Back to Journals » Cancer Management and Research » Volume 18

Hippo-YAP/TAZ Signaling in Hematological Malignancies: Molecular Mechanisms, Pathway Crosstalk and Therapeutic Potential
Authors Jiang W, Zhu C, Xu L ![]() , Ma N
, Ma N
Received 21 April 2026
Accepted for publication 26 June 2026
Published 7 July 2026 Volume 2026:18 618762
DOI https://doi.org/10.2147/CMAR.S618762
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Kattesh Katti
Wanfu Jiang1, Changling Zhu2, Li Xu3, Na Ma4
1School of Medicine, Anhui University of Science and Technology, Huainan, People’s Republic of China; 2Graduate School, Bengbu Medical University, Bengbu, People’s Republic of China; 3Department of Hematology, Anhui No. 2 Provincial People’s Hospital (Affiliated Hospital of Anhui Second Medical College), Hefei, People’s Republic of China; 4Department of Radiology, Anhui No. 2 Provincial People’s Hospital (Affiliated Hospital of Anhui Second Medical college), Hefei, People’s Republic of China
Correspondence: Li Xu, Department of hematology, Anhui No. 2 Provincial People’s Hospital Affiliated Hospital of Anhui Second Medical College, Hefei, People’s Republic of China, Email [email protected]; Na Ma, Department of Radiology, Anhui No. 2 Provincial People’s Hospital (Affiliated Hospital of Anhui Second Medical college, Hefei, People’s Republic of China, Email [email protected]
Abstract: The Hippo signaling pathway represents an evolutionarily conserved regulatory network. It plays a central role in controlling cell proliferation, apoptosis, differentiation, and tissue homeostasis. Increasing evidence indicates that dysregulation of this pathway promotes the development and progression of hematological malignancies. This includes leukemia, lymphoma, and multiple myeloma. Notably, aberrant activation of downstream effectors—Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ)—plays a central role in this process. Abnormal YAP/TAZ signaling promotes malignant cell survival, proliferation, therapeutic resistance, and disease aggressiveness through extensive crosstalk with multiple oncogenic pathways, such as Phosphatidylinositol 3-kinase (PI3K)/ Protein kinase B (AKT), Transforming growth factor-beta (TGF-β), Wnt/β-catenin, and metabolic signaling networks. Notably, the biological functions of YAP/TAZ appear to be highly context-dependent, with both oncogenic and tumor-suppressive roles reported in different hematopoietic lineages and tumor microenvironments. In this review, we summarize the molecular architecture and regulatory mechanisms of the Hippo pathway, discuss its dysregulation and functional significance in major hematological malignancies, and highlight recent advances in Hippo-targeted therapeutic strategies, including YAP/TEA domain transcription factor (TEAD) inhibitors, upstream pathway modulators, and combination treatment approaches. We further outline current challenges and future opportunities for translating Hippo-based precision therapies into clinical practice. Despite promising preclinical findings, hematological malignancy-specific clinical evidence remains limited. Future studies are required to validate Hippo-targeted therapeutic strategies and establish clinically actionable biomarkers. A deeper understanding of Hippo signaling may provide novel insights into disease biology and accelerate the development of precision medicine approaches for hematological malignancies.
Keywords: hematological malignancies, hippo signaling pathway, leukemia, lymphoma, myeloma, precision medicine and therapeutic targets, tumor microenvironment, YAP and TAZ
Introduction
Hematological malignancies, including leukemia, lymphoma, and multiple myeloma (MM), comprise a heterogeneous group of clonal disorders arising from hematopoietic cells and remain a major cause of cancer-related morbidity and mortality worldwide.1,2 Despite substantial progress in targeted therapy, immunotherapy, and supportive care, many patients still experience disease relapse, therapeutic resistance, and poor long-term outcomes, particularly in biologically aggressive or refractory subtypes.3,4 Context-dependent functional heterogeneity of YAP/TAZ in hematological malignancies—where these effectors can act as either oncogenes or tumor suppressors depending on the specific lineage and microenvironment—has emerged as a critical consideration for therapeutic development.5 These persistent clinical challenges underscore the urgent need to identify novel molecular mechanisms that drive malignant progression and treatment failure.6,7
The Hippo pathway has garnered increasing attention due to its central role in regulating cell proliferation, apoptosis, stemness, and tissue homeostasis.8,9 Dysregulation of this pathway, especially aberrant activation of its downstream transcriptional coactivators YAP and TAZ, has been increasingly implicated in hematological malignancy pathogenesis.10,11 Growing evidence suggests that Hippo pathway dysfunction promotes malignant cell survival and proliferation.12,13 Additionally, it contributes to chemoresistance, microenvironmental adaptation, and disease progression.14,15 Therefore, a comprehensive understanding of Hippo signaling may provide important mechanistic insights and reveal new opportunities for precision-targeted therapeutic intervention in hematological cancers.16,17 Although numerous reviews have comprehensively summarized the role of Hippo signaling in solid tumors and general cancer biology, a focused synthesis of its involvement in hematological malignancies remains limited. In particular, recent discoveries regarding the context-dependent functions of YAP/TAZ, extensive signaling crosstalk, and emerging Hippo-targeted therapeutic strategies have not been systematically integrated within the framework of hematologic cancers.
In this review, we specifically focus on the molecular mechanisms and biological functions of Hippo signaling in hematological malignancies. We summarize current evidence regarding pathway dysregulation in leukemia, lymphoma, and multiple myeloma, discuss interactions with major oncogenic signaling networks, evaluate emerging therapeutic strategies and clinical advances, and highlight future opportunities for precision medicine in hematologic cancers.
Molecular Mechanisms of the Hippo Signaling Pathway
Core Components
The Hippo signaling pathway exerts its biological functions via an evolutionarily conserved serine/threonine kinase cascade and downstream transcriptional regulatory modules, which collectively regulate cell proliferation, apoptosis, differentiation, and tissue homeostasis in hematopoietic lineages. Its core kinase module consists of Mammalian sterile 20-like kinases 1 and 2 (MST1/2) and Large tumor suppressor kinases 1 and 2 (LATS1/2): as upstream initiators, MST1/2 phosphorylate and activate LATS1/2 upon recruitment of scaffold proteins; activated LATS1/2 then induce inhibitory phosphorylation of YAP/TAZ—the key downstream effectors of the pathway.18 Scaffold proteins are essential for the assembly and activation of the Hippo core kinase complex in hematopoietic cells. Salvador family WW domain-containing protein 1 (SAV1), a classic scaffold protein, bridges MST1/2 and LATS1/2 to enhance their physical interaction and kinase activation.19 In contrast, CORO7—a newly identified scaffold protein—binds to LATS1 and MST2 to promote formation of functional core kinase complexes.18 This enhances the efficiency and specificity of Hippo signaling in hematopoietic homeostasis. Loss of these scaffold proteins impairs Hippo kinase complex assembly in hematopoietic cells. Consequently, YAP/TAZ phosphorylation is reduced, leading to their aberrant nuclear accumulation. This promotes oncogenic transcriptional programs and drives malignant transformation of hematopoietic progenitors.
In the inactive state of the Hippo pathway, unphosphorylated YAP/TAZ translocate to the nucleus and bind to TEAD1-4 transcription factors, thereby driving the expression of oncogenic target genes associated with hematopoietic cell proliferation (eg., CCND1, MYC) and anti-apoptosis (eg., BCL2).20 Beyond canonical cell growth regulation, the Hippo pathway plays a unique role in hematopoietic innate immunity and inflammatory responses: innate immune and proinflammatory signals in the hematopoietic microenvironment activate the Hippo pathway via the Tak1-STRIPAK-Tao axis, which modulates cell fate decisions in hematopoietic lineages and maintains hematopoietic tissue homeostasis under inflammatory stress.13 Collectively, the core Hippo components (MST1/2, LATS1/2, YAP/TAZ) and scaffold proteins (SAV1, CORO7) form a complex regulatory network that integrates multiple extracellular signals in the hematopoietic microenvironment to govern physiological and pathological processes of hematopoietic cells.
Regulation of YAP/TAZ
YAP/TAZ are the key downstream effectors of the Hippo pathway in hematological malignancies, and their subcellular localization and transcriptional activity are tightly regulated by phosphorylation, degradation, and hematopoietic microenvironment-specific mechanical/metabolic signals. The canonical regulatory mechanism is mediated by the MST1/2-LATS1/2 kinase cascade: LATS1/2 directly phosphorylate YAP at Ser127 and TAZ at Ser89 in hematopoietic cells, which promotes 14-3-3 protein binding to YAP/TAZ, sequestering them in the cytoplasm and abrogating their nuclear translocation.21 Additionally, LATS1/2-induced phosphorylation of YAP at Ser381 triggers subsequent CK1-mediated phosphorylation, leading to ubiquitination and proteasomal degradation of YAP in malignant hematopoietic cells.22
Independent of the MST1/2-LATS1/2 cascade, YAP/TAZ activity in hematological malignancies is modulated by mechanical and metabolic signals unique to the hematopoietic microenvironment. Mechanical cues from bone marrow stromal matrix stiffness and altered cytoskeletal architecture modulate YAP/TAZ nuclear translocation. This occurs via Rho GTPases and actin cytoskeleton remodeling. Specifically, increased bone marrow ECM stiffness drives F-actin polymerization, which inhibits LATS1/2 activity. Consequently, YAP/TAZ undergo aberrant activation in leukemia and multiple myeloma cells.23 Metabolic signals, a hallmark of malignant hematopoietic cell reprogramming, also play a critical role in YAP/TAZ regulation: AMP-activated protein kinase (AMPK), a cellular energy sensor, directly phosphorylates YAP/TAZ to inhibit their transcriptional activity in glycolysis-dependent hematological malignant cells.24 Hexokinase 2 (HK2), a key glycolytic enzyme overexpressed in multiple myeloma and acute myeloid leukemia (AML) cells, links aerobic glycolysis to YAP/TAZ activation in malignant hematopoietic cells, highlighting the metabolic-Hippo crosstalk that sustains the survival of hematological tumor cells.
YAP/TAZ predominantly exert oncogenic functions in many hematological malignancies. Although this is generally the case, notable exceptions exist. TAZ exhibits context-dependent functional dichotomy in hematological malignancies, acting as a tumor suppressor in multiple myeloma by downregulating MYC expression, in contrast to its oncogenic role in leukemia and most lymphomas.25,26 These alterations are closely associated with tumorigenesis, disease progression, and chemoresistance.18 Notably, YAP/TAZ display hematopoietic context-dependent functional heterogeneity: their biological roles are strictly influenced by hematopoietic cell lineage, bone marrow/lymph node tumor microenvironment, and co-expression of other oncogenic factors. TAZ exemplifies this functional dichotomy: it acts as a tumor suppressor in multiple myeloma by downregulating MYC expression—a stark contrast to its oncogenic role in leukemia and most lymphomas.26 Elucidating the precise lineage- and microenvironment-specific regulatory mechanisms of YAP/TAZ in hematological malignancies is therefore critical for the development of subtype-specific targeted therapeutic strategies (Figure 1).
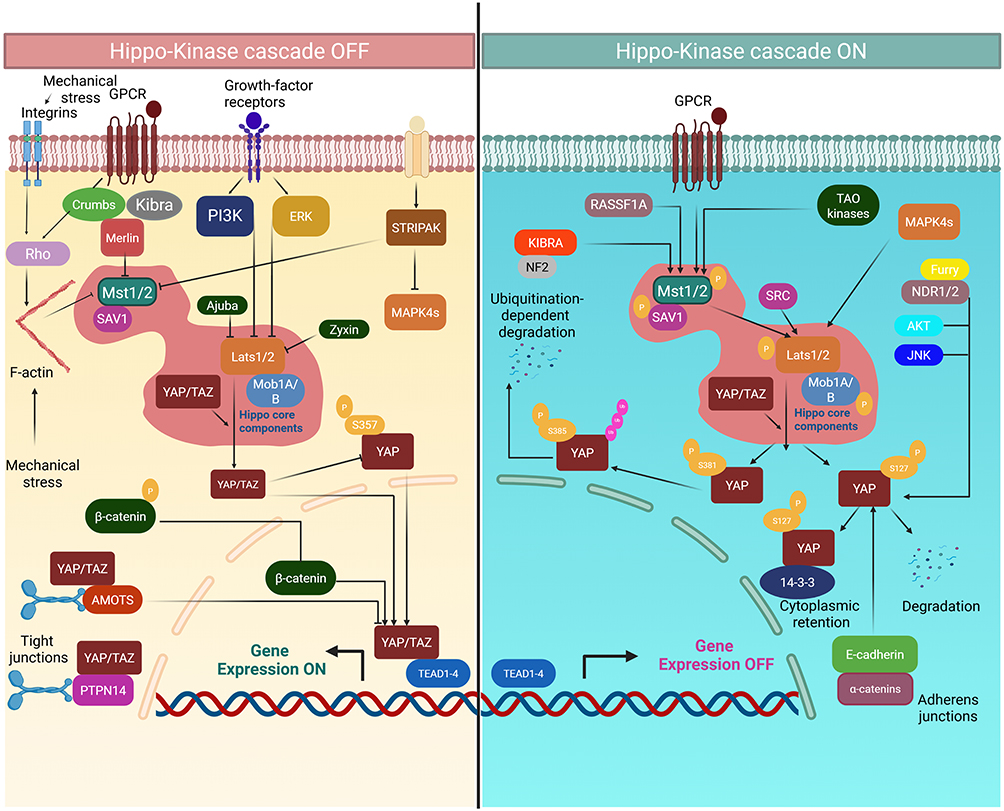 |
Figure 1 Nuclear translocation of YAP/TAZ and downstream transcriptional activation following Hippo pathway suppression.Under physiological conditions, the Hippo core kinase cascade (MST1/2–SAV1–LATS1/2–MOB1A/B) remains active and induces inhibitory phosphorylation of YAP and TAZ, leading to their cytoplasmic retention through 14-3-3 binding. Phosphorylated YAP is further subjected to CK1-mediated phosphorylation and ubiquitin-dependent proteasomal degradation. Upon Hippo pathway inactivation, triggered by mechanical stress, growth factor stimulation, or GPCR signaling, unphosphorylated YAP/TAZ translocate into the nucleus and interact with TEAD1–4 transcription factors. This process promotes the transcription of genes involved in cell-cycle progression, proliferation, and anti-apoptotic signaling, including CCND1, MYC, and BCL2, thereby driving malignant hematopoietic cell survival. Abbreviations: CK1, casein kinase 1; GPCR, G protein-coupled receptor; TEAD, TEA domain transcription factor. |
Hippo Pathway Cross-Talk with Other Signaling Pathways
The Hippo pathway does not act as an independent regulatory network but engages in extensive crosstalk with other key oncogenic signaling cascades, forming a complex regulatory network that regulates cell fate decisions and tumorigenesis. The crosstalk between the Hippo pathway and TGF-β, Wnt/β-catenin, PI3K/Akt, and Notch pathways has been well characterized, and these interactions play critical roles in hematological malignancies.
TGF-β/Mothers Against Decapentaplegic Homolog (Smad) Signaling
YAP/TAZ can bind to Smad proteins and enhance their transcriptional activity, thereby amplifying TGF-β/Smad signaling.13 TGF-β can also upregulate TAZ expression, forming a positive feedback loop that promotes fibrosis and epithelial-mesenchymal transition (EMT).13 In hematological malignancies, this crosstalk may contribute to the malignant transformation of hematopoietic cells and the remodeling of the tumor microenvironment.
Wnt/β-Catenin Signaling
TAZ acts as a transcriptional coactivator of β-catenin, enhancing the expression of Wnt target genes.27 The hierarchical crosstalk between the Hippo, TGF-β, and Wnt pathways regulates the switch between proliferative and invasive cellular phenotypes in tumors.27 Dysregulation of any of these pathways can affect the function of the others, accelerating tumor progression.
PI3K/Akt Signaling
This is the most well-characterized crosstalk partner of the Hippo pathway in cancer biology. PI3K/Akt signaling can directly inhibit LATS1/2 activity, leading to YAP/TAZ activation.28 Vascular endothelial growth factor receptor (VEGFR) can regulate the Hippo pathway via PI3K/Mitogen-activated protein kinase (MAPK) activation, further linking angiogenesis to Hippo signaling.28 This crosstalk is particularly important in hematological malignancies, as PI3K/Akt activation is a common oncogenic event that contributes to YAP/TAZ-mediated chemoresistance.
Notch Signaling
Notch1 signaling can upregulate YAP/TAZ expression in endothelial cells, and knockdown of Notch1 leads to reduced YAP/TAZ levels.13 In hematological malignancies, the Notch pathway is frequently dysregulated in T-cell leukemia and lymphoma, and its crosstalk with the Hippo pathway may play a critical role in malignant cell survival and proliferation.
In addition, metabolic signals and non-coding RNAs act as key mediators of the crosstalk between the Hippo pathway and other signaling cascades.13 This complex crosstalk network underscores the importance of understanding Hippo pathway regulation in hematological malignancies, as single-target therapies may be limited by compensatory activation of other oncogenic pathways (Figure 2 and Table 1). Importantly, such multilayered crosstalk creates a highly adaptive signaling network that may underlie therapeutic resistance and limit the efficacy of single-pathway-targeted strategies.
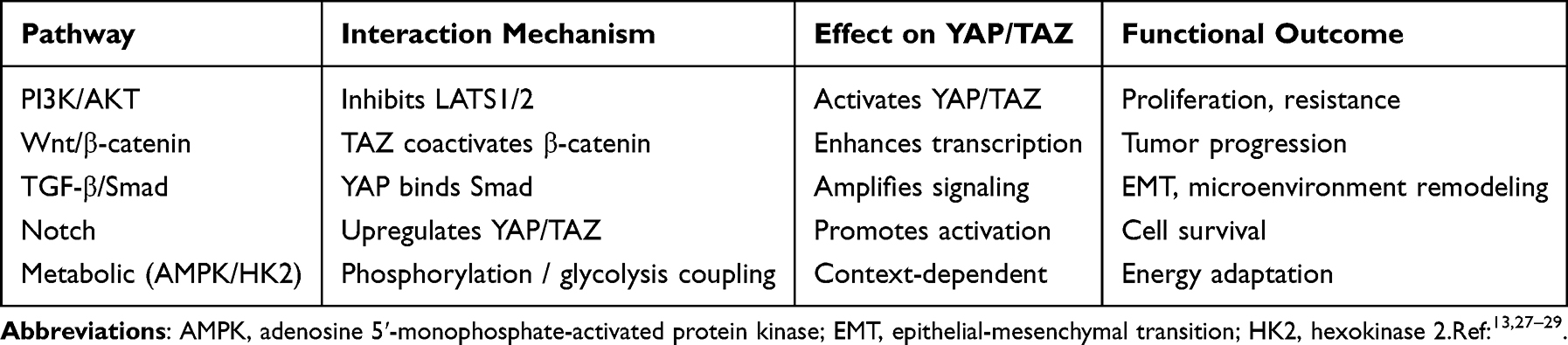 |
Table 1 Crosstalk Between Hippo Pathway and Major Oncogenic Signaling Pathways |
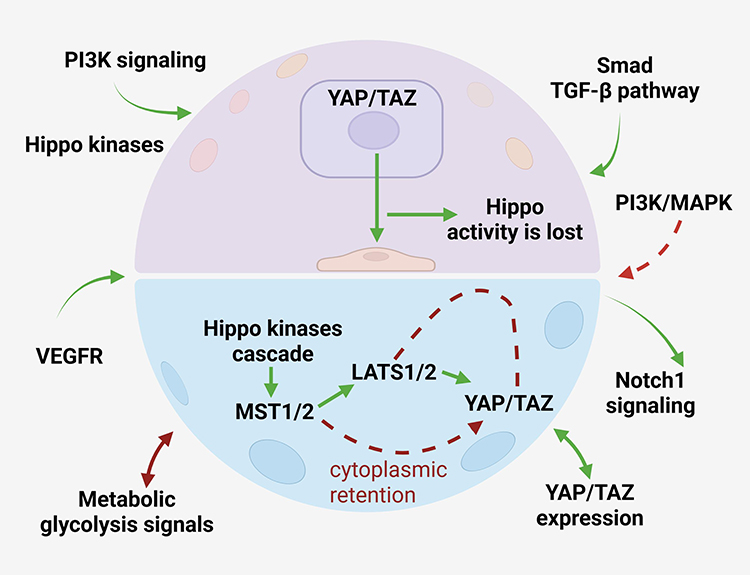 |
Figure 2 Crosstalk between Hippo signaling and major oncogenic pathways in hematological malignancies Dysregulated YAP/TAZ signaling extensively interacts with multiple oncogenic pathways. YAP/TAZ enhance TGF-β/Smad signaling through direct association with Smad proteins, strengthen Wnt/β-catenin signaling by functioning as transcriptional coactivators, and are positively regulated by PI3K/AKT signaling through LATS1/2 inhibition. In addition, Notch signaling can increase YAP/TAZ expression and amplify their oncogenic activity. Metabolic reprogramming, particularly glycolysis-related pathways, further contributes to aberrant Hippo signaling activation. These interconnected signaling networks collectively promote malignant proliferation, invasive phenotypic transition, microenvironmental adaptation, and disease progression in hematological cancers. Symbols: solid green arrows indicate activation, signaling transmission, or downstream biological effect; solid green double-headed arrows indicate bidirectional crosstalk or reciprocal regulation between signaling pathways; solid brown double-headed arrows indicate bidirectional metabolic signaling or feedback regulation; red dashed arrows indicate aberrant signaling associated with tumor progression; red dashed blunt-ended arrows indicate indirect inhibitory regulation. Abbreviations: AKT, v-akt murine thymoma viral oncogene homolog; EMT, epithelial–mesenchymal transition; PI3K, phosphatidylinositol 3-kinase; TGF-β, transforming growth factor-β. |
Hippo Pathway in Hematological Malignancies
Leukemias
The Hippo pathway plays a pivotal role in the regulation of normal hematopoiesis, and its dysregulation is closely associated with the initiation and progression of various leukemias. Genetic and epigenetic alterations in core Hippo pathway components are commonly detected in leukemic cells. Specifically, MST1/2 and LATS1/2 are frequently deleted or transcriptionally downregulated. In acute myeloid leukemia, RHOBTB2 enhances cell proliferation by stabilizing KLHL13, which inhibits Hippo pathway activity and activates YAP/TAZ transcriptional coactivators, thereby revealing a previously unrecognized RHOBTB2-KLHL13-Hippo axis in AML pathogenesis.30 This leads to aberrant nuclear accumulation of YAP/TAZ and subsequent activation of oncogenic target genes.18 This dysregulation drives the malignant transformation of hematopoietic stem/progenitor cells by promoting cell proliferation, inhibiting apoptosis, and impairing cellular differentiation.
The crosstalk between the Hippo pathway and other oncogenic signaling cascades further exacerbates leukemogenesis. For example, the PI3K/Akt pathway, which is commonly activated in acute myeloid leukemia (AML) and acute lymphoblastic leukemia (ALL), can inhibit LATS1/2 activity and enhance YAP/TAZ activation.28 Innate immune and proinflammatory signals in the leukemic microenvironment can also modulate Hippo pathway activity via the Tak1-STRIPAK-Tao axis, influencing cell fate decisions in hematopoietic lineages and promoting leukemic cell survival.13,31 Functional studies have confirmed the oncogenic role of YAP in leukemia: knockdown of YAP in Jurkat cells (a human T-cell leukemia cell line) significantly inhibits cell proliferation and induces apoptosis, and YAP inhibition enhances the therapeutic responsiveness of leukemic cells to conventional chemotherapeutic agents.31,32
Notably, the Hippo pathway is also involved in the development of therapeutic resistance in leukemia: YAP1 activity confers survival benefits to leukemic cells under chemotherapy stress, and targeting YAP may be a promising strategy to overcome refractoriness to standard treatments.32 In chronic myeloid leukemia, Hippo-YAP signaling is involved in imatinib resistance in Chronic myeloid leukemia (CML), and ectopic overexpression of STK4 enhances imatinib-induced apoptosis of CML cells.33,34 Furthermore, LATS2 expression is decreased in highly drug-resistant leukemic cells and is associated with unfavorable prognosis.25 Preclinical studies have shown that small-molecule agents targeting the Hippo pathway (eg., YAP/TAZ inhibitors) can effectively induce apoptosis in leukemic cells and inhibit tumor growth in xenograft models.18 These findings confirm the Hippo pathway as a potential therapeutic target in leukemia, especially for refractory subtypes. Collectively, these findings support the Hippo pathway as a critical driver of leukemogenesis and a promising therapeutic target, particularly in treatment-resistant disease.
Lymphomas
Lymphomas are a heterogeneous group of B-cell or T-cell malignancies, and the Hippo pathway has emerged as a key regulator of lymphoma progression and therapeutic resistance. “Dysregulation of the Hippo pathway, characterized by YAP/TAZ overexpression or nuclear accumulation, has been documented in major lymphoma subtypes, including Diffuse large B-cell lymphoma (DLBCL) and natural killer/T-cell lymphoma (NK/TCL), and correlates with tumor aggressiveness and poor clinical outcomes.35
In DLBCL—the most common non-Hodgkin lymphoma subtype—Hippo pathway dysregulation underlies rituximab resistance. Rituximab is a key component of standard first-line immunochemotherapy regimens for DLBCL. Notably, Semaphorin-3F, a secreted protein, can downregulate YAP/TAZ expression and restore rituximab sensitivity in DLBCL cells. This suggests that modulating the Hippo pathway may enhance the efficacy of anti-CD20 antibody therapy.35 Genetic alterations in Hippo pathway components (eg., LATS) are also common in B-cell lymphomas, and concurrent alterations of TP53 and Hippo pathway components lead to markedly poorer prognosis, suggesting that concurrent inactivation of the Hippo pathway and p53 signaling accelerates lymphoma progression.36 In NK/TCL, a highly aggressive lymphoma subtype, activation of MST1/2 significantly inhibits cell proliferation and induces apoptosis by suppressing YAP activity, confirming the therapeutic potential of Hippo pathway activation in aggressive lymphomas.37
Metabolic and oxidative stress signals also modulate Hippo pathway activity in lymphomas: increased reactive oxygen species (ROS) levels can activate MST1, strengthen Hippo signaling, and drive cell death in lymphoma cells.37 ROS function as both signaling molecules and cell death inducers. This dual role reveals the intricate link between cellular metabolism and the Hippo pathway in lymphoma biology. In addition, non-coding RNAs have emerged as important regulators of Hippo signaling in lymphoma. For example, hsa_circ_0000877 promotes DLBCL progression through the miR-370-3p/MAP4K4/Hippo signaling axis, highlighting the potential of non-coding RNA (ncRNA)-mediated Hippo regulation as a therapeutic target in lymphoma.38 Collectively, these findings confirm the critical role of the Hippo pathway in lymphoma pathogenesis and highlight its potential as both a prognostic biomarker and a therapeutic target. These observations highlight the Hippo pathway as both a prognostic biomarker and a potential therapeutic vulnerability in lymphoma.
Myelomas
Multiple myeloma (MM) is a plasma cell malignancy characterized by clonal expansion of abnormal plasma cells in the bone marrow. Hippo pathway dysregulation represents a pivotal mechanism underlying MM development, progression, and chemoresistance. Excessive YAP expression is a common event in MM cells and is associated with enhanced cell proliferation, survival, and resistance to conventional chemotherapeutic agents.26 Transcriptomic analysis of ~1000 MM patients from the CoMMpass study demonstrated that TAZ expression significantly decreases during disease progression from stage 1 to stage 3, showing an inverse correlation with disease severity. High TAZ expression is strongly associated with better prognosis in MM patients, further supporting its tumor-suppressive function in this malignancy.25 The functional role of YAP/TAZ in MM is complex and context-dependent: YAP functions as an oncogene by promoting MM cell proliferation and survival, whereas TAZ exhibits tumor-suppressive activity by downregulating MYC expression, revealing the functional heterogeneity of Hippo effectors in MM.26
The crosstalk between the Hippo pathway and metabolic pathways is a key regulatory mechanism in MM. PLA2G16, a lipid metabolism-related gene, modulates Hippo/YAP signaling in MM cells: inhibition of PLA2G16 leads to cell cycle arrest and apoptosis, and impairs glucose metabolism reprogramming in MM cells.39 This finding reveals the intricate link between lipid metabolism and the Hippo pathway, providing a basis for dual-target therapy targeting metabolic pathways and the Hippo pathway in MM. Non-coding RNAs also critically regulate the Hippo pathway in MM. For instance, long non-coding RNA (lncRNA) MALAT1 functions as a competing endogenous RNA (ceRNA) that sponges miR-181a-5p, thereby modulating Hippo-YAP signaling and promoting multiple myeloma cell proliferation and adhesion.40 This lncRNA-microRNA (miRNA)-Hippo axis forms a complex regulatory network that modulates MM cell behavior, and targeting this axis has shown promising preclinical efficacy.
YAP/TAZ are also involved in the regulation of oxidative stress responses in MM: TAZ can upregulate miR-224 to inhibit oxidative stress responses, thereby promoting MM cell survival.41 This finding further expands the functional scope of the Hippo pathway in MM and suggests that targeting the Hippo-oxidative stress axis may be a novel therapeutic strategy. Preclinical studies have confirmed that reactivation of the Hippo pathway can effectively inhibit MM cell growth and induce apoptosis, and combining Hippo pathway modulators with conventional chemotherapeutic agents or proteasome inhibitors may overcome chemoresistance in MM26(Table 2). Overall, Hippo pathway dysregulation represents a key mechanism underlying MM progression and drug resistance, supporting its relevance as a therapeutic target.
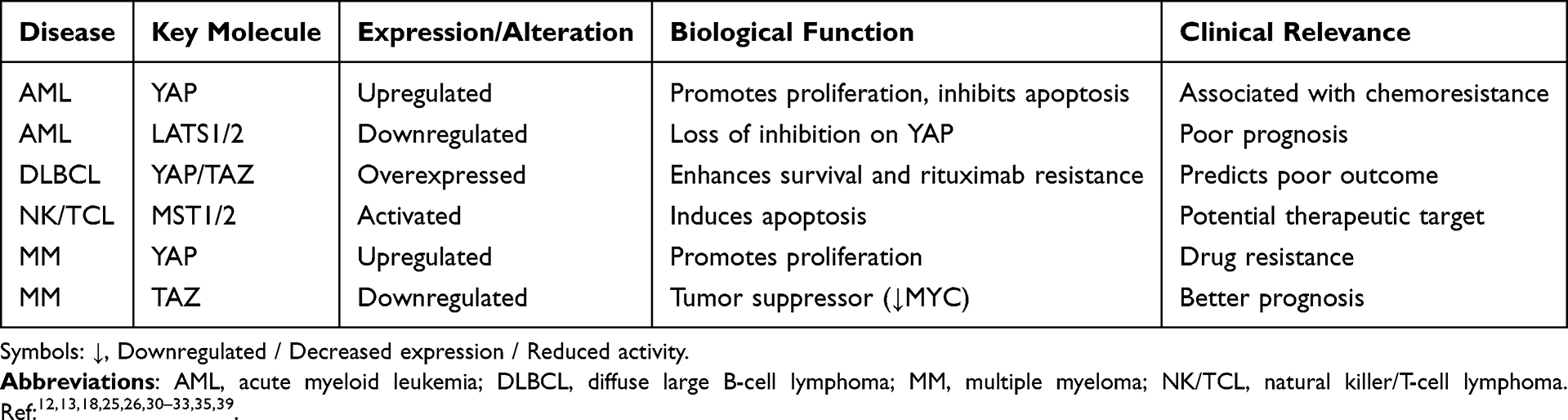 |
Table 2 Functional Roles of Hippo Pathway Components in Hematological Malignancies |
Context-Dependent Functions of YAP and TAZ
Accumulating evidence suggests that the biological functions of YAP and TAZ in hematological malignancies are highly context-dependent and cannot be simply categorized as uniformly oncogenic.42,43 While YAP/TAZ activation generally promotes proliferation, survival, and therapeutic resistance in many hematologic cancers, emerging studies indicate that these transcriptional coactivators may exert tumor-suppressive functions in specific cellular contexts.29,42,43
In leukemia and lymphoma, YAP/TAZ are predominantly considered oncogenic drivers.44–46 Elevated YAP expression has been associated with enhanced proliferation, inhibition of apoptosis, and poor clinical outcomes in acute myeloid leukemia (AML), diffuse large B-cell lymphoma (DLBCL), and T-cell acute lymphoblastic leukemia (T-ALL).45,47 Mechanistically, YAP/TAZ cooperate with TEAD transcription factors to induce the expression of genes involved in cell-cycle progression, metabolic adaptation, and survival signaling, thereby facilitating malignant transformation and disease progression.29,44,48
In contrast, studies in multiple myeloma (MM) have revealed a more complex regulatory landscape.26,46 Grieve et al demonstrated that TAZ functions as a tumor suppressor in MM by repressing MYC expression and limiting malignant plasma-cell growth. Loss of TAZ expression was associated with enhanced tumor progression, highlighting a biological role distinct from that observed in many other malignancies.26 Similarly, previous investigations have suggested that activation of specific Hippo pathway components may promote apoptosis rather than tumor growth in selected hematologic contexts.26,44,49
These observations emphasize that the functional consequences of Hippo pathway dysregulation depend on cellular lineage, genetic background, and microenvironmental factors.44,46,50 Therefore, therapeutic strategies targeting YAP/TAZ should be carefully tailored according to disease subtype.29,43 While YAP/TAZ inhibition may represent a promising approach for leukemia and lymphoma, indiscriminate suppression of Hippo signaling could be detrimental in malignancies where TAZ exerts tumor-suppressive functions.26,46,49 A more comprehensive understanding of the context-specific biology of YAP/TAZ will be essential for the development of precision Hippo-targeted therapies.44,50
Molecular Evidence
Accumulating molecular and clinical evidence has confirmed the critical role of the Hippo pathway in the pathogenesis of hematological malignancies, and YAP/TAZ have emerged as key molecular biomarkers and therapeutic targets. Systematic molecular profiling studies have revealed that genetic alterations in Hippo pathway-related genes (eg., MST1/2, LATS1/2, YAP/TAZ) are common across various hematological malignancies, and these alterations are closely associated with disease stage and prognosis.18,45 For example, abnormal YAP expression is a key driver of DLBCL progression, and YAP expression level can serve as a prognostic biomarker for DLBCL patients.45
Functional studies have confirmed the causal relationship between Hippo pathway dysregulation and hematological malignancies: Wang et al found that Hippo pathway alterations are associated with the malignant transformation of hematopoietic cells, and YAP/TAZ overexpression is sufficient to promote the formation of hematological tumors in animal models.18 Zhou et al elucidated the specific mechanism by which IGF-1R regulates DLBCL tumorigenesis via the Hippo-YAP pathway, confirming the Hippo pathway as a downstream effector of oncogenic receptors in lymphoma.45 In myeloproliferative neoplasms (MPNs), the expression of Hippo pathway-related genes is significantly deregulated, and these alterations are associated with the abnormal proliferation of hematopoietic cells.51
Targeted inhibition of YAP/TAZ has shown promising preclinical efficacy in hematological malignancies. Verteporfin, a small-molecule agent that disrupts the YAP-TEAD interaction, can effectively inhibit the transcriptional activity of YAP in various cancer cell lines, including hematological malignant cells.45 Non-coding RNAs also provide potential therapeutic targets: miR-103a-3p can regulate the Hippo/YAP axis to inhibit tumor glycolysis in solid tumors, and this mechanism may be conserved in hematological malignancies.52 These molecular findings not only confirm the Hippo pathway as a key oncogenic driver in hematological malignancies but also provide a wealth of potential molecular targets for the development of novel targeted therapies (Figure 3).
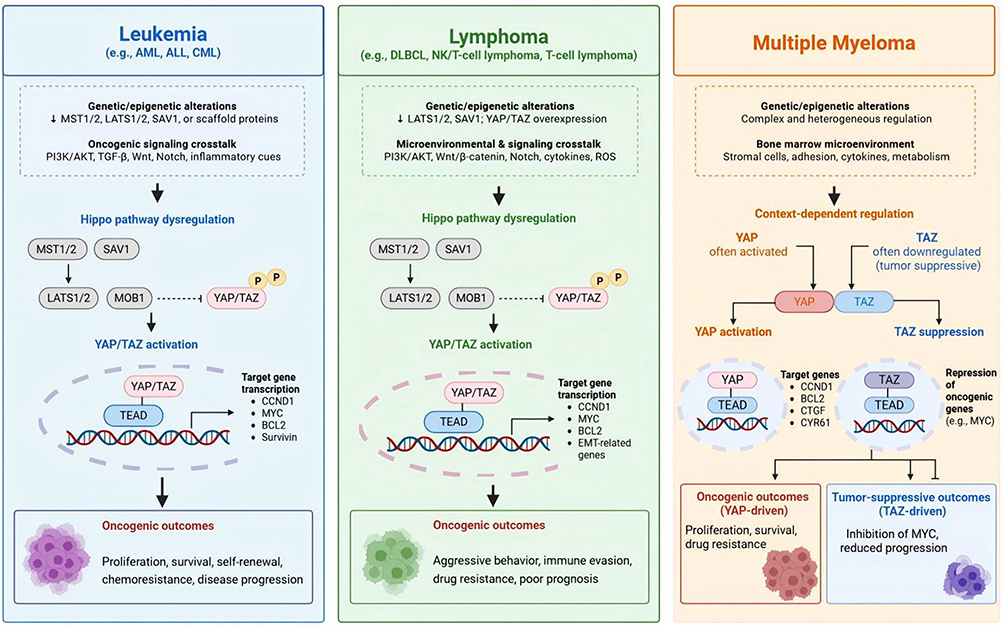 |
Figure 3 Hippo pathway dysregulation and context-dependent YAP/TAZ functions across major hematological malignancies Genetic and epigenetic alterations, together with oncogenic signaling crosstalk and microenvironmental cues, contribute to Hippo pathway dysregulation in leukemia, lymphoma, and multiple myeloma. In leukemia and lymphoma, loss of upstream Hippo kinase activity promotes YAP/TAZ nuclear accumulation and TEAD-dependent transcription, resulting in enhanced proliferation, survival, drug resistance, and disease progression. In multiple myeloma, Hippo signaling exhibits context-dependent regulation, with YAP frequently acting as an oncogenic factor whereas TAZ may exert tumor-suppressive functions through repression of MYC-associated programs.Symbols: ↓, inhibition, loss, or downregulation; →, activation, signaling transmission, or downstream biological effect; ⊣, inhibitory regulation; P, phosphorylation; dashed lines/arrows indicate indirect regulation or multistep signaling events; dashed blunt-ended lines indicate indirect inhibitory regulation rather than direct molecular interactions. Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; CML, chronic myeloid leukemia; DLBCL, diffuse large B-cell lymphoma; EMT, epithelial–mesenchymal transition; MM, multiple myeloma; NK/TCL, natural killer/T-cell lymphoma; ROS, reactive oxygen species; TEAD, TEA domain transcription factor. |
Therapeutic Implications
Targeting the Hippo Pathway in Cancer Therapy
Targeting the Hippo pathway has emerged as a promising therapeutic strategy for cancer, and preclinical studies have demonstrated its therapeutic potential in hematological malignancies. The therapeutic strategies for targeting the Hippo pathway can be divided into two main categories: direct targeting of YAP/TAZ and indirect targeting via upstream regulators or pathway crosstalk.
Direct targeting of YAP/TAZ focuses on inhibiting their transcriptional activity or nuclear translocation. Verteporfin is the most well-characterized YAP inhibitor. Originally developed as a photosensitizer for age-related macular degeneration, it disrupts the YAP-TEAD interaction and inhibits YAP-mediated transcriptional activity.5 Insights into the dual role of YAP/TAZ in hematological malignancies suggest that therapeutic strategies may require disease-specific tailoring. YAP activation rather than inhibition may prove beneficial in certain hematologic cancers where YAP functions as a tumor suppressor.5 Although verteporfin has shown promising preclinical efficacy in hematological malignant cell lines, its clinical application is limited by phototoxicity and poor stability, highlighting the need for the development of more specific and potent YAP/TAZ inhibitors.53 First-generation TEAD inhibitors have been developed. These agents target the YAP-TEAD binding interface or the TEAD palmitoylation site. They have shown efficacy in preclinical models of tumors with high YAP/TAZ activity. Their efficacy in hematological malignancies is currently under investigation.17,54
Indirect targeting of the Hippo pathway via upstream regulators provides an alternative strategy. Scaffold proteins (eg., SAV1, CORO7) and kinases (eg., MST1/2, LATS1/2) are potential upstream targets: restoring the expression or activity of these components can reactivate the Hippo tumor-suppressive kinase cascade.18 Phosphatases also play important roles in Hippo pathway regulation. Recent studies identified PTPN12 as a novel negative regulator of YAP/TAZ, and restoration of PTPN12 activity suppresses YAP/TAZ-mediated proliferation and migration in cancer cells, highlighting its potential therapeutic relevance.55 Non-coding RNAs also provide indirect targeting opportunities: targeting lncRNA MALAT1 or restoring miR-181a-5p expression can inhibit YAP activity in MM cells, and this strategy can be extended to other hematological malignancies with dysregulated non-coding RNA-Hippo axes.56
Combination therapy targeting the Hippo pathway and its crosstalk partners is a promising direction to overcome therapeutic resistance. The Hippo pathway interacts extensively with the PI3K/Akt, MAPK, and Notch pathways in hematological malignancies, and simultaneous inhibition of the Hippo pathway and these cascades can produce a synergistic anti-tumor effect.13 For example, combining YAP inhibitors with PI3K/Akt inhibitors can effectively inhibit the growth of AML cells with concurrent activation of PI3K/Akt and YAP.28 In addition, combining Hippo pathway modulators with conventional chemotherapeutic agents or immunotherapies (eg., rituximab, CAR-T therapy) may enhance their efficacy and overcome chemoresistance/immunoresistance in hematological malignancies.35
Clinical Trials and Drug Development
The rapid evolution of Hippo-targeted therapeutics has significantly accelerated the translational potential of this pathway in oncology. Although hematological malignancy-specific clinical trials remain limited, recent advances in solid tumors provide important proof-of-concept evidence supporting future extension into hematologic cancers. A Phase 1b/2a trial evaluated bemcentinib (BGB324), a first-in-class AXL receptor tyrosine kinase inhibitor downstream of YAP/TAZ, in combination with low-dose cytarabine in acute myeloid leukemia patients unfit for intensive chemotherapy, demonstrating the feasibility of targeting YAP/TAZ downstream effectors in hematologic malignancies.57 In particular, TEAD palmitoylation inhibitors have emerged as the most clinically advanced class of Hippo-targeted agents, directly suppressing YAP/TAZ-driven transcriptional programs.58
Among these, VT3989 is currently the leading first-in-class oral TEAD inhibitor. In the first-in-human phase 1/2 trial involving refractory solid tumors, especially mesothelioma, VT3989 demonstrated manageable toxicity, durable disease control, and clear early clinical proof-of-concept for pharmacologic blockade of the Hippo–YAP–TEAD axis.58 Importantly, the objective response rate reached 32% at clinically optimized dosing, with a disease control rate of 86%, highlighting the feasibility of TEAD inhibition in advanced malignancies.59 These findings strongly support the rationale for extending TEAD inhibitors into relapsed or refractory hematological malignancies characterized by YAP/TAZ hyperactivation.58 Notably, most current clinical evidence is derived from solid tumors, highlighting a critical gap in hematological malignancy-specific clinical validation and underscoring the need for dedicated clinical studies in this field.
Another promising agent, IK-930, has completed first-in-human Phase I evaluation in advanced solid tumors and further validates TEAD as a druggable transcriptional dependency node.60 Although these agents are currently evaluated mainly in solid tumors, their mechanistic relevance is highly translatable to hematological malignancies, particularly diffuse large B-cell lymphoma, acute leukemia, and multiple myeloma, where aberrant YAP/TAZ signaling contributes to survival advantage and therapeutic resistance.5
Beyond direct TEAD inhibition, future drug development is increasingly shifting toward rational combination strategies, including co-targeting PI3K/AKT, mTOR, MAPK, and immune checkpoint pathways.59 Given the extensive signaling crosstalk described in hematologic cancers, combination regimens integrating Hippo pathway inhibitors with rituximab, proteasome inhibitors, BCL-2 inhibitors, or CAR-T–based immunotherapy may provide superior efficacy and overcome adaptive resistance.59 These combination paradigms are particularly attractive for biomarker-enriched patient subsets with elevated nuclear YAP/TAZ activity. Recent clinical advances therefore indicate that Hippo pathway modulation is transitioning from a conceptual target into a clinically actionable therapeutic strategy61(Figure 4; Table 3). However, hematological malignancy-specific trials remain urgently needed.
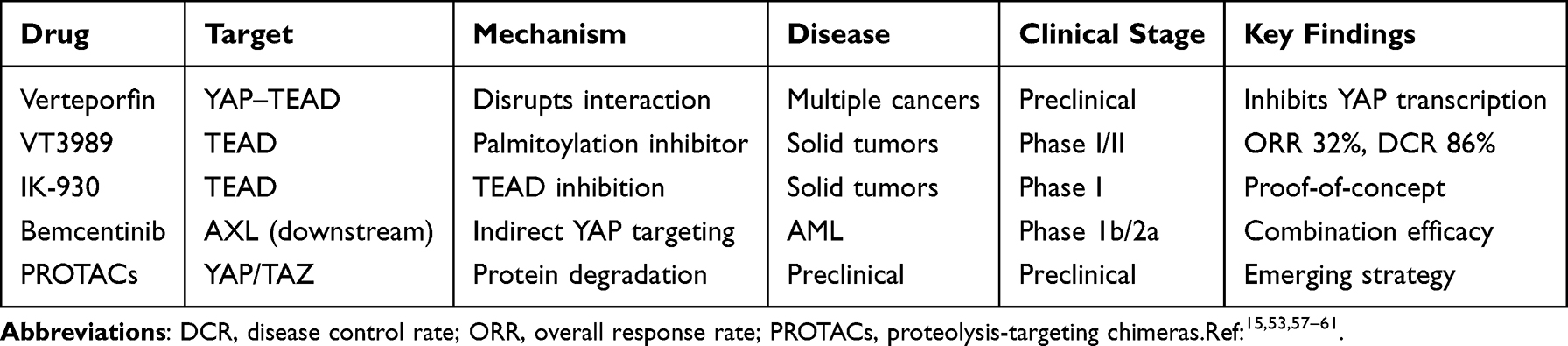 |
Table 3 Hippo Pathway-Targeted Therapeutic Strategies and Clinical Development |
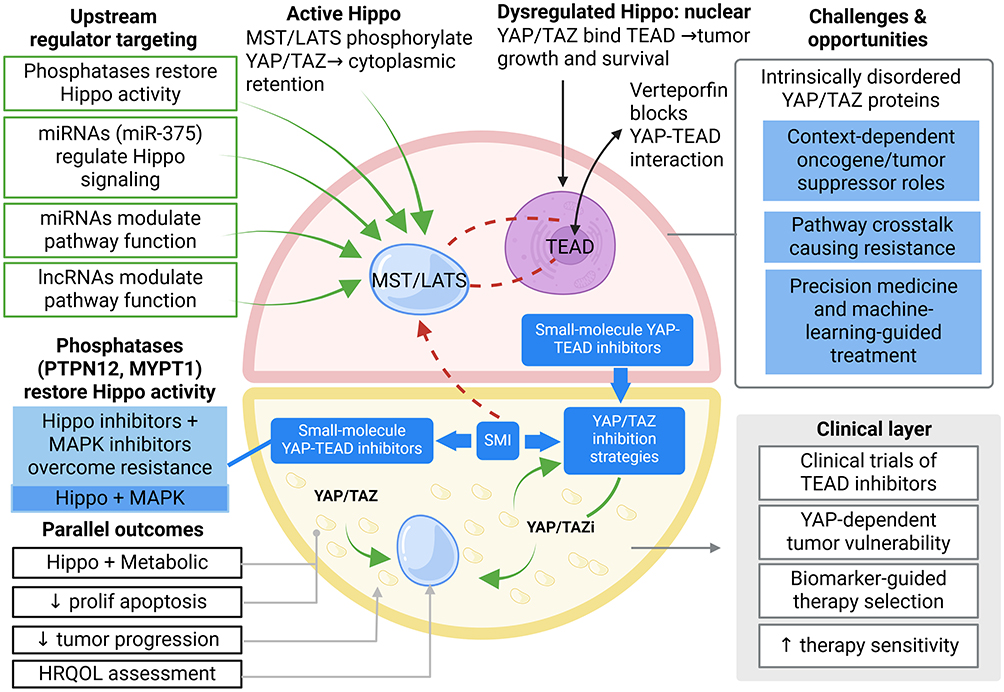 |
Figure 4 Therapeutic targeting strategies and translational opportunities of Hippo signaling in hematological malignancies Therapeutic interventions targeting the Hippo pathway include restoration of upstream kinase activity, inhibition of YAP/TAZ–TEAD transcriptional complexes, and combination therapies integrating Hippo pathway modulators with PI3K/AKT, MAPK, metabolic, or immunotherapeutic approaches. Emerging translational opportunities include biomarker-guided therapy selection, clinical development of TEAD inhibitors, precision medicine strategies, and machine-learning–assisted patient stratification. Key challenges include context-dependent YAP/TAZ functions, pathway crosstalk-mediated resistance, and the intrinsically disordered structure of YAP/TAZ proteins.Symbols: ↑, increased activity, enhanced sensitivity, or therapeutic benefit; ↓, reduced proliferation, tumor progression, or pathway activity; →, signaling activation or downstream consequence; green arrows indicate Hippo pathway activation/restoration; blue arrows indicate therapeutic interventions; red dashed arrows indicate aberrant signaling associated with tumor progression. Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; CML, chronic myeloid leukemia; DLBCL, diffuse large B-cell lymphoma; EMT, epithelial–mesenchymal transition; MM, multiple myeloma; NK/TCL, natural killer/T-cell lymphoma; ROS, reactive oxygen species; TEAD, TEA domain transcription factor. |
Challenges and Opportunities
Although targeting the Hippo pathway holds great promise for the treatment of hematological malignancies, there are several critical challenges that need to be addressed for successful clinical translation, along with promising opportunities to overcome these barriers.
Key Challenges
Context-Dependent Functional Heterogeneity of YAP/TAZ
YAP/TAZ can act as both oncogenes and tumor suppressors in hematological malignancies depending on the cell type and tumor microenvironment.13 For example, TAZ is a tumor suppressor in MM but an oncogene in most lymphomas and leukemias. This functional heterogeneity makes it difficult to develop universal Hippo-targeted therapies and requires subtype-specific treatment strategies.
Clinical Translation Barriers of TEAD and YAP/TAZ Inhibitors
Recent early-phase clinical advances of TEAD inhibitors such as VT3989 and IK-930 have provided proof-of-concept for targetability; however, hematological malignancy-specific efficacy, long-term safety, and resistance mechanisms remain largely undefined. YAP/TAZ are intrinsically disordered proteins with no well-characterized binding pockets, making it challenging to develop small-molecule inhibitors with high specificity and potency.53 Current inhibitors (eg., verteporfin) have limitations such as off-target effects and poor pharmacokinetic properties, and novel drug development strategies (eg., PROTACs, peptide inhibitors) are needed.53
Compensatory Activation of Other Signaling Pathways
The extensive crosstalk between the Hippo pathway and other oncogenic cascades (eg., PI3K/Akt, Wnt/β-catenin) leads to compensatory activation of these pathways upon Hippo pathway inhibition, resulting in therapeutic resistance. Single-target therapies are thus likely to be ineffective, and combination therapies are required.
Lack of Validated Clinical Biomarkers
Potential biomarkers may include nuclear YAP/TAZ localization, TEAD target gene signatures, pathway mutation profiles, and dynamic liquid biopsy–based monitoring approaches.Although YAP/TAZ expression is associated with prognosis in hematological malignancies, there are no validated clinical biomarkers to select patients who are most likely to benefit from Hippo-targeted therapies.13 The development of reliable biomarkers is critical for the clinical translation of Hippo-targeted therapies.
Promising Opportunities
Advances in Molecular Characterization and Precision Medicine
High-throughput sequencing and multi-omics analysis have enabled the comprehensive characterization of Hippo pathway alterations in individual patients, providing a basis for subtype-specific and personalized Hippo-targeted therapies.13 Machine learning and bioinformatics can integrate multi-omics data to predict patient response to Hippo-targeted therapies and identify potential combination targets.13
Novel Drug Development Strategies
Novel drug development approaches such as PROTACs, antibody-drug conjugates (ADCs), and gene therapy are being applied to target the Hippo pathway. For example, PROTACs targeting YAP/TAZ can achieve selective degradation of these proteins, and gene therapy to restore LATS1/2 expression is being explored in preclinical models.53
Combination Therapy with Existing Standard Treatments
Future rational combinations may include TEAD inhibitors with rituximab in DLBCL, BCL-2 inhibition in leukemia, or proteasome inhibitors in multiple myeloma.Combining Hippo-targeted therapies with conventional chemotherapeutic agents, immunotherapies, or other targeted agents can produce a synergistic anti-tumor effect and overcome therapeutic resistance.35 For example, combining YAP inhibitors with rituximab can restore sensitivity in refractory DLBCL.35
Exploitation of the Tumor Microenvironment
The Hippo pathway regulates the interaction between tumor cells and the tumor microenvironment (eg., stromal cells, immune cells).14 Targeting the Hippo pathway in the tumor microenvironment can modulate immune cell infiltration and activation, enhancing the efficacy of immunotherapies.14 This provides a new direction for the development of Hippo-targeted therapies in combination with immunotherapy.
Future Directions
Emerging Research Areas Focused on Hematological Malignancies
Functional Heterogeneity of YAP/TAZ in Distinct Hematopoietic Cell Lineages
Further studies are needed to elucidate the cell type-specific regulatory mechanisms of YAP/TAZ in normal and malignant hematopoietic cells, and to determine the functional consequences of YAP/TAZ activation/inhibition in different subtypes of hematological malignancies. Single-cell transcriptomic studies have begun to resolve the expression patterns of Hippo pathway components in distinct hematopoietic lineages, revealing that YAP/TAZ activity is tightly regulated in a cell-context dependent manner.5 This will provide a basis for the development of subtype-specific Hippo-targeted therapies.18
Crosstalk Between the Hippo Pathway and the Tumor Microenvironment in Hematology
The bone marrow and lymph node microenvironments play a critical role in the progression and therapeutic resistance of hematological malignancies. Future studies should focus on the role of the Hippo pathway in regulating the interaction between malignant hematopoietic cells and stromal cells, immune cells, and endothelial cells in the tumor microenvironment, and explore the potential of targeting the Hippo pathway to remodel the tumor microenvironment.14
Non-Coding RNA-Hippo Regulatory Networks
Non-coding RNAs (lncRNAs, miRNAs, circRNAs) are key regulators of the Hippo pathway in hematological malignancies.62 Future research should identify novel non-coding RNA-Hippo axes in different hematological malignancies and explore their potential as diagnostic biomarkers and therapeutic targets.62
Metabolic-Hippo Crosstalk in Hematological Malignancies
Hematological malignant cells rely on metabolic reprogramming for survival and proliferation, and the Hippo pathway is closely linked to metabolic pathways.63 Future studies should elucidate the specific metabolic-Hippo regulatory mechanisms in hematological malignancies and develop dual-target therapies targeting metabolic pathways and the Hippo pathway.39
Hippo Signaling and Transplantation-Associated Endothelial Injury
Recent studies suggest that Hippo signaling may play a role in endothelial dysfunction after allogeneic hematopoietic stem cell transplantation (allo-HSCT).64,65 Because endothelial injury is a central feature of transplant-associated thrombotic microangiopathy (TA-TMA) and sinusoidal obstruction syndrome/veno-occlusive disease (SOS/VOD), YAP/TAZ-mediated regulation of vascular homeostasis may represent a novel area of investigation.65–67 Understanding this pathway could provide new biomarkers and therapeutic targets for transplant-related complications.64,68,69
Technological Advances Driving Hippo Pathway Research in Hematology
Advanced molecular and computational technologies are providing new tools to study the Hippo pathway in hematological malignancies, deepening our understanding of its regulatory mechanisms and facilitating the development of novel therapeutic strategies.
CRISPR/Cas9 Gene Editing and in vivo Screening
CRISPR/Cas9 technology enables precise knockout/activation of Hippo pathway genes in hematopoietic cells and animal models, providing a powerful tool to study the functional role of the Hippo pathway in leukemogenesis and lymphomagenesis.70 CRISPR-based screening in hematologic malignancies has identified novel Hippo-related vulnerabilities, such as the RHOBTB2-KLHL13 axis in AML, which could be exploited for therapeutic intervention.30 In vivo CRISPR screening can identify novel Hippo-related vulnerabilities and synergistic drug targets in hematological malignancies, accelerating the development of combination therapies.70
Single-Cell RNA Sequencing (scRNA-Seq)
scRNA-seq can resolve the transcriptomic heterogeneity of hematological malignancies at the single-cell level, revealing the expression pattern and activity of the Hippo pathway in distinct tumor cell subpopulations and immune cells.71 This technology can identify rare malignant cell subpopulations with dysregulated Hippo pathway and provide a basis for precision targeting.71
High-Resolution/Live-Cell Imaging
Live-cell and super-resolution microscopy can track the real-time subcellular localization of YAP/TAZ in hematological malignant cells and their dynamic changes in response to extracellular signals (eg., mechanical stress, cytokines).72 This technology can elucidate the spatiotemporal regulation of the Hippo pathway in hematopoietic cells and identify the optimal timing for Hippo-targeted therapy.72
Multi-Omics Integration and Bioinformatics Modeling
Integrating genomics, transcriptomics, proteomics, and metabolomics data can map the complex regulatory network of the Hippo pathway and its crosstalk with other signaling cascades in hematological malignancies.73 Machine learning and computational modeling can predict the response to Hippo-targeted therapies and identify potential combination targets, facilitating the design of personalized treatment strategies73(Table 3).
Precision Medicine: The Core Direction of Hippo-Targeted Therapy in Hematology
The combination of the Hippo pathway and precision medicine is the key direction for the clinical translation of Hippo-targeted therapies in hematological malignancies. The development of precision medicine for Hippo-targeted therapy relies on three core pillars: validated clinical biomarkers, subtype-specific treatment strategies, and personalized combination therapies.
First, identifying and validating clinical biomarkers for Hippo pathway activity is critical for patient stratification. Potential biomarkers include: (i) expression levels of YAP/TAZ; (ii) genetic alterations in Hippo pathway components; and (iii) activity of Hippo pathway target genes.14 These biomarkers should be validated in large clinical cohorts to determine their prognostic and predictive value. Liquid biopsy technologies (eg., circulating tumor DNA, exosomes) can be used to detect Hippo pathway alterations in real time, enabling dynamic monitoring of treatment response and early detection of resistance.74
Second, subtype-specific treatment strategies are needed to address the functional heterogeneity of the Hippo pathway in different hematological malignancies. For example, YAP inhibition is a promising strategy for leukemia and DLBCL, while TAZ activation may be beneficial for MM.18 Subtype-specific therapies should be developed based on the functional role of the Hippo pathway in each subtype, and multi-omics analysis can be used to guide the selection of appropriate therapeutic targets.75
Third, personalized combination therapies should be designed based on the individual genetic and molecular profile of patients. For patients with concurrent activation of the Hippo and PI3K/Akt pathways, combination therapy with YAP and PI3K inhibitors is recommended.28 For patients with dysregulated non-coding RNA-Hippo axes, non-coding RNA-based interventions combined with small-molecule inhibitors may be more effective.62 Machine learning and bioinformatics can integrate multi-omics data to predict the optimal combination therapy for individual patients, maximizing the therapeutic effect and minimizing off-target effects.73
In addition, the integration of artificial intelligence (AI) and big data into Hippo pathway research and clinical translation will further promote the development of precision medicine. AI can be used to analyze large-scale clinical and molecular data to identify novel Hippo-related biomarkers and therapeutic targets, and to design personalized treatment strategies.76 Big data platforms can facilitate the sharing of clinical and molecular data from hematological malignancy patients, accelerating the validation of Hippo-targeted therapies and their clinical translation.71
Conclusion
In conclusion, the Hippo signaling pathway has emerged as a critical regulatory network in both normal hematopoiesis and hematological malignancies.18 Aberrant activation of its downstream effectors—YAP and TAZ—drives malignant proliferation, survival advantage, therapeutic resistance, and unfavorable clinical outcomes.14 The context-dependent functional heterogeneity of YAP/TAZ in hematological malignancies—where they can act as either oncogenes or tumor suppressors depending on the specific lineage and microenvironment—underscores the necessity of precision medicine approaches in Hippo-targeted therapy.5,25,77 Beyond its direct oncogenic effects, the extensive crosstalk between Hippo signaling and other pathways, including PI3K/AKT, TGF-β, Wnt/β-catenin, metabolic signaling, and non-coding RNA networks, further highlights its central role in hematologic tumor biology.13
Recent advances in preclinical studies have demonstrated the therapeutic promise of targeting the Hippo pathway through YAP/TAZ inhibition, modulation of upstream regulators, and rational combination strategies with existing chemotherapeutic agents, targeted therapies, or immunotherapies.5 Nevertheless, several challenges remain, including the context-dependent functional heterogeneity of YAP/TAZ, limited availability of highly specific inhibitors, compensatory activation of parallel oncogenic pathways, and the lack of clinically validated biomarkers for patient stratification.13,14,59,78
With the rapid development of single-cell sequencing, CRISPR-based functional screening, multi-omics integration, and artificial intelligence-assisted biomarker discovery, a more comprehensive understanding of Hippo pathway dysregulation in hematological malignancies is becoming increasingly achievable.70 Future efforts should focus on subtype-specific mechanisms, biomarker-guided patient selection, and personalized combination therapies to accelerate the clinical translation of Hippo-targeted strategies.18 Overall, the Hippo pathway represents a highly promising avenue for precision medicine and may provide new therapeutic opportunities for patients with refractory hematological malignancies.18
Despite substantial progress in understanding Hippo signaling biology, robust clinical evidence supporting Hippo-targeted therapies in hematological malignancies remains scarce.46,79,80 Most available data are derived from preclinical studies, and only a limited number of Hippo-associated therapeutic strategies have entered early-phase clinical evaluation.59,80,81 Future studies should prioritize validation of disease-specific mechanisms, development of predictive biomarkers, and well-designed clinical trials to establish the safety and efficacy of Hippo-targeted interventions in hematologic cancers.29
Data Sharing Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Acknowledgments
I sincerely thank my mentor Li Xu for the selfless help and support provided throughout the process of writing the manuscript. We appreciate the support of the Department of Hematology of the Anhui No.2 Provincial People’s Hospital.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Scientific Research Project of the Education Department of Anhui Province (2023AH053386, 2025AHGXZK31294).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pan D. The hippo signaling pathway in development and cancer. Dev Cell. 2010;19(4):491–18. doi:10.1016/j.devcel.2010.09.011
2. Pulte D, Jansen L, Brenner H. Changes in long term survival after diagnosis with common hematologic malignancies in the early 21st century. Blood Cancer J. 2020;10(5):56. doi:10.1038/s41408-020-0323-4
3. Hofmann WK, Trumpp A, Müller-Tidow C. Therapy resistance mechanisms in hematological malignancies. Int, J, Cancer. 2023;152(3):340–347. doi:10.1002/ijc.34243
4. Zhang J, Gu Y, Chen B. Mechanisms of drug resistance in acute myeloid leukemia. Onco Targets Ther. 2019;12:1937–1945. doi:10.2147/OTT.S191621
5. Baroja I, Kyriakidis NC, Halder G, Moya IM. Expected and unexpected effects after systemic inhibition of Hippo transcriptional output in cancer. Nat Commun. 2024;15(1):2700. doi:10.1038/s41467-024-46531-1
6. Wei AH, Roberts AW, Spencer A, et al. Targeting MCL-1 in hematologic malignancies: rationale and progress. Blood Rev. 2020;44:100672. doi:10.1016/j.blre.2020.100672
7. Bolomsky A, Vogler M, Köse MC, et al. MCL-1 inhibitors, fast-lane development of a new class of anti-cancer agents. J Hematol Oncol. 2020;13(1):173. doi:10.1186/s13045-020-01007-9
8. Ma S, Meng Z, Chen R, Guan KL. The hippo pathway: biology and pathophysiology. Annu Rev Biochem. 2019;88:577–604. doi:10.1146/annurev-biochem-013118-111829
9. Li Z, Lin J, Wu J, Suo J, Wang Z. The Hippo signalling pathway in bone homeostasis: under the regulation of mechanics and aging. Cell Prolif. 2024;57(10):e13652. doi:10.1111/cpr.13652
10. Noorbakhsh N, Hayatmoghadam B, Jamali M, Golmohammadi M, Kavianpour M. The Hippo signaling pathway in leukemia: function, interaction, and carcinogenesis. Cancer Cell Int. 2021;21(1):705. doi:10.1186/s12935-021-02408-7
11. Marsola A, Simões BP, Palma LC, Berzoti-Coelho MG, Burin SM, de Castro FA. Expression of Hippo signaling pathway and Aurora kinase genes in chronic myeloid leukemia. Med Oncol. 2018;35(3):26. doi:10.1007/s12032-018-1079-6
12. Safari S, Movafagh A, Zare-Adollahi D, et al. MST1/2 and YAP1 gene expression in acute myeloid leukemia. Leuk Lymphoma. 2014;55(9):2189–2191. doi:10.3109/10428194.2013.867493
13. Zhang J, Wu H, Ren X, et al. Hippo/YAP signaling’s multifaceted crosstalk in cancer. Front Cell Dev Biol. 2025;13:1595362. doi:10.3389/fcell.2025.1595362
14. Kim HS, Nam JS. The multifaceted role of YAP in the tumor microenvironment and its therapeutic implications in cancer. Exp Mol Med. 2025;57(10):2201–2213. doi:10.1038/s12276-025-01551-9
15. Liang J, Wang L, Wang C, et al. Verteporfin Inhibits PD-L1 through Autophagy and the STAT1-IRF1-TRIM28 signaling axis, exerting antitumor efficacy. Cancer Immunol Res. 2020;8(7):952–965. doi:10.1158/2326-6066.CIR-19-0159
16. Carter BZ, Mak PY, Tao W, et al. Targeting MCL-1 dysregulates cell metabolism and leukemia-stroma interactions and resensitizes acute myeloid leukemia to BCL-2 inhibition. Haematologica. 2022;107(1):58–76. doi:10.3324/haematol.2020.260331
17. Cunningham R, Hansen CG. The Hippo pathway in cancer: YAP/TAZ and TEAD as therapeutic targets in cancer. Clin Sci. 2022;136(3):197–222. doi:10.1042/CS20201474
18. Bala R, Madaan R, Bedi O, et al. Targeting the Hippo/YAP Pathway: a Promising Approach for Cancer Therapy and Beyond. MedComm. 2025;6(9):e70338. doi:10.1002/mco2.70338
19. Bae SJ, Ni L, Osinski A, Tomchick DR, Brautigam CA, Luo X. SAV1 promotes Hippo kinase activation through antagonizing the PP2A phosphatase STRIPAK. Elife. 2017;6.
20. Zhong B, Du J, Liu F, Sun S. The role of yes-associated protein in inflammatory diseases and cancer. MedComm. 2025;6(3):e70128. doi:10.1002/mco2.70128
21. Lim JY, Choi EH, Kim Y, et al. Identification of YAP regulators through high-throughput screening and NanoBiT-based validation-drug repositioning for cancer therapy. Anim Cells Syst. 2025;29(1):325–338. doi:10.1080/19768354.2025.2489389
22. Zhao B, Li L, Tumaneng K, Wang CY, Guan KL. A coordinated phosphorylation by Lats and CK1 regulates YAP stability through SCF(beta-TRCP). Genes Dev. 2010;24(1):72–85. doi:10.1101/gad.1843810
23. Barral L, Lespinasse N, Martin Cardozo C, et al. Targeting BMP and TAZ/TEAD mechanotransduction pathways impairs acute myeloid leukemia chemoresistance. Leukemia. 2026;40:1151–1162. doi:10.1038/s41375-026-02904-7
24. Wang G, Lai Y, Chen X, et al. Hexokinase 2 promotes tumor development and progression. Am J Cancer Res. 2025;15(10):4499–4515. doi:10.62347/ZYNN3077
25. Bruno T, Catena V, Corleone G, et al. Che-1/miR-590-3p/TAZ axis sustains multiple myeloma disease. Leukemia. 2024;38(4):877–882. doi:10.1038/s41375-024-02168-z
26. Grieve S, Wajnberg G, Lees M, et al. TAZ functions as a tumor suppressor in multiple myeloma by downregulating MYC. Blood Adv. 2019;3(22):3613–3625. doi:10.1182/bloodadvances.2019000374
27. Ajongbolo AO, Langhans SA. YAP/TAZ-associated cell signaling - at the crossroads of cancer and neurodevelopmental disorders. Front Cell Dev Biol. 2025;13:1522705. doi:10.3389/fcell.2025.1522705
28. Garcia KC, Khan AA, Ghosh K, et al. PI3K regulates TAZ/YAP and mTORC1 axes that can be synergistically targeted. JCI Insight. 2026;11(6). doi:10.1172/jci.insight.191600.
29. Piccolo S, Panciera T, Contessotto P, Cordenonsi M. YAP/TAZ as master regulators in cancer: modulation, function and therapeutic approaches. Nat Cancer. 2023;4(1):9–26. doi:10.1038/s43018-022-00473-z
30. Liu Y, Zhou F, Wang L, Xue Y, Wang W. RHOBTB2 enhances cell proliferation of acute myeloid leukemia by modulating Hippo-YAP1 signaling and dependent of KLHL13. Sci Rep. 2025;15(1):36428. doi:10.1038/s41598-025-20492-x
31. Wu R, Yang H, Wan J, et al. Knockdown of the Hippo transducer YAP reduces proliferation and promotes apoptosis in the Jurkat leukemia cell. Mol Med Rep. 2018;18(6):5379–5388. doi:10.3892/mmr.2018.9556
32. Feng P, Zhang J, Zhang J, et al. Deacetylation of YAP1 Promotes the Resistance to Chemo- and Targeted Therapy in FLT3-ITD(+) AML Cells. Front Cell Dev Biol. 2022;10:842214. doi:10.3389/fcell.2022.842214
33. Chorzalska A, Kim JF, Roder K, et al. Long-term exposure to imatinib mesylate downregulates hippo pathway and activates yap in a model of chronic myelogenous leukemia. Stem Cells Dev. 2017;26(9):656–677. doi:10.1089/scd.2016.0262
34. Li H, Huang Z, Gao M, et al. Inhibition of YAP suppresses CML cell proliferation and enhances efficacy of imatinib in vitro and in vivo. J Exp Clin Cancer Res. 2016;35(1):134. doi:10.1186/s13046-016-0414-z
35. Li Q, Ma N, Li X, et al. Reverse effect of Semaphorin-3F on rituximab resistance in diffuse large B-cell lymphoma via the Hippo pathway. Chin Med J. 2023;136(12):1448–1458. doi:10.1097/CM9.0000000000002686
36. Baghdadi H, Heidari R, Zavvar M, et al. Long Non-Coding RNA Signatures in Lymphopoiesis and Lymphoid Malignancies. Noncoding RNA. 2023;9(4). doi:10.3390/ncrna9040044.
37. Zheng J, Yu H, Zhou A, et al. It takes two to tango: coupling of Hippo pathway and redox signaling in biological process. Cell Cycle. 2020;19(21):2760–2775. doi:10.1080/15384101.2020.1824448
38. Zhan C, Zhou H, Zhang W, Si C. Hsa_circ_0000877 facilitates the progression of diffuse large B-cell lymphoma by miR-370-3p/mitogen-activated protein kinase kinase kinase kinase 4/Hippo pathway. Anticancer Drugs. 2022;33(10):1091–1102. doi:10.1097/CAD.0000000000001366
39. Li H, Zhang Y, Mou X, Huang B, Fan X. Interference with PLA2G16 promotes cell cycle arrest and apoptosis and inhibits the reprogramming of glucose metabolism in multiple myeloma cells by modulating the Hippo/YAP signaling pathway. Anticancer Drugs. 2024;35(10):902–911. doi:10.1097/CAD.0000000000001642
40. Sun Y, Jiang T, Jia Y, Zou J, Wang X, Gu W. LncRNA MALAT1/miR-181a-5p affects the proliferation and adhesion of myeloma cells via regulation of Hippo-YAP signaling pathway. Cell Cycle. 2019;18(19):2509–2523. doi:10.1080/15384101.2019.1652034
41. Abegunde SO, Grieve S, Reiman T. TAZ upregulates MIR-224 to inhibit oxidative stress response in multiple myeloma. Cancer Rep. 2023;6(10):e1879.
42. LeBlanc L, Ramirez N, Kim J. Context-dependent roles of YAP/TAZ in stem cell fates and cancer. Cell Mol Life Sci. 2021;78(9):4201–4219. doi:10.1007/s00018-021-03781-2
43. Luo J, Deng L, Zou H, et al. New insights into the ambivalent role of YAP/TAZ in human cancers. J Exp Clin Cancer Res. 2023;42(1):130. doi:10.1186/s13046-023-02704-2
44. Moroishi T, Hansen CG, Guan KL. The emerging roles of YAP and TAZ in cancer. Nat Rev Cancer. 2015;15(2):73–79. doi:10.1038/nrc3876
45. Zhou X, Chen N, Xu H, et al. Regulation of Hippo-YAP signaling by insulin-like growth factor-1 receptor in the tumorigenesis of diffuse large B-cell lymphoma. J Hematol Oncol. 2020;13(1):77. doi:10.1186/s13045-020-00906-1
46. Anderson AM, Bailetti AA, Rodkin E, De A, Bach EA. A Genetic Screen Reveals an Unexpected Role for Yorkie Signaling in JAK/STAT-Dependent Hematopoietic Malignancies in Drosophila melanogaster. G3. 2017;7(8):2427–2438. doi:10.1534/g3.117.044172
47. Zhao T, Wang Z, Fang J, et al. HTLV-1 activates YAP via NF-κB/p65 to promote oncogenesis. Proc Natl Acad Sci U S A. 2022;119(9).
48. Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the Roots of Cancer. Cancer Cell. 2016;29(6):783–803. doi:10.1016/j.ccell.2016.05.005
49. Cottini F, Hideshima T, Xu C, et al. Editorial Expression of Concern: rescue of Hippo coactivator YAP1 triggers DNA damage-induced apoptosis in hematological cancers. Nat Med. 2026;32(3):1161. doi:10.1038/s41591-026-04295-9
50. Panciera T, Azzolin L, Cordenonsi M, Piccolo S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol. 2017;18(12):758–770. doi:10.1038/nrm.2017.87
51. Ma X, Zhou Z, Gu S, et al. Advances in the Diagnosis and Treatment of Myeloproliferative Neoplasms (MPNs). Cancers. 2025;17(19):3142. doi:10.3390/cancers17193142
52. Sun Z, Zhang Q, Yuan W, et al. MiR-103a-3p promotes tumour glycolysis in colorectal cancer via hippo/YAP1/HIF1A axis. J Exp Clin Cancer Res. 2020;39(1):250. doi:10.1186/s13046-020-01705-9
53. Lao Z, Chen X, Pan B, Fang B, Yang W, Qian Y. Pharmacological regulators of Hippo pathway: advances and challenges of drug development. FASEB j. 2025;39(6):e70438. doi:10.1096/fj.202401895RR
54. Pobbati AV, Kumar R, Rubin BP, Hong W. Therapeutic targeting of TEAD transcription factors in cancer. Trends Biochem Sci. 2023;48(5):450–462. doi:10.1016/j.tibs.2022.12.005
55. Sarmasti Emami S, Ge A, Zhang D, et al. Identification of PTPN12 Phosphatase as a Novel Negative Regulator of Hippo Pathway Effectors YAP/TAZ in Breast Cancer. Int J Mol Sci. 2024;25(7):4064. doi:10.3390/ijms25074064
56. Yang C, Liang Y, Shu J, et al. Long non‑coding RNAs in multiple myeloma (Review). Int J Oncol. 2023;62(6). doi:10.3892/ijo.2023.5517.
57. Loges S, Heuser M, Chromik J, et al. Bemcentinib as monotherapy and in combination with low-dose cytarabine in acute myeloid leukemia patients unfit for intensive chemotherapy: a phase 1b/2a trial. Nat Commun. 2025;16(1):2846. doi:10.1038/s41467-025-58179-6
58. Yap TA, Kwiatkowski DJ, Dagogo-Jack I, et al. YAP/TEAD inhibitor VT3989 in solid tumors: a phase 1/2 trial. Nat Med. 2025;31(12):4281–4290. doi:10.1038/s41591-025-04029-3
59. Battina R, Rahhal R, Wellstein A, Riegel AT, Sharif GM. Targeting TEAD in cancer. Front Oncol. 2025;15:1692512. doi:10.3389/fonc.2025.1692512
60. Zhang Z, He P, Yang L, Gong J, Qin R, Wang M. Posttranslational modifications of YAP/TAZ: molecular mechanisms and therapeutic opportunities. Cell Mol Biol Lett. 2025;30(1):83. doi:10.1186/s11658-025-00760-4
61. Harvey KF, Tang TT. Targeting the Hippo pathway in cancer. Nat Rev Drug Discov. 2025;24(11):852–869. doi:10.1038/s41573-025-01234-0
62. Joulani M, Aghajamal N, Nouri M, et al. Non-coding RNAs and Hippo signaling in non-small cell lung cancer: emerging roles as biomarkers and therapeutic targets. Discov Oncol. 2025;16(1):1365. doi:10.1007/s12672-025-03099-6
63. Yu M, Yang D, Chen X, et al. Metabolic reprogramming in cancer: dysregulation of glucose, lipid, and amino acid pathways and therapeutic opportunities. Mol Biomed. 2026;7(1). doi:10.1186/s43556-026-00427-2.
64. Mavrikou I, Castelli M, Touloumenidou T, et al. Genetic susceptibility in sinusoidal obstruction syndrome/veno-occlusive disease: a case-control study. Int J Mol Sci. 2025;26(14):6712. doi:10.3390/ijms26146712
65. Luft T, Dreger P, Radujkovic A. Endothelial cell dysfunction: a key determinant for the outcome of allogeneic stem cell transplantation. Bone Marrow Transplant. 2021;56(10):2326–2335. doi:10.1038/s41409-021-01390-y
66. Vythoulkas D, Tsirigotis P, Griniezaki M, Konstantellos I, Lazana I. Endothelial dysfunction syndromes after allogeneic stem cell transplantation. Cancers. 2023;15(3):680. doi:10.3390/cancers15030680
67. Bonifazi F, Barbato F, Ravaioli F, et al. Diagnosis and Treatment of VOD/SOS After Allogeneic Hematopoietic Stem Cell Transplantation. Front Immunol. 2020;11:489. doi:10.3389/fimmu.2020.00489
68. Moreno-Castaño AB, Salas MQ, Palomo M, et al. Early vascular endothelial complications after hematopoietic cell transplantation: role of the endotheliopathy in biomarkers and target therapies development. Front Immunol. 2022;13:1050994. doi:10.3389/fimmu.2022.1050994
69. Wang KC, Yeh YT, Nguyen P, et al. Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis. Proc Natl Acad Sci U S A. 2016;113(41):11525–11530. doi:10.1073/pnas.1613121113
70. He Y, Tu X, Xue Y, et al. CRISPR screening redefines therapeutic target identification and drug discovery with precision and scalability. J Pharm Anal. 2026;16(2):101357. doi:10.1016/j.jpha.2025.101357
71. Hsu CY, Askar S, Alshkarchy SS, et al. AI-driven multi-omics integration in precision oncology: bridging the data deluge to clinical decisions. Clin Exp Med. 2025;26(1):29. doi:10.1007/s10238-025-01965-9
72. Wehling L, Keegan L, Fernández-Palanca P, et al. Spatial modeling reveals nuclear phosphorylation and subcellular shuttling of YAP upon drug-induced liver injury. Elife. 2022;11.
73. Marouf AA, Rokne JG, Alhajj R. Integrating multi-omics and medical imaging in artificial intelligence-based cancer research: an umbrella review of fusion strategies and applications. Cancers. 2025;17(22):3638. doi:10.3390/cancers17223638
74. Hao X, Liu Z, Ma F, et al. Exosome-based liquid biopsy in early screening and diagnosis of cancers. Dose Response. 2025;23(2):15593258251344480. doi:10.1177/15593258251344480
75. Khoury R, Raffoul C, Khater C, Hanna C. Precision medicine in hematologic malignancies: evolving concepts and clinical applications. Biomedicines. 2025;13(7). doi:10.3390/biomedicines13071654
76. Li D, Shi S, Yu Z, Xu P, Zhang C. AI accelerate the identification of druggable targets by 3D structures of proteins and compounds. NPJ Precis Oncol. 2026;10(1).
77. Klaihmon P, Lorthongpanich C, Kheolamai P, Saisaard W, Issaragrisil S. Inhibition of LATS kinases reduces tumorigenicity and increases the sensitivity of human chronic myelogenous leukemia cells to imatinib. Sci Rep. 2024;14(1):3993. doi:10.1038/s41598-024-54728-z
78. Nguyen CDK, Yi C. YAP/TAZ Signaling and Resistance to Cancer Therapy. Trends Cancer. 2019;5(5):283–296. doi:10.1016/j.trecan.2019.02.010
79. Allegra A, Pioggia G, Innao V, Musolino C, Gangemi S. New Insights into YES-associated protein signaling pathways in hematological malignancies: diagnostic and therapeutic challenges. Cancers. 2021;13(8):1981. doi:10.3390/cancers13081981
80. Dey A, Varelas X, Guan KL. Targeting the Hippo pathway in cancer, fibrosis, wound healing and regenerative medicine. Nat Rev Drug Discov. 2020;19(7):480–494. doi:10.1038/s41573-020-0070-z
81. Chapeau EA, Sansregret L, Galli GG, et al. Direct and selective pharmacological disruption of the YAP-TEAD interface by IAG933 inhibits Hippo-dependent and RAS-MAPK-altered cancers. Nat Cancer. 2024;5(7):1102–1120. doi:10.1038/s43018-024-00754-9
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.