Back to Journals » Journal of Inflammation Research » Volume 19
Gut Microbiota Mitigates Chronic Itch and Cutaneous Inflammation in DNFB-Induced Atopic Dermatitis Mice
Authors Xiong Z ![]() , Dong X, Yuan Y, Lu L, Deng X
, Dong X, Yuan Y, Lu L, Deng X
Received 15 October 2025
Accepted for publication 31 January 2026
Published 6 February 2026 Volume 2026:19 573709
DOI https://doi.org/10.2147/JIR.S573709
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Anish R. Maskey
Zichen Xiong, Xinming Dong, Yi Yuan, Lin Lu, Xueting Deng
Medical Centre for Digestive Diseases, The Second Affiliated Hospital of Nanjing Medical University, Nanjing, Jiangsu, 210011, People’s Republic of China
Correspondence: Xueting Deng, Medical Centre for Digestive Diseases, The Second Affiliated Hospital of Nanjing Medical University, Nanjing, Jiangsu, 210011, People’s Republic of China, Email [email protected]
Background: Chronic itch is the most prominent symptom of atopic dermatitis (AD), which severely impacts the quality of life of patients and persists even after medication. Gut microbiota dysbiosis is considered to contribute to AD, however, the roles of gut microbiota in the modulation of chronic pruriceptive processing currently remain unclear. The present study aimed to elucidate the potential regulatory role of the gut microbiota in AD-associated chronic itch.
Methods: In this study, the 1-fluoro-2,4-dinitrobenzene (DNFB)-induced mouse model of AD-associated chronic itch was established. Differences in gut microbiota composition between model and healthy controls were analyzed using high-throughput 16S rRNA gene sequencing. In addition, we performed oral fecal microbiota transplantation (FMT) from model mice to antibiotic cocktail-treated healthy mice and observed whether they could induce itch behavior. Furthermore, feces from healthy mice were transplanted into model mice to evaluate their effects on itch symptoms and skin inflammation.
Results: The DNFB induced significantly itch behaviors and an altered gut microbiota composition. The gut microbiota from chronic itch mice through oral administration could induce itch behaviors in antibiotic cocktail-treated healthy mice. While, oral FMT from healthy mice to chronic itch mice not only significantly alleviated scratching behavior but also ameliorated skin damage and inflammation. Following FMT administration from healthy donors, remarkable alterations were observed in the metabolomic profiles of mice with DNFB-induced chronic itch.
Conclusion: These findings highlight the potential link between gut microbiota dysbiosis and chronic itching in AD, suggesting that targeting the gut microbiota may be a therapeutic strategy for chronic itch.
Keywords: gut microbiota, chronic itch, atopic dermatitis, fecal microbiota transplantation, inflammation
Introduction
Atopic dermatitis (AD) is a recurrent, chronic, non-infectious inflammatory skin disease characterized by persistent itching of the skin.1–3 The prevalence of AD is approximately 20% in children and about 10% in adults.4,5 According to clinicians, persistent itching is the chief complaint of 87% of AD patients and the first symptom of AD, regardless of its specific phenotype. Despite treatment, it remains poorly relieved, disrupting daily activities, causing insomnia, and significantly impairing quality of life.6–10 Although not fully understood, it is widely believed that genetic disorders, epidermal barrier defects, immune response alterations, and disturbances in the skin microbiota balance are the most important mechanisms underlying the development of AD.11–13 However, the relationship between gut microbiota and chronic itch in AD has yet to be fully elucidated, leaving key mechanisms incompletely understood. Moreover, the development of safe and effective antipruritic therapies has proven challenging, resulting in inadequate management of chronic itch for many AD patients in clinical settings.
Studies have reported the presence of gut microbiota dysbiosis in AD.14,15 Infants and children with AD exhibit significantly lower gut microbiota diversity compared to healthy controls, which is closely associated with an increased risk of developing AD.16,17 The gut microbiota in early life is associated with the age of onset, severity, flare-ups, remission, and phenotype of AD. In addition to the diversity of the gut microbiota itself, interactions between specific gut microbiota, the established immune system, and the coordination between the gut microbiota and the host may play a role in the development of AD.18 Furthermore, metabolites of the gut microbiota, namely short-chain fatty acids, have been found to be significantly reduced in AD patients.19,20 These metabolites can circulate through the bloodstream and affect inflammation in distant parts of the body.
Chronic itching is a key feature of AD. While several studies have been conducted on the role of gut microbiota in AD, the exact impact of gut microbiota on chronic itching in AD remains unclear. Investigating the effect of gut microbiota on AD related itch phenotypes and the possible molecular mechanisms could contribute to the development of new treatment strategies for alleviating chronic itching in AD patients.
Recent studies increasingly indicate that the gut microbiota may be involved in the onset and modulation of chronic itch. For example, significant alterations in gut microbial composition have been observed in a mouse model of chronic itch.21 Clinical evidence further suggests that gut dysbiosis often accompanies chronic itch in multiple diseases and is associated with itch severity.22 In addition, studies in patients with liver cirrhosis complicated by chronic itch have demonstrated that those with more severe itch exhibit significantly different fecal and salivary microbial profiles compared with patients with mild itch, and these alterations are not directly concordant with the decompensated status of cirrhosis.23 These findings suggest that specific alterations in the gut microbiota may be independent of overall disease severity and closely associated with the development of the itch phenotype. Notably, other studies have reported that patients with primary biliary cholangitis accompanied by pruritus do not exhibit significant differences in baseline gut microbiota composition compared with control groups.24,25 This suggests that whether the gut microbiota can serve as a primary driving factor of chronic itch requires further investigation, and the causal relationship between the two remains unclear.
The present study sought to comprehensively examine the involvement of the gut microbiota in AD-related chronic itch. Accordingly, gut microbiota composition in AD mice was initially compared with that of healthy controls using 16S rRNA amplicon sequencing to characterize global structural differences. Subsequently, antibiotic-mediated gut microbiota depletion and fecal microbiota transplantation (FMT) were employed to further validate the causal effects of microbiota alterations on the chronic itch phenotype. Given that 1-fluoro-2,4-dinitrobenzene (DNFB) has been widely used to establish experimental models of atopic dermatitis and has been shown to reliably and reproducibly induce chronic itch–related behaviors, a DNFB-induced AD mouse model was selected for this study.26,27 Overall, this design advances insight into gut microbiota–mediated mechanisms underlying AD-related chronic itch and supports microbiome-based therapeutic strategies.
Methods
Materials
Vancomycin hydrochloride (V105495), ampicillin sodium (A105483), neomycin sulfate (N109017), and metronidazole (M109874) were obtained from Aladdin (Shanghai, China). These antibiotics were used for microbiota depletion prior to fecal microbiota transplantation. 1-fluoro-2,4-dinitrobenzene (DNFB) and olive oil were purchased from MCE (USA). DNFB was applied to induce atopic dermatitis-like symptoms in mice, with olive oil and acetone serving as its solvents during the sensitization and challenge phases. Acetone was sourced from China National Pharmaceutical Group Chemical Reagent Co., Ltd. (China).
Animals
Specific-pathogen-free 8-week-old male BALB/c mice (weight 19–23 grams) were acquired from Zhejiang Vital River Laboratory Animal Technology Co., Ltd. All animals were housed in a 23±2°C environment with a 12-hour light/dark cycle, and were given free access to food and water. The experiments strictly followed the guidelines of the Nanjing Medical University Animal Welfare and Use Committee (license number IACUC-2406065) and the Animal Ethics Review Committee.
Before the start of the experiments, mice were acclimatized for at least 7 days. They were then randomly assigned to different experimental and control groups using a computer-generated random number sequence. To minimize potential confounding factors, animals from each group were randomly distributed across cages, and experimental procedures and measurements were performed in a randomized order. The experimental unit was defined as an individual mouse. Inclusion and exclusion criteria were predefined during the experimental design stage. All healthy mice with successful model establishment were included in the final analysis; mice were to be excluded if unexpected death, severe infection, or failure of model induction occurred during the experiment. No animals were excluded in the present study. A total of 64 mice were used in this study and were randomly assigned to the experimental groups, with eight mice per group. Sample size determination was based on previously published literature and the experimental experience of our research group with similar animal models, to ensure reliable and reproducible results within ethical constraints. Group sizes varied (n = 6–8 per group) depending on the experimental assay, and the exact number of animals used for each analysis is indicated in the corresponding figure legends. The experimental allocation was performed by a separate investigator who was not involved in treatment administration, outcome assessment, or data analysis. To minimize observer bias, behavioral assessments, histological analyses, and data analyses were all performed in a blinded manner by investigators who were unaware of the group allocations.
Establishment of the Chronic Itching Model and Evaluation of Scratching Behavior
According to the schematic, DNFB dissolved in a 4:1 mixture of acetone and olive oil was repeatedly applied to the shaved skin of the mice to establish the chronic itching model.27 Two days before the application of DNFB, hair removal was performed on the mouse’s abdomen and the dorsal neck region, approximately 2 cm2 in area. During the sensitization phase, 50 μL of 0.5% DNFB solution was applied to the depilated abdominal skin. On the fifth day after sensitization, the challenge phase began, and 30 μL of 0.25% DNFB solution was applied to the depilated neck and back skin every other day (on days 6, 9, 11, and 13). The control group mice underwent the same procedure, but only solvent was applied.
Behavioral assessments were conducted on experimental days 1, 5, 7, 10, 12, and 14. Before each assessment, mice were individually placed in transparent plastic cages and allowed to acclimatize to the environment for 30 mins. After the acclimation period, spontaneous scratching behavior was continuously recorded for 1 h using a video monitoring system. Quantitative measures of scratching behavior included the number of scratching events and the total scratching duration. Scratching behavior was defined using a standardized operational criterion: a single scratching bout was defined as a mouse lifting a hind paw and repeatedly scratching the DNFB-treated skin area until the hind paw was withdrawn and returned to the floor or transitioned to another behavior, such as paw licking. If the interval between two consecutive scratching movements was ≥1 s, they were counted as two independent scratching events. All behavioral data were analyzed based on video playback by investigators who were blinded to the group allocations.
Fecal Collection and 16S rRNA Gene Sequencing Analysis
Fresh fecal samples were collected, placed in sterile centrifuge tubes, immediately frozen, and stored at −80°C. DNA extraction from fecal samples and 16S rRNA analysis were conducted by Shenzhen BGI Genomics Co., Ltd. PCR amplification was used to amplify the bacterial 16S rRNA gene targeting the V3-V4 region. Alpha-diversity analysis was evaluated using the Shannon entropy index, Simpson’s diversity index, Pielou’s evenness index, and observed species count. Beta-diversity was determined through principal coordinate analysis (PCoA). Statistical significance was tested by analysis of similarities (ANOSIM). A heatmap cluster analysis at the genus level was performed based on bacterial abundance data. The analysis was conducted using the pheatmap package in R (version 4.3.2), with the Euclidean distance metric and the complete linkage method. The top 10 most abundant genera were selected, and their average relative abundances along with the statistical significance of differences between groups were displayed.
Non-Targeted Metabolomics Detection Method
Metabolomics detection was performed using the Q Exactive HF high-resolution mass spectrometer from BGI Genomics (Shenzhen, China), based on liquid chromatography-tandem mass spectrometry (LC-MS/MS) technology. To maximize metabolite detection coverage, data acquisition was performed in both positive (ESI⁺) and negative (ESI−) electrospray ionization modes, with mass spectrometry data collected in full-scan mode. To ensure data quality and system stability, quality control samples were periodically inserted at fixed intervals throughout the sample run to monitor instrument response reproducibility and data consistency. Raw mass spectrometry data were processed uniformly using Compound Discoverer 3.1 software. The analysis workflow included peak extraction, peak alignment, peak integration, and noise reduction, with all parameter settings following the software’s recommended standard procedures and kept consistent across all samples. Metabolite identification was based on accurate mass, retention time, and MS/MS fragmentation patterns, and involved cross-referencing multiple databases, including the BGI in-house standard library (BGI Library), mzCloud, and ChemSpider (integrating HMDB, KEGG, and LipidMaps). Peak extraction and metabolite identification were performed on the raw mass spectrometry data to obtain peak areas and identification results. The exported results were further preprocessed using metaX to generate compounds and quantitative values suitable for formal analysis.
Antibiotic Cocktail Therapy and Fecal Microbiota Transplantation
The antibiotic cocktail (ABX) was composed of vancomycin hydrochloride (0.5 g/L), ampicillin sodium (1 g/L), neomycin sulfate (1 g/L), and metronidazole (1 g/L), which were mixed into the drinking water of the mice at the specified concentrations. Fresh fecal samples from mice were collected prior to antibiotic treatment. One gram of feces was placed in a sterilized screw-cap EP tube containing 10 mL of sterile PBS, vortexed thoroughly, and centrifuged at 4°C and 2000 r.p.m. for 5 minutes. The pellet was discarded, and the supernatant was transferred to a new sterile tube and stored at −80°C for future use. Mice were continuously treated with ABX for 14 days. A 1-day washout period (ABX cessation) was applied before the first FMT. The experimental group of mice was then orally gavaged with the supernatant or PBS for 7 consecutive days, at a dose of 100 μL per mouse.
Histological and Immunohistochemical Analyses
Skin lesion tissues and colon tissues from each group of mice were fixed overnight in 4% paraformaldehyde, followed by routine dehydration, paraffin embedding, and sectioning. Paraffin sections were stained with hematoxylin and eosin (H&E) according to the manufacturer’s instructions. Quantitative analysis of epidermal thickness was performed using ImageJ software. Epidermal thickness was defined as the vertical distance from the basal layer to the top of the stratum corneum and was measured in micrometers (μm). For each section, three measurement points were randomly selected for multipoint assessment, and the mean value was calculated as the epidermal thickness.
Skin tissue sections were stained with toluidine blue for mast cells and processed for immunohistochemistry to evaluate the expression and spatial distribution of IL-4, IL-13, and IL-31. Mast cell toluidine blue staining was performed according to the manufacturer’s instructions to evaluate mast cell infiltration. For immunohistochemistry, sections were deparaffinized, rehydrated, subjected to heat-induced antigen retrieval, and blocked with serum, followed by incubation with primary antibodies (IL-4: Proteintech, 84758-5-RR; IL-13: Affinity, DF6813; IL-31: Omnimabs, OM281946), then with the corresponding secondary antibodies and developed.
Evaluation of Skin Lesions
Clinical skin scores were recorded based on four criteria: erythema/bleeding, scarring/dryness, edema, and scaling/erosion. Each criterion was scored from 0 to 3 based on lesion severity (0: none; 1: mild; 2: moderate; 3: severe, with higher scores indicating more severe symptoms). The assessment of each item was based on clinical features such as the extent and intensity of lesions, and the presence of exudation or structural changes in the skin. The clinical skin score was then calculated as the sum of the four individual scores, yielding a total score ranging from 0 to 12.
Enzyme-Linked Immunosorbent Assay Analysis
Lipopolysaccharides (LPS), IL-6, IL-1β, TNF-α, and IgE were measured using ELISA kits (obtained from Xinbosheng Biotechnology Co., Ltd., Shenzhen, China, and Huamei Bioengineering Co., Ltd., Wuhan, China). All experimental procedures were performed strictly in accordance with the manufacturers’ instructions. Optical density (OD) at 450 nm was measured using a microplate reader (Agilent, USA) to determine analyte concentrations.
Statistical Analysis
All data were analyzed using GraphPad Prism (version 9.5.0) and expressed as mean ±SEM. Comparisons between two independent experimental groups were performed using a two-tailed unpaired Student’s t-test. Comparisons among three or more groups were performed using one-way ANOVA, followed by post-hoc correction using Fisher’s least significant difference method. Multiple group comparisons with two independent variables were performed using two-way ANOVA, followed by post-hoc testing with Tukey’s honest significant difference method. The sample size for each experiment and detailed statistical analysis methods is provided in the figure legends. A p-value < 0.05 was considered statistically significant. Data were first assessed for normality using the Shapiro–Wilk test and for homogeneity of variance using Levene’s test. If the assumptions were not met, appropriate non-parametric tests were applied (eg, Mann–Whitney U-test or Kruskal–Wallis test).
Results
Successful Establishment of the DNFB-Induced Chronic Itch Model in Mice
To establish a mouse model of DNFB-induced chronic itch, a standardized protocol was used involving skin sensitization followed by repeated DNFB challenges (Figure 1A). Behavioral assessments were conducted at designated time points to evaluate pruritic responses, such as scratching frequency and total scratching duration. Compared with control mice (NC), DNFB-treated mice exhibited significantly increased scratching frequency and prolonged scratching duration (Figure 1B). In addition, DNFB-induced chronic itch was associated with prominent dermatitis-like skin lesions, including erythema, dryness, excoriation, and scaling, as illustrated by representative images (Figure 1C). Quantitative analysis of clinical skin scores revealed significantly greater dermatitis severity in the DNFB group compared to controls (Figure 1D). H&E staining showed marked epidermal hyperplasia and inflammatory cell infiltration in the DNFB group (Figure 1E), along with significantly increased epidermal thickness compared to the control group (Figure 1F). There was no statistically significant difference on focal epithelial damage or submucosal inflammatory cell infiltration in the colorectal tissues between the DNFB group and NC group (Figure 1G).
 |
Figure 1 Successful establishment of the DNFB-induced chronic itch model in mice. (A) Schematic illustration of the experimental protocol used to establish DNFB-induced chronic itch in mice. (B) Behavioral assessment schedule evaluating pruritic responses: (i) number of scratching bouts; (ii) total scratching time. (C) Representative images showing dermatitis-like skin lesions in mice. (D) Quantitative analysis of dermatitis severity based on clinical skin scores across groups. (E and F) Histological examination of skin tissue (scale bar = 200 μm) (E) and the quantification of epidermal thickness in each group (F). (G) H&E-stained colon tissue sections from each group (scale bar = 200 μm). *P < 0.05, **P < 0.01; n = 8. |
Alterations in Gut Microbiota Composition and LPS Levels in DNFB-Induced Chronic Itch Mice
To explore the differences in gut microbiota composition between chronic itching mice and healthy wild-type mice, fecal samples were collected from the mice and sequenced to analyze the characteristics of their gut microbiota. As shown in Figure 2A, the species richness, Shannon diversity index, Simpson diversity index, and Pielou’s evenness index were decreased in the chronic itching mice, suggesting that alpha-diversity analysis was significant differences between the two groups. Subsequently, beta-diversity analysis, which quantifies the similarity or difference between groups, revealed a significant difference in the gut microbiota composition between chronic itching mice and wild-type mice (Figure 2B).
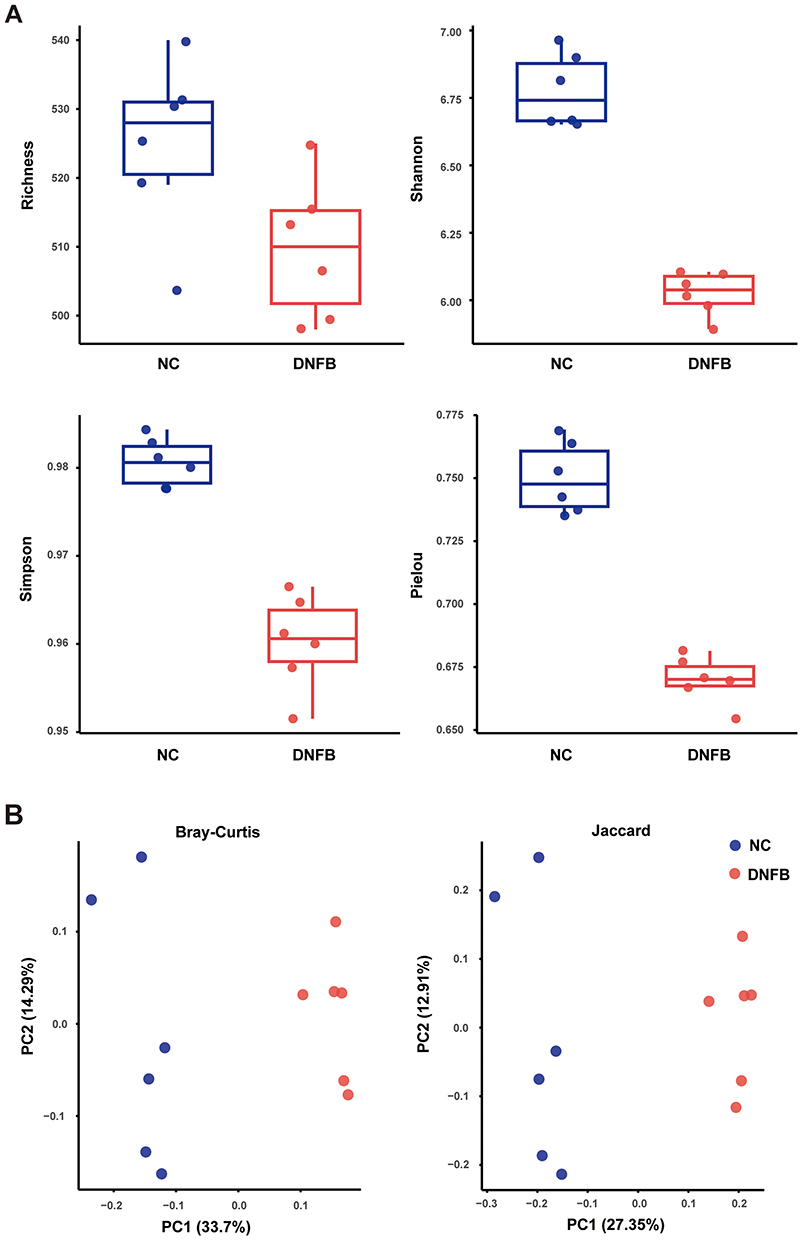 |
Figure 2 Analysis of alpha-diversity and beta-diversity of gut microbiota in chronic itching mice and their littermate wild-type controls. (A) Alpha-diversity analysis: Comparison of (i) species richness, (ii) Shannon entropy, (iii) Simpson diversity index, and (iv) Pielou’s evenness index between the control group (NC) and chronic itching (DNFB) mice. (B) Beta-diversity analysis: Principal coordinate analysis (PCoA) based on Bray-Curtis and Jaccard distances illustrates gut microbiota structural differences between NC and DNFB groups. n = 6. |
Bray-Curtis matrix analysis showed significant differences in dominant species between the two groups. Further analysis of the Jaccard index revealed significant differences between chronic itching mice and their littermate wild-type controls. Comparison of genus-level relative abundance across groups revealed that the relative abundances of Prevotellamassilia (P < 0.05) was decreased in DNFB-treated mice compared to controls, while Flintibacter (P < 0.01), Anaerotignum (P < 0.01), and Schaedlerella (P < 0.01) were increased (Figure 3A and B). Meanwhile, the serum LPS concentration was significantly increased in DNFB-treated mice (Figure 3C).
 |
Figure 3 Differential abundance analysis of microbial taxa and serum LPS levels between controls and mice with chronic itch. (A) Heatmap of genus-level microbial abundance: Hierarchical clustering-based heatmap illustrating genus-level microbial profiles in NC and DNFB mice. Rows represent genera ordered by hierarchical clustering, and columns represent individual samples. Color scale: yellow indicates low abundance, red indicates high abundance. (B) Bar plot of differentially abundant key genera: The top 10 genera by abundance were selected to display group-wise mean relative abundances and corresponding statistical significance. (C) Serum LPS levels in NC and DNFB-treated mice. **P < 0.01; n = 6. |
Fecal Microbiota Transplantation from Mice with DNFB-Induced Chronic Itch Can Induce Chronic Itching in Healthy Mice
To further investigate the relationship between the development of chronic itching and gut microbiota, we performed FMT from DNFB-induced mice into healthy mice to explore the influence of abnormal gut microbiota on chronic itching phenotype. As shown in Figure 4A, mice were first pretreated with an antibiotic cocktail for 7 days to deplete the gut microbiota. Following this, they received normal drinking water for 1 day to permit recovery of the gut microenvironment. Finally, a 7-day FMT was conducted, after which scratching frequency and duration were evaluated. The findings indicated that compared with the control group (NC+ABX), FMT from DNFB-treated mice (NC+ABX+DNFB_F) induced an increased number of scratches and a longer scratching duration (P < 0.05, Figure 4B). Notably, no skin damage was observed in the NC+ABX+DNFB_F group (Supplementary Figure S1).
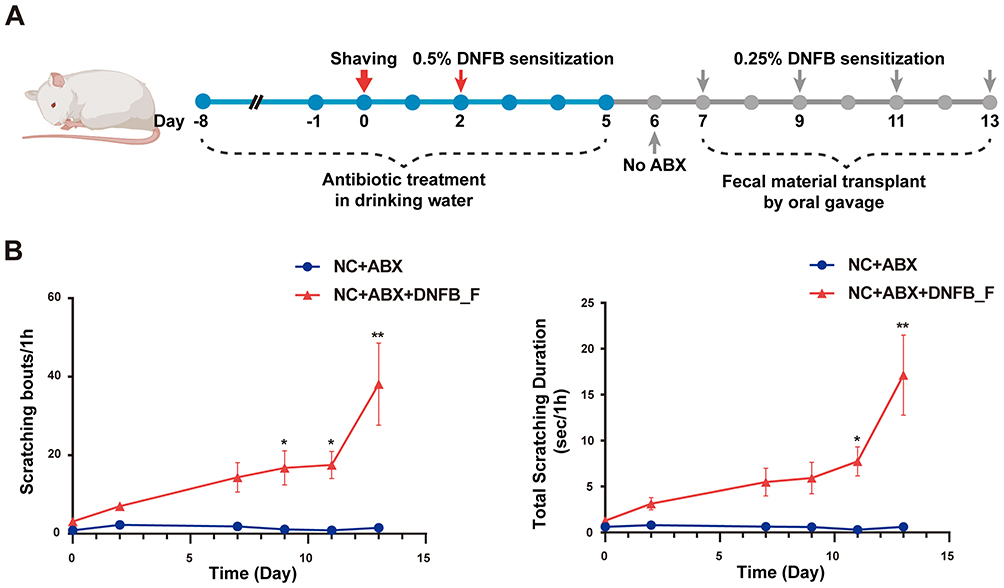 |
Figure 4 Effect of fecal microbiota transplantation from chronic itch to healthy mice on the chronic itching phenotype. (A) Experimental design diagram. Both groups of mice were treated with broad-spectrum antibiotics (ABX) in drinking water for 7 days, followed by FMT for 7 days with a 1-day interval. The experimental group received FMT derived from DNFB-induced chronic itch mice (NC+ABX+DNFB_F), whereas the control group received saline (NC+ABX). (B) Time points for behavioral assessment of scratching: (i) Number of scratching bouts; (ii) Duration of scratching. *P < 0.05, **P < 0.01; n = 8. |
Fecal Microbiota Transplantation from Healthy Mice Alleviates Scratching Behavior and Inflammatory Responses in DNFB-Induced Chronic Itch Mice
To explore whether remodeling the gut microbiota in DNFB mice has a regulatory effect on chronic itching, we performed FMT from healthy mice into DNFB-induced chronic itch mice (DNFB+ABX+NC_F). The experimental procedure was consistent with the previous protocol: recipient mice received a 14-day antibiotic pretreatment, followed by a 1-day interval, after which the 7-day FMT was commenced. The results showed that FMT from healthy mice (DNFB+ABX+NC_F) significantly reduced the number of scratches and the duration of scratching in DNFB-induced chronic itch mice (P < 0.05, Figure 5). To further evaluate the effects of FMT derived from healthy mice on skin pathological damage and local inflammatory responses in DNFB-induced chronic itch mice, histological and immunohistochemical analyses were performed on skin tissues from different groups. The results showed that, compared with the DNFB group, gut microbiota transplantation from healthy donors significantly ameliorated skin lesions and reduced epidermal thickening and inflammatory infiltration (Figure 6A–D).
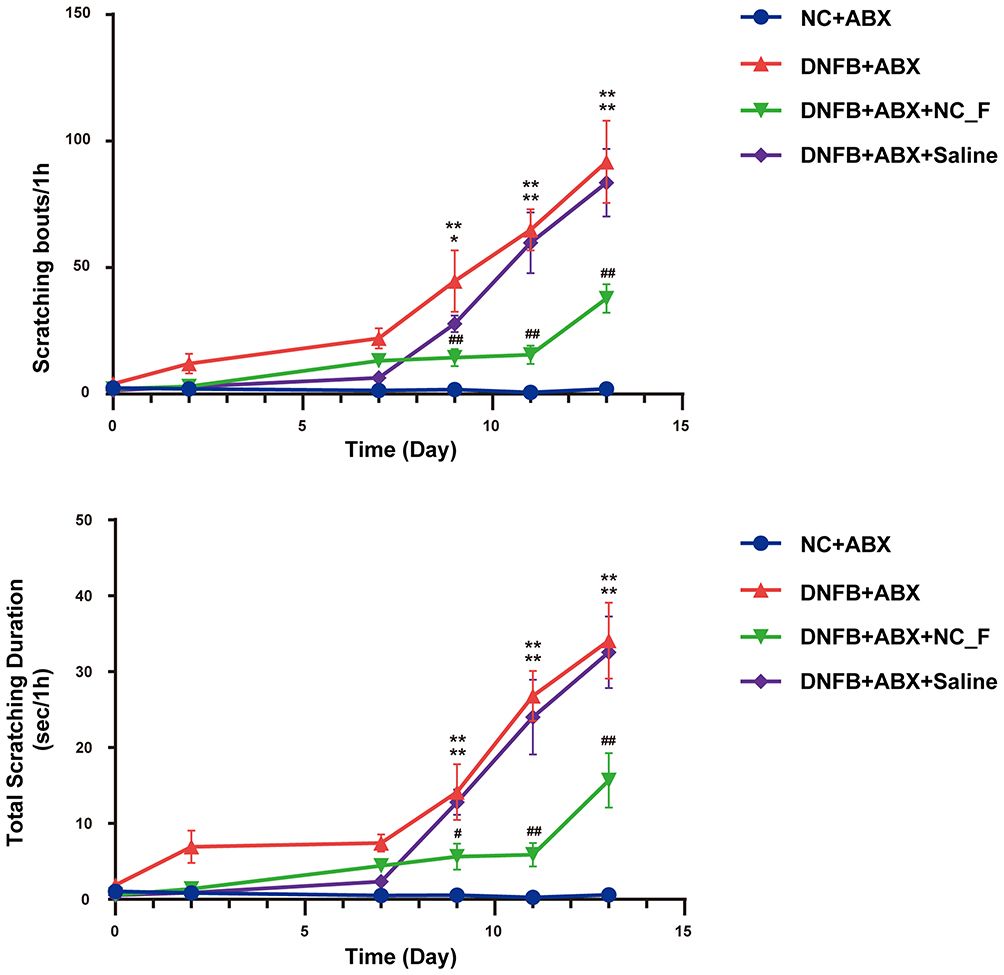 |
Figure 5 Fecal microbiota transplantation from healthy donor mice alleviates chronic itching in DNFB-induced chronic itch mice. *P < 0.05, **P < 0.01, compared with controls (NC+ABX); #P < 0.05, ##P < 0.01, compared with mice with chronic itch (DNFB+ABX); n = 8. |
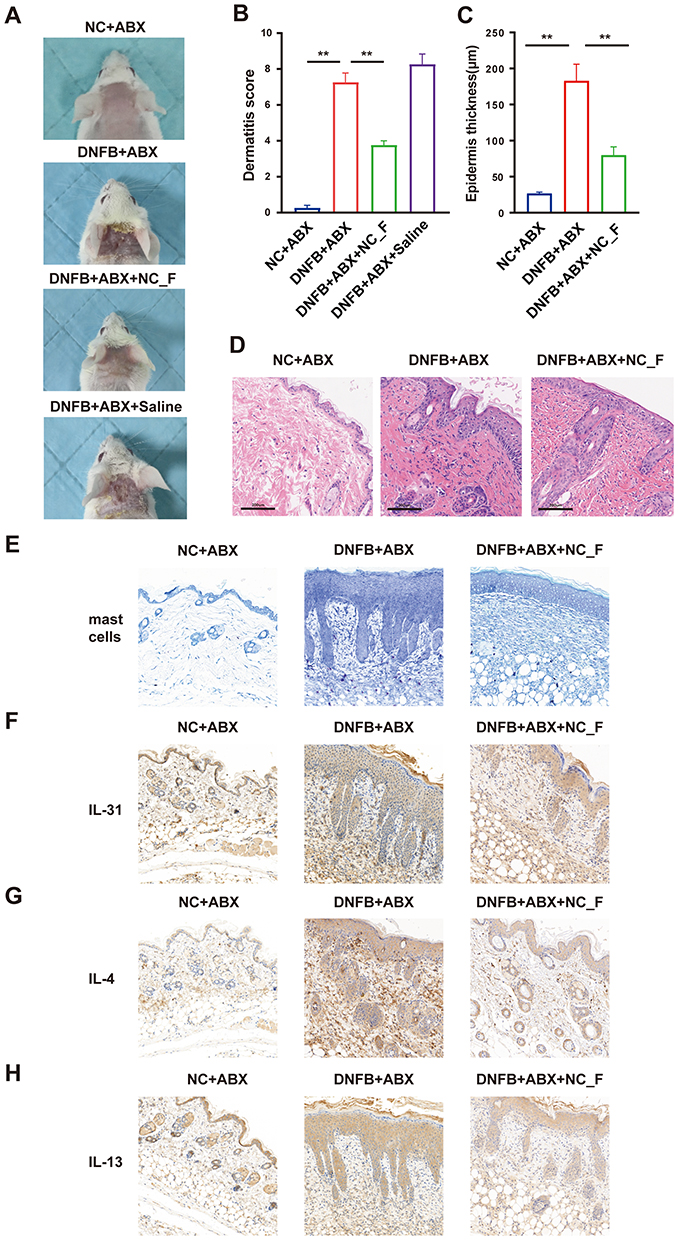 |
Figure 6 Fecal microbiota transplantation from healthy donors attenuates skin lesions in mouse model of chronic itch. (A) Representative images of skin lesions; (B) Statistical summary of dermatitis scores. (C and D) Statistical analysis of epidermal thickness (C) and histopathological examination of skin tissues (D). (E) Representative staining of mast cells in skin tissues. (F–H) immunohistochemical staining of IL-31, IL-4, and IL-13 in skin tissues. *P < 0.05, **P < 0.01; n = 8. |
In light of the critical involvement of mast cell infiltration and various pruritogenic and type 2 inflammatory mediators in chronic itch pathogenesis, we next assessed related indicators in the skin. The results showed an increase in mast cell accumulation in the skin of DNFB-induced chronic itch mice, which was attenuated after transplantation of microbiota from healthy mice (Figure 6E). Meanwhile, immunohistochemical staining revealed that IL-31, IL-4 and IL-13 expressions were upregulated in the skin of the DNFB group, which was efficiently inhibited by FMT from healthy mice (Figure 6F–H), suggesting that restore gut microbiota has potential to improve skin structural impairment and inhibits local inflammatory in chronic itch.
As shown in Figure 7, compared with the control group, serum levels of IgE, IL-6, IL-1β, and TNF-α were significantly elevated in mice with DNFB-induced chronic itch, indicating a pronounced allergic immune response and systemic inflammatory activation. FMT from healthy mice markedly decreased serum levels of these inflammatory markers compared to the mice with DNFB-induced chronic itch. These results suggested that partially restored the gut microbiota may contribute to alleviate systemic inflammation in chronic itch.
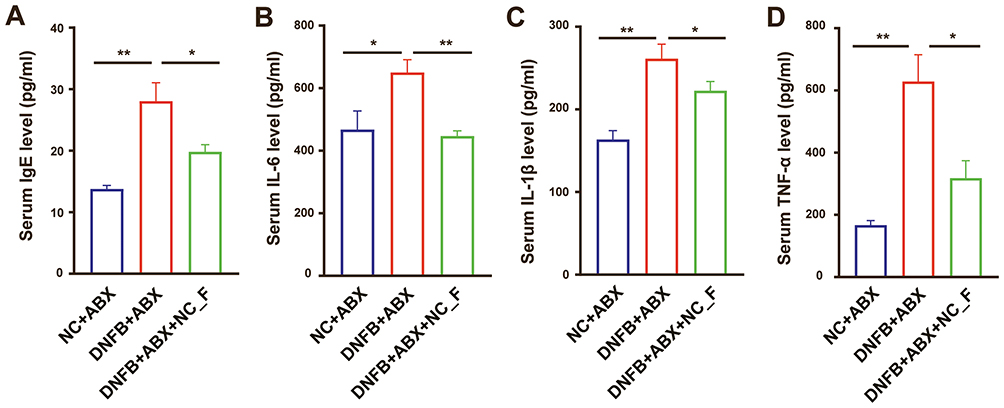 |
Figure 7 Fecal microbiota transplantation from healthy mice attenuates inflammatory responses in mice with chronic itch AD. (A–D) Serum levels of IgE, IL-6, IL-1β, and TNF-α measured by ELISA in each group of mice. *P < 0.05, **P < 0.01; n = 8. |
The Impact of FMT on the Metabolomics of Mice with DNFB-Induced Chronic Itch
Furthermore, we observed the untargeted metabolomics differences between chronic itch mice receiving fecal transplantation from healthy wild-type donor mice (DNFB+ABX+NC_F) and the chronic itch mice. In Figure 8, the results showed that the base peak chromatograms of quality control (QC) samples were highly overlapping, and correlation analysis demonstrated strong consistency among QC samples. In addition, the coefficients of variation (CV) for most metabolites in QC samples were below 30%, indicating good analytical reproducibility. Overall, these findings confirmed the robustness of the Liquid Chromatography–Mass Spectrometry platform and support the reliability of downstream metabolomic analyses. Untargeted metabolomics analysis was used to select differential metabolites between the two groups based on a t-test (criteria: p < 0.05, Variable Importance in Projection (VIP) ≥ 1, Fold Change ≥ 1.2 or ≤ 0.83), and a volcano plot of these metabolites was generated. Simultaneously, a volcano plot of differential metabolites was generated based on VIP and fold change (Figure 9A). A total of 254 differential metabolites were identified, with 51 metabolites upregulated and 203 metabolites downregulated in chronic itch mice receiving fecal microbiota transplantation compared to the chronic itch group, based on multiple testing correction (Figure 9B). Pathway enrichment analysis of differential metabolites based on the KEGG database indicated significant enrichment in several metabolic pathways, including linoleic acid metabolism, carbon metabolism, central carbon metabolism in cancer, bile secretion, and biosynthesis of amino acids (Figure 9C). Among them, the linoleic acid metabolism pathway demonstrated the most pronounced enrichment, coupled with a high enrichment factor (RichFactor), indicating substantial metabolic alterations in this pathway following FMT treatment. To further elucidate the directional trends of these pathways, a differential abundance score-based analysis was performed. As shown in Figure 9D, the linoleic acid metabolism, carbon metabolism, and amino acid biosynthesis as well as cofactor biosynthesis pathways were generally downregulated after treatment, whereas the bile secretion pathway was significantly upregulated in response to the intervention.
 |
Figure 8 Metabolomic quality control (QC) analysis of a chronic itch mouse model modulated by fecal microbiota transplantation from healthy donors. (A) Base peak chromatograms: The base peak chromatograms of QC samples in both positive and negative ion modes were highly overlapping, indicating good stability of instrument response intensity and retention time. (B) QC correlation analysis: Spearman correlation analysis among QC samples revealed overall correlation coefficients approaching 1, indicating minimal system error and good data reproducibility. (C) Coefficients of variation of metabolites in QC samples: The distribution of metabolite CV in QC samples showed that most metabolites had CV values below 30%, indicating good analytical stability and reliability. |
 |
Figure 9 Fecal microbiota transplantation from healthy donors modulates the metabolomic profile in a mouse model of chronic itch. (A) Volcano plot of differential metabolites: red dots indicate significantly upregulated metabolites (P < 0.05 and FC ≥ 2), blue dots indicate significantly downregulated metabolites (P < 0.05 and FC ≤ 0.5). Gray dots represent metabolites with no significant change. (B) Bar chart of differential metabolites between groups. (C) Bubble chart of pathway enrichment analysis for differential metabolites: bubble size indicates the number of metabolites, and color gradient reflects the p-value. Larger and darker bubbles represent pathways with stronger enrichment and higher statistical significance. (D) Differential pathway analysis based on DA scores: The Y-axis lists the metabolic pathways, and the X-axis shows the DA scores, which reflect the relative metabolic trends in the treatment group compared to the chronic itch mice model (positive values indicate pathway upregulation, while negative values indicate downregulation). |
Discussion
Our study reveals the critical role of the gut microbiota in AD-associated chronic itching. Transplantation of fecal microbiota from DNFB-induced chronic itch mice into antibiotic-treated healthy mice induced chronic itching in the recipients. Furthermore, FMT from healthy donors into mice with DNFB-induced chronic itch effectively alleviated pathological scratching, epidermal thickening, and reduced inflammation in the recipient mice. We conclude that altered gut microbiota has a role in the pathogenesis of AD-associated chronic pruritus, highlighting it as a promising target for therapeutic interventions.
Gut microbiota, comprising the vast community of commensal microorganisms residing in the gastrointestinal tract, has been shown to play critical roles in numerous physiological and pathological processes. The role of gut microbiota in the pathophysiology of AD has gained increasing attention.28,29 Previous studies have reported dysbiosis of the gut microbiota in AD patients.30–32 The link between gut microbiota dysbiosis and chronic itch in AD is supported by our experiments. Transplantation of gut microbiota from AD-induced chronic itch model mice into healthy recipient mice successfully provoked significant scratching behavior in the recipients. This finding suggests that AD-associated pruritus is not merely a localized cutaneous symptom but is systemically regulated by gut microbial ecology. Dysregulated microbiota alone can act as a pathogenic factor, independent of primary skin inflammation, to drive itch-related behavior.
Interesting, transferring gut microbiota from DNFB-treated donors to healthy recipient mice did not result in obvious skin lesions. This observation suggests that without direct chemical irritation of the skin, fecal microbiota transplantation alone cannot entirely reproduce the morphological features of atopic dermatitis. In DNFB-induced models, the development of skin lesions relies on several factors, such as repeated hapten exposure, localized disruption of the skin barrier, and continuous activation of skin immune responses. Although gut microbiota-mediated phenotypic transfer is partial and does not fully recapitulate the complete disease state, we observed a significant increase in scratching behavior following fecal microbiota transplantation. This result aligns with existing literature and suggests that specific gut microbial alterations may operate independently of overall disease severity and are closely linked to the emergence of the itch phenotype. These findings are consistent with the concept that the gut microbiota can serve as a primary driver in the pathogenesis of chronic itch.23,24
LPS, a surface glycolipid of Gram-negative bacteria, is widely recognized for its role in acute inflammatory responses. Beyond this acute context, LPS is also implicated in the development of chronic inflammation. In such chronic settings, the primary source of LPS is typically not an external infection, but rather an increase in endogenous production, which is often influenced by the gut microbiota. Our results showed a significant elevation in serum LPS levels in DNFB-treated mice, suggesting that microbiota dysbiosis may contribute to the initiation of systemic inflammatory responses. However, the functional contribution of specific microbial taxa to LPS elevation and the mechanistic links to chronic itch remain to be fully elucidated, warranting further experimental investigation.
Moreover, in the DNFB-induced disease model with ABX administration, compared to the mice treated with saline (DNFB+ABX+saline), mice treated with FMT from healthy mice (DNFB+ABX+NC_F) showed a significant reduction in scratching behavior, as well as the inflammatory responses. The findings suggest that the amelioration of scratching behavior is mainly due to FMT-mediated recovery of gut microbiota function. In this study, the results provide experimental evidence suggesting that modulation of the gut microbiota may contribute to the attenuation of certain pathological features observed in DNFB-induced chronic itch. This supports further investigation into its potential as a targeted therapeutic strategy. It is noteworthy that microbiota depletion can prevent the development chronic painful conditions with nerve injury, chemotherapy and diabetic neuropathy,33 while the ABX effects on chronic itch remain more explores. Importantly, antibiotic-mediated microbiota depletion may influence baseline itch-related responses and immune homeostasis, which could in turn affect the sensitivity of the DNFB-induced chronic itch model. Although these baseline effects were not specifically dissected in the present study, they warrant further investigation in future work.
Notably, linoleic acid metabolism exhibited the most significant enrichment, with a relatively high enrichment factor. Furthermore, DA score-based directional analysis revealed a marked reduction in metabolites associated with this pathway following FMT treatment, suggesting a potential downregulation of linoleic acid metabolism in response to microbial intervention. Linoleic acid is a polyunsaturated fatty acid of the n-6 series, and its metabolites (such as 13-HODE and 9-HODE) have been shown to induce inflammatory pain by activating TRPV1 ion channels. Given that both pain and itch are transmitted via overlapping TRP channels, these findings suggest that linoleic acid metabolites may drive itch through similar mechanisms;34,35 however, this hypothesis was not directly tested in the present study. In addition, previous studies have shown that dysregulated linoleic acid metabolism is associated with impaired skin barrier integrity and increased transepidermal water loss, which may exacerbate cutaneous inflammation and pruritus.36 The alterations in linoleic acid-related metabolites observed after FMT in this study suggested that the gut microbiota may alleviate AD-associated chronic itch by modulating linoleic acid metabolism. It should be noted that the exploration of the underlying mechanisms in the present study remains at a preliminary stage and requires further validation and clarification. Further mechanistic studies are warranted to elucidate the downstream molecular pathways linking microbial regulation of lipid metabolism with itch sensation and cutaneous inflammation.
Conclusion
This study demonstrates that the gut microbiota plays a crucial regulatory role in AD-related chronic itching. Our experimental results indicate that dysbiosis or restoration of gut microbiota composition significantly influences both scratching behavior and inflammation responses in mice with DNFB-induced chronic itch. These findings provide a new perspective for elucidating the mechanisms underlying chronic itching and may serve as a theoretical foundation for developing microbiota-targeted therapeutic strategies.
Abbreviations
ABX, antibiotic cocktail; AD, atopic dermatitis; CV, coefficients of variation; DA-score, differential abundance-score; DNFB, 1-fluoro-2,4-dinitrobenzene; FMT, fecal microbiota transplantation; LPS, lipopolysaccharide; PCoA, principal coordinate analysis; QC, coefficients of variation.
Data Sharing Statement
The data that support the finding of this study are available from the corresponding author upon reasonable request.
Ethics Approval Statements
The study was designed, conducted, and reported in accordance with the ARRIVE guidelines. All animal procedures strictly followed the regulations of the Animal Welfare and Use Committee of Nanjing Medical University (license No. IACUC-2406065) and the Animal Ethics Review Committee. Mice were anesthetized with isoflurane to minimize pain and distress, and euthanized by carbon dioxide (CO2) inhalation in accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals (2020).
Acknowledgments
We thank the Second Affiliated Hospital of Nanjing Medical University for providing the experimental platform, all participating researchers, and the animal subjects used in this study, as well as the support provided by the National Natural Science Foundation of China.
Author Contributions
Xueting Deng contributed to the conceptualization and design of the study, supervision, and project administration; Zichen Xiong contributed to methodology, investigation, data curation, formal analysis, visualization, and drafting of the original manuscript; Xinming Dong contributed to methodology and investigation; Yi Yuan contributed to formal analysis and data interpretation; Lin Lu contributed to investigation; Xueting Deng and Zichen Xiong contributed to reviewing and editing the manuscript. All authors took part in either drafting or revising the manuscript, gave final approval of the version to be published, have agreed on the journal to which the article has been submitted, and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This work was supported by the National Natural Science Foundation of China (grant number 82171222).
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Asher MI, Montefort S, Björkstén B, et al. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC phases one and three repeat multicountry cross-sectional surveys. Lancet. 2006;368(9537):733–17. doi:10.1016/s0140-6736(06)69283-0
2. Arrais M, Lulua O, Quifica F, Rosado-Pinto J, Gama JMR, Taborda-Barata L. Prevalence of asthma, allergic rhinitis and eczema in 6-7-year-old schoolchildren from Luanda, Angola. Allergol Immunopathol. 2019;47(6):523–534. doi:10.1016/j.aller.2018.12.002
3. Mandlik DS, Mandlik SK. Atopic dermatitis: new insight into the etiology, pathogenesis, diagnosis and novel treatment strategies. Immunopharmacol Immunotoxicol. 2021;43(2):105–125. doi:10.1080/08923973.2021.1889583
4. Mallol J, Crane J, von Mutius E, Odhiambo J, Keil U, Stewart A. The international study of asthma and allergies in childhood (ISAAC) phase three: a global synthesis. Allergol Immunopathol. 2013;41(2):73–85. doi:10.1016/j.aller.2012.03.001
5. Leung TF, Wong GW. The Asian side of asthma and allergy. Curr Opin Allergy Clin Immunol. 2008;8(5):384–390. doi:10.1097/ACI.0b013e3283103a8e
6. Chrostowska-Plak D, Reich A, Szepietowski JC. Relationship between itch and psychological status of patients with atopic dermatitis. J Eur Acad Dermatol Venereol. 2013;27(2):e239–42. doi:10.1111/j.1468-3083.2012.04578.x
7. Silverberg JI, Gelfand JM, Margolis DJ, et al. Patient burden and quality of life in atopic dermatitis in US adults: a population-based cross-sectional study. Ann Allergy Asthma Immunol. 2018;121(3):340–347. doi:10.1016/j.anai.2018.07.006
8. Sroka-Tomaszewska J, Trzeciak M. Molecular mechanisms of atopic dermatitis pathogenesis. Int J Mol Sci. 2021;22(8):4130. doi:10.3390/ijms22084130
9. Hussein MN, Abu-Raghif AR, Ridha-Salman H, et al. Flavonoids from Conyza Canadensis mitigates CDNB-induced mouse model of atopic dermatitis. Comp Clin Pathol. 2025;34(5):987–1010. doi:10.1007/s00580-025-03709-3
10. Habbas AH, Abu-Raghif AR, Ridha-Salman H, Hussein MN. Therapeutic effect of bosentan on 2, 4-dinitrochlorobenzene (DNCB)-induced atopic dermatitis mouse model. Archives of Dermatological Res. 2025;317(1):436. doi:10.1007/s00403-025-03955-z
11. Tominaga M, Takamori K. Peripheral itch sensitization in atopic dermatitis. Allergol Int. 2022;71(3):265–277. doi:10.1016/j.alit.2022.04.003
12. Langan SM, Irvine AD, Weidinger S. Atopic dermatitis. Lancet. 2020;396(10247):345–360. doi:10.1016/s0140-6736(20)31286-1
13. Czarnowicki T, He H, Krueger JG, Guttman-Yassky E. Atopic dermatitis endotypes and implications for targeted therapeutics. J Allergy Clin Immunol. 2019;143(1):1–11. doi:10.1016/j.jaci.2018.10.032
14. De Pessemier B, Grine L, Debaere M, Maes A, Paetzold B, Callewaert C. Gut-skin axis: current knowledge of the interrelationship between microbial dysbiosis and skin conditions. Microorganisms. 2021;9(2). doi:10.3390/microorganisms9020353
15. Penders J, Thijs C, van den Brandt PA, et al. Gut microbiota composition and development of atopic manifestations in infancy: the KOALA birth cohort study. Gut. 2007;56(5):661–667. doi:10.1136/gut.2006.100164
16. Hu C, van Meel ER, Medina-Gomez C, et al. A population-based study on associations of stool microbiota with atopic diseases in school-age children. J Allergy Clin Immunol. 2021;148(2):612–620. doi:10.1016/j.jaci.2021.04.001
17. Abrahamsson TR, Jakobsson HE, Andersson AF, Björkstén B, Engstrand L, Jenmalm MC. Low diversity of the gut microbiota in infants with atopic eczema. J Allergy Clin Immunol. 2012;129(2):434–40, 440.e1–2. doi:10.1016/j.jaci.2011.10.025
18. Lee SY, Lee E, Park YM, Hong SJ. Microbiome in the gut-skin axis in atopic dermatitis. Allergy Asthma Immunol Res. 2018;10(4):354–362. doi:10.4168/aair.2018.10.4.354
19. Park DH, Kim JW, Park HJ, Hahm DH. Comparative analysis of the microbiome across the gut-skin axis in atopic dermatitis. Int J Mol Sci. 2021;22(8). doi:10.3390/ijms22084228
20. Nylund L, Nermes M, Isolauri E, Salminen S, de Vos WM, Satokari R. Severity of atopic disease inversely correlates with intestinal microbiota diversity and butyrate-producing bacteria. Allergy. 2015;70(2):241–244. doi:10.1111/all.12549
21. Li Y, Zhang W, Sun T, et al. The role of gut microbiota in chronic itch-evoked novel object recognition-related cognitive dysfunction in mice. Front Med Lausanne. 2021;8:616489. doi:10.3389/fmed.2021.616489
22. Jiao B, Cao X, Zhang C, et al. Alterations of the gut microbiota in patients with postherpetic neuralgia. AMB Express. 2023;13(1):108. doi:10.1186/s13568-023-01614-y
23. Bajaj JS, Fagan A, Sikaroodi M, et al. Alterations in skin microbiomes of patients with cirrhosis. Clin Gastroenterol Hepatol. 2019;17(12):2581–2591.e15. doi:10.1016/j.cgh.2019.03.028
24. Hegade VS, Pechlivanis A, McDonald JAK, et al. Autotaxin, bile acid profile and effect of ileal bile acid transporter inhibition in primary biliary cholangitis patients with pruritus. Liver Int. 2019;39(5):967–975. doi:10.1111/liv.14069
25. Hegade VS, Kendrick SF, Dobbins RL, et al. Effect of ileal bile acid transporter inhibitor GSK2330672 on pruritus in primary biliary cholangitis: a double-blind, randomised, placebo-controlled, crossover, phase 2a study. Lancet. 2017;389(10074):1114–1123. doi:10.1016/s0140-6736(17)30319-7
26. Qu Y, Wang G, Sun X, Wang K. Inhibition of the warm temperature-activated Ca(2+)-permeable transient receptor potential vanilloid TRPV3 channel attenuates atopic dermatitis. Mol Pharmacol. 2019;96(3):393–400. doi:10.1124/mol.119.116962
27. Wang Y, Tan L, Jiao K, et al. Scutellarein attenuates atopic dermatitis by selectively inhibiting transient receptor potential vanilloid 3 channels. Br J Pharmacol. 2022;179(20):4792–4808. doi:10.1111/bph.15913
28. Kong HH, Oh J, Deming C, et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012;22(5):850–859. doi:10.1101/gr.131029.111
29. Penders J, Gerhold K, Stobberingh EE, et al. Establishment of the intestinal microbiota and its role for atopic dermatitis in early childhood. J Allergy Clin Immunol. 2013;132(3):601–607.e8. doi:10.1016/j.jaci.2013.05.043
30. Fang Z, Li L, Zhang H, Zhao J, Lu W, Chen W. Gut microbiota, probiotics, and their interactions in prevention and treatment of atopic dermatitis: a review. Front Immunol. 2021;12:720393. doi:10.3389/fimmu.2021.720393
31. de Andrade P, Maria ESJ, Carregaro V, et al. Efficacy of probiotics in children and adolescents with atopic dermatitis: a randomized, double-blind, placebo-controlled study. Front Nutr. 2021;8:833666. doi:10.3389/fnut.2021.833666
32. Xu B, Ling S, Xu X, et al. A new formulation of probiotics attenuates calcipotriol-induced dermatitis by inducing regulatory dendritic cells. Front Immunol. 2021;12:775018. doi:10.3389/fimmu.2021.775018
33. Ma P, Mo R, Liao H, et al. Gut microbiota depletion by antibiotics ameliorates somatic neuropathic pain induced by nerve injury, chemotherapy, and diabetes in mice. J Neuroinflammation. 2022;19(1):169. doi:10.1186/s12974-022-02523-w
34. Patwardhan AM, Akopian AN, Ruparel NB, et al. Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents. J Clin Invest. 2010;120(5):1617–1626. doi:10.1172/jci41678
35. Alsalem M, Wong A, Millns P, et al. The contribution of the endogenous TRPV1 ligands 9-HODE and 13-HODE to nociceptive processing and their role in peripheral inflammatory pain mechanisms. Br J Pharmacol. 2013;168(8):1961–1974. doi:10.1111/bph.12092
36. Elias PM, Brown BE, Ziboh VA. The permeability barrier in essential fatty acid deficiency: evidence for a direct role for linoleic acid in barrier function. J Invest Dermatol. 1980;74(4):230–233. doi:10.1111/1523-1747.ep12541775
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.