Back to Journals » International Journal of Nanomedicine » Volume 18
Harnessing Stem Cell-Derived Extracellular Vesicles for the Regeneration of Degenerative Bone Conditions
Received 7 June 2023
Accepted for publication 23 September 2023
Published 29 September 2023 Volume 2023:18 Pages 5561—5578
DOI https://doi.org/10.2147/IJN.S424731
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Zhiwei Jia,1 Shunxin Zhang,2 Wei Li3
1Department of Orthopedics, Dongzhimen Hospital, Beijing University of Chinese Medicine, Beijing, 101100, People’s Republic of China; 2Department of Ultrasound, 2nd Medical Center of PLA General Hospital, Beijing, 100853, People’s Republic of China; 3Department of Sports Medicine, Fourth Medical Center of PLA General Hospital, Beijing, 100048, People’s Republic of China
Correspondence: Wei Li, Department of Sports Medicine, Fourth Medical Center of PLA General Hospital, Beijing, 100048, People’s Republic of China, Email [email protected] Shunxin Zhang, Department of Ultrasound, 2nd Medical Center of PLA General Hospital, Beijing, 100853, People’s Republic of China, Email [email protected]
Abstract: Degenerative bone disorders such as intervertebral disc degeneration (IVDD), osteoarthritis (OA), and osteoporosis (OP) pose significant health challenges for aging populations and lack effective treatment options. The field of regenerative medicine holds promise in addressing these disorders, with a focus on utilizing extracellular vesicles (EVs) derived from stem cells as an innovative therapeutic approach. EVs have shown great potential in stimulating biological responses, making them an attractive candidate for rejuvenating degenerative bone disorders. However, a comprehensive review summarizing the current state of this field and providing a clear assessment of EV-based therapies in degenerative bone disorders is currently deficient. In this review, we aim to fill the existing gap by outlining the current knowledge on the role of EVs derived from different types of stem cells, such as mesenchymal stem cells, embryonic stem cells, and induced pluripotent stem cells, in bone regeneration. Furthermore, we discuss the therapeutic potential of EV-based treatments for IVDD, OA, and OP. By substantiating the use of stem cell-derived EVs, we highlight their promising potential as a cell-free strategy to improve degenerative bone disorders.
Keywords: nanomedicine, extracellular vesicles, stem cells, regeneration, therapy
Introduction
Degenerative bone disorders, encompassing conditions such as intervertebral disc degeneration (IVDD), osteoarthritis (OA), and osteoporosis (OP), exert a profound impact on the well-being of individuals. These disorders pose a substantial healthcare burden, particularly within aging populations.1 These insidious conditions are characterized with the relentless decay of bone and cartilage, causing excruciating pain, debilitating loss of function, and a significant reduction in mobility.2–4 Unfortunately, the available treatment options for degenerative bone disorders remain frustratingly limited, often falling short of delivering long-term relief and restoration. The current medical context underscores the imperative to promptly attend to these unmet medical requirements, inciting an increasing enthusiasm for the investigation of novel therapeutic strategies.
In recent years, the landscape of regenerative medicine has been transformed by the emergence of stem cell-derived extracellular vesicles (EVs) therapies, presenting a promising approach for improving degenerative bone disorders. These diminutive yet potent membrane-enclosed vesicles, released by stem cells, have emerged as the effective factors responsible for the regenerative outcomes witnessed in stem cell therapies. With their bioactive cargo, these EVs derived from stem cells release a complex of regenerative signals, directing a coordinated interaction within the complicated microenvironment of deteriorating bone. This captivating phenomenon has captured the attention of both researchers and clinicians, as stem cell-derived EVs show their remarkable potential in reshaping the landscape of regenerative medicine.5 These EVs contain a diverse cargo of bioactive molecules, including proteins, lipids, and RNAs, which can be transferred to recipient cells. Through this transfer, stem cell-derived EVs participate in crucial molecular signaling, influencing cellular behavior and initiating regenerative processes within target tissues.6 For instance, Rong et al made a significant discovery when they conducted a study that shed light on the remarkable capabilities of neural stem cell-derived EVs. Their research revealed that these tiny vesicles, released by neural stem cells, played a pivotal role in the upregulation of autophagy marker proteins LC3B and beclin-1. As a result, a cascade of events unfolded, leading to the enhancement of autophagosome formation, which in turn contributed to the promotion of functional recovery in individuals with spinal cord injuries.7
By harnessing the inherent regenerative attributes of stem cells and utilizing the distinctive cargo encapsulated within their secreted EVs, researchers and clinicians aspire to surmount the constraints frequently linked to conventional therapeutic modalities. In contrast to conventional methods primarily centered on symptom management, this pioneering strategy aims to exploit the innate healing potential of the human body. The utilization of EVs derived from stem cells heralds a huge transformation in medical intervention, ushering in a new frontier of prospects for addressing a diverse array of diseases. In this extensive review, we perform an intriguing exploration to investigate the captivating domain of therapies involving EVs derived from stem cells, with a particular emphasis on their prospective applications in revitalizing degenerated bone tissues. With the objective of elucidating the present state of understanding within this domain, we discuss and summarize the intricate mechanisms that govern the regenerative processes, and we accentuate the therapeutic ramifications for three prominent conditions: IVDD, OA, and OP.
Isolation and Characterization of EVs, and Their Roles in Intercellular Communication
In this section, we focus on investigating the methods and techniques for EVs isolation and characterization, their delivered cargo and their involvement in intercellular communication. We aim to thoroughly unravel the complex tapestry of EVs. This endeavor entails an in-depth exploration of their definition, distinctive features, molecular mechanisms, cargo composition, and the extraordinary biological consequences that arise from the horizontal transfer of their cargo to recipient cells.
Definition and Characteristics of EVs
Within the captivating sphere of cellular communication, an exceptional category of entities comes to the fore—EVs. These captivating microstructures, delicately ensconced within membranes, harbor the potential to reveal a promising frontier in therapeutics. Secreted by cells into the extracellular space, these nanoscale vesicles embody a captivating diversity that defies conventional expectations. The extensive spectrum of EVs is classified into various subtypes, including the captivating microvesicles, the enigmatic exosomes, and the mysterious apoptotic bodies (Figure 1). Each of these subtypes boasts its distinctive characteristics and origins, perpetually enthralling the scientific community.8
 |
Figure 1 Schematic illustration of the biogenesis of microvesicles, exosomes, and apoptotic bodies. Created with Medpeer (https://medpeer.cn/). |
From the preservation of homeostasis to the orchestration of intricate disease pathways, EVs assume a central role, mediating the behavior and destiny of cells.9 These diminutive messengers encapsulate within them a universe of potential, containing nucleic acids, lipids, and proteins, each possessing a distinctive composition functionally customized to their specific cells of origin.10 It is through this exquisite cargo that EVs wield their power, mediating targeted cellular responses and sculpting the destiny of neighboring cells.
The exploration of EVs and the intricate mechanisms dictating their generation and functions reveals a captivating panorama. It spans the nuanced regulation of normal tissue physiology to the complex repercussions of pathological alterations and the intricate process of organ remodeling.8 With each revelation, the potential of EVs as innovative diagnostic and therapeutic tools becomes increasingly conspicuous, catalyzing advanced investigations aimed at harnessing their extraordinary capabilities.11
Composition of EVs: Proteins, Lipids, and RNAs
Within the captivating filed of EVs, a series of diverse cargo composition unfolds, reflecting the cellular origin of these remarkable vesicles. Proteins, lipids, and nucleic acids encapsulated within EVs amalgamate to define their functional attributes and confer upon them the capability to functionally regulate the behavior of recipient cells.12
Proteins assume a prominent role within the payload of EVs, their presence serving as a testament to the intricate molecular interaction within these vesicles. Enzymes, growth factors, receptors, and signaling molecules coalesce, engaging actively in cellular processes that propel tissue regeneration.12 Previous studies have unveiled the intriguing composition of saliva-derived extracellular vesicles (saliva-Exos), highlighting the presence of the ubiquitin-conjugating enzyme E2O (UBE2O). These saliva-Exos, enriched with UBE2O, exhibit a remarkable ability to reduce the expression of SMAD family member 6 (SMAD6), consequently triggering the activation of bone morphogenetic protein 2 (BMP2). This intricate cascade sets the stage for the stimulation of angiogenesis, enhancing the growth of new blood vessels and fostering tissue regeneration.13
Lipids emerge as another vital component within the repertoire of EV cargo, their presence playing an essential role in maintaining the structural integrity of these nanoscale vesicles. Moreover, specific lipid molecules carried within EVs hold the power to modulate cellular responses, orchestrating inflammation, angiogenesis, and immune regulation, thereby shaping the regenerative milieu.12 The regenerative potential of EVs finds its foundational basis within the delicate equilibrium of these lipids, indicating a complex process of molecular interactions that intricately mediate the fate of recipient cells.
Adding an additional layer of intricacy to the functional versatility of EVs, various categories of RNA molecules establish their residence within these captivating vesicles. Among these RNA molecules are messenger RNA (mRNA), microRNA (miRNA), and various other non-coding RNA species, all enclosed within EVs, ready for transfer to recipient cells.14,15 Once internalized, these transferred RNA molecules hold the key to unlocking regenerative processes. They can either be translated into proteins, actively participating in cellular functions, or take on the role of gene expression regulators through mechanisms such as RNA interference. This dynamic RNA transfer facilitates the modulation of cellular phenotypes, triggering the initiation of regenerative pathways and presenting an array of possibilities in the quest for tissue regeneration.
Upon their release, the cargo contained within EVs intermingles with cellular machinery, triggering intricate signaling cascades that govern a multitude of cellular functions. The proteins transferred from EVs interact with cell surface receptors, initiating sequences of events that govern cellular behavior. These proteins may even penetrate the realms of intracellular pathways, possessing the capability to regulate intricate molecular processes and conduct the regenerative potential concealed within each cell.12
Interplay of Metabolic Effects, Inflammation, Cellular Crosstalk, and Osteogenic Activity
Osteogenic activity, the process of bone formation, is intricately regulated by a complex interplay of metabolic factors, inflammation, and cellular crosstalk. Recent research has uncovered the profound influence of metabolic factors on osteogenic processes.16 Obesity, characterized by chronic low-grade inflammation, has been associated with impaired bone formation.17 Adipose tissue-derived hormones (adipokines), such as leptin and adiponectin, play a role in regulating bone metabolism. High levels of leptin, as observed in obesity, can suppress osteoblast function, while adiponectin may have a protective effect on bone health.18–20 Of note, individuals with diabetes often exhibit an increased risk of osteoporosis and fractures. Hyperglycemia, insulin resistance, and advanced glycation end-products (AGEs) have been implicated in impairing osteoblast function and promoting osteoclast activity, contributing to reduced bone density.21 Hormonal imbalances, such as those seen in menopause or andropause, can significantly impact bone health.22 Estrogen deficiency in postmenopausal women leads to increased bone resorption, while in men, decreased testosterone levels are associated with lower bone mineral density.23
Furthermore, inflammation is a critical component of the bone remodeling process and can have both positive and negative effects on osteogenesis. Recent studies have elucidated the role of macrophage polarization in bone regeneration. M1 pro-inflammatory macrophages can promote osteoclast differentiation and bone resorption, while M2 anti-inflammatory macrophages support osteoblast differentiation and bone formation.24 The balance between these macrophage subsets is crucial for effective bone healing. Inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), can inhibit osteogenic differentiation and induce osteoclastogenesis. Conversely, anti-inflammatory cytokines like interleukin-10 (IL-10) may enhance osteogenesis.25
Moreover, the crosstalk between different cell types within the bone microenvironment plays a pivotal role in regulating osteogenic activity. Recent studies have highlighted the significance of direct communication between osteoblasts and osteoclasts. Osteoblast-derived factors, such as osteoprotegerin (OPG) and receptor activator of nuclear factor kappa-B ligand (RANKL), regulate osteoclast formation and activity, influencing bone resorption. The presence of a well-developed vascular network is essential for bone formation.26 In addition, recent findings emphasize the role of endothelial cells and angiogenic factors in supporting osteogenesis by providing a conducive microenvironment for bone-forming cells.27
Active Role of EVs in Stem Cell Therapy
EVs derived from stem cells undertake pivotal roles in stem cell therapy. This intricate process serves as the foundation of stem cell therapy, furnishing the driving force behind its regenerative and healing potential.28 In recent years, intensive interest has attracted to the extraordinary role of EVs as the central mediators of molecular signaling in the domain of stem cell therapy, captivating the scientific community.29 These captivating vesicles encapsulate the power to govern and regulate a variety of cellular processes, thus mediating the fate of neighboring cells through the transformative cargo they carry.
EVs exhibit the remarkable capacity to exert influence over a range of vital cellular processes, spanning from differentiation and migration to proliferation and immune regulation. For instance, in a remarkable study by Ge et al, it was discovered that EVs derived from MSCs hold the remarkable capability to enhance endothelial cell migration and stimulate angiogenesis, propelling the healing of damaged tissues and fostering the growth of new blood vessels.30 This excellent ability to influence neighboring cells within damaged or degenerated tissues sets EVs apart as potent catalysts for tissue regeneration.31
EVs offer several unique advantages over traditional signaling molecules and lipids when it comes to their role in stem cell therapy. EVs carry a selective cargo of proteins, nucleic acids, lipids, and metabolites.32 This cargo is protected within the vesicle, allowing for precise delivery to target cells. In contrast, traditional signaling molecules, like growth factors or cytokines, may have off-target effects or be rapidly degraded in the extracellular environment.
Furthermore, EVs protect their cargo from degradation by enzymes and environmental factors, increasing the stability and longevity of the signaling molecules they transport. This protection ensures that the cargo reaches its intended destination intact, which can be challenging with unprotected molecules.33,34
Importantly, EVs generally have lower immunogenic potential compared to some traditional signaling molecules. This is because EVs can carry self-antigens, reducing the risk of provoking an immune response when used in therapeutic applications.35 Traditional recombinant proteins or small molecules may be seen as foreign by the recipient’s immune system.
Notably, EVs can naturally home to specific tissues or cell types due to the presence of surface ligands or receptors on their membranes. This targeting ability can enhance the efficiency of paracrine signaling and stem cell therapy.33 In contrast, traditional signaling molecules often require additional modifications or carriers for specific tissue targeting.
Functionally, EVs carry a wide range of bioactive molecules, allowing them to influence multiple signaling pathways simultaneously.33 This versatility makes them potent regulators of various cellular processes, including cell proliferation, differentiation, immune modulation, and tissue repair. Traditional signaling molecules often have a more limited range of biological activities.
Moreover, EVs may have a more favorable safety profile compared to high doses of traditional signaling molecules.33 EVs can exert their effects at lower concentrations, reducing the risk of side effects or adverse reactions associated with high doses of recombinant proteins or growth factors. In addition, EVs can be isolated and customized from specific cell sources, such as patient-derived stem cells.33 This opens the door to personalized medicine approaches, tailoring EV-based therapies to individual patient needs.
Transfer of EVs’ Cargo to Target Cells and Its Biological Effects
EVs, including exosomes and microvesicles, are nanosized vesicles secreted by various cell types. They play a crucial role in cell-cell communication by transferring bioactive molecules such as proteins, lipids, nucleic acids, and metabolites from donor cells to recipient cells.33,36 Like ethereal messengers, EVs carry within them a cargo that holds the power to transform the destiny of recipient cells. This cargo, once released upon internalization, becomes an agent of change, directly interacting with cellular machinery and triggering cascades of signaling events that shape cellular behavior.37 The transferred proteins within EVs take on the role of orchestrators, engaging with cell surface receptors or delving into intracellular pathways, regulating cellular functions (Figure 2).37 The interaction between EVs and recipient cells involves intricate mechanisms that include receptor-mediated interactions, endocytosis, and intracellular signaling pathways.
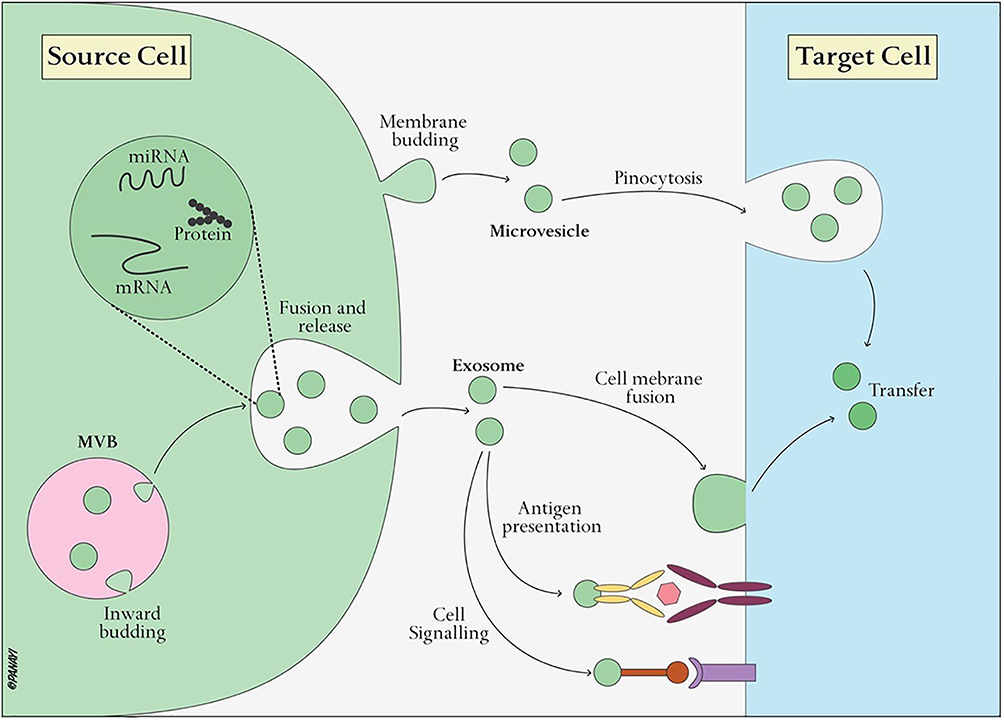 |
Figure 2 Illustration of transfer of EVs cargo to target cells and their biological effects. Reproduced from Xie X, Xiong Y, Panayi AC, et al. Exosomes as a Novel Approach to Reverse Osteoporosis: A Review of the Literature. Front Bioeng Biotechnol. 2020;8:594247; Copyright 2020, Frontiers. Creative Commons.37 |
EVs carry specific surface molecules or ligands that can interact with receptors on the recipient cell surface. These ligand-receptor interactions are highly selective and enable targeted communication.36,38 For example, tetraspanins, integrins, and adhesion molecules on EV surfaces can interact with corresponding receptors on recipient cells.38 Furthermore, EVs can carry ICAMs on their surfaces, which can bind to lymphocyte function-associated antigen 1 (LFA-1) receptors on immune cells, facilitating immune cell adhesion and subsequent immune response modulation.39
Moreover, larger EVs, such as microvesicles, can be engulfed by recipient cells through phagocytosis, a process where the cell membrane surrounds and engulfs the EV.40 This process is often used by immune cells to capture EVs released by pathogens or damaged cells. EVs can also be internalized through micropinocytosis, a non-specific form of endocytosis where cells engulf small particles, including exosomes.41 Some studies suggest that specific interactions between EV surface proteins and receptors on recipient cells can trigger clathrin-mediated endocytosis. This mechanism allows for the selective uptake of EVs.42
The transference of EVs’ cargo facilitates an interaction between discrete cell populations within injured or degenerated tissues, nurturing a regenerative milieu in which cells cooperate to promote repair and regeneration.43 The effects triggered by the transferred cargo are profound, encompassing the promotion of cell proliferation, the modulation of immune responses, the stimulation of angiogenesis, and the enhancement of extracellular matrix synthesis. Each of these effects constitutes a vital component on the process of tissue repair and regeneration.44
Sources of Stem Cell-Derived EVs for Bone Regeneration
Mesenchymal Stem Cells (MSCs)-Derived EVs
MSCs hold tremendous potential as a remarkable source of EVs that could revolutionize the field of bone regeneration.45 Operating as stromal cells, MSCs exhibit the remarkable capability to undergo differentiation into bone, cartilage, and adipose tissue. These versatile cells are found in various tissues, such as bone marrow, adipose tissue, the umbilical cord, and placental tissue, providing readily accessible sources for isolation and subsequent culture expansion. (Figure 3).46 The EVs generated by MSCs comprise a complex composition of biomolecules that closely parallel the distinctive differentiation capabilities of their progenitor cells.47
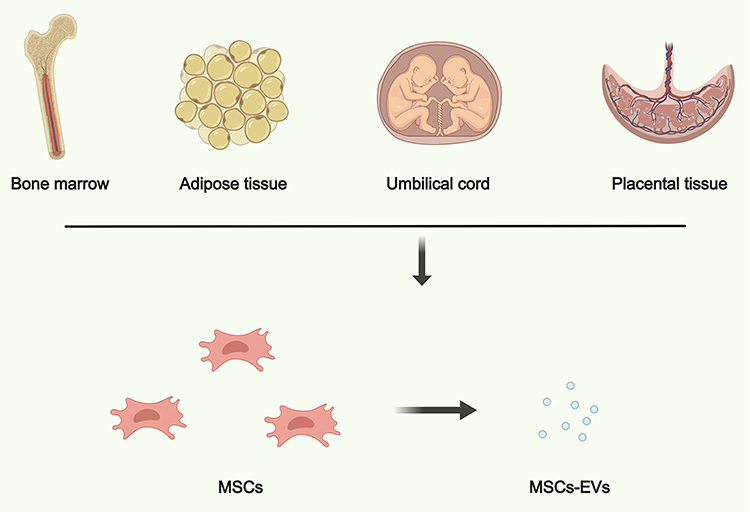 |
Figure 3 An overview of EVs derived from the frequently utilized sources, including bone marrow, adipose tissue, umbilical cord, and placental tissue. Created with Medpeer (https://medpeer.cn/). |
Bone marrow is a rich source of MSCs, hematopoietic stem cells (HSCs), and other cell types. EVs can be isolated from the supernatant of cultured bone marrow-derived cells.48 Bone marrow-derived EVs often carry a diverse cargo of proteins, miRNAs, and other bioactive molecules. They are known for their potential to modulate immune responses, promote tissue repair, and support hematopoiesis.49 These EVs have shown promise in regenerative medicine, especially in the context of bone regeneration, wound healing, and immunomodulation. They may also have therapeutic potential in conditions like graft-versus-host disease (GVHD) and autoimmune disorders.12
Furthermore, EVs can be isolated from adipose-derived stem/stromal cells (ADSCs) or mature adipocytes.50 Adipose-derived EVs are known to carry growth factors, cytokines, and adipokines, which can influence processes such as angiogenesis, inflammation, and metabolism.48 These EVs have been explored for their potential in promoting tissue regeneration, particularly in bone repair, plastic and reconstructive surgery, and musculoskeletal disorders.51
Moreover, the umbilical cord, specifically Wharton’s Jelly, is a valuable source of MSCs and their derived EVs.52 These cells can be isolated from the umbilical cord tissue. Umbilical cord-derived EVs are known for their relatively high proliferative capacity and immunomodulatory properties. They carry a range of bioactive molecules that can modulate inflammation and support tissue repair.53 These EVs have been studied for their potential in various therapeutic applications, including the treatment of skeletal degenerative diseases, wound healing, and immune-related disorders.54
In addition, the placenta is a temporary organ that develops during pregnancy to provide essential nutrients, oxygen, and waste removal for the developing fetus. It is composed of various cell types, including trophoblasts, MSCs, and endothelial cells, all of which can contribute to the release of MSCs-derived EVs.55 Placental EVs carry a diverse cargo, including proteins, lipids, nucleic acids (such as miRNAs, mRNAs, and DNA), and growth factors. This cargo is essential for regulating fetal development, maternal-fetal immune interactions, and tissue repair processes. Placental EVs often exhibit strong immunomodulatory properties.56 They can suppress maternal immune responses to prevent rejection of the fetus while also promoting tolerance. Of note, placental EVs are rich in growth factors and angiogenic factors, making them potential candidates for promoting tissue repair and regeneration in various medical fields, including bone repair and tissue engineering.57
Compared to the transplantation of MSCs themselves, the utilization of EVs presents a compelling alternative that mitigates certain risks associated with cell-based therapies. Concerns such as unwanted differentiation, immune rejection, and teratoma formation can be effectively decreased by utilizing MSC-derived EVs.12 These naturally secreted products have emerged as a promising therapeutic modality, enabling their isolation under scalable culture systems and subsequent storage as an “off-The-shelf” product. Moreover, their delivery can be achieved through minimally invasive procedures, further enhancing their clinical applicability.
Embryonic Stem Cells (ESCs)-Derived EVs
ESCs possess extraordinary attributes that set them apart, including their remarkable pluripotent nature, which enables them to differentiate into any cell type found in the human body.58 This inherent versatility makes ESCs a promising alternative to explore for various medical applications, particularly in bone regeneration.59
EVs originating from ESCs are not mere carriers; they are laden with a diverse array of bioactive molecules that hold immense potential for stimulating bone regeneration.60 These EVs contain an array of growth factors, morphogens, and developmental regulators, possessing the capacity to shape the destiny and actions of recipient cells. Through this capability, they can actively stimulate osteogenic differentiation and contribute to the regeneration of deteriorated bone tissue.
An exemplary study conducted by Gong et al demonstrated the tangible benefits of harnessing ESC-EVs for therapeutic purposes.61 In their research, the application of ESC-EVs proved to be a rescue mission for senescent MSCs, effectively preventing age-related bone loss in aging mice. The transcriptome analysis conducted as part of the study yielded fascinating results, unveiling that treatment with ESC-EVs led to a significant upregulation in the expression of genes associated with anti-aging, stem cell proliferation, and osteogenic differentiation within MSCs. This remarkable finding suggests that ESC-EVs hold enormous potential for reversing the senescence of MSCs and addressing age-related osteogenic dysfunction.
Induced Pluripotent Stem Cells (iPSCs)-Derived EVs
iPSCs have undergone a process of reprogramming to assume a state resembling that of embryonic stem cells.62 This reprogramming is accomplished through the introduction of specific transcription factors associated with pluripotency. Similar to ESCs, iPSCs possess the extraordinary ability to self-renew indefinitely and differentiate into any cell type found in the body.63 This exceptional potential has paved the way for iPSC-derived EVs (iPSC-EVs) to emerge as a promising cell-free platform for regenerative therapy.
The EVs secreted by iPSCs demonstrate regenerative effects similar to those derived from ESCs. Moreover, they exhibit shared surface markers and molecular signatures.64 These vesicles encapsulate a wide array of essential components, including proteins, lipids, mRNAs, miRNAs, and growth factors associated with crucial cellular processes such as proliferation, anti-apoptosis, migration, differentiation, and ECM production. Notably, signaling molecules contained within iPSC-EVs, such as Wnt proteins, transforming growth factor-β, and ECM proteins, have demonstrated positive regulatory effects on bone formation and regeneration.65 These inherent properties make iPSC-EVs a powerful tool for enhancing regenerative processes within the bone.
In a prior study, the potential of iPSC-EVs to rejuvenate senescent nucleus pulposus cells (NPCs) and attenuate the progression of IVDD was demonstrated.66 Mechanistically, the therapeutic effects of iPSC-EVs were attributed, in part, to the delivery of exosomal miR-105-5p to the senescent NPCs, thereby activating the Sirt6 signaling pathway. These remarkable findings underscore the potential of iPSCs as a reliable and scalable source for obtaining EVs, while also highlighting the immense therapeutic promise of iPSC-EVs in the context of degenerative bone disorders.
Therapeutic Potential of Stem Cell-Derived EVs in Degenerative Bone Disorders
IVDD
IVDD is a multifaceted condition characterized by the progressive deterioration of the discs that connect adjacent vertebrae in the spine. The development of IVDD involves a complex interplay of intrinsic and extrinsic factors. Among these factors, aging plays a significant role, as it brings about natural changes that negatively affect disc health over time. These changes include reduced nutrient transport into the discs, cellular senescence, and increased calcification of the matrix. Excessive or abnormal mechanical loading further accelerates the damage to intervertebral discs by triggering inflammatory responses and the activation of enzymes that degrade the ECM.67 Inflammatory cytokines, such as TNF-α and IL-1, play a pivotal role in propagating disc cell apoptosis and the breakdown of the ECM. The delicate balance between matrix synthesis and degradation becomes disrupted. Catabolic enzymes, including matrix metalloproteinases (MMPs), are upregulated, leading to excessive degradation of ECM components such as collagen and proteoglycans.68 Additionally, the capacity of disc cells to synthesize new matrix diminishes as their numbers and functionality decline over time.67,69
Current treatment options for IVDD are largely limited to providing symptomatic relief, rather than modifying the underlying disease process. To effectively reverse or decelerate disc degeneration, a multimodal approach is required, one that focuses on restoring matrix homeostasis, supporting disc cell nutrition and viability, and mitigating inflammation and abnormal mechanical stressors.70 In light of these challenges, stem cell-derived EVs have emerged as a promising therapeutic approach for intervertebral disc degeneration.71 Stem cell-derived EVs offer a cell-free alternative to stem cell transplantation, harnessing the rejuvenated potential of stem cells while avoiding potential drawbacks associated with direct cell injection.72,73
The regenerative effects of stem cell-derived EVs are mediated through various mechanisms.74,75 Firstly, the cargo carried by EVs, consisting of growth factors, cytokines, and ECM components, can stimulate the synthesis of new ECM molecules, promoting tissue remodeling and restoring disc homeostasis. These bioactive molecules contribute to the enhancement of disc cell viability and function.76 Secondly, stem cell-derived EVs exert a modulatory effect on inflammatory responses within the degenerated disc. They can attenuate excessive inflammation, reduce the secretion of pro-inflammatory cytokines, and establish an anti-inflammatory microenvironment. By suppressing inflammatory processes, EVs create a favorable environment for tissue regeneration while limiting further disc degeneration.77 Thirdly, EVs play a crucial role in promoting cell survival and inhibiting apoptosis in the degenerated disc. Through the transfer of specific molecules, such as anti-apoptotic factors and microRNAs, EVs protect disc cells from apoptotic signals, enhancing their survival.78 Furthermore, stem cell-derived EVs have the potential to induce the differentiation of endogenous progenitor cells toward a discogenic lineage. Through paracrine signaling and the delivery of specific factors, EVs can direct the differentiation of resident disc cells or recruited progenitor cells into disc-like cells, thereby promoting tissue revitalization and functional recovery.79
The remarkable potential of stem cell-derived EVs is their ability to harness these diverse mechanisms of action to address the complex challenges associated with intervertebral disc degeneration. By delivering a rich cargo of bioactive molecules, including growth factors, cytokines, and ECM components, EVs stimulate the synthesis of new matrix, promote tissue remodeling, and restore the delicate balance within the disc microenvironment.80 This multifaceted approach addresses the underlying matrix imbalance and supports the viability and functionality of disc cells. The Wang et al group conducted a study demonstrating an innovative approach to deliver MSCs-EVs for the treatment of disc degeneration. They developed a functional hydrogel composed of quaternized chitosan and oxidized starch, which served as a delivery system for the MSCs-EVs. The results of the study showed that the MSCs-EVs released from this hydrogel had a regenerative effect on senescent NPs, promoting ECM remodeling, and partially restoring the structures of the NPs and annulus fibrosus (Figure 4).81 These findings represent a significant advance in disc degeneration therapy by targeting NPs cell senescence. The use of the functional hydrogel as a delivery system enables controlled release and localization of the MSCs-EVs, ensuring their targeted action within the degenerated disc.
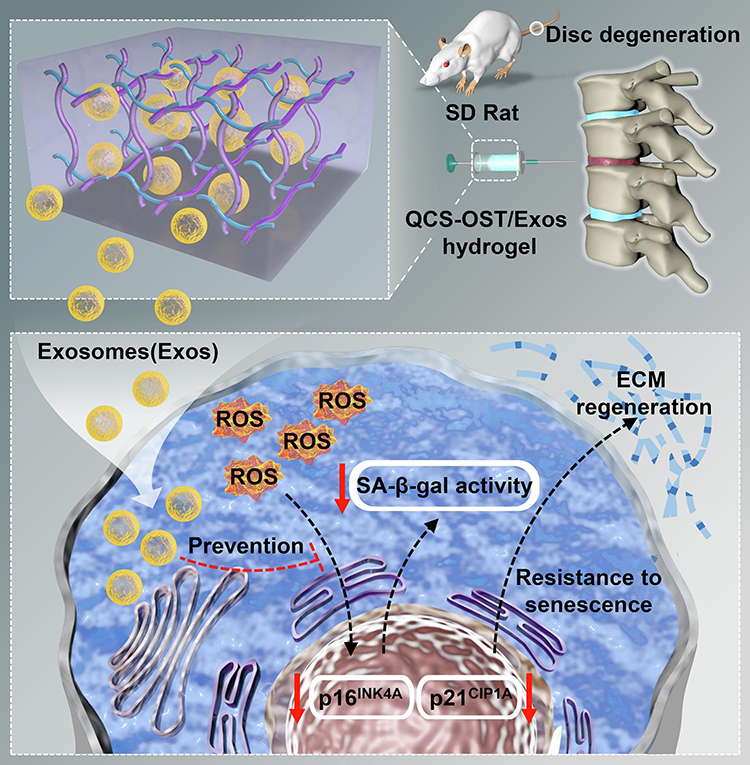 |
Figure 4 Schematic illustration of the stem cells-derived EVs for IVDD therapy. The therapeutic approaches aimed at addressing cellular senescence in nucleus pulposus cells could potentially decelerate the development of disc degeneration. Reprinted from Int J Biol Macromol, 232, Guan M, Liu C, Zheng Q, et al. Exosome-laden injectable self-healing hydrogel based on quaternized chitosan and oxidized starch attenuates disc degeneration by suppressing nucleus pulposus senescence.123479, Copyright 2023, Creative Commons.81 |
Furthermore, the anti-inflammatory properties of stem cell-derived EVs offer an effective means to modulate the inflammatory processes that contribute to disc degeneration. By reducing the secretion of pro-inflammatory cytokines and creating an anti-inflammatory milieu, EVs help alleviate the destructive inflammatory responses, creating an environment conducive to tissue regeneration and healing.82 In addition to their role in modulating inflammation and matrix homeostasis, stem cell-derived EVs exhibit a remarkable ability to promote cell survival and inhibit apoptosis within the degenerated disc.83 Through the transfer of specific molecules, such as anti-apoptotic factors and miRNAs, EVs confer protective effects on disc cells, shielding them from apoptotic signals and enhancing their survival. This preservation of disc cellularity is vital for maintaining the overall health and function of the intervertebral disc.84
Furthermore, EVs derived from stem cells hold the potential to harness the regenerative capabilities of endogenous progenitor cells. Through the delivery of paracrine signals and specific factors, these EVs can modulate the differentiation of resident disc cells or recruited progenitor cells toward a discogenic lineage. This directed differentiation promotes the regeneration of disc-like cells, facilitating tissue repair and functional recovery.85
Osteoarthritis (OA)
OA is a debilitating degenerative joint disease characterized by the gradual destruction of articular cartilage and surrounding tissues, leading to structural changes and a loss of mobility. The development of OA is influenced by a combination of mechanical factors, genetic influences, aging processes, and chronic inflammation, all of which interact to disrupt normal cartilage homeostasis.86
Chronic inflammation plays a crucial role in the pathogenesis and progression of OA. In osteoarthritic joints, pro-inflammatory cytokines such as interleukin-1 beta (IL-1β) and tumor necrosis factor alpha (TNF-α) are overexpressed.87 These cytokines stimulate chondrocytes and synoviocytes to release matrix metalloproteinases (MMPs), aggrecanases, and other catabolic factors. The collective action of these enzymes and factors damages the cartilage matrix, leading to fibrillation, osteophyte formation, and overall cartilage matrix degradation.88 Conversely, the production of anti-inflammatory and anabolic regulators, such as IL-4, IL-10, and insulin-like growth factor-1 (IGF-1), is suppressed, impeding matrix repair and tissue regeneration.89
The imbalance between cartilage matrix synthesis and breakdown is a key driver of joint degeneration in OA. MMPs and aggrecanases play a critical role in the degradation of collagen fibers and proteoglycans, including aggrecan, within the cartilage matrix. Proteases such as MMP-1, MMP-3, MMP-7, MMP-9, and MMP-13, are major contributors to accelerated matrix degradation.90 Over time, the excessive breakdown of the matrix coupled with insufficient anabolic activity depletes cartilage of its crucial macromolecules, resulting in fissures, fibrillation, and ultimately, full-thickness cartilage loss, which characterizes end-stage OA.91
In recent years, EVs have emerged as a promising therapeutic approach for the treatment of OA.92,93 These EVs, derived from various stem cell sources, offer a cell-free alternative with regenerative properties similar to stem cell transplantation. Stem cell-derived EVs exert their beneficial effects in OA through multiple mechanisms.94 Firstly, EVs carry a cargo of bioactive molecules, including growth factors, cytokines, and anti-inflammatory factors, which can modulate the local microenvironment within the diseased joint. These molecules have the potential to counteract inflammation, promote tissue repair, and enhance the synthesis of crucial ECM components, such as type II collagen and proteoglycans, within the damaged cartilage.95 Secondly, EVs can regulate chondrocyte metabolism and function. Through the transfer of specific miRNAs via EVs, gene expression in recipient cells, including chondrocytes, can be modulated. This miRNA-mediated regulation suppresses the activity of catabolic enzymes and inflammatory mediators, while promoting anabolic processes in chondrocytes.47 This regulation helps attenuate cartilage degradation, enhance matrix synthesis, and support the survival of chondrocytes, ultimately contributing to the preservation of cartilage health.96 For instance, Sun et al reported a EVs-based therapy for cartilage regeneration for OA treatment. In this study, the researchers demonstrated that EVs derived from TGFβ3-preconditioned BMSCs presented enriched specific miRNAs that could be transferred to native BMSCs to promote chondrogenesis. Mechanistically, the overexpression of miR-455, released from the EVs derived from TGFβ3-preconditioned MSCs, was able to enhance OA alleviation and cartilage regeneration by activation of the SOX11/FOXO signaling pathway (Figure 5).97
 |
Figure 5 (A) Schematic illustration of the EVs-based strategy. (B) Heatmap of clustering dysregulated miRNA expression profiles with microarray in the different groups. (C) Injection of EVs-conjugated composite for cartilage regeneration in rat OA model. (D) Chondrogenesis was defined with immunostaining of different biomarkers. (E) Intra-articular delivery of the labeled EVs were examined with in vivo fluorescence imaging. Reproduced from Sun Y, Zhao J, Wu Q, et al. Chondrogenic primed extracellular vesicles activate miR-455/SOX11/FOXO axis for cartilage regeneration and osteoarthritis treatment. NPJ Regen Med. 2022;7(1):53. Creative Commons.97 |
Furthermore, stem cell-derived EVs possess remarkable immunomodulatory properties that can regulate immune responses within the osteoarthritic joint.96 These EVs have the ability to suppress the activation of immune cells, such as macrophages, and inhibit the release of pro-inflammatory cytokines. By doing so, they reduce the overall inflammatory burden, creating a more favorable environment for tissue regeneration and repair.98
Osteoporosis (OP)
OP, a debilitating skeletal disorder, manifests as a decline in bone mineral density and the deterioration of bone microarchitecture. Consequently, bones become fragile and prone to fractures. The pathogenesis of OP involves a multifaceted interplay among several factors, including the aging process, hormonal fluctuations, lifestyle decisions, and genetic predisposition. These elements disrupt the intricate equilibrium between bone formation and resorption.99
At the center of OP is a fundamental imbalance in the two pivotal processes that regulate bone turnover: bone formation by osteoblasts and bone resorption by osteoclasts.100 When osteoclastic activity surpasses osteoblastic bone formation over an extended period, there is a net loss of bone tissue, mineral content, and matrix proteins such as collagen.101 Consequently, the microarchitectural structure of bones deteriorates, compromising their strength and load-bearing capacity.
Stem cell-derived EVs have emerged as a promising cell-free alternative with regenerative properties that could potentially stimulate bone formation and improve bone quality in OP.102 Firstly, EVs carry a diverse cargo of bioactive molecules, including growth factors, cytokines, and extracellular matrix proteins, which have the potential to enhance osteoblast activity and bone formation. These molecules can promote the proliferation and differentiation of osteoblasts, as well as the synthesis of bone matrix components, ultimately leading to increased bone mineralization and improved bone strength.103
Moreover, stem cell-derived EVs possess the remarkable ability to modulate osteoclast activity and bone resorption.104 EVs can transport specific miRNAs that target genes involved in osteoclast differentiation and function, effectively inhibiting their activity. By regulating the delicate balance between osteoblast-mediated bone formation and osteoclast-mediated bone resorption, stem cell-derived EVs have the potential to restore bone homeostasis and mitigate bone loss in OP.105 In a prior study conducted by Yang et al, a remarkable discovery was made regarding the potential of bioactive glass nanoparticles (BGN) in inducing the expression of EVs secreted by MSCs. These EVs were found to possess a unique ability to suppress osteoclast differentiation and reverse the pathogenesis of osteoporosis, shedding light on a promising therapeutic approach.106 Of note, within these functional EVs, a key factor was identified: a long non-coding RNA called NRON. By binding to these crucial factors, NRON effectively disrupted their movement into the nucleus, ultimately leading to the inhibition of osteoclast differentiation (Figure 6).
 |
Figure 6 (A) Schematic illustration of fabrication and function of BGN/EVs in OA therapy. (B) Representative H&E and Masson staining image of femurs, with a scale bar indicating a length of 500 μm. Reprinted from Biomaterials, vol 283, Yang Z, Liu X, Zhao F, et al. Bioactive glass nanoparticles inhibit osteoclast differentiation and osteoporotic bone loss by activating lncRNA NRON expression in the extracellular vesicles derived from bone marrow mesenchymal stem cells. 121438, Copyright 2022, with permission from Elsevier.106 |
The regenerative properties of stem cell-derived EVs offer a promising strategy for innovative and effective treatments for individuals affected by OP. By stimulating osteoblast-mediated bone formation and modulating osteoclast activity, these EVs hold the potential to restore bone homeostasis, enhance bone strength, and reduce fracture risk. Continued research and development in this field may pave the way for advanced therapies that improve skeletal health and enhance the quality of life for those living with OP.
Advantages of EV-Based Therapies Over Stem Cell Transplantation
One of the remarkable advantages of utilizing EV-based therapies is their cell-free nature, providing a solution to the inherent limitations associated with cell availability. Traditional stem cell transplantation often relies on obtaining an adequate number of viable and functional cells, which can be a considerable challenge, particularly when utilizing the patient’s own cells.107 The process of isolating, expanding, and characterizing stem cells can be laborious, time-consuming, and resource-intensive, posing significant obstacles to widespread implementation.
In contrast, EVs provide a more accessible and readily available alternative. These minuscule vesicles can be easily extracted from stem cells and produced in significant quantities, thereby ushering in a revolution in regenerative medicine.108 By harnessing the regenerative and regenerative potential encapsulated within EVs, the limitations posed by cell availability can be effectively overcome. The immediate availability of EVs, without the need for extensive cell expansion or laborious characterization processes, represents a paradigm shift in regenerative therapies.109 This advantage is promising in expediting treatment timelines, providing relief to patients who are in urgent need of therapeutic intervention.
Another significant advantage of EV-based therapies is their inherent ability to circumvent the potential concern of tumorigenicity often associated with stem cell transplantation.110 While stem cells possess the remarkable capacity for self-renewal and differentiation, which is crucial for their regenerative potential, this property raises the risk of uncontrolled cell proliferation and the formation of tumors. By utilizing EVs as therapeutic agents instead of stem cells, the risk of tumorigenicity is significantly minimized. EVs, as carriers of bioactive molecules such as growth factors and regulatory RNAs, play a pivotal role in mediating the regenerative effects.111 These bioactive molecules are responsible for eliciting therapeutic responses within the recipient tissue or organ. Importantly, they achieve this without the direct presence of live cells, thereby reducing the potential for abnormal cell growth and tumor formation.
Challenges and Opportunities
Although the potential of EVs as therapeutics for bone degenerative disorders is promising, there are several key challenges that should be addressed to fully harness their potential and translate them into clinical practice.
First and foremost, a deeper understanding of the underlying mechanisms by which EVs exert their effects and optimizing strategies for their targeted delivery to skeletal tissues are critical areas for ongoing research. Unraveling the intricate molecular pathways and cargo components responsible for the osteogenic and bone-modulatory effects of EVs will provide valuable insights into their bioactivity and guide the development of strategies to enhance their therapeutic potency.
Further investigations are needed to better define the uptake mechanisms of EVs by target cells and the subsequent signaling events they trigger. Comparative studies exploring the effects of EVs derived from different cell sources may uncover key factors influencing their functional outcomes. Efforts to improve the efficiency of EV transport and uptake at sites of bone disease will be crucial for maximizing their therapeutic effectiveness.
Exploring approaches to modify EV surface properties for targeted delivery and investigating biomaterial systems and nanoplatforms to protect EVs during circulation and enable controlled release at specific target sites may further improve their therapeutic outcomes. These research endeavors are essential for advancing the clinical potential of EVs in the field of regenerative medicine for bone disorders.
Moreover, regulatory and manufacturing considerations present challenges that demand collaborative efforts across multiple disciplines to facilitate the development of EV-based interventions. Standardizing methods for isolating, quantifying, and characterizing EVs is of utmost importance to ensure the production of clinical-grade products with well-defined specifications and quality controls. Regulatory approval necessitates comprehensive evidence of safety, purity, potency, and reproducibility based on good manufacturing practices. Rigorous clinical trials are essential to determine optimal dosing, treatment windows, and the long-term effects of EV therapies. Collaborations among research institutions, industry partners, and regulatory agencies will foster progress in this field, ensuring compliance with regulatory requirements while driving innovation. Such cooperative efforts will be instrumental in advancing the clinical application of EVs in regenerative medicine for bone disorders and beyond.
In addition, continued investigations in understanding EVs biology, optimizing scalable production methodologies, developing advanced delivery systems, and conducting robust clinical trials is crucial for realizing the full potential of EVs in regenerating bone tissue.
Despite the challenges that must be overcome, EVs represent a promising therapeutic approach for degenerative skeletal disorders. Their inherent regenerative capacity and ability to stimulate endogenous regenerative processes make them a compelling avenue for treatment. Interdisciplinary efforts currently underway to address manufacturing, delivery, safety, and regulatory issues will play a crucial role in advancing the clinical application of EV technologies for the treatment of degenerative bone disorders.
This review not only seeks to consolidate the existing knowledge regarding stem cell-derived EV therapies for degenerative bone disorders but also aims to highlight critical knowledge gaps that warrant further investigation. By addressing these gaps and surmounting the challenges on the horizon, we anticipate facilitating the translation of these remarkable therapeutic approaches into clinical practice.
Acknowledgments
We would like to thank Medpeer software (https://image.medpeer.cn/) for the picture source assistance during preparation of Figures 1 and 3.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Van Gastel N, Carmeliet G. Metabolic regulation of skeletal cell fate and function in physiology and disease. Nat Metab. 2021;3(1):11–20. doi:10.1038/s42255-020-00321-3
2. Barnett R. Osteoarthritis. Lancet. 2018;391(10134):1985. doi:10.1016/S0140-6736(18)31064-X
3. Cheng Z, Xiang Q, Wang J, Zhang Y. The potential role of melatonin in retarding intervertebral disc ageing and degeneration: a systematic review. Ageing Res Rev. 2021;70:101394. doi:10.1016/j.arr.2021.101394
4. Prestwood KM, Pilbeam CC, Raisz LG. Treatment of osteoporosis. Annu Rev Med. 1995;46:249–256. doi:10.1146/annurev.med.46.1.249
5. Weng Z, Zhang B, Wu C, et al. Therapeutic roles of mesenchymal stem cell-derived extracellular vesicles in cancer. J Hematol Oncol. 2021;14(1):136. doi:10.1186/s13045-021-01141-y
6. Zhao M, Liu S, Wang C, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate mitochondrial damage and inflammation by stabilizing mitochondrial DNA. ACS Nano. 2021;15(1):1519–1538. doi:10.1021/acsnano.0c08947
7. Rong Y, Liu W, Wang J, et al. Neural stem cell-derived small extracellular vesicles attenuate apoptosis and neuroinflammation after traumatic spinal cord injury by activating autophagy. Cell Death Dis. 2019;10(5):340. doi:10.1038/s41419-019-1571-8
8. Kalluri R, Lebleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
9. Van Niel G, D’angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
10. Gupta D, Zickler AM, El Andaloussi S. Dosing extracellular vesicles. Adv Drug Deliv Rev. 2021;178:113961. doi:10.1016/j.addr.2021.113961
11. He C, Zheng S, Luo Y, Wang B. Exosome theranostics: biology and translational medicine. Theranostics. 2018;8(1):237–255. doi:10.7150/thno.21945
12. Matsuzaka Y, Yashiro R. Therapeutic strategy of mesenchymal-stem-cell-derived extracellular vesicles as regenerative medicine. Int J Mol Sci. 2022;23(12):6480. doi:10.3390/ijms23126480
13. Mi B, Chen L, Xiong Y, et al. Saliva exosomes-derived UBE2O mRNA promotes angiogenesis in cutaneous wounds by targeting SMAD6. J Nanobiotechnology. 2020;18(1):68. doi:10.1186/s12951-020-00624-3
14. Wu X, Liu Y, Wei W, Liu ML. Extracellular vesicles in autoimmune vasculitis - Little dirts light the fire in blood vessels. Autoimmun Rev. 2019;18(6):593–606. doi:10.1016/j.autrev.2018.12.007
15. Alexandru N, Procopciuc A, Vilcu A, Comarita IK, Bӑdilӑ E, Georgescu A. Extracellular vesicles-incorporated microRNA signature as biomarker and diagnosis of prediabetes state and its complications. Rev Endocr Metab Disord. 2022;23(3):309–332. doi:10.1007/s11154-021-09664-y
16. Peng H, Hu B, Xie LQ, et al. A mechanosensitive lipolytic factor in the bone marrow promotes osteogenesis and lymphopoiesis. Cell Metab. 2022;34(8):1168–82 e6. doi:10.1016/j.cmet.2022.05.009
17. Ambrosi TH, Scialdone A, Graja A, et al. Adipocyte accumulation in the bone marrow during obesity and aging impairs stem cell-based hematopoietic and bone regeneration. Cell Stem Cell. 2017;20(6):771–84 e6. doi:10.1016/j.stem.2017.02.009
18. Xu ZH, Xiong CW, Miao KS, et al. Adipokines regulate mesenchymal stem cell osteogenic differentiation. World J Stem Cells. 2023;15(6):502–513. doi:10.4252/wjsc.v15.i6.502
19. Wang Y, Zhang X, Shao J, Liu H, Liu X, Luo E. Adiponectin regulates BMSC osteogenic differentiation and osteogenesis through the Wnt/beta-catenin pathway. Sci Rep. 2017;7(1):3652. doi:10.1038/s41598-017-03899-z
20. Zhang ZM, Jiang LS, Jiang SD, Dai LY. Osteogenic potential and responsiveness to leptin of mesenchymal stem cells between postmenopausal women with osteoarthritis and osteoporosis. J Orthop Res. 2009;27(8):1067–1073. doi:10.1002/jor.20846
21. Fu T, Zhu Q, Lou F, Cai S, Peng S, Xiao J. Advanced glycation end products inhibit the osteogenic differentiation potential of adipose-derived stem cells in mice through autophagy. Cell Signal. 2023;108:110694. doi:10.1016/j.cellsig.2023.110694
22. Han L, Wang B, Wang R, Gong S, Chen G, Xu W. The shift in the balance between osteoblastogenesis and adipogenesis of mesenchymal stem cells mediated by glucocorticoid receptor. Stem Cell Res Ther. 2019;10(1):377. doi:10.1186/s13287-019-1498-0
23. Zhang B, Li Y, Zhou Q, Ding Y. Estrogen deficiency leads to impaired osteogenic differentiation of periodontal ligament stem cells in rats. Tohoku J Exp Med. 2011;223(3):177–186. doi:10.1620/tjem.223.177
24. Xiong Y, Chen L, Yan C, et al. M2 Macrophagy-derived exosomal miRNA-5106 induces bone mesenchymal stem cells towards osteoblastic fate by targeting salt-inducible kinase 2 and 3. J Nanobiotechnology. 2020;18(1):66. doi:10.1186/s12951-020-00622-5
25. Xiong Y, Mi BB, Lin Z, et al. The role of the immune microenvironment in bone, cartilage, and soft tissue regeneration: from mechanism to therapeutic opportunity. Mil Med Res. 2022;9(1):65. doi:10.1186/s40779-022-00426-8
26. Cappariello A, Loftus A, Muraca M, Maurizi A, Rucci N, Teti A. Osteoblast-derived extracellular vesicles are biological tools for the delivery of active molecules to bone. J Bone Miner Res. 2018;33(3):517–533. doi:10.1002/jbmr.3332
27. Liu Y, Tian H, Hu Y, et al. Mechanosensitive Piezo1 is crucial for periosteal stem cell-mediated fracture healing. Int J Biol Sci. 2022;18(10):3961–3980. doi:10.7150/ijbs.71390
28. Yin Y, Chen H, Wang Y, Zhang L, Wang X. Roles of extracellular vesicles in the aging microenvironment and age-related diseases. J Extracell Vesicles. 2021;10(12):e12154. doi:10.1002/jev2.12154
29. Maacha S, Bhat AA, Jimenez L, et al. Extracellular vesicles-mediated intercellular communication: roles in the tumor microenvironment and anti-cancer drug resistance. Mol Cancer. 2019;18(1):55. doi:10.1186/s12943-019-0965-7
30. Ge L, Xun C, Li W, et al. Extracellular vesicles derived from hypoxia-preconditioned olfactory mucosa mesenchymal stem cells enhance angiogenesis via miR-612. J Nanobiotechnology. 2021;19(1):380. doi:10.1186/s12951-021-01126-6
31. Wu Q, Wang J, Tan WLW, et al. Extracellular vesicles from human embryonic stem cell-derived cardiovascular progenitor cells promote cardiac infarct healing through reducing cardiomyocyte death and promoting angiogenesis. Cell Death Dis. 2020;11(5):354. doi:10.1038/s41419-020-2508-y
32. Xiong Y, Chen L, Yan C, et al. Circulating exosomal miR-20b-5p inhibition restores wnt9b signaling and reverses diabetes-associated impaired wound healing. Small. 2020;16(3):e1904044. doi:10.1002/smll.201904044
33. Dai J, Su Y, Zhong S, et al. Exosomes: key players in cancer and potential therapeutic strategy. Signal Transduct Target Ther. 2020;5(1):145. doi:10.1038/s41392-020-00261-0
34. Lin Z, Xiong Y, Meng W, et al. Exosomal PD-L1 induces osteogenic differentiation and promotes fracture healing by acting as an immunosuppressant. Bioact Mater. 2022;13:300–311. doi:10.1016/j.bioactmat.2021.10.042
35. Zeng X, Zhang YD, Ma RY, et al. Activated Drp1 regulates p62-mediated autophagic flux and aggravates inflammation in cerebral ischemia-reperfusion via the ROS-RIP1/RIP3-exosome axis. Mil Med Res. 2022;9(1):25. doi:10.1186/s40779-022-00383-2
36. Hu Q, Zhang S, Yang Y, et al. Extracellular vesicles in the pathogenesis and treatment of acute lung injury. Mil Med Res. 2022;9(1):61. doi:10.1186/s40779-022-00417-9
37. Xie X, Xiong Y, Panayi AC, et al. Exosomes as a novel approach to reverse osteoporosis: a review of the literature. Front Bioeng Biotechnol. 2020;8:594247. doi:10.3389/fbioe.2020.594247
38. Zhdanov VP. Ligand-receptor-mediated attachment of lipid vesicles to a supported lipid bilayer. Eur Biophys J. 2020;49(5):395–400. doi:10.1007/s00249-020-01441-0
39. Chand S, Demarino C, Gowen A, et al. Methamphetamine induces the release of proadhesive extracellular vesicles and promotes syncytia formation: a potential role in HIV-1 neuropathogenesis. Viruses. 2022;14(3):550. doi:10.3390/v14030550
40. Li Y, Wu Y, Federzoni EA, et al. CD47 cross-dressing by extracellular vesicles expressing CD47 inhibits phagocytosis without transmitting cell death signals. Elife. 2022;2022:11.
41. Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;2014:3.
42. Cronqvist T, Erlandsson L, Tannetta D, Hansson SR. Placental syncytiotrophoblast extracellular vesicles enter primary endothelial cells through clathrin-mediated endocytosis. Placenta. 2020;100:133–141. doi:10.1016/j.placenta.2020.07.006
43. Garcia-Martin R, Wang G, Brandao BB, et al. MicroRNA sequence codes for small extracellular vesicle release and cellular retention. Nature. 2022;601(7893):446–451. doi:10.1038/s41586-021-04234-3
44. Kang M, Huang CC, Lu Y, et al. Bone regeneration is mediated by macrophage extracellular vesicles. Bone. 2020;141:115627. doi:10.1016/j.bone.2020.115627
45. Sonoda S, Yamaza T. Extracellular vesicles rejuvenate the microenvironmental modulating function of recipient tissue-specific mesenchymal stem cells in osteopenia treatment. Front Endocrinol. 2023;14:1151429. doi:10.3389/fendo.2023.1151429
46. Lei Q, Gao F, Liu T, et al. Extracellular vesicles deposit PCNA to rejuvenate aged bone marrow-derived mesenchymal stem cells and slow age-related degeneration. Sci Transl Med. 2021;13(578). doi:10.1126/scitranslmed.aaz8697
47. Yin B, Ni J, Witherel CE, et al. Harnessing tissue-derived extracellular vesicles for osteoarthritis theranostics. Theranostics. 2022;12(1):207–231. doi:10.7150/thno.62708
48. Hu Q, Lyon CJ, Fletcher JK, Tang W, Wan M, Hu TY. Extracellular vesicle activities regulating macrophage- and tissue-mediated injury and repair responses. Acta Pharm Sin B. 2021;11(6):1493–1512. doi:10.1016/j.apsb.2020.12.014
49. Liu M, Sun Y, Zhang Q. Emerging role of extracellular vesicles in bone remodeling. J Dent Res. 2018;97(8):859–868. doi:10.1177/0022034518764411
50. Rome S. Muscle and adipose tissue communicate with extracellular vesicles. Int J Mol Sci. 2022;23(13):7052. doi:10.3390/ijms23137052
51. Kahn CR, Wang G, Lee KY. Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. J Clin Invest. 2019;129(10):3990–4000. doi:10.1172/JCI129187
52. Arrigoni C, D’arrigo D, Rossella V, Candrian C, Albertini V, Moretti M. Umbilical Cord MSCs and their secretome in the therapy of arthritic diseases: a research and industrial perspective. Cells. 2020;9(6):1343. doi:10.3390/cells9061343
53. Lelek J, Zuba-Surma EK. Perspectives for future use of extracellular vesicles from umbilical cord- and adipose tissue-derived mesenchymal stem/stromal cells in regenerative therapies-synthetic review. Int J Mol Sci. 2020;21(3):799. doi:10.3390/ijms21030799
54. Vatsa P, Negi R, Ansari UA, Khanna VK, Pant AB. Insights of extracellular vesicles of mesenchymal stem cells: a prospective cell-free regenerative medicine for neurodegenerative disorders. Mol Neurobiol. 2022;59(1):459–474. doi:10.1007/s12035-021-02603-7
55. Andjus P, Kosanovic M, Milicevic K, et al. Extracellular vesicles as innovative tool for diagnosis, regeneration and protection against neurological damage. Int J Mol Sci. 2020;21(18):6859. doi:10.3390/ijms21186859
56. Maqsood M, Kang M, Wu X, Chen J, Teng L, Qiu L. Adult mesenchymal stem cells and their exosomes: sources, characteristics, and application in regenerative medicine. Life Sci. 2020;256:118002. doi:10.1016/j.lfs.2020.118002
57. Li T, Fu Y, Guo Z, et al. A new cell-free therapeutic strategy for liver regeneration: human placental mesenchymal stem cell-derived extracellular vesicles. J Tissue Eng. 2022;13:20417314221132093. doi:10.1177/20417314221132093
58. Dorronsoro A, Santiago FE, Grassi D, et al. Mesenchymal stem cell-derived extracellular vesicles reduce senescence and extend health span in mouse models of aging. Aging Cell. 2021;20(4):e13337. doi:10.1111/acel.13337
59. Malekpour K, Hazrati A, Zahar M, et al. The potential use of mesenchymal stem cells and their derived exosomes for orthopedic diseases treatment. Stem Cell Rev Rep. 2022;18(3):933–951. doi:10.1007/s12015-021-10185-z
60. Zhang Y, Hao Z, Wang P, et al. Exosomes from human umbilical cord mesenchymal stem cells enhance fracture healing through HIF-1alpha-mediated promotion of angiogenesis in a rat model of stabilized fracture. Cell Prolif. 2019;52(2):e12570. doi:10.1111/cpr.12570
61. Gong L, Chen B, Zhang J, et al. Human ESC-sEVs alleviate age-related bone loss by rejuvenating senescent bone marrow-derived mesenchymal stem cells. J Extracell Vesicles. 2020;9(1):1800971. doi:10.1080/20013078.2020.1800971
62. Chan CKF, Gulati GS, Sinha R, et al. Identification of the Human Skeletal Stem Cell. Cell. 2018;175(1):43–56 e21. doi:10.1016/j.cell.2018.07.029
63. Tu J, Huo Z, Yu Y, et al. Hereditary retinoblastoma iPSC model reveals aberrant spliceosome function driving bone malignancies. Proc Natl Acad Sci USA. 2022;119(16):e2117857119. doi:10.1073/pnas.2117857119
64. Spitzhorn LS, Megges M, Wruck W, et al. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res Ther. 2019;10(1):100. doi:10.1186/s13287-019-1209-x
65. Cheng Z, Peng HL, Zhang R, Fu XM, Zhang GS. Rejuvenation of cardiac tissue developed from reprogrammed aged somatic cells. Rejuvenation Res. 2017;20(5):389–400. doi:10.1089/rej.2017.1930
66. Sun Y, Zhang W, Li X. Induced pluripotent stem cell-derived mesenchymal stem cells deliver exogenous miR-105-5p via small extracellular vesicles to rejuvenate senescent nucleus pulposus cells and attenuate intervertebral disc degeneration. Stem Cell Res Ther. 2021;12(1):286. doi:10.1186/s13287-021-02362-1
67. Wang F, Cai F, Shi R, Wang XH, Wu XT. Aging and age related stresses: a senescence mechanism of intervertebral disc degeneration. Osteoarthritis Cartilage. 2016;24(3):398–408. doi:10.1016/j.joca.2015.09.019
68. Chao-Yang G, Peng C, Hai-Hong Z. Roles of NLRP3 inflammasome in intervertebral disc degeneration. Osteoarthritis Cartilage. 2021;29(6):793–801. doi:10.1016/j.joca.2021.02.204
69. Wang W, Jing X, Du T, et al. Iron overload promotes intervertebral disc degeneration via inducing oxidative stress and ferroptosis in endplate chondrocytes. Free Radic Biol Med. 2022;190:234–246. doi:10.1016/j.freeradbiomed.2022.08.018
70. Kamali A, Ziadlou R, Lang G, et al. Small molecule-based treatment approaches for intervertebral disc degeneration: current options and future directions. Theranostics. 2021;11(1):27–47. doi:10.7150/thno.48987
71. Hao Y, Ren Z, Yu L, et al. p300 arrests intervertebral disc degeneration by regulating the FOXO3/Sirt1/Wnt/beta-catenin axis. Aging Cell. 2022;21(8):e13677. doi:10.1111/acel.13677
72. Cheng X, Zhang G, Zhang L, et al. Mesenchymal stem cells deliver exogenous miR-21 via exosomes to inhibit nucleus pulposus cell apoptosis and reduce intervertebral disc degeneration. J Cell Mol Med. 2018;22(1):261–276. doi:10.1111/jcmm.13316
73. Xia C, Zeng Z, Fang B, et al. Mesenchymal stem cell-derived exosomes ameliorate intervertebral disc degeneration via anti-oxidant and anti-inflammatory effects. Free Radic Biol Med. 2019;143:1–15. doi:10.1016/j.freeradbiomed.2019.07.026
74. Liao Z, Luo R, Li G, et al. Exosomes from mesenchymal stem cells modulate endoplasmic reticulum stress to protect against nucleus pulposus cell death and ameliorate intervertebral disc degeneration in vivo. Theranostics. 2019;9(14):4084–4100. doi:10.7150/thno.33638
75. Lu K, Li HY, Yang K, et al. Exosomes as potential alternatives to stem cell therapy for intervertebral disc degeneration: in-vitro study on exosomes in interaction of nucleus pulposus cells and bone marrow mesenchymal stem cells. Stem Cell Res Ther. 2017;8(1):108. doi:10.1186/s13287-017-0563-9
76. Chen Y, Tang L. Stem cell senescence: the obstacle of the treatment of degenerative disk disease. Curr Stem Cell Res Ther. 2019;14(8):654–668. doi:10.2174/1574888X14666190906163253
77. Hu YC, Zhang XB, Lin MQ, et al. Nanoscale treatment of intervertebral disc degeneration: mesenchymal stem cell exosome transplantation. Curr Stem Cell Res Ther. 2023;18(2):163–173. doi:10.2174/1574888X17666220422093103
78. Ma M, Cui G, Liu Y, et al. Mesenchymal stem cell-derived extracellular vesicles, osteoimmunology and orthopedic diseases. PeerJ. 2023;11:e14677. doi:10.7717/peerj.14677
79. Xia Y, Yang R, Hou Y, et al. Application of mesenchymal stem cell-derived exosomes from different sources in intervertebral disc degeneration. Front Bioeng Biotechnol. 2022;10:1019437. doi:10.3389/fbioe.2022.1019437
80. Sun Y, Li X, Yang X, Chen B, Zhang W. Small extracellular vesicles derived from adipocytes attenuate intervertebral disc degeneration in rats by rejuvenating senescent nucleus pulposus cells and endplate cells by delivering exogenous NAMPT. Oxid Med Cell Longev. 2021;2021:9955448. doi:10.1155/2021/9955448
81. Guan M, Liu C, Zheng Q, et al. Exosome-laden injectable self-healing hydrogel based on quaternized chitosan and oxidized starch attenuates disc degeneration by suppressing nucleus pulposus senescence. Int J Biol Macromol. 2023;232:123479. doi:10.1016/j.ijbiomac.2023.123479
82. Bhujel B, Shin HE, Choi DJ, Han I. Mesenchymal stem cell-derived exosomes and intervertebral disc regeneration: review. Int J Mol Sci. 2022;23(13). doi:10.3390/ijms23137306
83. Krut Z, Pelled G, Gazit D, Gazit Z. Stem cells and exosomes: new therapies for intervertebral disc degeneration. Cells. 2021;10(9):2241. doi:10.3390/cells10092241
84. Dai Z, Xia C, Zhao T, et al. Platelet-derived extracellular vesicles ameliorate intervertebral disc degeneration by alleviating mitochondrial dysfunction. Mater Today Bio. 2023;18:100512. doi:10.1016/j.mtbio.2022.100512
85. Liang W, Han B, Hai Y, Sun D, Yin P. Mechanism of action of mesenchymal stem cell-derived exosomes in the intervertebral disc degeneration treatment and bone repair and regeneration. Front Cell Dev Biol. 2021;9:833840. doi:10.3389/fcell.2021.833840
86. Xiang XN, Zhu SY, He HC, Yu X, Xu Y, He CQ. Mesenchymal stromal cell-based therapy for cartilage regeneration in knee osteoarthritis. Stem Cell Res Ther. 2022;13(1):14. doi:10.1186/s13287-021-02689-9
87. Greene MA, Loeser RF. Aging-related inflammation in osteoarthritis. Osteoarthritis Cartilage. 2015;23(11):1966–1971. doi:10.1016/j.joca.2015.01.008
88. Lieberthal J, Sambamurthy N, Scanzello CR. Inflammation in joint injury and post-traumatic osteoarthritis. Osteoarthritis Cartilage. 2015;23(11):1825–1834. doi:10.1016/j.joca.2015.08.015
89. Sanchez-Lopez E, Coras R, Torres A, Lane NE, Guma M. Synovial inflammation in osteoarthritis progression. Nat Rev Rheumatol. 2022;18(5):258–275. doi:10.1038/s41584-022-00749-9
90. Goldring MB, Otero M. Inflammation in osteoarthritis. Curr Opin Rheumatol. 2011;23(5):471–478. doi:10.1097/BOR.0b013e328349c2b1
91. Li T, Peng J, Li Q, Shu Y, Zhu P, Hao L. The mechanism and role of ADAMTS protein family in osteoarthritis. Biomolecules. 2022;12:7.
92. Cosenza S, Ruiz M, Toupet K, Jorgensen C, Noel D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep. 2017;7(1):16214. doi:10.1038/s41598-017-15376-8
93. Jin Y, Xu M, Zhu H, et al. Therapeutic effects of bone marrow mesenchymal stem cells-derived exosomes on osteoarthritis. J Cell Mol Med. 2021;25(19):9281–9294. doi:10.1111/jcmm.16860
94. Tao SC, Huang JY, Gao Y, et al. Small extracellular vesicles in combination with sleep-related circRNA3503: a targeted therapeutic agent with injectable thermosensitive hydrogel to prevent osteoarthritis. Bioact Mater. 2021;6(12):4455–4469. doi:10.1016/j.bioactmat.2021.04.031
95. Withrow J, Murphy C, Liu Y, Hunter M, Fulzele S, Hamrick MW. Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis. Arthritis Res Ther. 2016;18(1):286. doi:10.1186/s13075-016-1178-8
96. Zhang S, Teo KYW, Chuah SJ, Lai RC, Lim SK, Toh WS. MSC exosomes alleviate temporomandibular joint osteoarthritis by attenuating inflammation and restoring matrix homeostasis. Biomaterials. 2019;200:35–47. doi:10.1016/j.biomaterials.2019.02.006
97. Sun Y, Zhao J, Wu Q, et al. Chondrogenic primed extracellular vesicles activate miR-455/SOX11/FOXO axis for cartilage regeneration and osteoarthritis treatment. NPJ Regen Med. 2022;7(1):53. doi:10.1038/s41536-022-00250-7
98. Liu Z, Zhuang Y, Fang L, Yuan C, Wang X, Lin K. Breakthrough of extracellular vesicles in pathogenesis, diagnosis and treatment of osteoarthritis. Bioact Mater. 2023;22:423–452. doi:10.1016/j.bioactmat.2022.10.012
99. Mundy GR. Osteoporosis and inflammation. Nutr Rev. 2007;65(12 Pt 2):S147–51. doi:10.1301/nr.2007.dec.S147-S151
100. Bellavia D, Salamanna F, Raimondi L, et al. Deregulated miRNAs in osteoporosis: effects in bone metastasis. Cell Mol Life Sci. 2019;76(19):3723–3744. doi:10.1007/s00018-019-03162-w
101. Zhu L, Tang Y, Li XY, et al. Osteoclast-mediated bone resorption is controlled by a compensatory network of secreted and membrane-tethered metalloproteinases. Sci Transl Med. 2020;12(529). doi:10.1126/scitranslmed.aaw6143
102. Liu JH, Chen CY, Liu ZZ, et al. Extracellular vesicles from child gut microbiota enter into bone to preserve bone mass and strength. Adv Sci. 2021;8(9):2004831. doi:10.1002/advs.202004831
103. Wang Y, Yao J, Cai L, et al. Bone-targeted extracellular vesicles from mesenchymal stem cells for osteoporosis therapy. Int J Nanomedicine. 2020;15:7967–7977. doi:10.2147/IJN.S263756
104. Wang ZX, Luo ZW, Li FX, et al. Aged bone matrix-derived extracellular vesicles as a messenger for calcification paradox. Nat Commun. 2022;13(1):1453. doi:10.1038/s41467-022-29191-x
105. Zhang W, Huang P, Lin J, Zeng H. The role of extracellular vesicles in osteoporosis: a scoping review. Membranes. 2022;12:3.
106. Yang Z, Liu X, Zhao F, et al. Bioactive glass nanoparticles inhibit osteoclast differentiation and osteoporotic bone loss by activating lncRNA NRON expression in the extracellular vesicles derived from bone marrow mesenchymal stem cells. Biomaterials. 2022;283:121438. doi:10.1016/j.biomaterials.2022.121438
107. Li Y, Wang F, Liang H, et al. Efficacy of mesenchymal stem cell transplantation therapy for type 1 and type 2 diabetes mellitus: a meta-analysis. Stem Cell Res Ther. 2021;12(1):273. doi:10.1186/s13287-021-02342-5
108. Keshtkar S, Azarpira N, Ghahremani MH. Mesenchymal stem cell-derived extracellular vesicles: novel frontiers in regenerative medicine. Stem Cell Res Ther. 2018;9(1):63. doi:10.1186/s13287-018-0791-7
109. Qiu G, Zheng G, Ge M, et al. Functional proteins of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res Ther. 2019;10(1):359. doi:10.1186/s13287-019-1484-6
110. Xia L, Zhang C, Lv N, et al. AdMSC-derived exosomes alleviate acute lung injury via transferring mitochondrial component to improve homeostasis of alveolar macrophages. Theranostics. 2022;12(6):2928–2947. doi:10.7150/thno.69533
111. Xiong M, Zhang Q, Hu W, et al. The novel mechanisms and applications of exosomes in dermatology and cutaneous medical aesthetics. Pharmacol Res. 2021;166:105490. doi:10.1016/j.phrs.2021.105490
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.