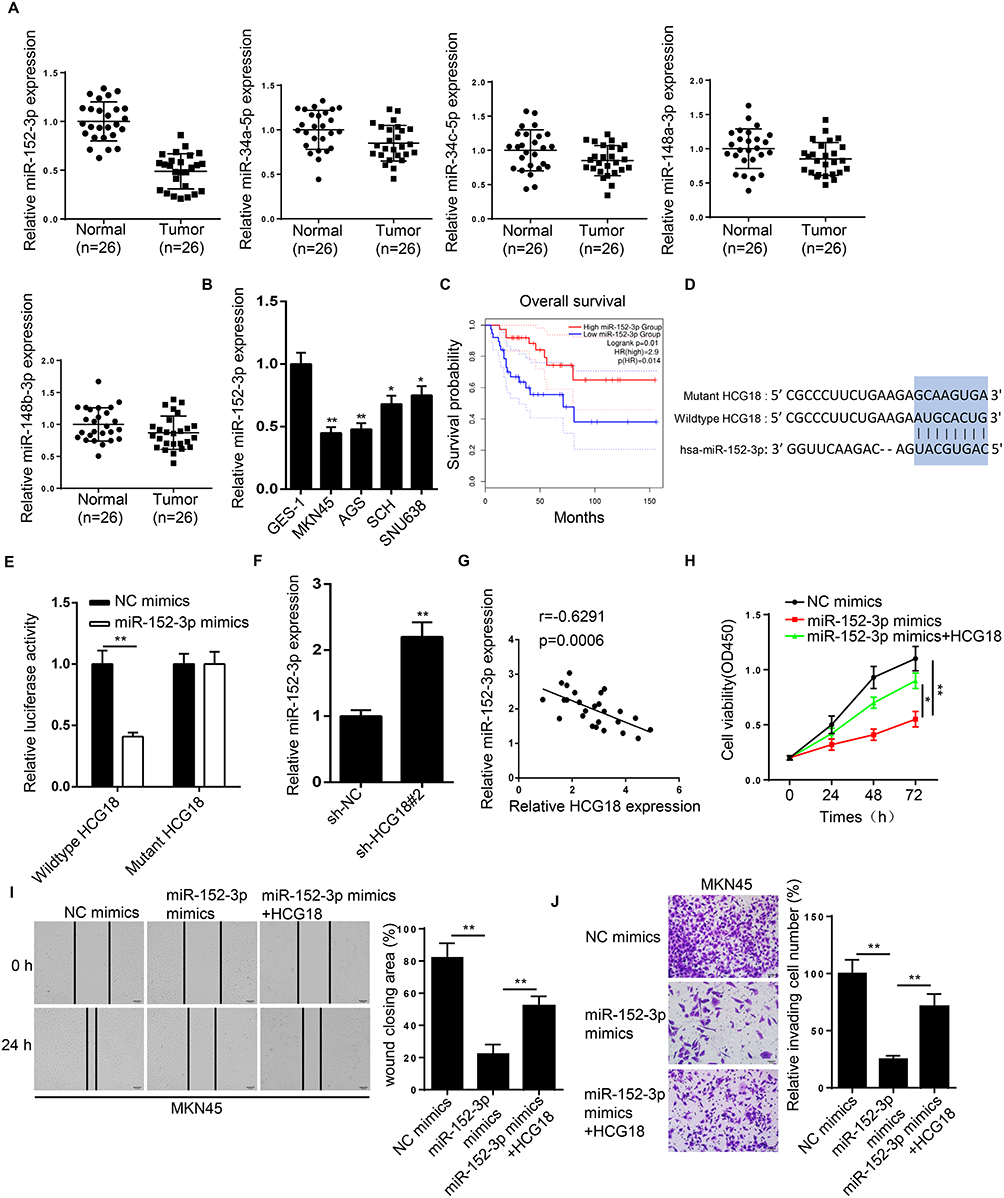 |
Figure 4 HCG18 promotes GC progression via miR-152-3p. (A) RT-qPCR showed the relative expression of miR-152-3p, miR-34a-5p, miR-34c-5p, miR-148a-3p and miR-148b-3p in GC tissues (n=26) and adjacent normal tissues (n=26). (B) RT-qPCR showed the relative expression of miR-152-3p in human gastric mucosa cells (GES-1) and GC cells (MKN45, AGS, SCH and SNU638). (C) Kaplan-Meier survival analysis showed the correlation between miR-152-3p expression and prognosis of GC patients. (D) Starbase showed the binding site between HCG18 and miR-152-3p. (E) Luciferase reporter assay was adopted to verify the binding ability between miR-152-3p and HCG18 in MKN45 cells. (F) RT-qPCR showed the relative expression of miR-152-3p in MKN45 cells transfected with sh-HCG18#2. (G) Pearson correlation analysis shows a negative correlation between HCG18 and miR-152-3p in patients with GC tumors. (H–J) CCK-8, wound healing and transwell assays showed the proliferation, migration and invasion of MKN45 cells transfected with NC mimics and miR-152-3p mimics. *p<0.05, **p<0.01. |
To verify the binding ability between miR-152-3p and HCG18, mutant sequences were designed and presented in Figure 4D. Luciferase reporter assay revealed that miR-152-3p mimics reduced the relative luciferase activity of the MKN45 cells transfected with the wild-type HCG18-fused luciferase gene, whereas no significant differences were observed in cells harboring the mutant HCG18 (Figure 4E). In addition, knockdown of HCG18 increased miR-152-3p expression (Figure 4F). Pearson correlation analysis further demonstrated that HCG18 expression was negatively correlated with miR-152-3p expression in GC tissues (Figure 4G). Moreover, CCK-8, wound healing and transwell assays revealed that miR-152-3p mimics inhibited the proliferation, migration and invasion of GC cells, while this effect was abrogated by HCG18 overexpression (Figure 4H–J). Based on these results, it was confirmed that HCG18 executed its oncogenic role in GC cells by regulating miR-152-3p.
HCG18 Promotes DNAJB12 Expression by Acting as a ceRNA of miR-152-3p in GC Cells
6 potential target genes (DNAJB12, COX6A1, PNPLA6, PDRG1, RNF145 and NDUFAF6) were selected out from starBase under a specific condition (CLIP Data: strict stringency; Degradome Data: high stringency; Pan-cancer number: 8). The expressions of target genes were determined in GC tissues and DNAJB12 was the most significantly upregulated (Figure 5A). In addition, the expression of DNAJB12 was also higher in GC cell lines than that in normal gastric cells (Figure 5B). The predicted binding sites between DNAJB12 and miR-152-3p were displayed in Figure 5C. Luciferase reporter assay illustrated that the luciferase activity of cells with wild-type 3ʹUTR of DNAJB12 was significantly reduced by miR-152-3p overexpression whilst no alteration was observed in cells with mutant 3ʹUTR of DNAJB12 (Figure 5D). In addition, we found that the overexpression of miR-152-3p downregulated DNAJB12 expression (Figure 5E and F), and DNAJB12 expression was negatively correlated with miR-152-3p expression in GC tissues (Figure 5G). Moreover, knockdown of HCG18 reduced the expression of DNAJB12 (Figure 5H and I), and there was a positive correlation between the expression of HCG18 and DNAJB12 (Figure 5J). Meanwhile, the expression of DNAJB112 was increased by the silencing of miR-152-3p, but this effect was partially abolished by knockdown of HCG18 in MKN45 cells (Figure 5K and L). Likewise, miR-152-3p inhibitor enhanced the luciferase activity of wild-type DNAJB112 reporters, while knockdown of sh-HCG18#2 counteracted such promotion (Figure 5M). Collectively, these findings revealed that HCG18 upregulated DNAJB12 expression through competitively binding with miR-152-3p.
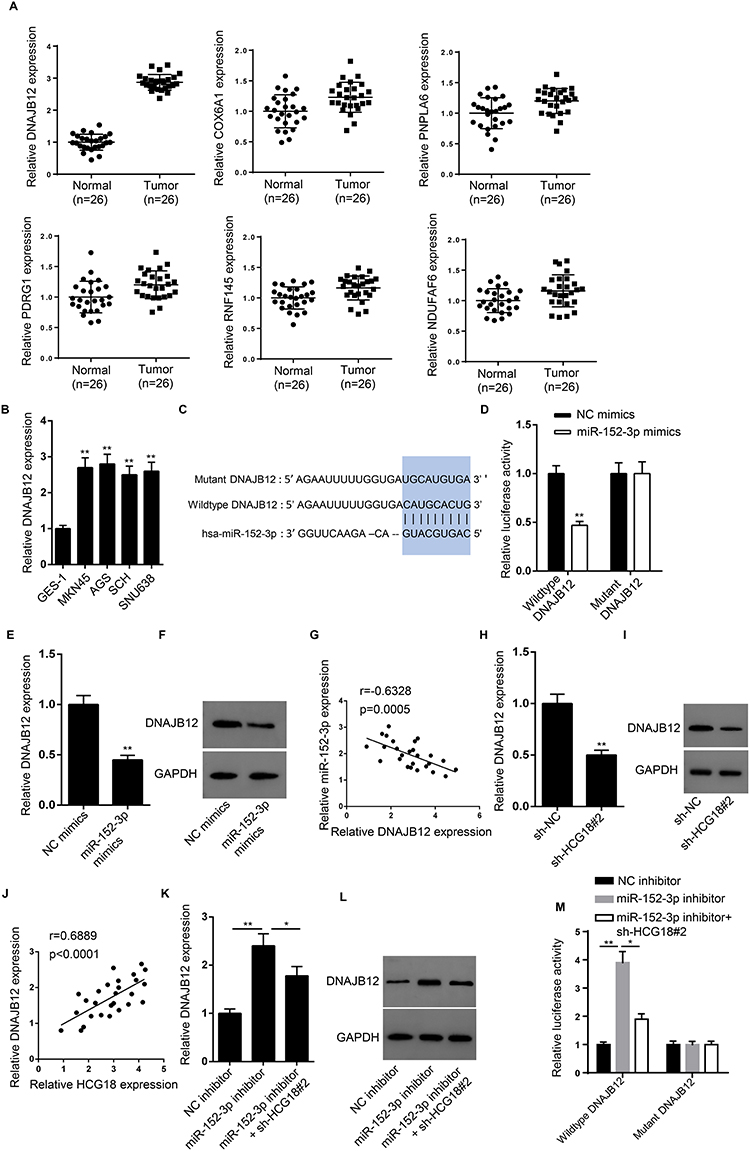 |
Figure 5 HCG18 promotes DNAJB12 expression by acting as a ceRNA of miR-152-3p in GC cells. (A) The relative expression of 6 potential target genes (DNAJB12, COX6A1, PNPLA6, PDRG1, RNF145 and NDUFAF6) was detected in GC tissues (n=26) and adjacent normal tissues (n=26) by RT-qPCR assay. (B) The relative expression of DNAJB12 in human gastric mucosa cells and GC cells was detected by RT-qPCR. (C) Starbase showed the binding site between DNAJB12 and miR-152-3p. (D) Luciferase reporter assay was used to confirm the binding ability between miR-152-3p and DNAJB12 in MKN45 cells. (E and F) RT-qPCR and Western blot showed the mRNA and protein levels of DNAJB12 in MKN45 cells transfected with NC mimics and miR-152-3p mimics. (G) The correlation between DNAJB12 and miR-152-3p in GC tissues was analyzed by Spearman analysis. (H and I) RT-qPCR and Western blot showed the mRNA and protein levels of DNAJB12 in MKN45 cells transfected with sh-NC and sh-HCG18#2. (J) The correlation between DNAJB12 and HCG18 in GC tissues was analyzed by Spearman analysis. (K and L) RT-qPCR and Western blot showed the mRNA and protein levels of DNAJB12 in MKN45 cells transfected with NC inhibitor, miR-152-3p inhibitor and miR-152-3p inhibitor+sh-HCG18#2. (M) Luciferase activity of wild-type DNAJB12 vectors was monitored by luciferase reporter assay in MKN45 cells transfected with NC inhibitor, miR-152-3p inhibitor or miR-152-3p inhibitor+sh-HCG18#2. *p<0.05, **p<0.01. |
Overexpression of DNAJB12 Partially Restored HCG18 Knockdown-Attenuated Progression of GC
RT-qPCR and Western blot showed that DNAJB12 was significantly upregulated in MKN45 cells transfected with DNAJB12 overexpression plasmid (Figure 6A and B). Next, the levels of epithelial-mesenchymal transition (EMT) proteins (E-cadherin and Vimentin) and proliferation-related protein (PCNA) were measured, and the results indicated that overexpression of DNAJB12 increased the expression of PCNA and Vimentin but decreased the expression of E-cadherin (Figure 6C and D). To explore whether HCG18 accelerated GC progression through regulating DNAJB12 expression, rescue assays were performed. CCK-8, wound healing and transwell assays revealed that overexpression of DNAJB12 abolished the inhibitory effect of HCG18 knockdown on the proliferation, migration, and invasion of GC cells (Figure 6E–G). In conclusion, these results demonstrated that HCG18 promoted GC progression by regulating DNAJB12 expression.
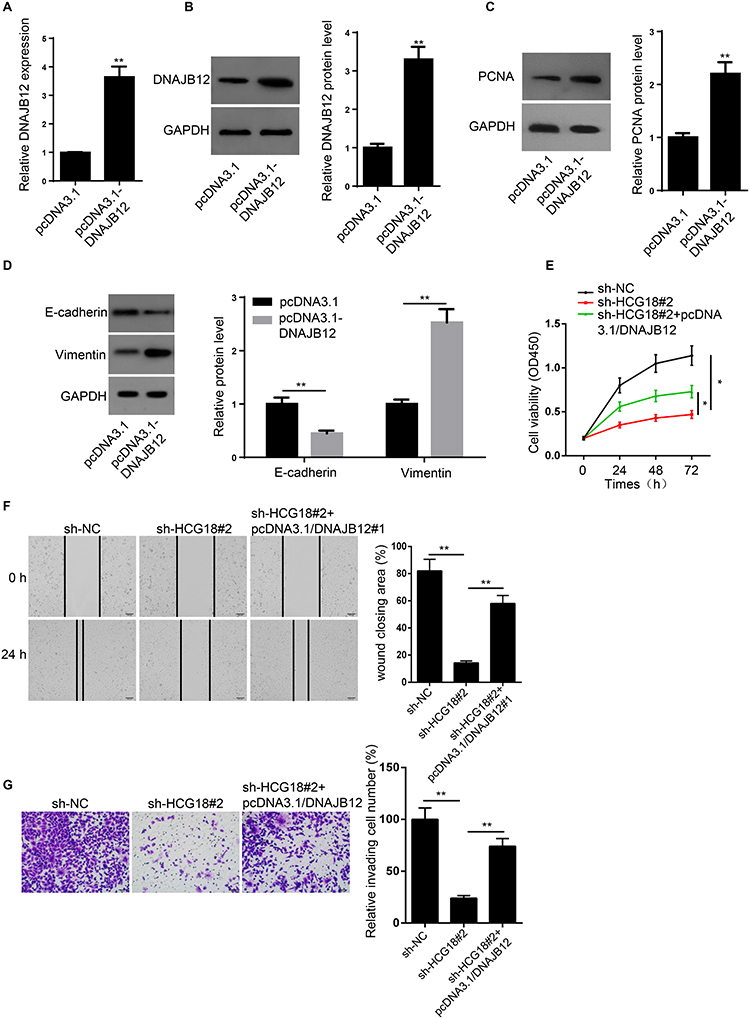 |
Figure 6 Overexpression of DNAJB12 partially restored HCG18 knockdown-attenuated progression of GC. (A and B) RT-qPCR and Western blot showed the mRNA and protein levels of DNAJB12 in MKN45 cells transfected with pcDNA3.1and pcDNA3.1/DNAJB12. (C and D) Western blot showed the protein levels of PCNA, E-cadherin and vimentin in MKN45 cells transfected with DNAJB12 and pcDNA3.1/DNAJB12. (E–G) CCK-8, wound healing and transwell assays were used to evaluate the proliferation, migration and invasion abilities of MKN45 cells transfected with sh-NC, sh-HCG18#2 and sh-HCG18#2 + pcDNA3.1/DNAJB12. *p<0.05, **p<0.01. |
Discussion
Accumulating researches have demonstrated that lncRNAs play vital roles in the development and progression of various types of cancers, including GC.16 For instance, LINC00858 was upregulated in GC tissues and patients with high expression of LINC00858 had poor overall survival.17 LINC00152 promoted the migration and invasion of GC cells through inhibiting ERK/MAPK signaling.18 LncRNA MALAT1 promoted cell proliferation and contributed cisplatin resistance in GC via PI3K/AKT pathway.19 In the present study, it was demonstrated that HCG18 was upregulated in GC tissues and cell lines, and high expression of HCG18 was associated with poor prognosis in patients with GC. Moreover, loss-of-function assays demonstrated that knockdown of HCG18 inhibited the proliferation, migration, and invasion of GC cells. These results suggested that HCG18 acted as an oncogenic role in GC progression.
Transcription factor has been reported to promote or inhibit the development and progression of various cancers through regulating the expression of lncRNAs. For example, Huang et al reported that transcription factor YY1 promoted the tumorigenesis of lung cancer progression by activating lncRNA PVT1.20 Gao et al indicated that SP1-induced lncRNA TINCR overexpression inhibited the migration and invasion of lung adenocarcinoma cells.21 Hepatocyte nuclear factor 1 alpha (HNF1A) was reported to increase the expression of CASC2, which further regulated pancreatic cancer progression.22 In our study, we investigated the upstream regulatory mechanism of HCG18 and discovered that HNF1A could directly bind to the promoter region of HCG18 to increase the expression of HCG18, and knockdown of HNF1A inhibited the proliferation, migration, and invasion of GC cells. In sum, we confirmed that HNF1A acted as a transcription activator to promote the expression of HCG18 in GC.
Increasing evidence indicated that lncRNA could function as a competing endogenous RNA (ceRNA) to regulate the expression of target genes of miRNA via competitively binding miRNA in GC.23,24 Tao et al reported that lncRNA OIP5-AS1 could function as a ceRNA in regulating HMGA2 through competitive binding to miR-367-3p in GC.25 Xie et al demonstrated that lncRNA OIP5-AS1 upregulated ANO1 expression by sponging miR-422a to promote GC cell growth.26 miR-152-3p was reported to be downregulated and serve as a tumor suppressor in several cancers, such as glioma, breast cancer, and prostate cancer.27–29 In our study, miR-152-3p was demonstrated to be a target gene of HCG18, and knockdown of HCG18 upregulated the expression of miR-152-3p. The absorbing effect of HCG18 on miR-152-3p in GC was first discovered. In addition, we discovered that miR-152-3p inhibited DNAJB12 expression by direct interaction. The silencing of miR-152-3p increased the expression of DNAJB12, while this effect was reversed by HCG18 knockdown. Furthermore, the expression of HCG18 was negatively correlated with miR-152-3p but positively correlated with DNAJB12 in GC tissues. Moreover, rescue assays demonstrated that overexpression of DNAJB12 partially abolished the inhibitory effect of HCG18 knockdown on the proliferation, migration, and invasion of GC cells. In sum, these results demonstrated that HCG18 promoted GC progression by acting as a ceRNA of miR-152-3p to upregulate DNAJB12 expression.
In conclusion, the present study for the first time reported the potential molecular mechanisms of HCG18 in the tumorigenesis of GC. Our study identified the HNF1A-induced upregulation of HCG18 promoted cell proliferation, migration, and invasion by competitively binding with miR-152-3p and upregulating DNAJB12 expression in GC. However, the limitations of the present study are required to be addressed in future studies. Firstly, whether other microRNAs or downstream effectors are crucial to HCG18-regulated phenotypes of GC. Secondly, in vivo experiments are required to further validate these findings in follow-up studies.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Shimizu D, Kanda M, Kodera Y. Review of recent molecular landscape knowledge of gastric cancer. Histol Histopathol. 2018;33(1):11–26. doi:10.14670/HH-11-898
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.21387
3. Wadhwa R, Taketa T, Sudo K, et al. Modern oncological approaches to gastric adenocarcinoma. Gastroenterol Clin North Am. 2013;42(2):359–369. doi:10.1016/j.gtc.2013.01.011
4. Derieux S, Svrcek M, Manela S, et al. Evaluation of the prognostic impact of pathologic response to preoperative chemotherapy using mandard’s tumor regression grade (TRG) in gastric adenocarcinoma. Dig Liver Dis. 2019.
5. Pei Y, Tang Z, Cai M, et al. The E2F3/miR-125a/DKK3 regulatory axis promotes the development and progression of gastric cancer. Cancer Cell Int. 2019;19(1):212. doi:10.1186/s12935-019-0930-y
6. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
7. Karimi P, Islami F, Anandasabapathy S, et al. Gastric cancer: descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol Biomarkers Prev. 2014;23(5):700–713. doi:10.1158/1055-9965.EPI-13-1057
8. Zheng P, Li H, Xu P, et al. High lncRNA HULC expression is associated with poor prognosis and promotes tumor progression by regulating epithelial-mesenchymal transition in prostate cancer. Arch Med Sci. 2018;14(3):679–686. doi:10.5114/aoms.2017.69147
9. Zhang C, Su C, Song Q, et al. LncRNA PICART1 suppressed non-small cell lung cancer cells proliferation and invasion by targeting AKT1 signaling pathway. Am J Transl Res. 2018;10(12):4193–4201.
10. Xu Z, Huang B, Zhang Q, et al. NOTCH1 regulates the proliferation and migration of bladder cancer cells by cooperating with long non-coding RNA HCG18 and microRNA-34c-5p. J Cell Biochem. 2019;120(4):6596–6604. doi:10.1002/jcb.27954
11. Wang W, Zhao Z, Yang F, et al. An immune-related lncRNA signature for patients with anaplastic gliomas. J Neurooncol. 2018;136(2):263–271. doi:10.1007/s11060-017-2667-6
12. Abba M, Mudduluru G, Allgayer H. MicroRNAs in cancer: small molecules, big chances. Anticancer Agents Med Chem. 2012;12(7):733–743. doi:10.2174/187152012802650273
13. Wang Y, Qin H. miR-338-3p targets RAB23 and suppresses tumorigenicity of prostate cancer cells. Am J Cancer Res. 2018;8(12):2564–2574.
14. Chen L, Gao H, Liang J, et al. miR-203a-3p promotes colorectal cancer proliferation and migration by targeting PDE4D. Am J Cancer Res. 2018;8(12):2387–2401.
15. You W, Zhang X, Ji M, et al. MiR-152-5p as a microRNA passenger strand special functions in human gastric cancer cells. Int J Biol Sci. 2018;14(6):644–653. doi:10.7150/ijbs.25272
16. Chan JJ, Tay Y. Noncoding RNA: RNA regulatory networks in cancer. Int J Mol Sci. 2018;19(5):1310. doi:10.3390/ijms19051310
17. Ai W, Li F, Yu HH, Liang ZH, Zhao HP. Up-regulation of long noncoding RNA LINC00858 is associated with poor prognosis in gastric cancer. J Gene Med. 2020;e3179.
18. Hu X-L, Wang J, He W, et al. Down-regulation of lncRNA Linc00152 suppressed cell viability, invasion, migration, and epithelial to mesenchymal transition, and reversed chemo-resistance in breast cancer cells. Eur Rev Med Pharmacol Sci. 2018;22(10):3074–3084. doi:10.26355/eurrev_201805_15067
19. Dai Q, Zhang T, Li C. LncRNA MALAT1 regulates the cell proliferation and cisplatin resistance in gastric cancer via PI3K/AKT pathway. Cancer Manag Res. 2020;12:1929–1939. doi:10.2147/CMAR.S243796
20. Huang T, Wang G, Yang L, et al. Transcription factor YY1 modulates lung cancer progression by activating lncRNA-PVT1. DNA Cell Biol. 2017;36(11):947–958. doi:10.1089/dna.2017.3857
21. Gao YW, Ma F, Xie Y-C, et al. Sp1-induced upregulation of the long noncoding RNA TINCR inhibits cell migration and invasion by regulating miR-107/miR-1286 in lung adenocarcinoma. Am J Transl Res. 2019;11(8):4761–4775.
22. Yu Y, Liang S, Zhou Y, et al. HNF1A/CASC2 regulates pancreatic cancer cell proliferation through PTEN/Akt signaling. J Cell Biochem. 2019;120(3):2816–2827. doi:10.1002/jcb.26395
23. Guo LL, Song CH, Wang P, Dai LP, Zhang JY, Wang KJ. Competing endogenous RNA networks and gastric cancer. World J Gastroenterol. 2015;21(41):11680–11687. doi:10.3748/wjg.v21.i41.11680
24. Zhang M, Du X. Noncoding RNAs in gastric cancer: research progress and prospects. World J Gastroenterol. 2016;22(29):6610–6618. doi:10.3748/wjg.v22.i29.6610
25. Tao Y, Wan X, Fan Q, et al. Long non-coding RNA OIP5-AS1 promotes the growth of gastric cancer through the miR-367-3p/HMGA2 axis. Dig Liver Dis. 2020;52(7):773–779. doi:10.1016/j.dld.2019.11.017
26. Xie R, Liu L, Lu X, et al. LncRNA OIP5-AS1 facilitates gastric cancer cell growth by targeting the miR-422a/ANO1 axis. Acta Biochim Biophys Sin (Shanghai). 2020;52(4):430–438. doi:10.1093/abbs/gmaa012
27. Shi J, Zhang Y, Qin B, et al. Long non-coding RNA LINC00174 promotes glycolysis and tumor progression by regulating miR-152-3p/SLC2A1 axis in glioma. J Exp Clin Cancer Res. 2019;38(1):395. doi:10.1186/s13046-019-1390-x
28. Ge S, Wang D, Kong Q, et al. Function of miR-152 as a tumor suppressor in human breast cancer by targeting PIK3CA. Oncol Res. 2017;25(8):1363–1371. doi:10.3727/096504017X14878536973557
29. Feng F, Liu H, Chen A, et al. miR-148-3p and miR-152-3p synergistically regulate prostate cancer progression via repressing KLF4. J Cell Biochem. 2019;120(10):17228–17239. doi:10.1002/jcb.28984
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.