Back to Journals » Neuropsychiatric Disease and Treatment » Volume 18
HIV, HSV, SARS-CoV-2 and Ebola Share Long-Term Neuropsychiatric Sequelae
Authors Büttiker P ![]() , Stefano GB
, Stefano GB ![]() , Weissenberger S
, Weissenberger S ![]() , Ptacek R
, Ptacek R ![]() , Anders M
, Anders M ![]() , Raboch J
, Raboch J ![]() , Kream RM
, Kream RM
Received 14 July 2022
Accepted for publication 2 September 2022
Published 5 October 2022 Volume 2022:18 Pages 2229—2237
DOI https://doi.org/10.2147/NDT.S382308
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Pascal Büttiker,1 George B Stefano,1 Simon Weissenberger,2 Radek Ptacek,1 Martin Anders,1 Jiri Raboch,1 Richard M Kream1
1Department of Psychiatry, First Faculty of Medicine, Charles University and General University Hospital in Prague, Prague, Czech Republic; 2Department of Psychology, University of New York in Prague, Prague, Czech Republic
Correspondence: Pascal Büttiker, Altenbach 26, Vaduz, 9490, Liechtenstein, Tel +41 78 653 8442, Email [email protected]
Abstract: Long COVID, in which disease-related symptoms persist for months after recovery, has led to a revival of the discussion of whether neuropsychiatric long-term symptoms after viral infections indeed result from virulent activity or are purely psychological phenomena. In this review, we demonstrate that, despite showing differences in structure and targeting, many viruses have highly similar neuropsychiatric effects on the host. Herein, we compare severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), human immunodeficiency virus 1 (HIV-1), Ebola virus disease (EVD), and herpes simplex virus 1 (HSV-1). We provide evidence that the mutual symptoms of acute and long-term anxiety, depression and post-traumatic stress among these viral infections are likely to result from primary viral activity, thus suggesting that these viruses share neuroinvasive strategies in common. Moreover, it appears that secondary induced environmental stress can lead to the emergence of psychopathologies and increased susceptibility to viral (re)infection in infected individuals. We hypothesize that a positive feedback loop of virus-environment-reinforced systemic responses exists. It is surmised that this cycle of primary virulent activity and secondary stress-induced reactivation, may be detrimental to infected individuals by maintaining and reinforcing the host’s immunocompromised state of chronic inflammation, immunological strain, and maladaptive central-nervous-system activity. We propose that this state can lead to perturbed cognitive processing and promote aversive learning, which may manifest as acute, long-term neuropsychiatric illness.
Keywords: HIV-1, SARS virus, virus latency, neuropsychiatry, interoception
Introduction
COVID-19 and its causative agent, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), have left a mark on the early 21st century. Confronted with a novel pandemic scenario, researchers and scientists worldwide were compelled to work together in analyzing and containing the rapidly spreading disease. This viral pandemic continues to have interlinked effects on a global scale, and its ultimate toll is only beginning to be understood. Beyond the physical and mental strain directly caused by the virus, governmental measures to contain the spread of the disease—such as social-distancing rules, economic lockdowns and the fear-evoking and often uncertain flow of information—may have negatively affected people’s wellbeing.1 Furthermore, the unified focus in fighting against this pandemic has exhausted scientific and medical resources. As such, the extensive attention being paid to this newly encountered pathogen has caused the scientific and medical management of many other diseases to be deferred. For example, in the case of human immunodeficiency virus (HIV), ongoing research has stagnated, and disruptions have occurred in medical supply chains and the intervention programs necessary to contain and treat the disease.2
Despite the harmful effects of SARS-CoV-2 and the economic, medical and societal measures associated with the pandemic, highly focused research on SARS-CoV-2 has vastly increased the understanding of viruses and their infectious strategies. Countless studies on COVID-19 were conducted, and reviews were published at unprecedented pace, thereby enabling new knowledge and understanding of human-virus interactions.3,4
An emerging yet still vastly disregarded aspect of viral infections is their associated long-term neurological/neuropsychiatric symptoms, which can manifest as both neurological illness and psychopathological expression.1,5,6 The observation of long COVID, also called post-acute sequelae of SARS-CoV-2, has recently revived the focus on the dangers associated with ongoing persistent infection after disease recovery.6,7 Long-term neuropsychiatric symptoms after viral infection are well-known phenomena that have been thoroughly documented and studied in diseases such HIV-1,8 herpes simplex virus 1 (HSV-1)9 and recently Ebola virus disease (EVD).10,11
Common Viral Strategies for Systemic Infection and Neuroinvasion: Primary Effects of Viral Activity
Neuroinvasion of the central nervous system (CNS) and the ability of viruses to “hide” in CNS-resident structures may be a major cause of subliminal infection and persistent disease. Given the scarcely accessible nature of CNS cells behind the blood–brain barrier (BBB) and their cytolytic sensitivity, neuronal cells can provide an ideal protective environment supporting reservoirs of persistent viral replication and latency.12,13 To exploit this protective niche, viruses have been suggested to use diverse routes of neuroinvasion. HIV-1, for example, targets immune cells, such as lymphocytes (eg, CD4+ T cells) and monocytes (eg, macrophages and dendritic cells) that express a high surface density of CD4 receptors, which viruses may use for facilitated transport across the BBB as a direct entrance route.13,14 CD4-expressing immune cells in the brain, such as astrocytes, pericytes, and microglia, may then be recruited and subsequently harbor the viral genome within the CNS.14–16 Protected by the BBB, the viral genome can then silently replicate in microglia and other CNS-resident structures, thus potentially expanding viral reservoirs and causing persistent deep structural infection and neuronal dysfunction.12,14 This phenomenon of dormant infection and reactivation has been extensively studied in HIV-1, thus providing a potential explanation for the chronicity of infection and associated cognitive symptoms despite antiretroviral therapy.8,12
Similar neurotropic characteristics are increasingly being described for other viruses, such as HSV-1,17 EVD,18 and members of the coronavirus family, such as the mouse hepatitis virus, Middle East respiratory syndrome coronavirus (MERS-CoV) and SARS-CoV-2.7,19,20 For example, in vitro analysis of infected human organoids has revealed the expression of angiotensin-converting enzyme 2 (ACE2), the main receptor for SARS-CoV-2, in lymph-node-resident macrophages,21 multiple brain areas, and CNS-resident glial cells, such as astrocytes, thus supporting similar mechanisms of neurovirulence across these viruses.22 Although the presence of such neuroinvasive pathways remains inconclusive for many viruses, secondary CNS infection via virally induced loss of BBB integrity with elevated CNS influx of inflammatory molecules and viral particles has been widely accepted.23,24 Thus, subsequent cytokine storms and associated systemic inflammation leading to multiple organ damage, cardiac dysfunction, and CNS inflammation may be a major culprit of interoceptive disturbance and cognitive malfunction (Figure 1).25
 |
Figure 1 Virus-induced enhancement of blood-brain barrier immune cell trafficking. The figure illustrates how a viral organ infection may stimulate white blood cell (eg, macrophage) and chemical trafficking across the BBB. (1) For example, SARS-CoV-2 infection in the periphery (ie, lung/gut) appears to generate an inflammatory microenvironment, releasing increased levels of activated immune cells that can travel to the brain, altering endothelial integrity, and inducing a similar inflammatory phenomenon there. (2) The increased macrophage excitation, number of immune cells and stimulated cytokine release causes this endothelial disruption, diminishing the immune privilege of the brain. (3) Enhanced trafficking of uninfected and possibly infected macrophages may also facilitate the transport of viral particles into the brain thereby altering microglia and astrocytes homeostasis, building viral reservoirs and affecting cellular interaction. (4) Infection-induced cytokine storms in the brain may further aid viral replication by damaging host DNA, stimulating an even greater rate of RNA polymerase errors and thus, enhanced levels of mutation, causing systemic and persistent infection. We surmise this infection stimulated inflammatory neuronal environment alters mitochondrial function either directly or indirectly in that it requires greater levels of metabolic substrates (eg, oxygen and glucose) to sustain normal functionality, thereby competing with normal neuronal energy demands. As a result, stress-induced viral reactivation and subsequent induction of HIF-1a and mitochondrial ROS may lead to abnormal metabolite generation and distribution, for example, in the form of increased mitochondrial GABA retention and extracellular or cytoplasmic glutamate accumulation, causing functional perturbation of brain networks and disruption in cognitive processing. |
In line with these findings, we suggest that many viruses have shared neurovirulence and CNS-persistence potential, despite fundamental differences in their specific targeting and immune responses, and that viral latency and chronic CNS inflammation are the likely causes, in part, of long-term neuropsychiatric symptoms.23,26 Specifically, we propose that viral latency and reactivation in CNS-harbored infected cells hijack the cellular reproductive machinery and energy expenditure and have deleterious effects on host DNA, metabolic tone and cellular communication, activation and proliferation.7,27–29 The viral interference in metabolic regulatory processes may directly alter intercellular communication and the composition of the extracellular microenvironment, thereby exerting multiple effects on host phenotypic expression and viral replication.27,30,31 Such dysregulated metabolite generation has been demonstrated in latent HIV-1 infection, in which stress-induced reactivation of the virus is associated with upregulated glycolysis,32 and the induction of hypoxia-inducible factor 1a (HIF-1a) and mitochondrial reactive oxygen species (ROS).33,34 These state-dependent shifts in metabolic processing in infected and neighboring cells, and subsequent alterations in metabolite generation, such as increased mitochondrial ROS, gamma aminobutyric acid (GABA) retention and glutamate excitotoxicity, may therefore be responsible for the functional perturbation of brain networks and cognitive disruption dependent on the viral infection phase.35–38 Indeed, targeting mitochondria may represent a conserved evolutionary strategy given their heteroplasmic informational reservoir.39 Thus, viruses, including SARS-CoV-2, exhibit extensive artificial intelligence processes, enabling, for example, mitochondrial hijacking.40
Common Neuropsychiatric Sequelae of Viral Infection: Secondary Effects of Viral Activity
Interestingly, virally induced neurological or neuropsychiatric effects are similar among viruses.26 Often, the sequelae of infections with viruses suggested to be neurovirulent—such as HIV-1, HSV-1, EVD, MERS-CoV, and SARS-CoV-2—have been reported to include encephalopathies (eg, meningitis or encephalitis, seizures, and strokes) with altered mental status; brain fog; fatigue; and increased neuropsychiatric incidence of anxiety, depression and post-traumatic stress (Table 1).5,9,11,41–43 The significance level of the following presented data is p <0.05.
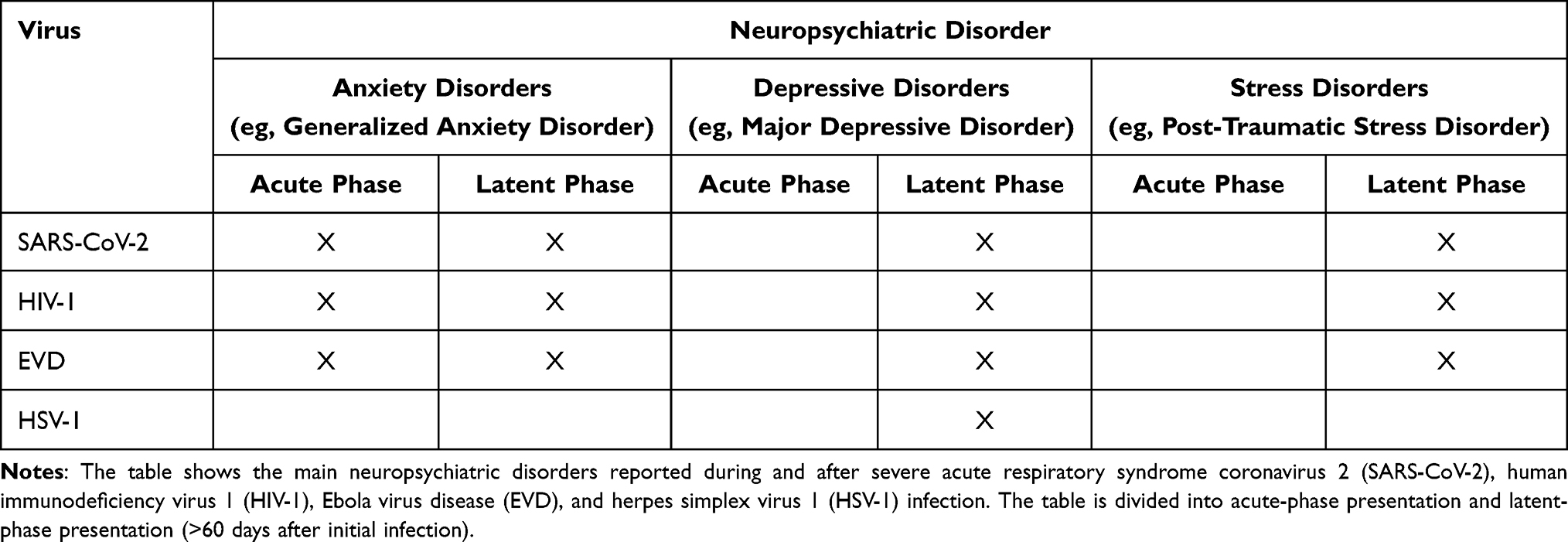 |
Table 1 Neuropsychiatric Disorders Associated with Viral Infections |
Notably, a recent follow-up study of 197 EVD survivors two years after their discharge from the Ebola treatment center has reported a prevalence of anxiety, depression, and post-traumatic stress disorder (PTSD) of (n = 49, 24.9%), (n = 93, 47.2%), and (n = 43, 21.8%), respectively, with older survivors (≥30 years) demonstrating an increased likelihood of developing anxiety (AOR = 3.04, 95% CI 1.2–7.7; p = 0.019) and depression (AOR = 8.5, 95% CI 2.68–27.01; p = 0.001) in comparison to younger survivors (<30 years).11 In addition, decreased exercise after treatment has been associated with greater risks of anxiety and depression (AOR = 2.63, 95% CI 1.25–5.54; p = 0.011).11 Furthermore, other studies have reported similar neuropsychiatric incidents during and after SARS-CoV-2 infection.41,44–46 In a systematic analysis of 72 studies including 3559 cases of coronavirus disease (ie, SARS-CoV, MERS-CoV, and SARS-CoV-2) between the ages of 12 and 68 years, Rogers et al have reported a prevalence of anxiety (35.7%; 95% CI 27.6–44.2) and depression (32.6%; 95% CI 24.7–40.9) during acute infection with symptoms of anxiety (12.3%; 95% CI 7.7–17.7), depression (10.5%; 95% CI 7.5–14.1) and trauma (30.4%; 95% CI 23.9–37.3) persisting months (60 days to 12 years) after the illness.47 A 6-month retrospective COVID-19 cohort study has reported an array of additional neurological issues, such as dementia, encephalitis, intracranial hemorrhage, ischemic stroke, parkinsonism, and myoneural junction and muscle diseases.48
HIV-associated neurological disorders and neuropsychiatric disorders have been observed during the course of life-long HIV-1 infections. Singer and Thames have reported that approximately 24% of the patients with HIV infections display comorbid encephalopathy (eg, aseptic meningitis), with cognitive, motor, and behavioral abnormalities in some cases, despite antiretroviral therapy.49 Furthermore, progressive dementia, changes in mental status, and a substantial prevalence of mood disorders have been reported in HIV infection.49,50 A study conducted by Celesia et al including 251 HIV-infected individuals demonstrated the presence of anxiety in 47% of the subjects (n = 118). Interestingly, within the anxiety group only 21.2% (n = 25; p = 0.047) of the subjects were currently receiving antiretroviral therapy compared to 32.3% (n = 43; p = 0.047) within the group without anxiety.51 It has been shown that up to 20% of HIV-associated anxiety manifests as generalized anxiety disorder.49 PTSD has been estimated to occur in up to 54% of HIV-infected individuals, with the highest prevalence among minority groups and individuals experiencing persistent pain.49 A study conducted by Smith et al including 145 HIV/AIDS diagnosed individuals that have been experiencing persistent HIV-associated pain demonstrated that 53.8% (n = 78) of the participants met the diagnostic criteria for PTSD.52 Major depressive disorder (MDD) is another highly prevalent clinical presentation in HIV infection.49 A high correlation between anxiety symptoms and depressive symptoms during the course of a life-long HIV infection has been established (p < 0.01).53 Interestingly, very limited data on neuropsychiatric symptoms are available for HSV despite its high occurrence; HSV-1 infection rates are as high as 80% in the general population.9 MDD is the only studied and verified neuropsychiatric sequelae of HSV-1 infection that we could find in the literature to date.54 In a UK Biobank cohort study, Ye et al demonstrated significant associations between depression status and HSV-1 antibody (OR = 1.09, 95% CI 1.02–1.16; p = 0.024) and seropositivity (OR = 1.28, 95% CI 1.12–1.47; p < 0.003).54
Although the common neuropsychiatric effects of different viruses indicate the existence of shared neurovirulent processes, other factors may be at play. Mental disorders, such as those described above, are diagnosed based on disease-specific criteria, as defined in diagnostic tools such as the Diagnostic and Statistical Manual of Mental Disorders.55 By implementing psychological tests, life history records and therapist-guided interviews, the psychiatric assessment of mental disorders concentrates largely on externally observable behavior, possibly leaving a physiological causation masked.55,58 This aspect must be considered when gathering and reporting data, because the observable behavior may be reducible to multiple factors. In a viral pandemic, anxiety, depression, and trauma can be environmentally (eg, exteroceptive) induced in infected and non-infected people alike.56,59 Therefore, psychopathological expression in infected individuals does not ultimately imply the presence of virulent activity but can also result from unconscious and conscious stress induced, for example, by social stigma, individual behavior (eg, substance abuse), experience with invasive medical procedures, long-term isolation and medical segregation, inconsistent information or the awareness of one’s affliction.1,49,57 Thus, the frequently observed disease-associated loss of energy (eg, fatigue), diminished interest in one’s environment (eg, anhedonia), and diminished ability to think (eg, brain fog)—symptoms that may lead to the diagnosis of MDD in infected individuals60 may be reducible to purely biological phenomena (eg, infection-stimulated immunological responses), secondary environmentally induced psychological phenomena (eg, exteroceptive responses) or both.49 Likewise, misattributions can result in the diagnosis of other neuropsychiatric disorders.49 Especially trauma- and stressor-related disorders (eg, PTSD), which diagnostic criteria are strictly reliant on exteroceptive events (ie, distinct stressors), may be subject to misassessment.52
A Holistic Cycle of Neuropsychiatric Effects and Latent Viral Infection
Understanding the different processes affecting individuals’ health and behavioral outcomes is essential for successful disease treatment and containment. Both a virally induced endogenous response that alters neuronal functioning in the CNS and an exteroceptive evolved fear of the disease can establish a neuronal level of activity for maladaptive perception and learning.23 Furthermore, chronic psychological stress may increase individuals’ susceptibility to viral infection or reinfection through proinflammatory responses and suppression of cellular immunity.61 We surmise that these effects can be detrimental, particularly in infected individuals. We further suggest that, in a positive feedback loop of cyclical reinforcement, psychological stress may foster the reactivation and aggravation of viral activity in infected individuals, thereby increasing acute psychopathological expression and susceptibility to neuropsychiatric sequelae (Figure 2).
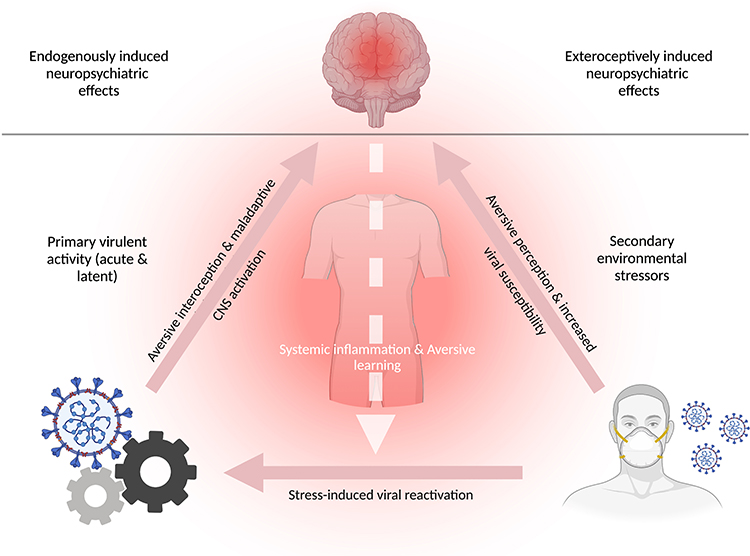 |
Figure 2 A positive feedback loop of primary virulent activity and secondary stress-induced reactivation with neuropsychiatric sequelae. The figure illustrates two distinct factors (ie, levers) that, after stimulation, may produce neuropsychiatric illness. The figure is divided into exteroceptively induced neuropsychiatric effects (top right) and endogenously induced neuropsychiatric effects (top left). The human mask (bottom right) represents external factors (ie, lever 1), such as social stigma, governmental restrictions, individual- and public behavior, or fear induced by inconsistent information and the awareness of one’s affliction; these stressors may cause secondary non-virulent emergence of neuropsychiatric sequelae (eg, anxiety) that can increase susceptibility to viral infection, owing to inflammation-associated immunosuppression, in infected and non-infected individuals. The cogwheels (bottom left) represent virulent activity and the reactivation of latent viruses in the CNS (ie, lever 2), which, after stress-induced activation, may cause shifts in the metabolic processing of infected and neighboring cells, and subsequently alter neuronal metabolite generation and the functionality of brain networks. In a bi-partite manner (ie, via primary virulent activity or secondary exteroceptive stressors), systemic inflammation and aversive perception can be generated, thus providing a basis favoring maladaptive learning. In infected individuals, the activation of each lever may trigger a positive feedback loop of virus-environment-reinforced systemic responses that maintains the host in an immunocompromised state of chronic inflammation (eg, cytokine storm), interoceptive aversion (eg, chronic sympathetic nervous system activation), and maladaptive CNS activity (eg, metabolic disruption of infected CNS-resident glial cells). Therefore, the stimulation of each cyclical element can lead to perturbed cognitive processing and promote aversive learning, which may manifest as acute and long-term neuropsychiatric illness. |
In a predictive coding model, virally induced inflammation and disease-associated interoceptive responses (eg, activation of the autonomic nervous system) foster aversive learning by conceptual overgeneralization in a top-down fashion.62 For example, virally induced chronic states of perceived pain or of anxiety that is not due to a distinct consciously perceived sensory cause may result in the host connecting the aversion to other non-aversive components of higher-order cognition.23 Consequently, the overgeneralization and association of aversive infection-related interoceptive responses to neutral sensory evidence can be funneled and integrated into various memory components.62 This maladaptive learning effect may be strengthened by the virulent changes in the neuronal milieu, thus giving rise to, for example, persistent and generalized anxiety, and maintaining the body in an immunocompromised state of chronic viral activity and aversive psychological reinforcement.23,62
In a bottom-up fashion, negative external stimuli (eg, shock-like exteroceptive experiences) may similarly prompt the reactivation of latent viral particles with subsequent shifts in metabolic processing in infected and neighboring cells.32,63 Such stress-induced responses may be driven psychologically (eg, by trauma or anxiety)64 or physiologically (eg, by muscle injury),65 thereby inducing effects such as alterations in redox balance and ROS production, and further accelerating the inflammatory cycle of subliminal viral infection.63 Stress-induced viral reactivation and associated upregulation of pro-inflammatory cytokines can modulate host circulatory barriers (eg, the BBB and gut endothelial cells) and increase their permeability (ie, leakage), thereby enabling unhindered diffusion of viral particles and other exo- and endo-toxins.66–70 In addition, circulatory barrier disruption may promote microbial dysbiosis and dysregulated interactions among bacteria, bacteriophages, the virus, and the host. Consequently, further upregulation of pro-inflammatory signaling may increasingly induce interoceptive aversion, such as through chronic activation of the sympathetic nervous system.70–73 Hence, environmentally induced upregulation of virulent activity not only strongly facilitates maladaptive learning but also may directly alter the functionality of affected brain networks, such as by causing acute anxiety states and persistent anxiety traits.32,74
Conclusion
The observation of long COVID, also known as post-acute sequelae of SARS-CoV-2, has recently accelerated scientific interest in the emergence of neuropsychiatric long-term persistent symptoms after viral infections. Although differences exist in viral harboring cells and targeting mechanisms, the neuropsychiatric effects of several viruses on the host are highly similar; the most commonly reported comorbidities with these infections include anxiety, depression, and post-traumatic stress. Although these mutual neuropsychiatric effects can result from primary viral activity and shared neurovirulent processes, secondary induced exteroceptive stress, such as disease-associated uncertainty and fear, may also be a culprit of psychopathological expression that can increase susceptibility to viral infection or reinfection. Importantly, in a positive feedback loop of virus-environment-reinforced systemic responses, we hypothesize that a cycle of primary virulent activity and secondary stress-induced reactivation maintains the host in an immunocompromised state of chronic inflammation, immunological strain, and maladaptive CNS activity. The stimulation of each cyclical element can then lead to perturbed cognitive processing and aversive learning, which may manifest as acute and long-term neuropsychiatric illness. In-depth research on the persistent effects of virulent activity in the CNS on cognitive processing and the adaptive role of mitochondria in health and disease (ie, latent vs active viral infection) is strongly recommended.
Acknowledgement
Pascal Büttiker is a PhD Candidate in the Dept. of Psychiatry, First Faculty of Medicine, Charles University in Prague, Czech Republic. BioRender was used to design figures and authors wish to thank the platform for the publication license. Additionally, we would like to thank the Department for hosting this project.
Data Sharing Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Cooperatio Program, research area Neuroscience and by the project MH CZ – DRO VFN64165.
Disclosure
All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Ptacek R, Ptackova H, Martin A, Stefano GB. Psychiatric manifestations of COVID-19 and their social significance. Med Sci Monit. 2020;26:e930340. doi:10.12659/MSM.930340
2. Chenneville T, Gabbidon K, Hanson P, Holyfield C. The impact of COVID-19 on HIV treatment and research: a call to action. Int J Environ Res Public Health. 2020;17(12):4548. doi:10.3390/ijerph17124548
3. Parasher A. COVID research: a year of scientific milestones. Nature. 2021. doi:10.1038/d41586-020-00502-w
4. Wang C, Wang Z, Wang G, Lau JY, Zhang K, Li W. COVID-19 in early 2021: current status and looking forward. Signal Transduct Target Ther. 2021;6(1):114. doi:10.1038/s41392-021-00527-1
5. Stefano GB, Ptacek R, Ptackova H, Martin A, Kream RM. Selective neuronal mitochondrial targeting in SARS-CoV-2 infection affects cognitive processes to induce “Brain Fog” and results in behavioral changes that favor viral survival. Med Sci Monit. 2021;27:e930886. doi:10.12659/MSM.930886
6. Wang F, Kream RM, Stefano GB. Long-term respiratory and neurological sequelae of COVID-19. Med Sci Monit. 2020;26:e928996. doi:10.12659/MSM.928996
7. Stefano GB, Buttiker P, Weissenberger S, Martin A, Ptacek R, Kream RM. Editorial: the pathogenesis of long-term neuropsychiatric COVID-19 and the role of microglia, mitochondria, and persistent neuroinflammation: a hypothesis. Med Sci Monit. 2021;27:e933015. doi:10.12659/MSM.933015
8. Thangaraj A, Periyasamy P, Liao K, et al. HIV-1 TAT-mediated microglial activation: role of mitochondrial dysfunction and defective mitophagy. Autophagy. 2018;14(9):1596–1619. doi:10.1080/15548627.2018.1476810
9. Qiao H, Guo M, Shang J, et al. Herpes simplex virus type 1 infection leads to neurodevelopmental disorder-associated neuropathological changes. PLoS Pathog. 2020;16(10):e1008899. doi:10.1371/journal.ppat.1008899
10. Lotsch F, Schnyder J, Goorhuis A, Grobusch MP. Neuropsychological long-term sequelae of Ebola virus disease survivors - A systematic review. Travel Med Infect Dis. 2017;18:18–23. doi:10.1016/j.tmaid.2017.05.001
11. Bah AJ, James PB, Bah N, Sesay AB, Sevalie S, Kanu JS. Prevalence of anxiety, depression and post-traumatic stress disorder among Ebola survivors in northern Sierra Leone: a cross-sectional study. BMC Public Health. 2020;20(1):1391. doi:10.1186/s12889-020-09507-6
12. Wallet C, De Rovere M, Van Assche J, et al. Microglial cells: the main HIV-1 reservoir in the brain. review. Front Cell Infect Microbiol. 2019;9(362). doi:10.3389/fcimb.2019.00362
13. Illanes-Alvarez F, Marquez-Ruiz D, Marquez-Coello M, Cuesta-Sancho S, Giron-Gonzalez JA. Similarities and differences between HIV and SARS-CoV-2. Int J Med Sci. 2021;18(3):846–851. doi:10.7150/ijms.50133
14. Osborne O, Peyravian N, Nair M, Daunert S, Toborek M. The paradox of HIV blood-brain barrier penetrance and antiretroviral drug delivery deficiencies. Trends Neurosci. 2020;43(9):695–708. doi:10.1016/j.tins.2020.06.007
15. Sawada M, Suzumura A, Marunouchi T. Down regulation of CD4 expression in cultured microglia by immunosuppressants and lipopolysaccharide. Biochem Biophys Res Commun. 1992;189(2):869–876. doi:10.1016/0006-291x(92)92284-5
16. Stefano GB, Bilfinger TV, Fricchione GL. The immune-neuro-link and the macrophage: postcardiotomy delirium, HIV-associated dementia and psychiatry. Prog Neurobiol. 1994;42(4):475–488. doi:10.1016/0301-0082(94)90048-5
17. Meyding-Lamade U, Strank C. Herpesvirus infections of the central nervous system in immunocompromised patients. Ther Adv Neurol Disord. 2012;5(5):279–296. doi:10.1177/1756285612456234
18. Keita AK, Koundouno FR, Faye M, et al. Resurgence of Ebola virus in 2021 in Guinea suggests a new paradigm for outbreaks. Nature. 2021;597(7877):539–543. doi:10.1038/s41586-021-03901-9
19. Tavcar P, Potokar M, Kolenc M, et al. Neurotropic viruses, astrocytes, and COVID-19. Front Cell Neurosci. 2021;15:662578. doi:10.3389/fncel.2021.662578
20. Yachou Y, El Idrissi A, Belapasov V, Ait Benali S. Neuroinvasion, neurotropic, and neuroinflammatory events of SARS-CoV-2: understanding the neurological manifestations in COVID-19 patients. Neurol Sci. 2020;41(10):2657–2669. doi:10.1007/s10072-020-04575-3
21. Xiang Q, Feng Z, Diao B, et al. SARS-CoV-2 induces lymphocytopenia by promoting inflammation and decimates secondary lymphoid organs. Front Immunol. 2021;12(1292). doi:10.3389/fimmu.2021.661052
22. Kumari P, Rothan HA, Natekar JP, et al. Neuroinvasion and encephalitis following intranasal inoculation of SARS-CoV-2 in K18-hACE2 mice. Viruses. 2021;13(1):132. doi:10.3390/v13010132
23. Büttiker P, Weissenberger S, Stefano GB, Kream RM, Ptacek R. SARS-CoV-2, trait anxiety, and the microbiome. Mini review. Front Psychiatry. 2021;12(1551). doi:10.3389/fpsyt.2021.720082
24. Stefano GB, Buttiker P, Kream RM. Reassessment of the blood-brain barrier: a potential target for viral entry into the immune-privileged brain. Germs. 2022;12(1):99–101. doi:10.18683/germs.2022.1310
25. Levine A, Sacktor N, Becker JT. Studying the neuropsychological sequelae of SARS-CoV-2: lessons learned from 35 years of neuroHIV research. J Neurovirol. 2020;26(6):809–823. doi:10.1007/s13365-020-00897-2
26. Stefano GB. Historical insight into infections and disorders associated with neurological and psychiatric sequelae similar to long COVID. Med Sci Monit. 2021;27:e931447. doi:10.12659/MSM.931447
27. Mayer KA, Stockl J, Zlabinger GJ, Gualdoni GA. Hijacking the supplies: metabolism as a novel facet of virus-host interaction. Front Immunol. 2019;10:1533. doi:10.3389/fimmu.2019.01533
28. Kirsch-Volders M, Fenech M. Inflammatory cytokine storms severity may be fueled by interactions of micronuclei and RNA viruses such as COVID-19 virus SARS-CoV-2. A hypothesis. Mutat Res Rev Mutat Res. 2021;788:108395. doi:10.1016/j.mrrev.2021.108395
29. Ollig J, Kloubert V, Taylor KM, Rink L. B cell activation and proliferation increase intracellular zinc levels. J Nutr Biochem. 2019;64:72–79. doi:10.1016/j.jnutbio.2018.10.008
30. Sanchez EL, Lagunoff M. Viral activation of cellular metabolism. Virology. 2015;479–480:609–618. doi:10.1016/j.virol.2015.02.038
31. Carty M, Guy C, Bowie AG. Detection of Viral Infections by Innate Immunity. Biochem Pharmacol. 2021;183:114316. doi:10.1016/j.bcp.2020.114316
32. Shytaj IL, Procopio FA, Tarek M, et al. Glycolysis downregulation is a hallmark of HIV-1 latency and sensitizes infected cells to oxidative stress. EMBO Mol Med. 2021;13(8):e13901. doi:10.15252/emmm.202013901
33. Duette G, Pereyra Gerber P, Rubione J, et al. Induction of HIF-1alpha by HIV-1 infection in CD4(+) T cells promotes viral replication and drives extracellular vesicle-mediated inflammation. mBio. 2018;9(5). doi:10.1128/mBio.00757-18
34. Kang S, Tang H. HIV-1 infection and glucose metabolism reprogramming of T cells: another approach toward functional cure and reservoir eradication. Front Immunol. 2020;11:572677. doi:10.3389/fimmu.2020.572677
35. Haroon E, Miller AH, Sanacora G. Inflammation, glutamate, and glia: a trio of trouble in mood disorders. Neuropsychopharmacology. 2017;42(1):193–215. doi:10.1038/npp.2016.199
36. Murphy MP. How mitochondria produce reactive oxygen species. Biochem J. 2009;417(1):1–13. doi:10.1042/BJ20081386
37. Kanellopoulos AK, Mariano V, Spinazzi M, et al. Aralar sequesters GABA into hyperactive mitochondria, causing social behavior deficits. Cell. 2020;180(6):1178–1197 e20. doi:10.1016/j.cell.2020.02.044
38. Bharadwaj S, Singh M, Kirtipal N, Kang SG. SARS-CoV-2 and glutamine: SARS-CoV-2 triggered pathogenesis via metabolic reprograming of glutamine in host cells. Front Mol Biosci. 2020;7:627842. doi:10.3389/fmolb.2020.627842
39. Stefano GB, Kream RM. Mitochondrial DNA heteroplasmy as an informational reservoir dynamically linked to metabolic and immunological processes associated with COVID-19 neurological disorders. Cell Mol Neurobiol. 2022;42(1):99–107. doi:10.1007/s10571-021-01117-z
40. Stefano GB, Kream RM. Viruses broaden the definition of life by genomic incorporation of artificial intelligence and machine learning processes. Curr Neuropharmacol. 2022;20:1888–1893. doi:10.2174/1570159X20666220420121746
41. Majolo F, Silva GLD, Vieira L, et al. Neuropsychiatric disorders and COVID-19: what we know so far. Pharmaceuticals. 2021;14(9):Sep. doi:10.3390/ph14090933
42. Iadecola C, Anrather J, Kamel H. Effects of COVID-19 on the nervous system. Cell. 2020;183(1):16–27e1. doi:10.1016/j.cell.2020.08.028
43. Wenzel J, Lampe J, Muller-Fielitz H, et al. The SARS-CoV-2 main protease M(pro) causes microvascular brain pathology by cleaving NEMO in brain endothelial cells. Nat Neurosci. 2021;24(11):1522–1533. doi:10.1038/s41593-021-00926-1
44. Bodnar B, Patel K, Ho W, Luo JJ, Hu W. Cellular mechanisms underlying neurological/neuropsychiatric manifestations of COVID-19. J Med Virol. 2021;93(4):1983–1998. doi:10.1002/jmv.26720
45. Hosen I, Al-Mamun F, Mamun MA. Prevalence and risk factors of the symptoms of depression, anxiety, and stress during the COVID-19 pandemic in Bangladesh: a systematic review and meta-analysis. Glob Ment Health. 2021;8:e47. doi:10.1017/gmh.2021.49
46. Alzahrani F, Alshahrani NZ, Abu sabah A, Zarbah A, Abu Sabah S, Mamun MA. Prevalence and factors associated with mental health problems in Saudi general population during the coronavirus disease 2019 pandemic: a systematic review and meta-analysis. Psych J. 2022;11(1):18–29. doi:10.1002/pchj.516
47. Rogers JP, Chesney E, Oliver D, et al. Psychiatric and neuropsychiatric presentations associated with severe coronavirus infections: a systematic review and meta-analysis with comparison to the COVID-19 pandemic. Lancet Psychiatry. 2020;7(7):611–627. doi:10.1016/S2215-0366(20)30203-0
48. Taquet M, Geddes JR, Husain M, Luciano S, Harrison PJ. 6-month neurological and psychiatric outcomes in 236 379 survivors of COVID-19: a retrospective cohort study using electronic health records. Lancet Psychiatry. 2021;8(5):416–427. doi:10.1016/S2215-0366(21)00084-5
49. Singer EJ, Thames AD. Neurobehavioral manifestations of human immunodeficiency virus/AIDS: diagnosis and treatment. Neurol Clin. 2016;34(1):33–53. doi:10.1016/j.ncl.2015.08.003
50. Alvarez-Carbonell D, Ye F, Ramanath N, et al. Cross-talk between microglia and neurons regulates HIV latency. PLoS Pathog. 2019;15(12):e1008249. doi:10.1371/journal.ppat.1008249
51. Celesia BM, Nigro L, Pinzone MR, et al. High prevalence of undiagnosed anxiety symptoms among HIV-positive individuals on cART: a cross-sectional study. Eur Rev Med Pharmacol Sci. 2013;17(15):2040–2046.
52. Smith MY, Egert J, Winkel G, Jacobson J. The impact of PTSD on pain experience in persons with HIV/AIDS. Pain. 2002;98(1–2):9–17. doi:10.1016/s0304-3959(01)00431-6
53. Shacham E, Morgan JC, Onen NF, Taniguchi T, Overton ET. Screening anxiety in the HIV clinic. AIDS Behav. 2012;16(8):2407–2413. doi:10.1007/s10461-012-0238-6
54. Ye J, Wen Y, Chu X, et al. Association between herpes simplex virus 1 exposure and the risk of depression in UK Biobank. Clin Transl Med. 2020;10(2):e108. doi:10.1002/ctm2.108
55. van Heugten-van der Kloet D, van Heugten T. The classification of psychiatric disorders according to DSM-5 deserves an internationally standardized psychological test battery on symptom level. Front Psychol. 2015;6:1108. doi:10.3389/fpsyg.2015.01108
56. Mamun MA. Exploring factors in fear of COVID-19 and its GIS-based nationwide distribution: the case of Bangladesh. BJPsych Open. 2021;7(5):e150. doi:10.1192/bjo.2021.984
57. Al Mamun F, Hosen I, Misti JM, Kaggwa MM, Mamun MA. Mental disorders of bangladeshi students during the COVID-19 pandemic: a systematic review. Psychol Res Behav Manag. 2021;14:645–654. doi:10.2147/PRBM.S315961
58. Taylor MA, Vaidya NA. Psychopathology in neuropsychiatry: DSM and beyond. J Neuropsychiatry Clin Neurosci. 2005;17(2):246–249. doi:10.1176/jnp.17.2.246
59. Saladino V, Algeri D, Auriemma V. The psychological and social impact of covid-19: new perspectives of well-being. Front Psychol. 2020;11:577684. doi:10.3389/fpsyg.2020.577684
60. Tolentino JC, Schmidt SL. DSM-5 criteria and depression severity: implications for clinical practice. original research. Front Psychiatry. 2018;9(450). doi:10.3389/fpsyt.2018.00450
61. Segerstrom SC, Miller GE. Psychological stress and the human immune system: a meta-analytic study of 30 years of inquiry. Psychol Bull. 2004;130(4):601–630. doi:10.1037/0033-2909.130.4.601
62. Buttiker P, Weissenberger S, Ptacek R, Stefano GB. Interoception, trait anxiety, and the gut microbiome: a cognitive and physiological model. Med Sci Monit. 2021;27:e931962. doi:10.12659/MSM.931962
63. Srivastava KK, Kumar R. Stress, oxidative injury and disease. Indian J Clin Biochem. 2015;30(1):3–10. doi:10.1007/s12291-014-0441-5
64. Schiavone S, Jaquet V, Trabace L, Krause KH. Severe life stress and oxidative stress in the brain: from animal models to human pathology. Antioxid Redox Signal. 2013;18(12):1475–1490. doi:10.1089/ars.2012.4720
65. Simioni C, Zauli G, Martelli AM, et al. Oxidative stress: role of physical exercise and antioxidant nutraceuticals in adulthood and aging. Oncotarget. 2018;9(24):17181–17198. doi:10.18632/oncotarget.24729
66. Erickson MA, Rhea EM, Knopp RC, Banks WA. Interactions of SARS-CoV-2 with the blood-brain barrier. Int J Mol Sci. 2021;22(5):2681. doi:10.3390/ijms22052681
67. Rhea EM, Logsdon AF, Hansen KM, et al. The S1 protein of SARS-CoV-2 crosses the blood-brain barrier in mice. Nat Neurosci. 2021;24(3):368–378. doi:10.1038/s41593-020-00771-8
68. Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, Katze MG. Into the eye of the cytokine storm. Microbiol Mol Biol Rev. 2012;76(1):16–32. doi:10.1128/MMBR.05015-11
69. Cox MA, Kahan SM, Zajac AJ. Anti-viral CD8 T cells and the cytokines that they love. Virology. 2013;435(1):157–169. doi:10.1016/j.virol.2012.09.012
70. Stefano GB, Pilonis N, Ptacek R, Raboch J, Vnukova M, Kream RM. Gut, microbiome, and brain regulatory axis: relevance to neurodegenerative and psychiatric disorders. Cell Mol Neurobiol. 2018;38(6):1197–1206. doi:10.1007/s10571-018-0589-2
71. Putra RD, Lyrawati D. Interactions between bacteriophages and eukaryotic cells. Scientifica. 2020;2020:3589316. doi:10.1155/2020/3589316
72. Van Belleghem JD, Dabrowska K, Vaneechoutte M, Barr JJ, Bollyky PL. Interactions between bacteriophage, bacteria, and the mammalian immune system. Viruses. 2018;11(1):10. doi:10.3390/v11010010
73. Ma Q, Xing C, Long W, Wang HY, Liu Q, Wang RF. Impact of microbiota on central nervous system and neurological diseases: the gut-brain axis. J Neuroinflammation. 2019;16(1):53. doi:10.1186/s12974-019-1434-3
74. Garry RF. Ebola virus can lie low and reactivate after years in human survivors. Nature. 2021;597(7877):478–480. doi:10.1038/d41586-021-02378-w
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.