")
Back to Journals » Drug Design, Development and Therapy » Volume 14
Hirudin Protects Against Kidney Damage in Streptozotocin-Induced Diabetic Nephropathy Rats by Inhibiting Inflammation via P38 MAPK/NF-κB Pathway
Authors Han J, Pang X , Zhang Y, Peng Z, Shi X, Xing Y
Received 9 April 2020
Accepted for publication 26 June 2020
Published 7 August 2020 Volume 2020:14 Pages 3223—3234
DOI https://doi.org/10.2147/DDDT.S257613
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Qiongyu Guo
Jiarui Han,1,2 Xinxin Pang,1,2 Yage Zhang,1,2 Zining Peng,1,2 Xiujie Shi,1,2 Yufeng Xing1,2
1Department of Nephropathy, Henan Provincial Hospital of Traditional Chinese Medicine, Zhengzhou, Henan Province, People’s Republic of China; 2Department of Nephropathy, The Second Hospital Affiliated to Henan University of Chinese Medicine, Zhengzhou, Henan Province, People’s Republic of China
Correspondence: Jiarui Han; Xinxin Pang
The Second Hospital Affiliated to Henan University of Chinese Medicine, 6, Dong Feng Road, Jinshui District, Zhengzhou, Henan Province 450002, People’s Republic of China
Tel +86-371-60979837
Email [email protected]; [email protected]
Background: Inflammation-induced podocyte apoptosis plays an important role in kidney injury during diabetic nephropathy (DN). Hirudin (HIR), a natural compound extracted from leeches, can inhibit inflammation. However, whether HIR can protect the kidneys against inflammation during DN is unknown. In the present study, we aimed to study the effects of HIR on kidney damage in a DN rat model and explore its anti-inflammatory properties.
Methods: A streptozotocin-induced DN rat model was generated, and HIR was administered subcutaneously. Immortal podocytes and primary peritoneal macrophages were used for vitro studies. Hematoxylin and eosin staining was used to evaluate renal pathological changes; quantitative polymerase chain reaction and immunoblotting were used to detect gene expression; and TUNEL staining was used to detect apoptotic cells.
Results: Our results showed that HIR protected against renal injury, as indicated by kidney weight/body weight, serum creatinine, renal pathological changes, blood urea nitrogen, and detection of urine proteins. Notably, HIR treatment reduced macrophage infiltration, pro-inflammatory cytokine expression, and podocyte apoptosis in the kidney tissues of DN rats. In vitro, high glucose (HG) induced the activation of M1 macrophages, which was accompanied by increased podocyte apoptosis. HIR could decrease HG-induced podocyte apoptosis and suppress pro-inflammatory cytokine expression in podocytes in vitro. This was achieved via inhibition of p38 MAPK/NF-κB activation in renal tissues and podocytes.
Conclusion: HIR could inhibit inflammation via the p38 MAPK/NF-κB pathway, prevent podocyte apoptosis, and protect against kidney damage in a DN rat model.
Keywords: hirudin, inflammation, apoptosis, diabetic nephropathy, kidney damage
Introduction
Diabetic nephropathy (DN) is one of the most serious microvascular complications of diabetes mellitus (DM) and a common cause of end-stage renal disease (ESRD).1 Data show that about 40% of patients with DM have varying degrees of DN, and ~25% of them will progress to uremia.1,2 Currently, DN is considered as a chronic inflammatory disease caused by various metabolic disorders. Therefore, inflammation plays a pivotal role in the development and progression of DN.3 Notably, previous studies have identified that inhibiting inflammation can protect the kidneys during DN and that p38/NF-κB is usually found to be involved in the regulation of inflammation.4,5 Wagner et al found that beraprost sodium was protective against DN by inhibiting the inflammation of kidney tissues in high-fat diet/streptozotocin-induced diabetic rats. Moreover, the molecular mechanism of protection was potentially related to the inhibition of the p38-MAPK signaling pathway;4 Zhang et al found that oridonin could not only improve renal function in DN rats, but could also inhibit the proliferation of mesangial cells. Again, the molecular mechanisms underlying this were found to be the suppression of inflammation via inhibition of the p38/NF-κB pathway.5 Therefore, the identification of drugs that can protect against kidney damage and limit inflammation in DN is of vital importance.
Hirudin (HIR) is an active ingredient extracted from the salivary glands of leeches. It is a small protein composed of 65–66 amino acids and has been found to be the strongest natural specific inhibitor of thrombin discovered to date.6,7 Interestingly, previous studies have found that HIR can exert anti-inflammatory effects via modulation of the p38/NF-κB pathway.8,9 Liu et al found that hirudin could significantly enhance flap viability and reduce inflammatory responses compared with a control group. This was achieved by inhibiting the expression of p-p38/p38, p-NF-κB p65/NF-κB p65, TNF-α, IL-6, and ICAM-1 levels.8 Additionally, hirudin was also found to inhibit the proliferation of human gliomas by blocking the ERK/MAPK pathway. However, we know very little about the ability of HIR to protect against inflammatory injury of the kidneys in DN. In the present study, we aimed to study the effect of HIR on ameliorating kidney damage and inhibiting inflammation via a p38/NF-κB pathway in a DN rat model.
Materials and Methods
Experimental Animal and Grouping
Here, we used 28 Sprague Dawley rats (half of which were male, 6−8 weeks old, 180–220 g) to conduct the animal tests. The 28 rats were divided into four groups with seven rats per group as follows: Control group, Control + HIR group, DN group, and DN + HIR group. Rats in the DN and DN + HIR groups were injected intraperitoneally with 100 mg/kg streptozotocin (STZ) (cat# 572,201, Sigma, USA)10 and rats in the Control and Control + HIR groups were injected intraperitoneally with the same amount of saline buffer. Three days after STZ injection, we measured blood glucose via the tail vein and rats with blood glucose of >16.7 mmol/L were considered DN rats. Rats in the Control + HIR and DN + HIR groups were subcutaneously injected with 5 U of hirudin11 (cat# 94,581, Sigma, USA) daily for 8 weeks. Rats in the Control and DN groups were subcutaneously injected with the same amount of saline buffer.
The animal experiments in this study were approved and supervised by the Animal Care and Use Committee in Henan Provincial Hospital of Traditional Chinese Medicine and were conducted according to the guidelines of the National Institution of Health.
The Quantification of Biochemical Markers
Eight weeks after treating with HIR, we harvested blood from the orbital venous plexus and centrifuged the blood to collect serum. We also harvested all urine for 24 hours. Serum glucose, serum creatinine, blood urea nitrogen, and 24-hour urine protein were quantified using a blood glucose meter (Bayer, Germany), SCR enzyme-linked immunosorbent assay (ELISA) kit (cat# YS04392B, YaJi Biological, China), BUN ELISA kit (cat# C013-2, YaJi Biological, China) and protein concentration assay kit (cat# K812-1000, AmyJet, China), respectively.
Histological Analysis of Kidney Sections
We euthanized the rats by cervical dislocation and confirmed death in the rats by observing respiration and heartbeat. We harvested the kidney tissue of the rats and prepared 5-μm histological sections as described previously.12 For hematoxylin and eosin (H&E) staining, frozen slices of rat kidney were slightly dried, stained with hematoxylin for 3 min, eosin for 10 seconds, dehydrated and sealed with neutral gum. For immunochemistry analysis, primary anti-CD68 (cat# 97,778, Cell Signaling Technology, USA) was incubated overnight at 4°C, and the second antibody (cat# 18,653, Cell Signaling Technology, USA) was incubated for 1 hour at room temperature. To quantify apoptosis, a TUNEL assay kit (cat# 11,966,006,001, Merck-Calbiochem, USA) was used as per the manufacturer’s instructions. Image J 3.0 (IBM, USA) was used to analyze the glomerular area and quantify the number CD68 positive and apoptotic cells.
RT-qPCR Analysis
We used an animal tissue/cell RNA extraction kit (cat# CW0580S, NuoYang Biological, China) to extract total RNA from tissues and cells according to manufacturer’s instructions. cDNA was prepared using a real-time (RT) reagent kit (cat# RR047A, Takara, Japan) according to manufacturer’s instructions. We prepared 20 μL of the RT-qPCR system as described in the quantitative polymerase chain reaction (qPCR) master mix kit instructions (cat# A600A, Promega, USA). The relative expression of genes was calculated by the 2–ΔΔCt method, and β-actin was used as a loading control. The primers for qPCR are shown in Table 1.
 |
Table 1 Sequence of qPCR Primers |
Cytokine ELISAs
The concentration of cytokines in tissue homogenates or cell culture medium was quantified via ELISA. For tissues, we added pre-chilled homogenate buffer solution (0.01 mol/L Tris–HCl, 0.1 mmol/L EDTA-2Na, 0.01 mol/L) to 1 g of tissue and used a homogenizer to obtain tissue homogenate. Homogenates were centrifuged (1500 x g, 15 minutes, 4°C) to collect supernatant for detection of cytokine concentrations. To detect cytokine concentrations in cell culture medium, we directly centrifuged the supernatant (1500 x g, 15 minutes, 4°C). The following ELISA kits were used for tissues: TNF-α (cat# K1052-100, AmyJet, China), IL-1β (cat# E-EL-R0012c, Elabscience, USA), and IL-6 (cat#ab119548, Abcam, UK); or for cell culture medium: TNF-α (cat# 208,348, Abcam, UK), IL-1β (cat# ab100705, Abcam, USA) and IL-6 (cat# ab10073, Abcam, UK).
Quantification of Apoptosis
Immortal podocytes (mouse) established by Professor Peter Mundel (Albert Einstein College of Medicine, New York, USA) and primary peritoneal macrophages were prepared and cultured according to previously described methods.13 In this study, cells were cultured with low glucose (1 g/L) Dulbecco's Modified Eagle Medium (DMEM) (cat# 10,567,022, Gibco, USA) supplemented with 10% fetal bovine serum (FBS) (cat# 16,140,071, Gibco, USA) at 37°C with 5% CO2, which was defined as a normal-glucose (NG) environment. A high-glucose (HG) environment was defined by the addition of 3 g/L glucose to NG medium.14 In a macrophage/podocyte co-culture system, we seeded 2 x 105 podocytes in the lower chamber of a polycarbonate insert cell culture device. We seeded 0 (HG group), 2 x 105 (HG + 1M group), 4 x 105 (HG + 2M group), or 8 x 105 (HG + 4M group) macrophages in the upper chamber and cultured for 48 hours. To analyze HIR treatment, we added 5 U/mL HIR to the HG group (HG + HIR group) and added 5 U/mL HIR to the HG + 4M group (HG + 4M + HIR group). In a culture of podocytes alone, 1 x 105 podocytes were seeded in a six-well culture dish. After 24 hours, we changed the cell culture medium to a HG medium and administered drugs (5 U/mL HIR, 5 μmol/L BAY11708215 or 20 μmol/L SB20358016) for 48 hours. Finally, cells were harvested to detect apoptosis using an Annexin V-FITC/PI Apoptosis Detection Kit (cat# 40302ES20, Yeasen, China) according to the manufacturer’s instructions.
Western Blot
Total protein was extracted from tissues and cells using a total protein extraction kit (cat# AMJ-KT0007, AmyJet, China) and proteins were quantified using a BCA kit (cat# KTD3001, AmyJet, China). Next, 50 μg total protein was separated on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel as described previously.17 In brief, after blocking with a 5% milk solution for 1 hour at room temperature, the primary antibodies (all Abcam, UK) to iNOS (cat# ab15232), CD206 (cat# ab8918), p38 (cat# ab170099), p-p38 (cat# 47,363), p65 (cat# 16,502), and p-p65 (cat# 86,299) were incubated overnight at 4°C. The secondary antibodies (cat# 6721 and # ab6789, Abcam, UK) were incubated for 1 hour at room temperature. We used Image J v3.0 (IBM, USA) to analyze the gray value and β-actin was used as a loading control.
Statistical Analysis
Data in the present study were analyzed by Graphpad prism v8.3.0. P values were calculated by Student’s t-test when only comparing two groups. For multiple groups, the P value was calculated by one-way analysis of variance (ANOVA) with Tukey post-hoc test. P < 0.05 indicated a significant difference.
Results
HIR Improved Renal Function and Reduced Renal Pathological Changes
In this study, we established a DN rat model by intraperitoneal injection of STZ and assessed the productive effects of subcutaneous HIR injections. After 8 weeks of treatment with HIR, we weighed the rats, harvested their blood, and collected their urine for 24 hours before they were euthanized. Compared with the rats in the control group, the average bodyweight of rats in the DN group was significantly lower and the serum glucose was significantly higher (Figure 1A and B). Notably, following an 8-week treatment with HIR, the weights of DN rats were significantly increased and the serum glucose of DN rats was significantly decreased. Investigating renal biochemical markers, we noted that the ratio of kidney weight/body weight, serum creatinine, blood urea nitrogen, and proteins in the urine of rats in the DN group were all significantly higher compared to the control group (Figure 1C–F). Notably, HIR treatment could significantly reduce these elevated renal biochemical markers in DN rats.
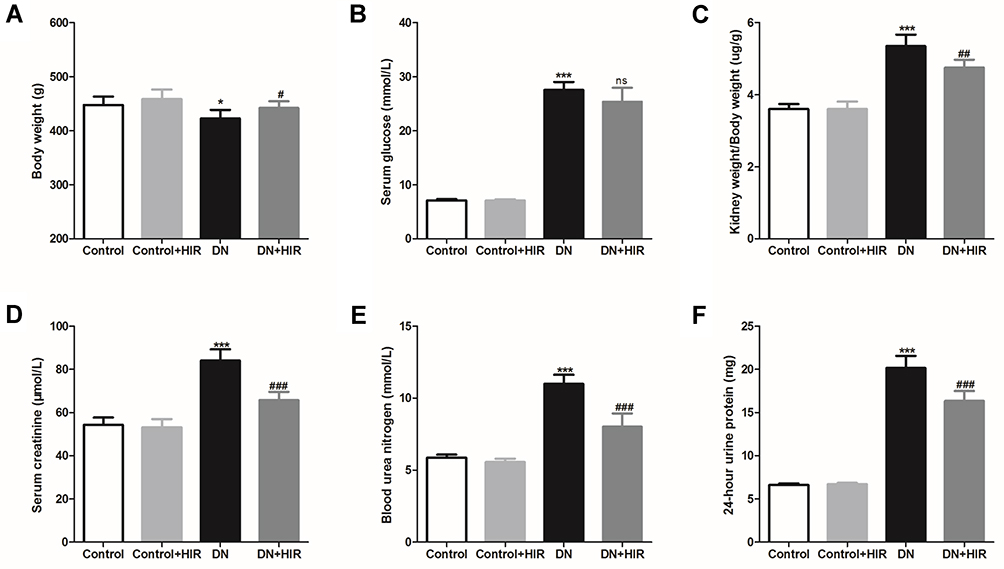 |
Figure 1 Effect of HIR on kidney biochemical markers in DN rats. (A–F) Body weight (A), serum glucose (B), kidney weight/body weight (C), serum creatinine (D), blood urea nitrogen (E), and 24-hour urine protein (F) were measured and compared in different group. Seven rats per group. *Indicates P < 0.05 and ***indicates P < 0.001 vs control group; ns indicates no significance, #indicates P < 0.05, ##indicates P < 0.01 and ###indicates P < 0.001 vs DN group. |
Additionally, we assessed differences in the renal pathology between groups using H&E staining (Figure 2A,B). The rats in the DN group displayed glomerular bulging, mesangial cell proliferation, mesangial thickening, and small renal capsule sac cavity, but these pathological changes were significantly reduced in DN rats treated with HIR (Figure 2C,D). Quantitative histopathology showed that the glomerular area of rats in the DN group was significantly greater compared to the control group and HIR could significantly reduce this elevated glomerular area in DN rats (Figure 2E).
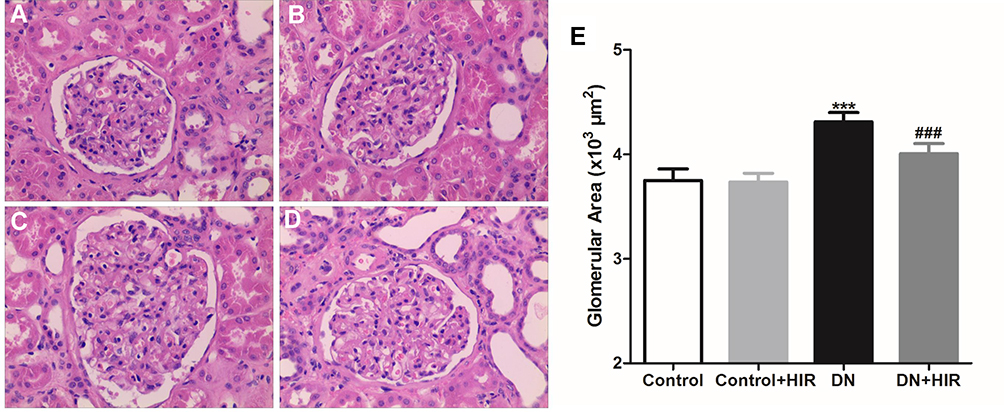 |
Figure 2 Effect of HIR on renal pathological changes in DN rats. (A–E) H&E staining was used to assess the changes of renal pathological in rat of control (A), control + HIR (B), DN (C), and DN + HIR (D) groups, and glomerular area was also detected and compared in different groups (E). Seven rats per group and each rat measures at least 10 glomeruli area. ***Indicates P < 0.001 vs control group, and ###indicates P < 0.001 vs DN group. |
HIR Reduced Macrophage Infiltration into Renal Tissues
Macrophage infiltration is a hallmark of pathology in the early stages of DN and one of the main causes of renal damage.18 We detected CD68 expression in kidney tissue by immunohistochemistry to indicate the quantity of macrophage infiltration and found that there was negligible macrophage infiltration in the glomeruli of normal rats (Figure 3A,B). Conversely, macrophage infiltration was identified in all glomeruli in DN rats (Figure 3C,D). Moreover, the number of CD68+ cells in rats in the DN group were significantly higher than the control group and the number of CD68+ cells in DN rats significantly decreased after treatment with HIR (Figure 3E).
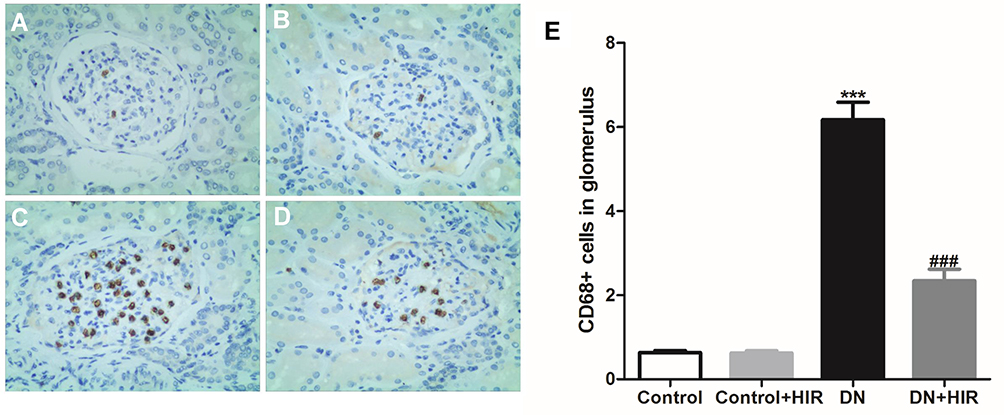 |
Figure 3 Effect of HIR on macrophage infiltration in glomerulus of DN rats. (A–E) CD68 protein was detected in glomerulus of control (A), control + HIR (B), DN (C), and DN + HIR (D) groups, and CD68+ positive cells were quantified (E). Seven rats per group and 10 visual fields for each slice for statistics. ***Indicates P < 0.001 vs control group, and ###indicates P < 0.001 vs DN group. |
HIR Inhibited Proinflammatory Cytokine Expression in Renal Tissues
Macrophages are not only recruited by proinflammatory cytokines, they also secrete proinflammatory cytokines. We harvested the renal tissues and detected the expression of proinflammatory cytokines in DN rats after treatment with HIR and found that the expression of TNF-α, IL-1β, and IL-6 mRNA in the renal tissues of rats in the DN group were all significantly higher than those in the control group (Figure 4A–C). HIR treatment could significantly reduce the elevated mRNA expression of proinflammatory cytokines in renal tissues of DN rats. Similarly, the protein concentrations of pro-inflammatory cytokines were quantified by ELISA. We found that TNF-α, IL-1β, and IL-6 protein in renal tissues of rats in the DN group were also all significantly higher than in rats in the control group (Figure 4D–F). HIR could significantly reduce the elevated production of these pro-inflammatory cytokines in the renal tissues of DN rats.
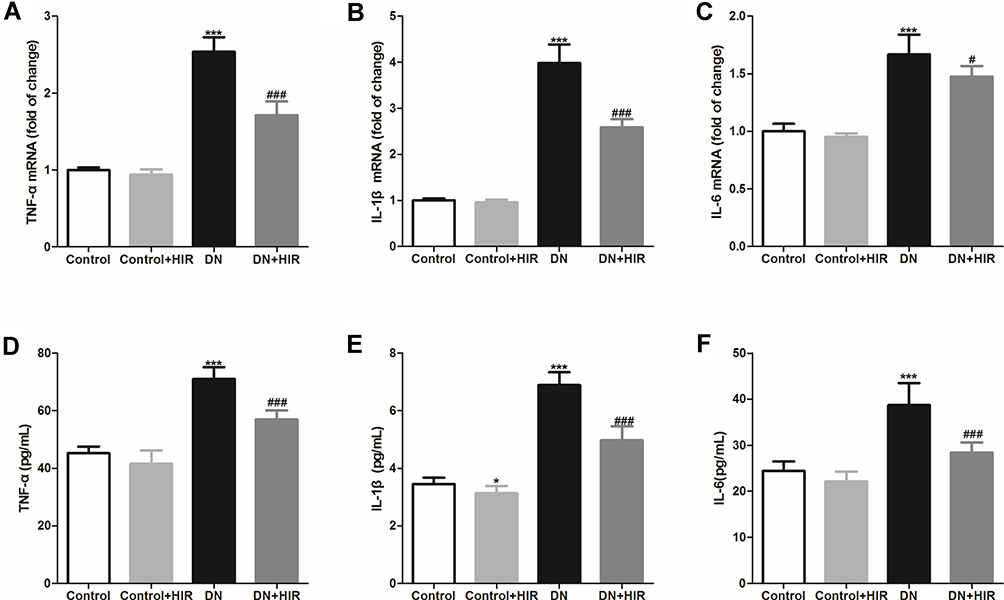 |
Figure 4 Effect of HIR on production of pro-inflammatory cytokines in renal tissues. (A–C) qPCR analysis of the indicated TNF-α (A), IL-1β (B), and IL-6 (C) mRNA in renal tissues of rats; (D–F) ELISA assay of the indicated TNF-α (D), IL-1β (E), and IL-6 (F) protein in renal tissues of rats. Seven rats per group. *Indicates P < 0.05 and ***indicates P < 0.001 vs control group; #indicates P < 0.05 and ###indicates P < 0.001 vs DN group. |
HIR Reduced Apoptosis in Renal Tissues
Podocyte apoptosis occurs in the early stages of DN and has been shown to be positively correlated with proteinuria and the decrease in renal function observed in patients with DN.19,20 Therefore, apoptosis was quantified in renal tissues using TUNEL staining. We found that there was negligible apoptosis in the glomeruli of control rats (Figure 5A,B). Conversely, high numbers of apoptotic cells were identified in all glomeruli in DN rats (Figure 5C,D). The number of apoptotic cells in rats of the DN group was significantly higher than in rats in the control group. Moreover, the numbers of apoptotic cells in DN rats significantly decreased following treatment with HIR (Figure 5E).
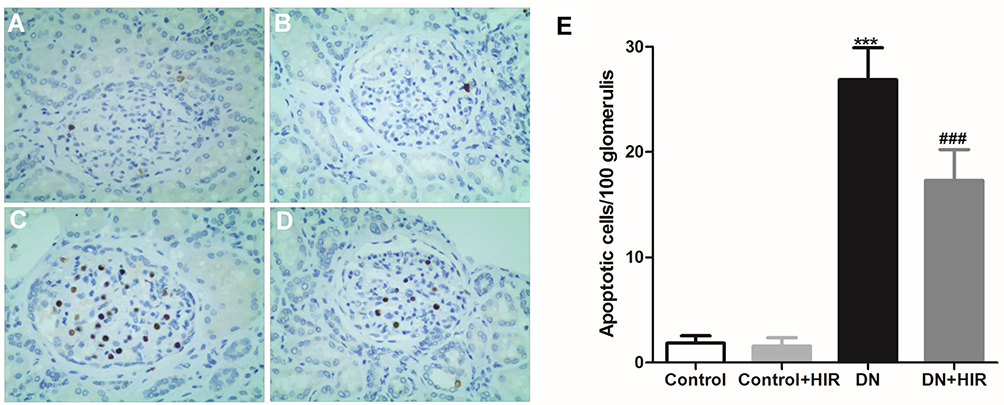 |
Figure 5 Effect of HIR on apoptosis in renal tissues. (A–E), TUNEL staining was used to detected the apoptotic cells in renal tissues of control (A), control + HIR (B), DN (C), and DN + HIR (D) group, and TUNEL positive cells were quantified (E). Seven rats per group and 10 visual fields for each slice for statistics. ***Indicates P < 0.001 vs control group, and ###indicates P < 0.001 vs DN group. |
HIR Reduced Podocyte Apoptosis When Co-Cultured with Macrophages in a High-Glucose Environment
To investigate the effect of macrophage infiltration on podocyte apoptosis in DN rats, we established a macrophage/podocyte co-culture system in a high-glucose (HG) environment. First, we found that the expression of iNOS mRNA in macrophages in an HG environment was significantly higher than macrophages in an NG environment, while CD206 mRNA was significantly lower (Figure 6A). The same trend was also found for the protein expression of iNOS and CD206 (Figure 6B). Moreover, compared with the NG environment, HG increased the expression of TNF-α, IL-1β, and IL-6 mRNA (Figure 6C). The apoptosis of podocytes significantly increased in a macrophage/podocyte co-culture system compared to podocytes alone in an HG environment (Figure 6D). Importantly, HIR could significantly inhibit the apoptosis of podocytes in HG. We also identified that the proportion of podocyte apoptosis was directly related to the number of macrophages in the macrophage/podocyte co-culture system, that is, the greater number of macrophages, the greater number of apoptotic podocytes (Figure 6D). However, HIR could also reduce HG-induced podocyte apoptosis in a macrophage/podocyte co-culture system (Figure 6D).
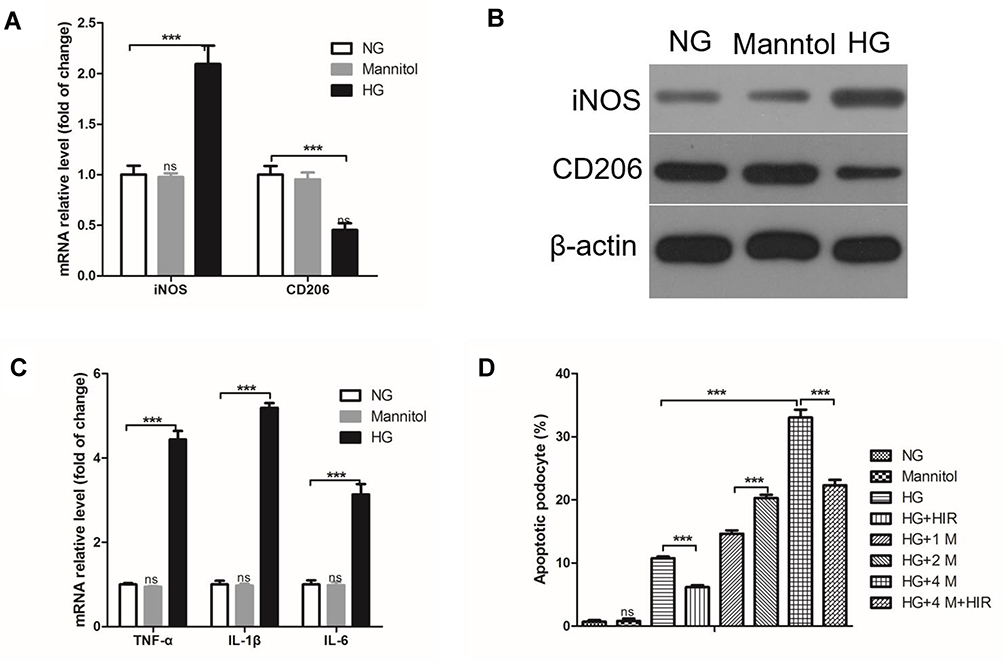 |
Figure 6 Effect of HIR on the apoptosis of podocytes co-cultured with macrophages in a high-glucose environment. (A) qPCR analysis of the indicated iNOS and CD206 mRNA in macrophages; (B) Western blot was used to detect the expression of iNOS and CD206 protein in macrophages; (C) HG induced increased expression of TNF-α, IL-1β, and IL-6 in macrophages; (D) apoptotic podocytes were quantified and compared. 1M, 2M, and 3M indicate the ratio of macrophages to podocytes is 1, 2, and 4, respectively. When comparing between two groups, ***indicates P < 0.001.Abbreviations: NG, normal glucose; HG, high glucose. |
HIR Suppressed Pro-Inflammatory Cytokine Expression in Podocytes
Initially, we found that the expression of TNF-α, IL-1β, and IL-6 mRNA and protein in podocytes was significantly increased in HG conditions and HIR could significantly reduce the production of these elevated pro-inflammatory cytokines (Figure 7). In order to explore the molecular mechanism underlying the anti-inflammatory effects of HIR, we used two compounds, SB203580 (an inhibitor of the p38 pathway) and BAY117085 (an inhibitor of NF-κB). We found that treatment with SB203580 or BAY117085 alone could also reduce the expression of HG-induced TNF-α, IL-1β, and IL-6 in podocytes, but their ability to inhibit these cytokines was lower compared to HIR. However, the combined treatment of SB203580 and BAY117085 could exert the same inhibitory effect on HG-induced expression of TNF-α, IL-1β, and IL-6 in podocytes compared to HIR.
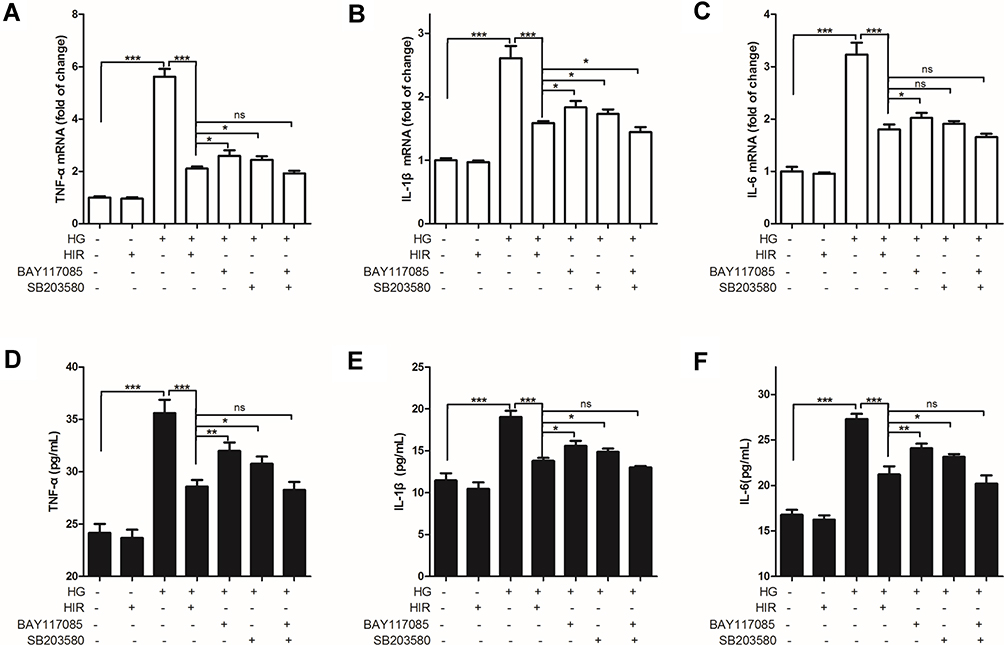 |
Figure 7 Effect of HIR on production of pro-inflammatory cytokines in podocytes. (A–C) qPCR analysis of the indicated TNF-α (A), IL-1β (B), and IL-6 (C) mRNA in podocytes; (D, F) ELISA assay of the indicated TNF-α (D), IL-1β (E), and IL-6 (F) protein in podocytes. Three independent replicates. When comparing between two groups, ns indicates no significance, *indicates P < 0.05, **indicates P < 0.01 and ***indicates P < 0.001. |
HIR Inhibited p38 Pathway and NF-κB Activation in Renal Tissues and Podocytes
First, we found that the expression of p-p38/p38, and p-p65/p65 proteins in the renal tissues of rats in the DN group were all significantly higher than those in rats in the control group. Notably, HIR could significantly reduce the expression of p-p38/p38 and p-p65/p65 protein in renal tissues of DN rats (Figure 8). In vitro, HG also induced the elevated expression of p-p38/p38 and p-p65/p65 protein in podocytes and HIR could significantly inhibit the expression of these proteins (Figure 9A). Importantly, HIR could exert the same inhibitory effect on HG-induced expression of p-p38/p38 and p-p65/p65 protein in podocytes as SB203580 and BAY117085, respectively (Figure 9B,C).
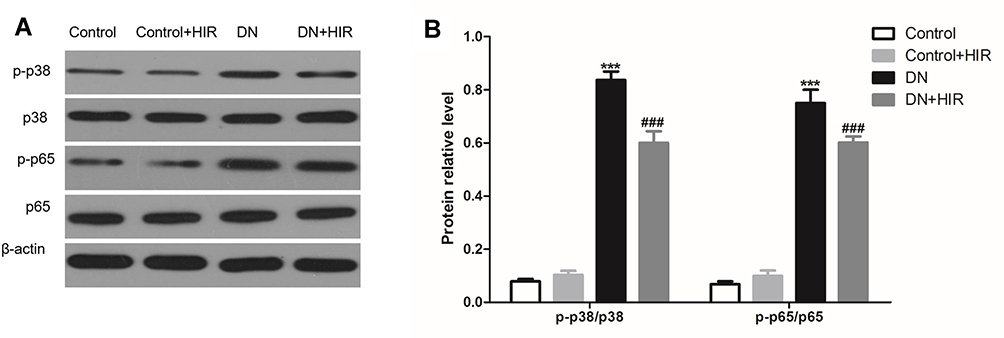 |
Figure 8 Effect of HIR on p38/NF-κB pathway in renal tissues. (A, B) Representative protein bands of p38, p-p38, p65, and p-p65 (A), and statistical comparison of gray values (B). Seven rats per group. ***Indicates P < 0.001 vs control group, and ###indicates P < 0.001 vs DN group. |
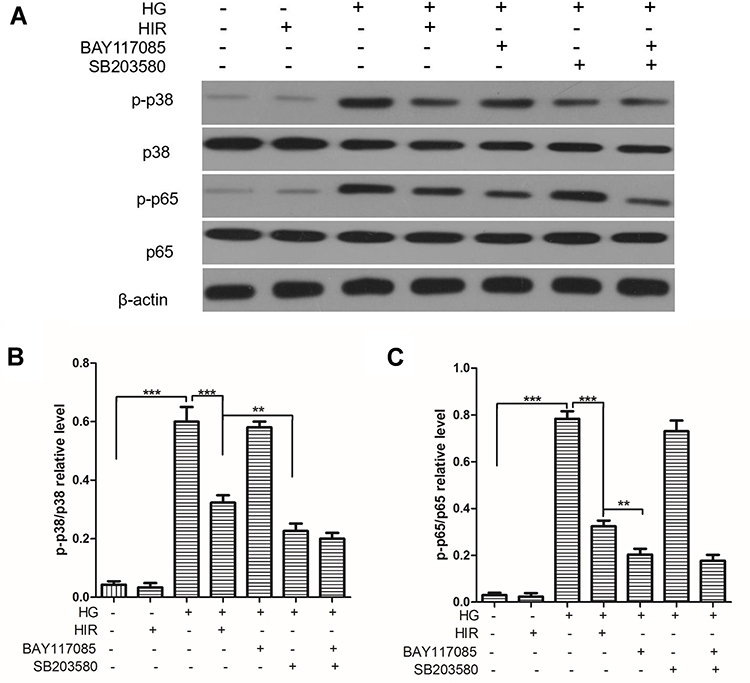 |
Figure 9 Expression of key proteins on p38/NF-kB pathway in podocytes. (A, B) Representative protein bands of p38, p-p38, p65, and p-p65 (A), and statistical comparison of gray values (B, C). When comparing between two groups, **indicates P < 0.01 and ***indicates P < 0.001. |
Discussion
HIR is the strongest natural thrombin inhibitor found to date and thrombin has been found to play an important role in kidney disease and regulation of inflammation. Thrombin was found to induce kidney damage and inhibition of thrombin could protect kidney function. Madhusudhan et al thought that coagulation proteases participating in roles other than homeostasis and thrombosis contribute to kidney disease. Moreover, substances activated by thrombin may have an effect on the microvascular circulation of the kidney by modulating the biological function of endothelial cells;21 Chen et al found that they could reduce renal damage caused by ischemia–reperfusion and improve renal function by inhibiting the activity of thrombin through nanoparticles specific to thrombin.22 In this study, we found that HIR could alleviate renal injury markers such as kidney weight/body weight, serum creatinine, blood urea nitrogen, and protein in the urine, while also reducing renal pathological changes in DN rats. Therefore, this suggested that HIR could protect the renal function of DN rats. However, it is important to note that the effects of HIR appear to be only partially protective, as proteinuria is still high in diabetic, HIR-treated rats. This may be related to the dose of HIR, but this is unknown as we only used one dose of HIR. Therefore, in future studies, the dose-dependent effects of HIR should be studied.
On the other hand, inflammation is found to be critical in the development of thrombin-induced diseases. In recent years, the link between inflammation and thrombin has been proven. Here, the coagulation cascade is initiated after vascular injury, which promotes inflammation, and inflammation in turn accelerates the production of thrombin, which eventually leads to the occurrence of disease.23–25 Simultaneously, DN is considered as a chronic inflammatory disease and is one of the most common microvascular complications of diabetes.3 In the present study, our results showed that HIR could markedly reduce macrophage infiltration and pro-inflammatory cytokine expression in renal tissues of DN rats. Macrophage infiltration is an important pathological feature in the progression of DN whereby they mediate the renal inflammatory response in DN.26 Renal biopsy of DM patients revealed that macrophage infiltration was present in DN patients and the aggregation of macrophages was positively correlated with serum creatinine, proteinuria levels, interstitial fibrosis, and glomerulosclerosis in DN patients.27 In addition, the pro-inflammatory cytokine release in the kidney tissue of DN rats arises from two sources.28,29 One is the abnormal glucose and lipid metabolism, hemodynamic disorders, etc. in DN patients that can damage the kidney cells which release a variety of chemokines and pro-inflammatory factors. The other is secreted by inflammatory cells recruited by these chemokines and pro-inflammatory factors. Therefore, the inhibition of inflammation by HIR might help to protect against kidney damage in rats with DN.
Podocytes are highly differentiated cells and their special anatomical structures are the material basis of the glomerular filtration membrane, which can effectively prevent the loss of protein and macromolecular substances in the blood and maintain normal glomerular filtration function.30,31 Therefore, once the structural and functional integrity of podocytes is disrupted, proteinuria eventually occurs.30,31 In this study, we found that HIR could significantly reduce podocyte apoptosis in DN rats. Previous studies have found that high glucose can directly induce the apoptosis of podocytes.32 In order to study the effects of HIR treatment on podocyte apoptosis, we established a high-glucose co-culture system of macrophages and podocytes. At first, we found that HG increased iNOS expression and decreased CD206 expression in macrophages. Macrophages can be polarized into M1 macrophages and M2 macrophages. M1 macrophages can promote inflammation by secreting pro-inflammatory factors (IL-1β, IL-6, IL-12, TNF-α), chemokines (MCP-1), and iNOS, while M2 macrophages exert anti-inflammatory effects by producing large amounts of anti-inflammatory IL-10 and inhibiting the secretion of pro-inflammatory factors IL-12, IL-1, and TNF-α.33,34 Our results showed that HG induced pro-inflammatory macrophage activation. More importantly, the number of macrophages was directly related to podocyte apoptosis in an HG environment and HIR could alleviate this. This suggested that HIR protected podocytes by suppressing inflammation, which may be related to the activation of M1 macrophages.
Emerging evidence from experimental and clinical studies indicated that the p38-MAPK and NF-kB pathway are involved in the development and progression of DN by regulating the secretion of inflammatory cytokines, namely, IL-1β, IL-6, and TNF-α.4,35 In our study, HIR was also found to suppress production of these pro-inflammatory cytokines in podocytes in vitro. Further investigation indicated that inhibition of p38-MAPK by SB203580 or NF-kB by BAY117085 could also cause pro-inflammatory cytokine production in podocytes in vitro. The inhibition of p38 or NF-kB alone by their inhibitors was not as effective as HIR at inhibiting cytokine production, but inhibition by both inhibitors simultaneously was equivalent to the inhibition observed with HIR. This suggested that HIR may inhibit podocyte inflammation via p38-MAPK and NF-kB signaling pathways. Notably, we confirmed this hypothesis and also found that HIR inhibited p-p38/p38 and p-p65/p65 expression in kidney tissues of DN rats.
Taken together, our results suggested that HIR could protect against podocyte apoptosis and inflammation induced by HG in a p38 MAPK/NF-κB-dependent manner, ultimately resulting in protection of kidney function in DN rats.
Disclosure
Jiarui Han and Xinxin Pang are the co-first authors for this study. The authors report no conflicts of interest in this work.
References
1. Serafinceanu C, Neculaescu C, Cimponeriu D, Timar R, Covic AC. Impact of gender and dialysis modality on early mortality risk in diabetic ESRD patients: data from a large single center cohort. Int Urol Nephrol. 2014;46(3):607. doi:10.1007/s11255-013-0589-6
2. Stack AG, Murthy BVR, Molony DA. Survival differences between peritoneal dialysis and hemodialysis among\”large\” ESRD patients in the United States. Kidney Int. 2004;65(6):2398–2408. doi:10.1111/j.1523-1755.2004.00654.x
3. Wada J, Makino H. Inflammation and the pathogenesis of diabetic nephropathy. Clin Sci. 2013;124(3):139–152. doi:10.1042/CS20120198
4. Wagner SA, Satpathy S, Beli P, Choudhary C. SPATA2 links CYLD to the TNF-alpha receptor signaling complex and modulates the receptor signaling outcomes. EMBO J. 2016;35(17):1868–1884. doi:10.15252/embj.201694300
5. Zhang C, Feng Y, Qu S, et al. Resveratrol attenuates doxorubicin-induced cardiomyocyte apoptosis in mice through SIRT1-mediated deacetylation of p53. Cardiovasc Res. 2011;90(3):538–545. doi:10.1093/cvr/cvr022
6. Stone SR, Hofsteenge J. Kinetics of the inhibition of thrombin by hirudin. Biochemistry. 1986;25(16):4622–4628. doi:10.1021/bi00364a025
7. Müller C, Haase M, Lemke S, Hildebrandt J-P. Hirudins and hirudin-like factors inHirudinidae: implications for function and phylogenetic relationships. Parasitol Res. 2017;116(1):313–325. doi:10.1007/s00436-016-5294-9
8. Liu P, Pan X, Yin G. Natural hirudin increases rat flap viability by anti-inflammation via PARs/p38/NF-κB pathway. BioMed Res Int. 2015;2015(12):1–7.
9. Zhao L. Hirudin inhibits cell growth via ERK/MAPK signaling in human glioma. Int J Clin Exp Med. 2016;8(11):20983–20987.
10. Patel SH, Sabbaghi A, Carroll CC. Streptozotocin-induced diabetes alters transcription of multiple genes necessary for extracellular matrix remodeling in rat patellar tendon. Connect Tissue Res. 2018;59(5):447–457. doi:10.1080/03008207.2018.1470168
11. Ying-Xin G, Guo-Qian Y, Jia-Quan L, Han X. Effects of natural and recombinant hirudin on superoxide dismutase, malondialdehyde and endothelin levels in a random pattern skin flap model. J Hand Surg Eur Vol. 2012;37(1):42–49. doi:10.1177/1753193411414628
12. Wang X, Gao L, Lin H, Song J, Li L. Mangiferin prevents diabetic nephropathy progression and protects podocyte function via autophagy in diabetic rat glomeruli. Eur J Pharmacol. 2018;824:170–178.
13. Pan Y, Zhu G, Wang Y, et al. Attenuation of high-glucose-induced inflammatory response by a novel curcumin derivative B06 contributes to its protection from diabetic pathogenic changes in rat kidney and heart. J Nutr Biochem. 2013;24;1:146–155.
14. Cao Y, Hao Y, Li H, et al. Role of endoplasmic reticulum stress in apoptosis of differentiated mouse podocytes induced by high glucose. Int J Mol Med. 2014;33(4):809–816. doi:10.3892/ijmm.2014.1642
15. Meng X, Martinez MA, Raymond-Stintz MA, Winter SS, Wilson BS. IKK inhibitor bay 11-7082 induces necroptotic cell death in precursor-B acute lymphoblastic leukaemic blasts. Br J Haematol. 2010;148(3):487–490. doi:10.1111/j.1365-2141.2009.07988.x
16. Zuo HJ, Lin JY, Liu ZY, et al. Activation of the ERK signaling pathway is involved in CD151-induced angiogenic effects on the formation of CD151-integrin complexes. Acta Pharmacol Sin. 2010;31(7):805–812. doi:10.1038/aps.2010.65
17. Jie T, Jingpu Z, Yun L. Mitochondrial sirtuin 4 resolves immune tolerance in monocytes by rebalancing glycolysis and glucose oxidation homeostasis. Front Immunol. 2018;9:419.
18. Wilson HM, Walbaum D, Rees AJ. Macrophages and the kidney. Curr Opin Nephrol Hypertens. 2004;13(3):285–290. doi:10.1097/00041552-200405000-00004
19. Verzola D, Gandolfo MT, Ferrario F, et al. Apoptosis in the kidneys of patients with type II diabetic nephropathy. Kidney Int. 2007;72(10):1262–1272. doi:10.1038/sj.ki.5002531
20. Menini S, Iacobini C, Oddi G, et al. Increased glomerular cell (podocyte) apoptosis in rats with streptozotocin-induced diabetes mellitus: role in the development of diabetic glomerular disease. Diabetologia. 2007;50(12):2591–2599. doi:10.1007/s00125-007-0821-y
21. Madhusudhan T, Kerlin BA, Isermann B. The emerging role of coagulation proteases in kidney disease. Nat Rev Nephrol. 2013;12(2):94.
22. Chen J, Vemuri C, Palekar RU, et al. Antithrombin nanoparticles improve kidney reperfusion and protect kidney function after ischemia-reperfusion injury. Am J Physiol Renal Physiol. 2015;308(7):F765–F773. doi:10.1152/ajprenal.00457.2014
23. Popovi M, Smiljani K, Dobutovi B, Syrovets T, Simmet T, Isenovi ER. Thrombin and vascular inflammation. Mol Cell Biochem. 2012;359(1–2):301–313.
24. Myles T, Nishimura T, Yun TH, et al. Thrombin activatable fibrinolysis inhibitor, a potential regulator of vascular inflammation. J Biol Chem. 2003;278(51):51059–51067. doi:10.1074/jbc.M306977200
25. Kitamoto Y, Arizono K, Fukui H, et al. Urinary thrombin: a novel marker of glomerular inflammation for the diagnosis of crescentic glomerulonephritis (prospective observational study). PLoS One. 2015;10(3):e0118704. doi:10.1371/journal.pone.0118704
26. Tesch GH. Role of macrophages in complications of type 2 diabetes. Clin Exp Pharmacol Physiol. 2010;34(10):1016–1019. doi:10.1111/j.1440-1681.2007.04729.x
27. Yonemoto S, Machiguchi T, Nomura K, Minakata T, Nanno M, Yoshida H. Correlations of tissue macrophages and cytoskeletal protein expression with renal fibrosis in patients with diabetes mellitus. Clin Exp Nephrol. 2006;10(3):186–192. doi:10.1007/s10157-006-0426-7
28. Sakai N, Wada T. Revisiting inflammation in diabetic nephropathy: the role of the Nlrp3 inflammasome in glomerular resident cells. Kidney Int. 2015;87(1):12–14. doi:10.1038/ki.2014.322
29. Wada J, Makino H. Innate immunity in diabetes and diabetic nephropathy. Nat Rev Nephrol. 2015;12(1):13–26. doi:10.1038/nrneph.2015.175
30. Imasawa T, Obre E, Bellance N, et al. High glucose repatterns human podocyte energy metabolism during differentiation and diabetic nephropathy. FASEB J. 2017;31(1):294–307.
31. Tian X, Ishibe S. Targeting the podocyte cytoskeleton: from pathogenesis to therapy in proteinuric kidney disease. Nephrol Dial Transplant. 2016;31(10):1577–1583.
32. Qian X, Tan J, Liu L, Chen S, Lu Y. MicroRNA-134-5p promotes high glucose-induced podocyte apoptosis by targeting bcl-2. Am J Transl Res. 2018;10(3):989–997.
33. Wynn TA, Barron L. Macrophages: master regulators of inflammation and fibrosis.
34. Tang PM-K, Nikolic-Paterson DJ, Lan H-Y. Macrophages: versatile players in renal inflammation and fibrosis. Nat Rev Nephrol. 2019;15(3):144–158. doi:10.1038/s41581-019-0110-2
35. Brunet A, Sweeney LB, Sturgill JF, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science (New York, N Y). 2004;303(5666):2011–2015. doi:10.1126/science.1094637
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.