Back to Journals » Infection and Drug Resistance » Volume 19
High-Resolution Melting Curve Analysis (HRMA) for the Identification of Class D β-Lactamases (CHDLs) in Pseudomonas aeruginosa Lung Infection
Authors Tahmasebi H ![]() , Dehbashi S
, Dehbashi S ![]() , Arabestani MR
, Arabestani MR
Received 6 May 2026
Accepted for publication 14 July 2026
Published 17 July 2026 Volume 2026:19 620945
DOI https://doi.org/10.2147/IDR.S620945
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Hazrat Bilal
Hamed Tahmasebi,1,2 Sanaz Dehbashi,3 Mohammad Reza Arabestani4,5
1School of Medicine, Shahroud University of Medical Sciences, Shahroud, Iran; 2Department of Bacteriology, Pasteur Institute of Iran, Tehran, Iran; 3Department of Laboratory Sciences, Varastegan Institute of Medical Sciences, Mashhad, Iran; 4Microbiology Department, Faculty of Medicine, Hamadan University of Medical Sciences, Hamadan, Iran; 5Nutrition Health Research Center, Institute of Health Sciences and Technology, Hamadan University of Medical Sciences, Hamadan, Iran
Correspondence: Mohammad Reza Arabestani, Microbiology Department, Faculty of Medicine, Hamadan University of Medical Sciences, Pajoohesh Junction, Hamadan, Iran, Tel +989188662009, Email [email protected]
Objective: Conventional approaches for laboratory detection of Carbapenem-hydrolyzing class D β-lactamases (CHDLs) from clinical isolates of Pseudomonas aeruginosa are laborious, slow, and have limited sensitivity. This study presents a novel high-resolution melting curve analysis (HRMA) assay for the rapid molecular characterization of CHDLs in P. aeruginosa lung infections.
Methods: Detection of blaOXA genes in CHDLs was performed using an HRMA assay. ABI Step One-Plus Manager Software version 3.2 and Precision Melt Analysis Software version 3.02 (Applied Biosystems) were used to analyze a wide range of HRMA data.
Results: Out of the 47 P. aeruginosa MBL-producing strains, 18 (38.2%) were blaOXA145 positive, 24 (51.0%) were blaOXA-161 positive, 20 (42.5%) were blaOXA-224 positive, 29 (61.7%) were blaOXA-539 positive, 7 (14.8%) were blaOXA-675 positive, and 19 (40.4%) were blaOXA848 positive. Distinct, non-overlapping melting peaks (eg, 81.70°C for blaOXA-145, 87.35°C for blaOXA-848) were achieved with an accuracy of ± 0.1– 0.5°C, enabling unambiguous genotype identification. Clinically, multi-locus sequence typing (MLST) of 100 isolates revealed that the dominant sequence types—ST09, ST15, ST111, and ST235—were significantly associated with MDR/XDR phenotypes, biofilm formation, and CHDL carriage (p < 0.05). ST235 and ST09 predominated among CHDL producers, and 57% of isolates were new or singleton sequence types, highlighting regional genetic diversity.
Conclusion: Novelty lies in the precise differentiation of six clinically relevant OXA alleles exclusively through unique melting temperature (Tm) shifts and melt curve morphologies, eliminating the need for fluorescent probes. In practice, HRMA offers significant advantages over conventional methods, such as the modified Hodge test or Sanger sequencing. As a closed-tube, single-step assay, it eliminates post-amplification handling, reduces cross-contamination risks, and provides actionable results within two hours. This marked reduction in turnaround time and reagent costs makes HRMA a highly scalable, user-friendly tool, ideally suited for routine clinical microbiology laboratories to expedite antimicrobial stewardship and epidemiological surveillance in P. aeruginosa pulmonary infections.
Keywords: Pseudomonas aeruginosa, high-resolution melting curve analysis, HRMA, New Delhi metallo-β-lactamase, NDM
Introduction
Respiratory tract infections (RTIs) include both upper and lower respiratory tract infections. Patients in the ICU may be exposed to Pseudomonas aeruginosa infections, either endogenous or exogenous.1–3 However, most RTIs are treated with antibiotics. P. aeruginosa has a considerable influence due to increasing resistance to broad-spectrum antibiotics and carbapenems. Reduced susceptibility to β-lactams and carbapenems is increasingly a problem worldwide.4–7
Class D β-lactamases (CHDLs) are usually found in Acinetobacter species; however, reports have described CHDLs in Enterobacteriaceae, especially P. aeruginosa, which produces blaOXA-48, which hydrolyses penicillins and carbapenems.5,8 CHDLs are classified into several clusters based on the similarity of the sequences: blaOXA-23, blaOXA-24, blaOXA-51, and blaOXA-58. Other less common groups, such as blaOXA-72 and blaOXA-143, are becoming clinically significant.9,10 Oxacillinase (OXA) enzymes are members of this group of carbapenemases that exhibit relatively low anti-carbapenem activity. Non-fermenters, Acinetobacter baumannii, and P. aeruginosa, as well as some rare Enterobacteriaceae strains, are mostly associated with these enzymes.11,12
Although CHDLs poorly hydrolyze carbapenems, they still have an effect and can be inhibited by clavulanic acid. There were extended-spectrum β-lactamases and, at least, carbapenemases among 102 different sequences of blaOXA identified over the years.13,14 There are numerous methods in clinical labs used to detect CHDLs: molecular methods, automated systems, determination of minimum inhibitory concentrations (MICs), selective agar, the modified Hodge test, and disc diffusion. Rapid identification of the causal organism and the gene conferring resistance allows prompt use of appropriate broad-spectrum therapy and saves money.11,15,16 Different MIC interpretations in the laboratory may mislead clinicians regarding the use of extended-spectrum cephalosporins, carbapenems, or neither.13,16,17
High-Resolution Melting Curve Analysis (HRMA) is a melting-curve method that measures the tendency of the strand to dissociate. Furthermore, the information capacity of HRMA is significantly improved, enabling the distinction of melt curve shapes derived from different amplicons despite having the same melting temperature.18,19 The emergence of HRMA technology has sparked renewed interest in melt curve analysis across numerous applications, including gene scanning, sequence matching, DNA methylation quantification, and linkage mapping. Due to its high accuracy and simplicity, high-resolution DNA melting has become increasingly popular.20,21
The idea behind HRMA technology is both simple and strong. Once amplified by PCR with a fluorescent dye, the solution is subjected to a gradual thermal denaturation gradient. As temperature increases, the amplicon will start to dissociate and break into single strands, consequently releasing bound fluorescence and reducing fluorescence intensity.22,23 The resulting melting curve is drawn from the ratio of –dF/dT versus T and shows the melting peak at 50% of DNA duplex dissociation. Crucially, each amplicon has a different Tm and melt-curve profile, allowing for the unique identification of DNA sequences without the need for post-amplification treatment, electrophoresis or expensive probe chemistry.24–26
There are several features of the HRMA that make it especially suitable for diagnostics. First, as a closed-tube, one-step assay, HRMA circumvents the risk of amplicon carry-over contamination characteristic of standard PCR techniques that require further handling after amplification.23,27 Second, this technique is fast, with test results obtained in 1.5–2 hours after sample preparation, and requires only standard real-time PCR equipment, which is becoming more widely available in clinical laboratories.28,29
In recent years, gene sequencing has been used to study populations infected with P. aeruginosa. The introduction of MLST techniques enabled molecular epidemiology to trace and monitor the global transmission of lethal or antibiotic-resistant bacterial pathogens.30,31 Epidemiological surveys discovered several high-risk P. aeruginosa clones – ST235, ST111, ST175 and ST253 that are usually resistant to carbapenems and possess enhanced virulence characteristics.32,33 In particular, P. aeruginosa clone ST235 is classified as an international high-risk clone because it tends to acquire multiple resistance genes encoding MBLs, KPC enzymes and CHDLs and is involved in infections of critically ill patients.32,33 The geographical variation in the occurrence and clonal distribution of P. aeruginosa producing CHDLs is remarkable, highlighting the importance of the local epidemiology of this pathogen for the development of effective infection control practices.34–36
Although CHDLs are of great importance in P. aeruginosa infections, the epidemiology of CHDLs in Iran has not been fully explored, and no studies have used high-resolution melt analysis to detect these resistance genes in clinical isolates from this region. Therefore, in this study, an attempt was made to fill this knowledge gap by characterizing CHDLs among P. aeruginosa clinical isolates collected from patients with lung infections in Hamadan province, western Iran. The aims of the current study included: (i) determining the prevalence of MDR and XDR phenotypes among P. aeruginosa clinical isolates with lung infection; (ii) developing and validating a new HRMA assay for rapid detection of six clinically important blaOXA genes; (iii) conducting a molecular epidemiology study on CHDL-positive isolates through MLST; and (iv) finding the correlation between CHDL carriage, antibiotic resistance profiles, ability to form biofilms, and clonality.
Materials and Methods
Design of Study
The current cross-sectional study design involved a prospective one-year study carried out from February 2020 to January 2021 at four affiliated hospital laboratories at Hamadan University of Medical Sciences, Hamadan, Iran. All patients with bacterial lung infection, regardless of sex, met the inclusion criteria for the present study. Exclusions included all patients from outpatient clinics and non-bacterial cases. Approval from the Institutional Human Ethics Committee of Hamadan University of Medical Sciences was obtained before conducting the study (IR.UMSHA.REC.1398.1007). Informed written consent was obtained from all adult participants, and parent/legal guardian consent from all child participants.
A total of 967 respiratory specimens were collected from hospitalized patients with clinically suspected lower respiratory tract infections. Respiratory specimens include 213 endotracheal aspirate specimens, 199 sputum specimens, 165 pleural fluid specimens, 169 throat swab specimens, 120 nasal swab specimens, and 101 bronchoalveolar lavage (BAL) fluids. All respiratory specimens were collected aseptically and transported immediately to the microbiology laboratory within 2 hours of collection.
The clinical specimens were initially isolated on MacConkey agar (Hi-Media, India) and Cetrimide agar (Hi-Media, India). Both media were incubated aerobically at 35 ± 2°C for 18–24 hours. Isolates presumed to be Pseudomonas aeruginosa were confirmed through colony characteristics (greenish pigmentation on cetrimide agar and non-lactose fermenting colonies on MacConkey agar) along with Gram staining (Gram-negative bacilli) and standard biochemical tests, which include the oxidase test, catalase test, growth at 42°C, nitrate reduction, arginine dihydrolase activity, and citrate utilization.37 The quality control strain used for Pseudomonas aeruginosa was P. aeruginosa ATCC 27853.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility patterns of all isolated P. aeruginosa strains were determined using the E-test method (bioMérieux, France) according to the Clinical and Laboratory Standards Institute 2023 (CLSI) M100 guidelines.35 Antimicrobials used for testing comprised amikacin (AMK), gentamicin (GEN), ciprofloxacin (CIP), norfloxacin (NOR), ceftazidime (CAZ), cefepime (CPE), ceftriaxone (CRO), cefoxitin (FOX), cefazolin (CZN), piperacillin (PIP), imipenem (IMI), meropenem (MER), ertapenem (ERT), and colistin (COL). The results were interpreted based on CLSI 2023 breakpoints.38
MDR strains were defined as those exhibiting non-susceptibility to at least one antimicrobial from three or more antimicrobial categories. XDR strains were defined as those exhibiting non-susceptibility to at least one antimicrobial in all but 2 or fewer antimicrobial categories. PDR strains were defined as those exhibiting non-susceptibility to all tested antimicrobials, including colistin.39
Imipenem-EDTA Disc Method (IED)
Two Imipenem discs, each containing 10 µg of Imipenem, were applied to Mueller-Hinton (MH) agar inoculated with a bacterial suspension at 0.5 McFarland turbidity. Afterward, one of the Imipenem discs was exposed to 10 µL of 0.5 M EDTA solution. After incubation for 16 hours at 35°C ± 2°C, the zone of inhibition around the Imipenem disc and the Imipenem + EDTA disc was evaluated. An increase in the zone of inhibition of more than 7 mm in the Imipenem + EDTA disc compared to the Imipenem disc alone suggests production of metallo-β-lactamase (MBL) by the bacterial suspension.24
Phenotypic Detection of Carbapenemases
The modified Hodge test (MHT) was performed according to CLSI guidelines as a screening test for carbapenemase production.40,41 Specifically, a 0.5 McFarland suspension of E. coli ATCC 25922 in sterile saline was prepared and diluted 1:10. The diluted suspension was then streaked in MHA to create a lawn. A 10 μg imipenem disk was placed at the center of the plate, and the test strain was streaked in a straight line from the edge of the disk towards the plate periphery. Growth after incubation at 35°C for 18–24 hours was considered a positive result if a cloverleaf-like zone of distortion was observed where the test organism streak intersected the E. coli lawn.
The Carba NP test was conducted with modifications as mentioned previously.40 The bacterial lysates were prepared by sonication of a bacterial suspension in 0.9% saline. The lysate was mixed with phenol red indicator solution containing imipenem in microtiter plates. A color change from red to yellow/orange indicated hydrolysis of imipenem and was considered a positive result. The positive control strains used in this test were Klebsiella pneumoniae ATCC BAA-1705 (KPC-producing strain) and Citrobacter freundii CRE 3123 (NDM producer).22
The imipenem-EDTA (IED) disc method was used to detect phenotypic MBL production. Two 10 μg imipenem discs were placed on an MHA plate that was inoculated with a 0.5 McFarland bacterial suspension. Ten microliters of 0.5 M EDTA solution was then added to one of the imipenem discs. Incubation of the plate at 35 ± 2°C for 18 hours, followed by measuring the diameter of the zone of inhibition for both discs, was performed. An increase in the zone diameter of ≥7 mm for the imipenem-EDTA disc compared to the imipenem disc alone was considered a positive test result.42
Biofilm Production Assay
The biofilm-forming ability of the bacterial isolates was analyzed using the Congo Red Agar (CRA) method as previously described.43 CRA medium was prepared by adding sucrose (5%) and Congo red (0.8 g/L) to brain heart infusion (BHI) agar. The test bacteria were streaked on CRA plates and incubated aerobically at 37°C for 24–48 hours. Interpretation of the colonies was carried out as follows: black, crystalline-like colonies reflected a high level of biofilm formation; black colonies showed moderate biofilm formation; red colonies reflected a low level of biofilm formation; and pink colonies reflected no biofilm formation.44
DNA Extraction
Genomic DNA was extracted from overnight cultures grown on Luria-Bertani (LB) agar plates using the QIAamp DNA Mini Kit (Qiagen, Germany). The steps involved slight modifications of the manufacturer’s protocol. Briefly, a single colony was first suspended in 200 μL of the enzymatic lysis buffer containing lysozyme (20 mg/mL) and incubated at 37°C for 30 min. Afterward, proteinase K and AL buffer were added, and the mixture was incubated at 56°C for 30 min. Ethanol was added, followed by the application of the lysate to the QIAamp spin column. The purified DNA was eluted in 50 μL of AE buffer and stored at −20°C. The DNA concentrations and purity were measured using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA). A260/A280 ratios of 1.8–2.0 were considered satisfactory.
Primer Design for blaOXA Gene Detection
Based on Table 1, two primer pairs can be used to amplify clinically relevant CHDL genes, specifically blaOXA-161 and blaOXA-848. The primer design process used Primer3Plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi) and OligoAnalyzer (Integrated DNA Technologies, USA). The following parameters were considered in the process of designing the primers: amplicon size ranging from 70 bp to 210 bp, melting temperature (Tm) between 58°C and 62°C, GC content from 40% to 60%, lack of hairpins and self-dimers, and low probability of having cross-reaction as assessed by BLAST analysis using the NCBI Nucleotide database.
 |
Table 1 Designed Oligonucleotide Sequences Used in This Study and Thermal Cycling Conditions |
HRMA Protocol and Optimization
Polymerase chain reaction (PCR) and high-resolution melting analysis (HRMA) were performed using an ABI StepOne Plus Real-Time PCR System (Applied Biosystems, USA) with HOT FIREPol EvaGreen HRMA Mix (Solis BioDyne, Estonia). The PCR mixture (20 μL) was composed of 10 μL of 2X EvaGreen HRMA Mix, 0.5 μM of each primer, 2 μL of template DNA (50 ng), and water to volume. For sensitivity determination, nine-fold dilution series of the genomic DNA of reference strains were prepared, ranging from 1.5 × 107 to 1.5 × 10−2 CFU/mL equivalent, and analyzed in triplicate.
The thermal cycling parameters included preliminary denaturation at 95°C for 12 minutes to activate hot-start polymerase, followed by 40 cycles with denaturation at 95°C for 15 seconds, annealing at 58–60 °C (depending on primer pair; see Table 1) for 20 seconds, and extension at 72°C for 20 seconds. Fluorescence was measured after each extension. Then, high-resolution melting analysis was performed by cooling the reaction to 60°C for 1 minute and slowly increasing the temperature from 60°C to 95°C at 0.1°C per step, with fluorescence readings after each step.
The HRMA data were analyzed with ABI StepOne Plus Software version 3.2 (Applied Biosystems) and Precision Melt Analysis Software version 3.02 (Applied Biosystems). Melting curves and a difference plot were created to visualize the melt profiles of individual blaOXA amplicons. The Tm value was determined as the temperature at which 50% of the amplicon denatured, ie, the maximum point of the derivative melting curve.
Analytical Validation of the HRMA Assay
HRMA test’s analytical sensitivity was assessed using nine-fold serial dilutions of DNA isolated from reference strains with known genotypic characteristics, in triplicate. The limit of detection (LOD) was set to the minimum DNA amount that yields reproducible melting curves with peaks within ±0.5°C of the expected melting temperature (Tm). Analytical specificity was assessed on DNA from non-target bacteria and P. aeruginosa without target genes. The efficiency of PCR amplification was calculated from the slope of the standard curve using the equation Efficiency (%) = (10^1/slope – 1) × 100%. We used K. pneumoniae ATCC 1705 (KPC-positive) and C. freundii CRE 3123 (OXA-48-positive) as positive controls.
Multi-Locus Sequence Typing (MLST)
The MLST was performed following the PubMLST P. aeruginosa scheme (https://pubmlst.org/bigsdb?db=pubmlst_paeruginosa_seqdef).39 In brief, the internal regions of seven housekeeping genes (acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE) were amplified by conventional PCR using primers as reported previously.46 PCR amplicons were purified using the QIAquick PCR purification kit (Qiagen) and bidirectionally sequenced with the BigDye Terminator v3.1 cycle sequencing kit on an ABI 3500 genetic analyzer (Applied Biosystems). The allele numbers and sequence types (STs) were identified by analyzing sequences using the P. aeruginosa MLST database.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism software version 8.0 (GraphPad Software, Inc., California, USA). In case of categorical variables, Fisher’s exact test or Chi-square test was used to measure the correlation. Student’s t-test was used in case of continuous variables. A two-way ANOVA was used to compare normally distributed data. Statistical significance was considered as P < 0.05 for all tests.
A dendrogram was constructed using a neighbor-joining algorithm from a distance matrix based on allelic profiles. Tree generation and viewing were performed using PHYLIP (http://pubmlst.org/analysis) and the iTOL web-based interface (http://itol.embl.de/itol.cgi) within MEGA 6. Genes’ presence /absence and antibiotic resistance phenotypes were coded as binary. P. aeruginosa strains were isolated from clinical samples using the heatmap package in R. Principal component analysis was done by the FactoMineR package in R, which is an open-source statistical environment.47 The data analyses were performed using HRMA software (Thermo Fisher Scientific, version 2018, Version 3.0.2) and BioEdit 7.4 (CareData, Inc., USA). Normalized data and various charts were created.
Results
Isolation and Identification of Bacteria
In a 1-year study, 967 specimens were analyzed from hospitalized patients with suspected lower respiratory tract infection at four tertiary care hospitals in Hamadan, Iran. Out of these, 548 non-Enterobacteriaceae Gram-negative bacilli were found. The biochemical test confirmed that 100 isolates (18.2%) were Pseudomonas aeruginosa, which were considered for further studies. The other 448 isolates contained non-fermenting Gram-negative bacilli and other non-Pseudomonas species and were not used in the research.
Antimicrobial Susceptibility Patterns
Data on the susceptibility patterns of 100 P. aeruginosa isolates are presented in Table S1 and Figure 1. It should be stated that all isolates (100%) were susceptible to colistin, as this antibiotic is rarely resistant among P. aeruginosa isolates in this geographic area. At the same time, there are numerous resistance cases for the commonly used antipseudomonal agents. The maximum resistance rates were found for cefoxitin (92%), ciprofloxacin (89%), norfloxacin (84%), and ceftriaxone (82%).
 |
Figure 1 Individual isolates were showing hierarchical clustering of isolates and factors. Binary factors such as antibiotics resistance pattern indicating presence as red and absence as blue. Clustering is based on the Wald-like test (D2) and for binary data. Abbreviations: C, Colistin; PIP, Piperacillin; GEN, Gentamycin; AMK, Amikacin; CIP, Ciprofloxacin; CAZ, Ceftazidime; CTX, Ceftriaxone; IMI, Imipenem; MER, Meropenem; ERT, Ertapenem; FOX, Cefoxitin; CFX, Cefazolin; CPE, Cefepime; MDR, multiple drug resistance; XDR, extensively drug-resistant. |
Resistance to carbapenems was also high: imipenem (51%), meropenem (48%), and ertapenem (52%) were resistant among those isolates (Table S1). Moderate resistance rates were observed with ceftazidime (45%), cefepime (43%), and piperacillin (38%). There were low resistance rates to gentamicin (31%) and amikacin (24%), as aminoglycosides retained relatively preserved antimicrobial activity against P. aeruginosa in this environment.
Using the criteria shown in Figure S1 and Table S1, 84 isolates (84%) were determined to be multidrug-resistant (MDR), while 39 isolates (39%) were extensively drug-resistant (XDR). None of the isolates was pandrug-resistant (PDR), as all remained susceptible to colistin. Thus, the considerable prevalence of MDR and XDR P. aeruginosa isolates reflects a high level of antimicrobial resistance.
Carbapenemase Phenotypic Detection
The results obtained from the phenotypic detection of carbapenemase assays are given in Table S1. Out of the 100 P. aeruginosa isolates, 47 (47%) were found to be positive for the production of metallo-β-lactamase (MBL) because there was an expansion in the size of the inhibition zone by ≥7 mm in the presence of EDTA by using the imipenem-EDTA disk method. Also, 49 (49%) isolates were positive by the modified Hodge test (MHT), indicating carbapenemase production. The Carba NP test yielded positive results for 52 (52%) isolates, indicating hydrolysis of imipenem by carbapenemases.
According to Table S1, KPC production was detected in 52 (52%) isolates, as indicated by positive Carba NP test results and negative MBL screening. Positive results in all three phenotypic assays were observed in 21 (21%) isolates, suggesting the co-production of MBL and KPC. The phenotypic assays showed good correlation, and the Carba NP test showed the highest sensitivity for detecting carbapenemase production in this isolate set.
Biofilm Forming Capacity
Biofilm-forming capacity was assessed using the Congo Red Agar (CRA) method in all 100 isolates. According to Table S1, 15 (15%) isolates were found to produce high levels of biofilm, as evidenced by black-colored colonies with a crystalline appearance in CRA medium. Moderate biofilm-forming capacity (black colonies without a crystalline appearance) was observed in 36 (36%) isolates. 9 (9%) isolates produced low biofilm, as indicated by red colonies. 40 (40%) isolates lacked biofilm-forming capacity and produced smooth, pink colonies in CRA medium. Overall, 60% of P. aeruginosa isolates produced biofilms.
Validation of blaOXA Gene-Specific Primers
Two primer pairs specific for HRMA detection of blaOXA genes and four other primers from our previous work were validated through a full analytical validation process. Each primer pair generated only one peak during HRMA testing of the amplicon produced with target-positive control DNA, with no primer dimers or nonspecific amplification observed. The specificity of each primer pair was confirmed by negative amplification results for the non-target blaOXA genes and for other bacterial strains unrelated to the target bacteria.
The analytical sensitivity, reported in Table 2, was determined using nine-fold dilutions of purified genomic DNA from reference strains. Linear range of detection of Primer Pair 1 (blaOXA-145, blaOXA-161, blaOXA-224) reached 1.5×100 CFU/mL (about 50 genomic copies/reaction). Linear range of detection of Primer Pair 2 (blaOXA-539, blaOXA-675, blaOXA-848) reached 1.5×101 CFU/mL. Efficiency of PCR amplification performed with all primers was between 99.9% and 108.0%, with correlation coefficients (R2) more than 0.98 for all standard curves.
 |
Table 2 Analytical Sensitivity and Specificity of Real-Time PCR |
HRMA-Based Differentiation of blaOXA Genes
The HRMA assay accurately distinguished all six blaOXA target genes based on Tm and melt curve morphology. All amplicons produced distinctive melt peaks without overlap, allowing easy differentiation. The Tm and corresponding values are provided in Table 2. According to Figure 2A, the Tm values for Primer Set 1 are: blaOXA-145 is 81.70°C (81.70–81.75°C range), blaOXA-161 is 83.80°C (83.80–83.85°C range), and blaOXA-224 is 84.90°C (84.90–84.95°C range). The Tm differences between adjacent melt peaks ranged from 1.05°C to 2.10°C, exceeding the HRMA platform’s ±0.5 °C resolution. According to Figure 3A, the Tm values for Primer Set 2 are: blaOXA-539 is 82.10°C (82.11–82.16°C range), blaOXA-675 is 86.60°C (86.66–87.01°C range), and blaOXA-848 is 87.30°C (87.39–87.44°C range).
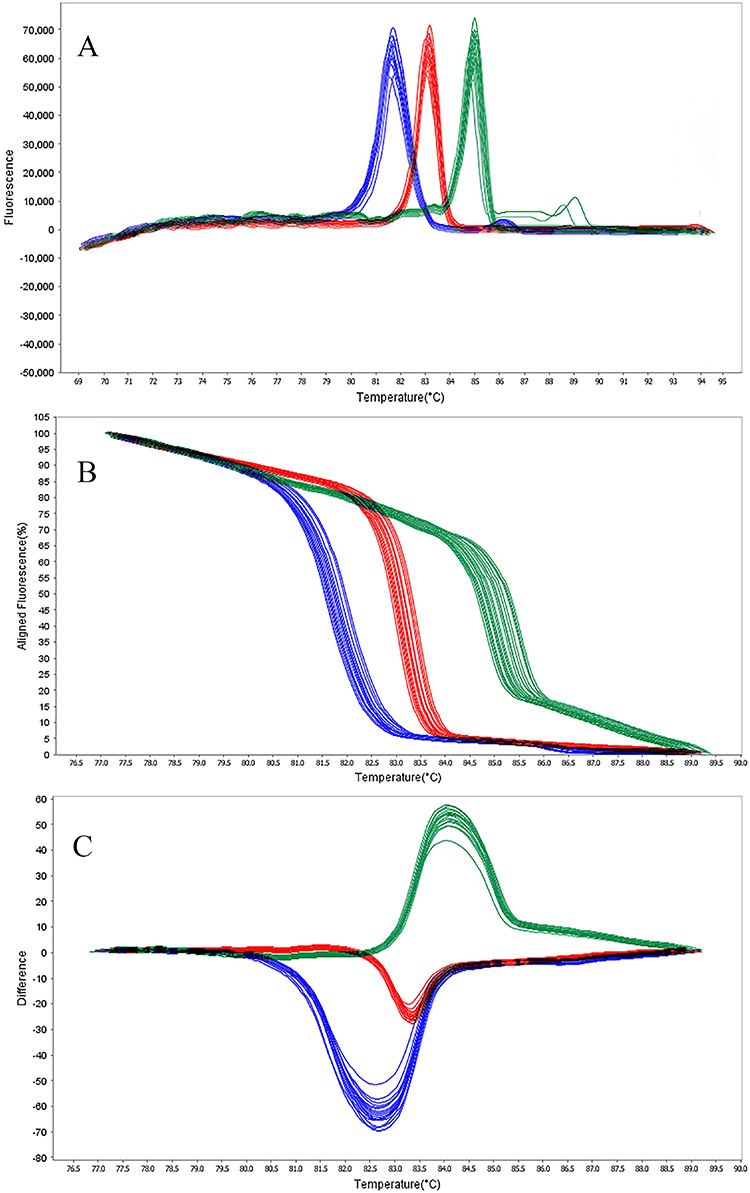 |
Figure 2 HRMA graphs corresponding to one high resolution melting analysis of a subset of NDM producing P. aeruginosa strain by blaOXA-145 (blue), blaOXA-161 (red), and blaOXA-224 (green) genes. DNA samples from all the dilutions involved in this study were prepared and amplified successfully using the EvaGreen dye-based method in the ABI instrument. Primers specific melting peaks (Tm) were obtained via HRMA analysis, allowing the differentiation of all investigated β-lactamase enzymes. Due to the positively saturating EvaGreen dye and the HRMA analysis, the resolution accuracy was ±0.1°C – 0.5°C. (A) Difference melt curve plot; (B) Aligned fluorescence plot (%); (C) Normalized difference plot. |
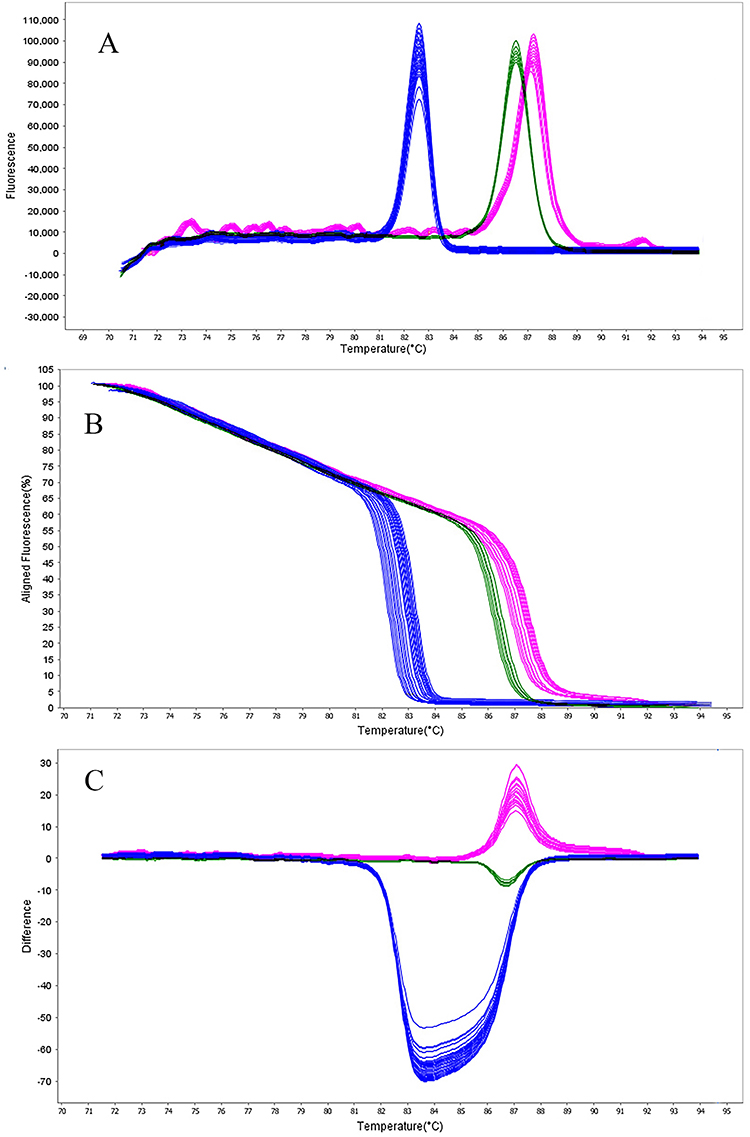 |
Figure 3 HRMA graphs corresponding to one high resolution melting analysis of a subset of NDM producing P. aeruginosa strain by blaOXA-539 (blue), blaOXA-675 (green), and blaOXA-848 (violet) genes. DNA samples from all the dilutions involved in this study were prepared and amplified successfully using the EvaGreen dye-based method in the ABI instrument. Primers specific melting peaks (Tm) were obtained via HRMA analysis, allowing the differentiation of all investigated β-lactamase enzymes. Due to the positively saturating EvaGreen dye and the HRMA analysis, the resolution accuracy was ±0.1°C – 0.5°C. (A) Difference melt curve plot; (B) Aligned fluorescence plot (%); (C) Normalized difference plot. |
In addition, Figures 2B, C, 3B and C show the results of normalized difference plots generated by the Precision Melt Analysis Software, which allowed more accurate discrimination between the different blaOXA genotypes. Each target gene produced a unique difference curve, which was easily recognizable against all other targets and negative control samples.
Prevalence of blaOXA Genes in P. Aeruginosa Isolates
The distribution of blaOXA genes among 100 Pseudomonas aeruginosa isolates was determined using HRMA and confirmed by conventional PCR followed by Sanger sequencing. Prevalence rates for all the strains are summarized in Table 2. As shown in Figure 2, out of the 47 MBL-producing isolates, the gene most commonly detected was blaOXA-539 present in 29 isolates (61.7%), blaOXA-161 – in 24 isolates (51.0%), blaOXA-224 – in 20 isolates (42.5%), blaOXA-145 – in 18 isolates (38.2%), blaOXA-848 – in 19 isolates (40.4%), and blaOXA-675 – in 7 isolates (14.8%) (Table S1).
As illustrated in Figure 3, among 52 KPC-producing isolates, the pattern of distribution was very similar: blaOXA-539 detected in 28 isolates (53.8%), blaOXA-161 – in 23 isolates (44.2%), blaOXA-224 – in 19 isolates (36.5%), blaOXA-145 – in 17 isolates (32.7%), blaOXA-848 – in 18 isolates (34.6%), and blaOXA-675 – in 6 isolates (11.5%). Importantly, 43 isolates (43%) had two or more blaOXA genes at the same time, of which the most common combination was the combination of blaOXA-539 and blaOXA-161 found in 18 isolates, while the second one was the combination of blaOXA-224 and blaOXA-145 in 12 isolates (Table S1).
Multi-Locus Sequence Typing Analysis
As shown in Figure 4, MLST typing successfully characterized all 100 isolates, demonstrating a highly diverse population structure represented by 95 distinct sequence types (STs). The most frequent STs included ST09 (12 isolates, 12%), ST235 (10 isolates, 10%), ST111 (9 isolates, 9%), and ST15 (8 isolates, 8%), which combined for 39% of the whole population. Moreover, the other 61 isolates carried 91 distinct rare STs, of which 57 (57%) were novel STs that had never been described in the PubMLST database.
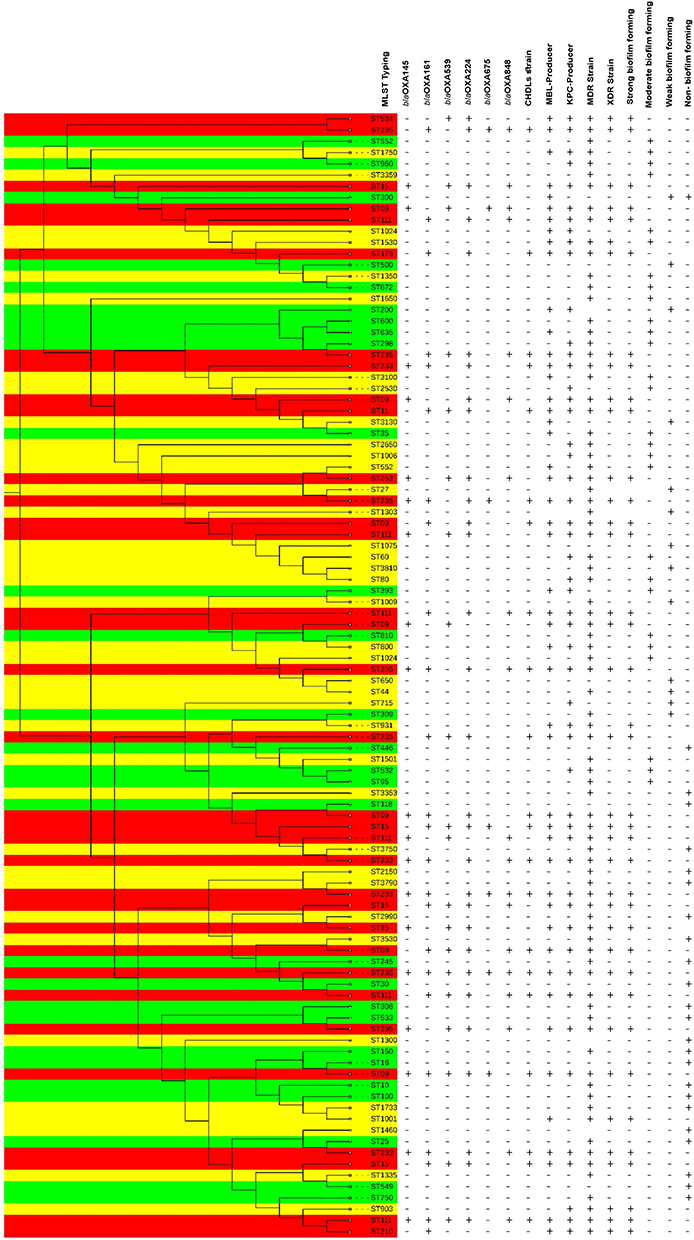 |
Figure 4 Dendrogram clustering of 100 lung infection isolates of P. aeruginosa based on sequence type (ST) profiles. Phylogenetic tree was constructed based on 500 base pairs of STs which were selected for multi locus sequence typing. The evolutionary history was inferred using the Neighbor-Joining method. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Red color: Dangerous STs; Green color: New STs in Iran; Yellow color: New STs in world. Abbreviations: MDR, Multiple drug resistance; XDR, Extensively drug-resistant; KPC, Klebsiella pneumoniae carbapenemase; MBL, Metallo-beta-lactamase; CHDLs, carbapenem-hydrolyzing class D OXA-type β-lactamases. |
Allelic profiling of seven MLST housekeeping genes revealed considerable allelic diversity. Thus, there were 15 different alleles in the gene acsA, 18 in the gene aroE, 14 in the gene guaA, 16 in the gene mutL, 20 in the gene nuoD, 12 in the gene ppsA, and 17 in the gene trpE. The gene nuoD proved to be the most polymorphic, with the largest number of allelic variants.
The phylogenetic analysis using the neighbor-joining method showed that ST235, ST111, and ST09 formed well-defined clusters, whereas ST15 showed greater dispersion within the phylogram. Novel 57 STs were distributed across the entire phylogenetic tree, indicating independent evolutionary origins rather than clonal expansion from a single ancestral strain.
Associations Between blaOXA Genes, Resistance Phenotypes, Biofilm Formation, and Sequence Types
The presence of blaOXA genes was significantly associated with increased resistance rates to carbapenems (imipenem: 78.3% vs 23.5%, meropenem: 75.0% vs 20.6%, ertapenem: 80.4% vs 26.5%), all p<0.001, when compared with negative controls. The number of co-harbored blaOXA genes was significantly positively correlated with the level of resistance to antimicrobials; isolates with the presence of three or more blaOXA genes had a significantly higher odds of having an XDR phenotype (odds ratio = 4.82, 95% CI: 2.14–10.86, p<0.001) (Figure 5).
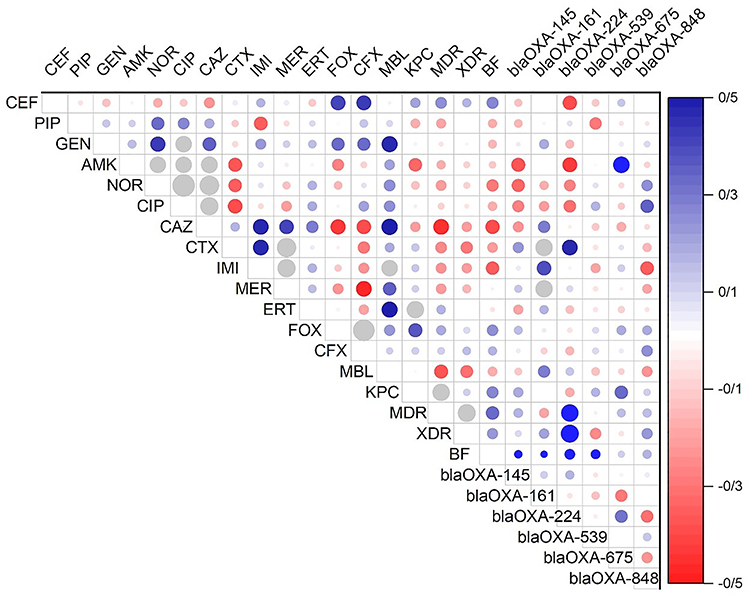 |
Figure 5 Correlation matrix of phenotypical (antibiotic resistance profile) and genotypical (OXA genes) characteristics with significant correlations (p< 0.05). There is no significant correlation between white spaces. The blue circles indicated a significant positive relationship, and the red indicated a significant negative relationship. The size and strength of color represent the numerical value of the Phi correlation coefficient. |
The production of biofilms was significantly associated with the carriage of blaOXA genes (p=0.003). Among the isolates classified as strong biofilm producers, 80.0% (12/15) had at least one blaOXA gene, compared with 55.6% (20/36) for moderate biofilm producers and 44.4% (4/9) for weak biofilm producers. The four major genes, including blaOXA-675, blaOXA-224, blaOXA-145, and blaOXA-539, accounted for more than 60% of the strong biofilm producers. On the other hand, there was no significant association between the carriage of blaOXA genes and resistance to colistin (p=0.742) and amikacin (p=0.186), which was consistent with their different resistance mechanisms to aminoglycosides and polymyxins.
The prevalent STs ST09, ST111, ST15, and ST235 showed statistically significant links with MDR/XDR phenotype (p<0.001), biofilm production (p=0.002), and CHDL possession (p<0.001). Interestingly, among all the prevalent STs, ST235 was particularly important: 90% of the ST235 strains (9/9 or 9/10, depending on the data) carried at least one blaOXA gene, and 80% of the strains (8/10) exhibited an XDR phenotype. Similarly, 83.3% of ST09 strains (10/12) and 77.8% of ST111 strains (7/9) were positive for blaOXA.
Discussion
The present study is the first comprehensive investigation of CHDLs in isolates of P. aeruginosa associated with pulmonary infections in western Iran, and, to the best of our knowledge, the first application of the HRMA technique for the detection of blaOXA genes in this organism in the Middle East. The data show a high rate of MDR (84%) and XDR (39%) phenotypes among P. aeruginosa isolates causing lung infections, and CHDLs play a significant role in carbapenem resistance. The HRMA method developed in the present study showed excellent analytical performance and allowed clear differentiation between six clinically important blaOXA genes solely from their melting curve patterns, without the use of fluorescent probes or additional steps in amplicon analysis.
Antimicrobial Resistance Patterns
The resistance pattern shown above confirms the growing global problem of Pseudomonas aeruginosa resistance. The high resistance levels seen for cefoxitin (92%) and strong resistance levels for ciprofloxacin (89%) and norfloxacin (84%) are similar to results from other Iranian centers and regional studies.48,49 Though resistance to carbapenems has raised concern, the results are consistent with neighboring countries’ statistics and can be attributed to selective pressure from carbapenem use.1,50
The antimicrobial resistance profiles observed in this study align with the escalating global challenge of P. aeruginosa resistance. The near-universal resistance to cefoxitin (92%) and high-level resistance to ciprofloxacin (89%) and norfloxacin (84%) are consistent with reports from other Iranian centers and regional surveillance data.48,49 The resistance rates to carbapenems, while concerning, are comparable to those documented in neighboring countries and reflect the selective pressure exerted by extensive carbapenem use in intensive care settings.1,50
Maintaining colistin susceptibility across all 100 isolates is important, as colistin resistance is rare in Pseudomonas aeruginosa compared with other Gram-negative nosocomial bacteria. However, colistin resistance has been reported in various geographic locations among P. aeruginosa strains, associated with the international high-risk clone ST235 and mutations in the pmrAB and phoPQ two-component regulatory systems.51 Thus, constant monitoring of colistin susceptibility patterns is important to maintain this last resource of antibiotics.
In this study, 47% and 53% of the isolates demonstrated MBL and KPC producer activity, respectively, results similar to those of Gill et al52 Moreover, Alam et al53 and Sisay et al54 have found that MBL and KPC producer P. aeruginosa are more frequent in wound isolates than in other isolates of P. aeruginosa. In turn, Muddassir et al55 reported that MBL-producing P. aeruginosa are more frequent among wound isolates. Differences in antimicrobial stewardship strategies, infection control systems, and the presence of selective pressure from carbapenems in healthcare institutions can explain such differences. The finding that 39% of isolates were XDR P. aeruginosa underscores the urgent need to improve antimicrobial stewardship and infection prevention programs in Iran.
Phenotypic Detection of Carbapenemases
The phenotypic tests used for carbapenemase detection in this study showed good performance, with the Carba NP test demonstrating the highest sensitivity (52% positive rate). However, the fact that 21% of the isolates produced both MBL and KPC enzymes indicates the intricacy of carbapenem resistance mechanisms in modern Pseudomonas aeruginosa isolates. The phenomenon of the production of two or more carbapenemases is gaining popularity and represents a major problem for both diagnostics and treatment.56
While the modified Hodge test is widely applied due to its ease and low cost, its sensitivity was found to be slightly lower than that of the Carba NP test in this study. This result is consistent with previous reports of the inconsistent sensitivity of the MHT test in detecting CHDL enzymes, especially OXA types with low carbapenemase activity.57 The Carba NP test, which directly detects carbapenemase activity by transforming the chromogenic substrate into carbapenem, is more sensitive and specific but requires specialized reagents and equipment.
Biofilm Formation and Virulence
The ability of 60% of the isolates to form biofilms, with 15% belonging to strong biofilm producers, has important clinical ramifications. P. aeruginosa biofilm infections are known to be particularly refractory to treatment by virtue of their enhanced resistance to antimicrobial agents and their immune-evasion abilities.58 The presence of a statistically significant association between the possession of blaOXA genes and biofilm production (p = 0.003) may indicate a relationship between resistance determinants and the virulence properties of P. aeruginosa.
Such a relationship could exist due to the co-expression of resistance and biofilm genes through common regulatory pathways, such as quorum-sensing systems (las, rhl, and pqs), which control both biofilm formation and the expression of resistance determinants.59,60 On the other hand, the acquisition of resistance genes and the increased biofilm-forming ability can be considered separate adaptive traits that emerge under the pressure exerted on the organism in the hospital environment and are therefore acquired independently. It is possible that specific resistance genes, such as blaOXA-675, blaOXA-224, blaOXA-145, and blaOXA-539, are linked to biofilm formation and biofilm-associated persistence.
Development and Performance of the HRMA Assay
The HRMA technique proposed in this study represents a considerable improvement in molecular detection of CHDLs in P. aeruginosa. The successful differentiation of all six blaOXA target genes using different melting temperatures for their PCR products, with a resolution of ±0.1–0.5 °C, demonstrates the feasibility of this technique for multiplex detection of resistance genes. There are significant advantages of the HRMA technique over current methods for molecular and phenotypic detection of CHDLs.
Since HRMA is a closed-tube one-step assay, there is no post-amplification manipulation, which prevents the risk of amplicon carry-over contamination.61 This aspect is especially relevant in a clinical setting, where many samples need to be tested simultaneously. Secondly, the non-use of probes specific to sequences of interest greatly reduces the cost of the reagents needed for the test. The use of a generic, saturated DNA-binding dye, such as EvaGreen, makes HRMA affordable because it can detect any PCR product regardless of its sequence.
Firstly, the turnaround time of about 2 hours from DNA extraction to result reporting represents a significant improvement over the 48–72 hours required for PCR and sequencing.62 Second, it allows for early diagnosis and, hence, the appropriate choice of antimicrobial treatment, contributing to the achievement of the goals of an antimicrobial stewardship program. Third, the HRMA assay has demonstrated very good analytical sensitivity with a detection limit of 1.5×100–1.5×101 CFU/mL. It is equivalent to, or superior to, real-time PCR with probes. High PCR efficiency (99.9–108.0%) and a high correlation coefficient (R2 > 0.98) confirm high amplification efficiency across the entire measurement range.
The precision of the Tm measurement in the current research, with intra-run and inter-run CV values less than 0.3%, is consistent with previous studies, which validate the applicability of the HRMA technique for microbial typing and screening for antibiotic resistance genes.63 Moreover, normalization of difference plots has improved visualization of genotype distinction, providing a more convenient visual output format.
Prevalence and Distribution of blaOXA Genes
The prevalence of blaOXA genes reported in this work contributes significantly to understanding the molecular epidemiology of CHDLs in P. aeruginosa from this region. The predominant occurrence of blaOXA-539 (61.7% among MBL producers) and blaOXA-161 (51.0%) is consistent with the findings of Fraile-Ribot et al, where blaOXA-224 and blaOXA-539 were among the prevalent genes in carbapenem-resistant isolates.10 On the other hand, the high prevalence of OXA-2 family has been previously reported in Spain.32 The blaOXA-539 gene encodes an OXA-2 variant with the amino acid duplication D149 (OXA-539) associated with increased resistance to ceftazidime and carbapenems in sequence type 235 P. aeruginosa57 and demonstrates one of the evolutionary adaptations of the OXA-2 β-lactamases.
The high prevalence of blaOXA-224 (42.5%) and blaOXA-145 (38.2%) is remarkable given their relatively recent identification. The enzymes encoded by the former and latter genes are representatives of the OXA-2 and OXA-10 enzyme families, respectively, with an expanded substrate range that includes carbapenems and extended-spectrum cephalosporins.64 The clinical importance of the above-mentioned enzymes is underscored by their contribution to the XDR phenotype and their ability to confer resistance to novel β-lactam/β-lactamase inhibitor combinations, such as ceftazidime-avibactam.65
The low prevalence of blaOXA-675 (14.8%) in our isolate collection is consistent with its relatively narrow geographical distribution, which is predominantly reported in Middle Eastern and South Asian countries.66 The blaOXA-848 gene, found in 40.4% of MBL producers, is a new member of the OXA-48-like family, recently identified in P. aeruginosa isolates from this geographical area.67 The presence of several blaOXA genes in 43% of our isolates implies horizontal gene transfer and the selection of increasingly resistant isolates.
Molecular Epidemiology and Clonal Structure
From the MLST results, it appears that there was genetic diversity among the P. aeruginosa strains, with 95 different sequence types (STs) among the 100 isolates. The dominance of ST09, ST235, ST111, and ST15 strains corresponds to their status as international high-risk clones characterized by carbapenem resistance and increased virulence.68 The last seven strains belonged to ST111 and ST235 as well. There are numerous cases of lung infections associated with these strains, and this ST lineage is highly resistant to antimicrobial drugs. ST235 also dominated lung infection isolates. According to de Sales et al, ST235 appears to be a dangerous strain in pulmonary infections.39
It is noteworthy that 57% of all isolates had unique or single STs. Thus, there is substantial genetic diversity among the strains and little evidence of clonal spread in the present case. This is a significant difference from cases reported in European countries, where several dominant clones accounted for most carbapenem-resistant P. aeruginosa infections.64 There may be selective pressure and infection control measures in Iranian medical institutions, or unknown P. aeruginosa reservoirs in the community.
Among the MDR and XDR strains, ST09, ST235, and ST111 also had a high rate. Additionally, those STs were found in isolates expressing blaOXA. It is interesting to note that various sequence types were present in the new STs from western Iran. Similar to our findings, Annear et al43 have observed several STs and variations in resistant and virulent isolates.; they also identified ST09, and ST235 mostly found in hyper pathogenic isolates. As reported by Irum et al, the high prevalence of those STs was not seen in our study.30 This difference could be due to the use of different clinical samples by Irum et al Our study only analyzed lung infection samples to assess novel STs.
According to our investigation into lung infections, biofilm-forming isolates were significantly enriched with potentially harmful STs, including ST09, ST15, and ST235. These findings support what Guzvinec et al42 observed for a group of P. aeruginosa isolates, namely that ST253 was more prevalent. Consistent with our present results, previous research has also stated that ST111, ST235, and ST233 are the most common STs among biofilm-forming P. aeruginosa isolates. This research was conducted in Brazil.39
Therefore, the strong relationships found between the dominant strains (ST09, ST111, ST15, ST235) and MDR/XDR phenotypes, biofilm production, and CHDL possession indicate the medical importance of these clonal complexes. In particular, ST235 strains had a 90% incidence of blaOXA and an 80% incidence of the XDR phenotype, thus confirming their classification as highly dangerous clones in this clinical setting. Furthermore, the fact that the new STs possessed the blaOXA genes and the MDR phenotype less often suggests that the development of resistance factors is currently underway among the local population of P. aeruginosa, with the dominant clones serving as carriers of these resistance factors.
Clinical Implications and Future Directions
The results of this research have several important implications for clinical practice and public health policy. A relatively high prevalence of MDR and XDR P. aeruginosa strains detected in respiratory tract infections underscores the pressing need for effective antimicrobial stewardship strategies and increased efforts in infection control and diagnostics of antibiotic-resistant pathogens in Iranian hospitals. The fact that there are very few treatment options for XDR infections (colistin and possibly a combination of antibiotics with adverse effects) emphasizes the necessity of prudent use of currently available antibiotics to keep them effective for longer.
The high-resolution melting analysis technique proposed in this research is a simple and cost-effective diagnostic tool for the identification of carbapenem-hydrolyzing beta-lactamases (CHDL), which can be easily implemented in routine practice in any clinical laboratory facility capable of conducting real-time PCR. Therefore, incorporation of this method for the diagnosis of CHDL into the daily practice of clinical microbiologists could be useful for proper therapy, infection control, and outbreak investigation, and could help obtain important data for local and regional antibiotic resistance surveillance programs.
Further research should focus on prospective validation of the effectiveness and cost-effectiveness of the new technique compared with currently used diagnostics, as well as its impact on patient outcomes and antimicrobial stewardship performance indicators. Furthermore, applying whole-genome sequencing techniques to the novel STs identified in this study would provide an opportunity to reveal additional information about their origins, gene pools and virulence characteristics. Finally, conducting surveillance studies on a larger scale would improve knowledge of the molecular epidemiology of CHDL-producing P. aeruginosa in Iran and the Middle East region.
Study Limitations
There are a few limitations to consider. First of all, the investigation was conducted in a limited geographic area for only one year, limiting the applicability of the results to other cases. Second, the use of CRA to detect biofilms is a qualitative method, whereas further research should employ quantitative methods, such as crystal violet microtiter plate assays. Third, the accuracy of the HRMA assay is determined by its validation against six blaOXA genes; even though those six are the most relevant to the region in question, there might be rare or new variants that will be undetectable Finally, the clinical outcomes of infection with CHDL-producing strains of P. aeruginosa were not studied.
In summary, the results indicate a high prevalence of CHDL-producing multidrug-resistant (MDR) and extensively drug-resistant (XDR) P. aeruginosa strains in pulmonary infections in western Iran. The most common blaOXA genes were blaOXA-539, blaOXA-161, and blaOXA-224. An HRMA assay was developed and proved to be a fast, precise, and relatively inexpensive tool for detecting and distinguishing among six clinically relevant blaOXA genes. The high prevalence of novel sequence types indicates significant genetic variability in P. aeruginosa in the region.
Abbreviations
CHDLs, Carbapenem-hydrolyzing class D β-lactamases; HRMA, High-Resolution Melting Curve Analysis; NDM, New Delhi Metallo-β-lactamase; OXA, oxacillinase; CLSI, Clinical & Laboratory Standards Institute; MHT, modified Hodge test.
Ethical Approval Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of Hamadan University of Medical Sciences (IR.UMSHA.REC.1398.1007).
Acknowledgments
This study was approved by the Ethics Committee of Hamadan University of Medical Sciences (Code No: IR.UMSHA.REC.1398.1007).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors of this article are grateful to Hamadan University of Medical Sciences for their financial support in conducting the research (Grant Number: 9812209730).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tchakal-Mesbahi A, Metref M, Singh VK, Almpani M, Rahme LG. Characterization of antibiotic resistance profiles in Pseudomonas aeruginosa isolates from burn patients. Burns. 2021;47(8):1833–21. doi:10.1016/j.burns.2021.03.005
2. Wood SJ, Kuzel TM, Shafikhani SH. Pseudomonas aeruginosa: infections, animal modeling, and therapeutics. Cells. 2023;12(1):199. doi:10.3390/cells12010199
3. Bozadzhieva L, Miteva D, Ilarionova L, Teneva T, Zdravkov B, Petrova G. Antibiotic use among children requiring respiratory support in Intensive Care Unit (ICU) from Sofia, Bulgaria: a single-center retrospective experience. Antibiotics. 2026;15(2):225. doi:10.3390/antibiotics15020225
4. Dehbashi S, Tahmasebi H, Zeyni B, Arabestani MR. Regulation of virulence and β-lactamase gene expression in Staphylococcus aureus isolates: cooperation of two-component systems in bloodstream superbugs. BMC Microbiol. 2021;21(1):192. doi:10.1186/s12866-021-02257-4
5. Vatansever C, Menekse S, Dogan O, et al. Co-existence of OXA-48 and NDM-1 in colistin resistant Pseudomonas aeruginosa ST235. Emerging Microbes Infect. 2020;9(1):152–154. doi:10.1080/22221751.2020.1713025
6. Tahmasebi H, Khazaei M, Arabestani MR. Role of antimicrobial peptide-based biomaterials in respiratory tract infections control. BMC Microbiol. 2026;26(1):552. doi:10.1186/s12866-026-05110-8
7. Tahmasebi H, Arabestani MR. Antimicrobial peptides against antimicrobial-resistant bacteria: focus on machine learning. Infect Drug Resist. 2026;19(602699). doi:10.2147/IDR.S602699
8. Tahmasebi H, Dehbashi S, Alikhani MY, Porbaran M, Arabestani MR. Prevalence and molecular typing of Metallo-β-lactamase-producing Pseudomonas aeruginosa with adhesion factors: a descriptive analysis of burn wounds isolates from Iran. Gene Rep. 2020;21:100853. doi:10.1016/j.genrep.2020.100853
9. Pérez A, Gato E, Pérez-Llarena J, et al. High incidence of MDR and XDR Pseudomonas aeruginosa isolates obtained from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J Antimicrob Chemother. 2019;74(5):1244–1252. doi:10.1093/jac/dkz030
10. Mentasti M, Prime K, Sands K, Khan S, Wootton M. Rapid detection of IMP, NDM, VIM, KPC and OXA-48-like carbapenemases from Enterobacteriales and Gram-negative non-fermenter bacteria by real-time PCR and melt-curve analysis. Eur J Clin Microbiol Infect Dis. 2019;38(11):2029–2036. doi:10.1007/s10096-019-03637-5
11. Zalas-Wiecek P, Prazynska M, Pojnar L, Palka A, Zabicka D. Ceftazidime/Avibactam and other commonly used antibiotics activity against enterobacterales and Pseudomonas aeruginosa isolated in Poland in 2015-2019. Infect Drug Resist. 2022;15:1289–1304. doi:10.2147/IDR.S344165
12. Grisold AJ, Luxner J, Bedenić B, et al. Diversity of oxacillinases and sequence types in carbapenem-resistant acinetobacter baumannii from Austria. Int J Environ Res Public Health. 2021;18(4):2171. doi:10.3390/ijerph18042171
13. Wachino JI, Jin W, Kimura K, Kurosaki H, Sato A, Arakawa Y. Sulfamoyl heteroarylcarboxylic acids as promising metallo-beta-lactamase inhibitors for controlling bacterial carbapenem resistance. mBio. 2020;11(2). doi:10.1128/mBio.03144-19
14. Husna A, Rahman MM, Badruzzaman ATM, et al. Extended-spectrum β-lactamases (ESBL): challenges and opportunities. Biomedicines. 2023;11(11):2937. doi:10.3390/biomedicines11112937
15. Lawrence J, O’Hare D, van Batenburg-Sherwood J, Sutton M, Holmes A, Rawson TM. Innovative approaches in phenotypic beta-lactamase detection for personalised infection management. Nat Commun. 2024;15(1):9070. doi:10.1038/s41467-024-53192-7
16. Kao K, Alocilja EC. A review of the diagnostic approaches for the detection of antimicrobial resistance, including the role of biosensors in detecting carbapenem resistance genes. Genes. 2025;16(7):794. doi:10.3390/genes16070794
17. Kaprou GD, Bergšpica I, Alexa EA, Alvarez-Ordóñez A, Prieto M. Rapid methods for antimicrobial resistance diagnostics. Antibiotics. 2021;10(2):209. doi:10.3390/antibiotics10020209
18. Fraile-Ribot PA, Mulet X, Cabot G, et al. In vivo emergence of resistance to novel cephalosporin-β-lactamase inhibitor combinations through the duplication of amino acid D149 from OXA-2 β-Lactamase (OXA-539) in sequence type 235 Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2017;61(9). doi:10.1128/aac.01117-17
19. Spinella K, Costa LD, Rocca DL, et al. High-Resolution Melting (HRM) analysis for screening edited lines: a case study in vitis spp. Int J Plant Biol. 2025;16(4):126. doi:10.3390/ijpb16040126
20. Aldea ACC, Diguṭă FC, Presacan O, Voaideṣ C, Toma RCC, Matei F. Detecting antibiotic resistance: classical, molecular, advanced bioengineering, and AI-enhanced approaches. Front Microbiol. 2025;16. doi:10.3389/fmicb.2025.1673343
21. van der Vlugt SEJ, Fros JJ, Martins Dos Santos VAP, Albada B, Asin-Garcia E. High-resolution melting analysis for rapid and cost-effective detection of unnatural base pairs. BioDesign Research. 2026;8(2):100090. doi:10.1016/j.bidere.2026.100090
22. Dehbashi S, Tahmasebi H, Alikhani MY, Keramat F, Arabestani MR. Distribution of class b and class a β-lactamases in clinical strains of Pseudomonas aeruginosa: comparison of phenotypic methods and high-resolution melting analysis (HRMA) assay. Infect Drug Resist. 2020;13:2037–2052. doi:10.2147/IDR.S255292
23. Phongroop K, Rattanasrisomporn J, Tangtrongsup S, Rungsipipat A, Piewbang C, Techangamsuwan S. High-resolution melting analysis for simultaneous detection and discrimination between wild-type and vaccine strains of feline calicivirus. Vet Q. 2023;43(1):1–12. doi:10.1080/01652176.2023.2272188
24. Andini N, Wang B, Athamanolap P, et al. Microbial typing by machine learned DNA melt signatures. Sci Rep. 2017;7(1):42097. doi:10.1038/srep42097
25. Tahmasebi H, Dehbashi S, Arabestani MR. New approach to identify colistin-resistant Pseudomonas aeruginosa by high-resolution melting curve analysis assay. Lett Appl Microbiol. 2020;70(4):290–299. doi:10.1111/lam.13270
26. Osadare IE, Abdilahi A, Reinicke M, et al. Multiplex real-time polymerase chain reaction and recombinase polymerase amplification: methods for quick and cost-effective detection of Vancomycin-Resistant Enterococci (VRE). Antibiotics. 2025;14(3):295. doi:10.3390/antibiotics14030295
27. Smiljanic M, Kaase M, Ahmad-Nejad P, Ghebremedhin B. Comparison of in-house and commercial real time-PCR based carbapenemase gene detection methods in Enterobacteriaceae and non-fermenting gram-negative bacterial isolates. Ann Clin Microbiol Antimicrob. 2017;16(1):48. doi:10.1186/s12941-017-0223-z
28. Ashrafi R, Bruneaux M, Sundberg L-R, Pulkkinen K, Ketola T. Application of high resolution melting assay (HRM) to study temperature-dependent intraspecific competition in a pathogenic bacterium. Sci Rep. 2017;7(1):980. doi:10.1038/s41598-017-01074-y
29. Espy MJ, Uhl JR, Sloan LM, et al. Real-time PCR in clinical microbiology: applications for routine laboratory testing. Clin Microbiol Rev. 2006;19(1):165–256. doi:10.1128/cmr.19.1.165-256.2006
30. Irum S, Naz K, Ullah N, et al. Antimicrobial resistance and genomic characterization of six new sequence types in multidrug-resistant Pseudomonas aeruginosa clinical isolates from Pakistan. Antibiotics. 2021;10(11):1386. doi:10.3390/antibiotics10111386
31. Shen C, Zeng J, Zheng D, et al. Molecular epidemiology and genomic dynamics of Pseudomonas aeruginosa isolates causing relapse infections. Microbiol Spectr. 2023;11(5):e0531222. doi:10.1128/spectrum.05312-22
32. Hocquet D, Colomb M, Dehecq B, et al. Ceftazidime-hydrolysing β-lactamase OXA-145 with impaired hydrolysis of penicillins in Pseudomonas aeruginosa. J Antimicrob Chemother. 2011;66(8):1745–1750. doi:10.1093/jac/dkr187
33. Kabic J, Fortunato G, Vaz-Moreira I, et al. Dissemination of metallo-β-lactamase-producing Pseudomonas aeruginosa in serbian hospital settings: expansion of ST235 and ST654 clones. Int J Mol Sci. 2023;24(2):1519. doi:10.3390/ijms24021519
34. Valzano F, Bella GL, Lopizzo T, et al. Resistance to ceftazidime-avibactam and other new β-lactams in Pseudomonas aeruginosa clinical isolates: a multi-center surveillance study. Microbiol Spectr. 2024;12(8):e0426623. doi:10.1128/spectrum.04266-23
35. Salleh MZ, Zuraina NMNN, Deris ZZ, Mohamed Z. Current trends in the epidemiology of multidrug-resistant and beta-lactamase-producing Pseudomonas aeruginosa in Asia and Africa: a systematic review and meta-analysis. PeerJ. 2025;13:e18986. doi:10.7717/peerj.18986
36. Kim N, Ko SY, Park SY, et al. Clonal distribution and its association with the carbapenem resistance mechanisms of carbapenem-non-susceptible Pseudomonas aeruginosa isolates From Korean Hospitals. Ann Lab Med. 2024;44(5):410–417. doi:10.3343/alm.2023.0369
37. Park Y, Koo SH. Epidemiology, molecular characteristics, and virulence factors of carbapenem-resistant Pseudomonas aeruginosa isolated from patients with urinary tract infections. Infect Drug Resist. 2022;15:141–151. doi:10.2147/idr.S346313
38. Rai S, Dash D, Agarwal N. Introducing the new face of CLSI M100 in 2023: an explanatory review. Indian J Med Microbiol. 2023;46:100432. doi:10.1016/j.ijmmb.2023.100432
39. de Sales RO, Migliorini LB, Puga R, Kocsis B, Severino P. A core genome multilocus sequence typing scheme for Pseudomonas aeruginosa. Front Microbiol. 2020;11:1049. doi:10.3389/fmicb.2020.01049
40. Gajdács M, Baráth Z, Kárpáti K, et al. No correlation between biofilm formation, virulence factors, and antibiotic resistance in Pseudomonas aeruginosa: results from a laboratory-based in vitro study. Antibiotics. 2021;10(9):1134. doi:10.3390/antibiotics10091134
41. Porbaran M, Tahmasebi H, Arabestani M. A comprehensive study of the relationship between the production of β -lactamase enzymes and iron/siderophore uptake regulatory genes in clinical isolates of acinetobacter baumannii. Int J Microbiol. 2021;2021:1–8. doi:10.1155/2021/5565537
42. Guzvinec M, Izdebski R, Butic I, et al. Sequence types 235, 111, and 132 predominate among multidrug-resistant Pseudomonas aeruginosa clinical isolates in Croatia. Antimicrob Agents Chemother. 2014;58(10):6277–6283. doi:10.1128/aac.03116-14
43. Annear D, Black J, Govender S. Multi-locus sequence typing of carbapenem resistant Pseudomonas aeruginosa isolates from patients presenting at port elizabeth hospitals, South Africa. Afr J Infect Dis. 2017;11(2):68–74. doi:10.21010/ajid.v11i2.9
44. Tahmasebi H, Dehbashi S, Arabestani MR. High resolution melting curve analysis method for detecting of carbapenemases producing Pseudomonas aeruginosa. J Krishna Instit of Med Sci Univ. 2018;7(4):70–77.
45. Sezadehghani A, Dehbashi S, Tahmasebi H, Arabestani MR. Detection of bla OXA-145, bla OXA-224, bla OXA-539, and bla OXA-675 genes and carbapenem-hydrolyzing class D β -lactamases (CHDLs) in clinical isolates of Pseudomonas aeruginosa collected from West of Iran, Hamadan. Int J Microbiol. 2022;2022:1–10. doi:10.1155/2022/3841161
46. Curran B, Jonas D, Grundmann H, Pitt T, Dowson CG. Development of a multi-locus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J Clin Microbiol. 2004;42(12):5644–5649. doi:10.1128/jcm.42.12.5644-5649.2004
47. Tahmasebi H, Dehbashi S, Nasaj M, Arabestani MR. Molecular epidemiology and collaboration of siderophore-based iron acquisition with surface adhesion in hypervirulent Pseudomonas aeruginosa isolates from wound infections. Sci Rep. 2022;12(1):7791. doi:10.1038/s41598-022-11984-1
48. Potron A, Poirel L, Nordmann P. Emerging broad-spectrum resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: mechanisms and epidemiology. Int J Antimicrob Agents. 2015;45(6):568–585. doi:10.1016/j.ijantimicag.2015.03.001
49. Xie X, Liu Z, Huang J, et al. Molecular epidemiology and carbapenem resistance mechanisms of Pseudomonas aeruginosa isolated from a hospital in Fujian, China. Front Microbiol. 2024;15:1431154. doi:10.3389/fmicb.2024.1431154
50. Hrabák J, Walková R, Studentová V, Chudácková E, Bergerová T. Carbapenemase activity detection by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J Clin Microbiol. 2011;49(9):3222–3227. doi:10.1128/jcm.00984-11
51. Oliver A, Mulet X, López-Causapé C, Juan C. The increasing threat of Pseudomonas aeruginosa high-risk clones. Drug Resist Updat. 2015;21-22:41–59. doi:10.1016/j.drup.2015.08.002
52. Gill CM, Lasko MJ, Asempa TE, Nicolau DP. Evaluation of the EDTA-modified carbapenem inactivation method for detecting metallo-beta-lactamase-producing Pseudomonas aeruginosa. J Clin Microbiol. 2020;58(6). doi:10.1128/JCM.02015-19
53. Alam MM, Islam MN, Hawlader MDH, et al. Prevalence of multidrug resistance bacterial isolates from infected wound patients in Dhaka, Bangladesh: a cross-sectional study. Int J Surg Open. 2021;28:56–62. doi:10.1016/j.ijso.2020.12.010
54. Sisay M, Worku T, Edessa D. Microbial epidemiology and antimicrobial resistance patterns of wound infection in Ethiopia: a meta-analysis of laboratory-based cross-sectional studies. BMC Pharmacol Toxicol. 2019;20(1):35. doi:10.1186/s40360-019-0315-9
55. Muddassir M, Munir S, Raza A, et al. Epidemiology and high incidence of metallo-beta-lactamase and AmpC-beta-lactamases in nosocomial Pseudomonas aeruginosa. Iran J Basic Med Sci. 2021;24(10):1373–1379. doi:10.22038/IJBMS.2021.57293.12748
56. Meletis G. Carbapenem resistance: overview of the problem and future perspectives. Ther Adv Infect Dis. 2016;3(1):15–21. doi:10.1177/2049936115621709
57. Tamma PD, Goodman KE, Harris AD, et al. Comparing the outcomes of patients with carbapenemase-producing and non-carbapenemase-producing carbapenem-resistant enterobacteriaceae bacteremia. Clin Infect Dis. 2017;64(3):257–264. doi:10.1093/cid/ciw741
58. Codjoe FS, Donkor ES. Carbapenem resistance: a review. Med Sci. 2017;6(1). doi:10.3390/medsci6010001
59. Poole K. Pseudomonas aeruginosa: resistance to the max. Front Microbiol. 2011;2:65. doi:10.3389/fmicb.2011.00065
60. Dehbashi S, Tahmasebi H, Alikhani MY, Vidal JE, Seifalian A, Arabestani MR. The healing effect of Pseudomonas Quinolone Signal (PQS) with co-infection of Staphylococcus aureus and Pseudomonas aeruginosa: a preclinical animal co-infection model. J Infect Public Health. 2024;17(2):329–338. doi:10.1016/j.jiph.2023.12.016
61. Lister PD, Wolter DJ, Hanson ND. Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin Microbiol Rev. 2009;22(4):582–610. doi:10.1128/cmr.00040-09
62. Livermore DM. Current epidemiology and growing resistance of gram-negative pathogens. Korean J Intern Med. 2012;27(2):128–142. doi:10.3904/kjim.2012.27.2.128
63. Vasala A, Hytönen VP, Laitinen OH. Modern tools for rapid diagnostics of antimicrobial resistance. Front Cell Infect Microbiol. 2020;10:308. doi:10.3389/fcimb.2020.00308
64. Nordmann P, Dortet L, Poirel L. Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol Med. 2012;18(5):263–272. doi:10.1016/j.molmed.2012.03.003
65. Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046–5054. doi:10.1128/aac.00774-09
66. Bush K, Bradford PA. Epidemiology of β-Lactamase-Producing Pathogens. Clin Microbiol Rev. 2020;33(2). doi:10.1128/cmr.00047-19
67. Walsh TR, Weeks J, Livermore DM, Toleman MA. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect Dis. 2011;11(5):355–362. doi:10.1016/s1473-3099(11)70059-7
68. Kumarasamy KK, Toleman MA, Walsh TR, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010;10(9):597–602. doi:10.1016/s1473-3099(10)70143-2
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.