Back to Journals » Journal of Inflammation Research » Volume 18
Gut Microbiota and Metabolite Changes Induced by Tacrolimus: Implications for Skin Transplant Immunology in Mice
Authors Wang J, Zhang X, Cui C, Li M, Xie Z, Yang L, Ding D, Li X ![]() , Zhao M
, Zhao M ![]()
Received 9 May 2025
Accepted for publication 30 September 2025
Published 9 October 2025 Volume 2025:18 Pages 14059—14073
DOI https://doi.org/10.2147/JIR.S539452
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Junpeng Wang,1,* Xiaofan Zhang,2,* Cuiyun Cui,3 Mengjun Li,4 Zixuan Xie,4 Lei Yang,5 Degang Ding,1 Xin Li,4 Ming Zhao6
1Department of Urology, Henan Provincial People’s Hospital, Zhengzhou University People’s Hospital, Zhengzhou, People’s Republic of China; 2Medical Discipline and Research Office, The Second Affiliated Hospital of Zhengzhou University, Zhengzhou, People’s Republic of China; 3Department of Blood Transfusion, Henan Provincial People’s Hospital, Zhengzhou University People’s Hospital, Zhengzhou, People’s Republic of China; 4Department of Pathophysiology, School of Basic Medical Sciences, Zhengzhou University, Zhengzhou, People’s Republic of China; 5Department of Pathology, Yale University, New Haven, CT, USA; 6Department of Organ Transplantation, Zhujiang Hospital, Southern Medical University, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xin Li, Email [email protected] Ming Zhao, Email [email protected]
Background: Tacrolimus is the most widely used immunosuppressive therapy in solid organ transplantation. However, whether it can inhibit transplant graft rejection by altering the composition and metabolism of gut microbiota remains unclear.
Methods: In this study, a skin transplantation mouse model was established to explore the effects of tacrolimus on gut microbiota and its metabolites. Additionally, we investigated the protective effect and potential mechanism of feces from mice treated with tacrolimus on skin allografts.
Results: Tacrolimus did not significantly affect gut microbiota α-diversity but altered β-diversity, with specific changes in microbial composition. LEfSe analysis identified 19 microbial taxa with reduced and 12 with elevated relative abundance in the Tac group (mice treated with tacrolimus) compared to the Ctrl group (mice with no treatment). Metabolomic analysis identified 33 differential fecal metabolites (17 upregulated and 16 downregulated) in the Tac group compared to the Ctrl group. FMT from tacrolimus-treated mice significantly prolonged skin allograft survival, reduced inflammatory cell infiltration, and improved graft histopathology. This protective effect was associated with increased Treg cell proportions and decreased Th17 cell proportions in draining lymph nodes and mesenteric lymph node.
Conclusion: Overall, our data may provide a basis for establishing gut microbiota-based therapies for allograft rejection.
Keywords: graft rejection, transplantation, intestinal flora, tacrolimus, intestinal metabolic profile
Introduction
Over the past half-century, organ transplantation has become the standard of care for patients with advanced organ failure. Although immunosuppressive drugs have significantly reduced the incidence of early allogeneic graft rejection, both chronic allogeneic graft rejection and drug-related side effects (primarily life-threatening infections, organ toxicity, diabetes mellitus, hypertension, and malignant tumors) remain major threats to the survival of both grafts and recipients.1,2
Tacrolimus, a calcineurin phosphatase inhibitor, has been widely used in immunosuppressive therapy for solid organ transplants since 1994 and is utilized in more than 93% of transplant recipients.3 Tacrolimus triggers immunosuppression by interacting with immunophilins such as FK506-binding protein (FKBP) to form the FKBP-FK506 complex, which in turn inhibits calcium-regulated neural phosphatase in T cells, prevents IL-2 transcription, and inhibits T lymphocyte activation.4 However, the oral bioavailability of tacrolimus varies from 4% to 89%, with an average of about 25%, and it has a narrow therapeutic index.5 Additionally, there is a poor correlation between tacrolimus dosage and whole blood concentration, resulting in significant individual variability in the dosage required to achieve target blood drug levels.6–9 Therefore, close monitoring of the dosage of tacrolimus is necessary to maintain blood drug concentration. Excessive doses of tacrolimus may elevate the risk of infection and malignancy, whereas insufficient doses can precipitate graft rejection, which in turn increases the risk of graft loss and reduces patient survival.10,11
Previous research indicates that changes occur in the gut microbiota after organ transplantation,12–16 suggesting a close relationship and mutual influence between organ transplantation, gut microbiota, and the immune system. The gut microbiota consists of trillions of microorganisms that engage in many bidirectional regulatory functions with the host and can influence host immune function through bacterial components and metabolites.17,18 Emerging evidence highlights that gut microbiota dysbiosis is closely linked to both acute and chronic rejection. Skin-restricted commensal Staphylococcus epidermidis was shown to accelerate skin graft rejection.19 For cardiac allografts, Bifidobacterium pseudolongum exerts a protective effect on allograft survival by enhancing the production of homeostatic CCL19 and anti-inflammatory IL-10, whereas Desulfovibrio desulfuricans promotes increased IL-6 and TNF-α production and reduces allograft survival.20 Additionally, Pseudomonas airway infection was reported to induces marked lung neutrophilia in a G-CSF-dependent manner, which could promote CD4⁺ T cell activation, thus triggering acute cellular graft rejection.21 Notably, gut microbial metabolites are found to be key mediators of host-microbiota crosstalk in transplantation immunity. Alistipes could delay skin graft rejection by the presence of anti-inflammatory metabolite sulfobacin B.22,23 Butyrate, a short-chain fatty acid (SCFA), may improve transplant outcomes via its Treg-inducing properties.24 Moreover, in a kidney transplant model, administration of SCFA acetate has been demonstrated to prolong allograft survival in a Treg- and GPR43-dependent manner.25 It is reported that tacrolimus is a macrolide antibiotic that is likely to affect bacteria.26 Recent research findings indicate that tacrolimus can induce changes in the composition and function of the intestinal microbiota.27,28 Combining tacrolimus with the fecal microbiota transplantation can enhance the development of T cells in the colonic mucosa, draining lymph nodes, and peripheral blood, and increase the survival of skin allografts.27 However, it is not yet known whether tacrolimus can exert an inhibitory effect on transplant rejection by altering the composition and metabolism of intestinal microbiota.
Therefore, in this study, we investigated the impact of tacrolimus on the intestinal microbiota in mice by establishing a mouse skin transplant model. We further performed a non-targeted metabolomics assay to examine the effects of tacrolimus on intestinal metabolites. Additionally, we analyzed immune cells in the peripheral blood, spleen, mesenteric lymph nodes, and draining lymph nodes to reveal the potential roles and mechanisms of the intestinal microbiota and its metabolites in transplant rejection.
Materials and Methods
Establishment of a Murine Skin Graft Model
A mouse allograft skin graft rejection model was established, as previously reported, using female BALB/c mice as donors and female C57BL/6 mice as recipients.29,30 Given that the density of dendritic cells (DCs) in trunk skin is much higher than that in tail skin, which enables trunk skin to elicit a more robust transplant rejection and thus makes it a more reliable rejection model, we used mouse dorsal skin to establish the skin transplantation model in the present study.31 All animals were housed in a facility at Zhengzhou University that was free from specific pathogens. All groups of mice were fed the same diet. The donor mice were intraperitoneally anesthetized with 10% chloral hydrate (40 μL/10 g), and the hair on their backs was removed and disinfected. A 1 cm × 1 cm full-thickness skin graft was excised from the back of the donor mice, followed by the removal of the subcutaneous fascia and fat. The recipient mice were anesthetized as described above, and the hair on their backs was also removed and disinfected. A skin area of approximately 1 cm × 1 cm was excised from the back of the recipient mice, and the resulting wound served as the graft bed. The donor skin was retrieved and laid flat on the graft bed. The skin graft was sutured to the edge of the graft bed using 5–0 silk sutures. After disinfection, the wound was bandaged and secured with a sterile wound dressing. The transplanted mice were placed in a clean cage on top of a microwavable heating pad until they completely woke up from anesthesia. The transplanted skin of the recipient mice was closely observed starting from the 5th day after transplantation, and the observation indexes mainly included the color gloss, the presence of bruises, the hardness, and the presence of skin shedding. If the grafted skin of the recipient mice developed scabs, hardened and atrophied, turned completely black, or even fell off, it could be judged as graft rejection. Conversely, if the grafted skin of the recipient mice was red, soft, or showed hair growth, it could be considered that the grafted skin had survived or had become tolerant. This research received approval from the Institutional Animal Care and Use Committee at Zhengzhou University, with experiments conducted in strict compliance with the Guidelines for Laboratory Animal Care.
Sequencing of the 16S rRNA Gene of Mouse Intestinal Flora
Mice were divided into two groups: the tacrolimus-treated group and the normal saline-treated group. Each group comprised four mice. The tacrolimus gavage dose was 10 mg/kg/day (volume of 0.5 mL), while the saline gavage dose was 0.5 mL, with both groups being gavaged once daily. Mice in the same experimental group were cohoused in specific pathogen-free (SPF) cages. After 7 consecutive days of gavage followed by a 2-day rest, fecal samples from both groups of mice were collected. The abdomen of each mouse was gently massaged to facilitate defecation, and fresh fecal pellets were collected into sterile centrifuge tubes. These samples were then stored at −80°C for subsequent use. 16S rRNA gene sequencing was performed using the Illumina MiSeq platform. Experiments were designed according to the published literature.27
LC-MS Untargeted Metabolomics of Intestinal Metabolites in Mice
The non-targeted metabolomics assay, based on LC-MS, was performed using an Agilent 1290 Infinity LC ultra-high-performance liquid chromatography system coupled with a Triple TOF 6600 mass spectrometer.
Fecal Transplantation
To prepare bacterial solution for fecal transplantation, we used tacrolimus solution (10 mg/kg/day) and saline for gavage in mice. Each group comprised four mice. After 7 consecutive days of gavage followed by a 2-day rest, the feces of the two groups were collected separately. The feces were diluted with sterile saline (approximately 12.5 mg of feces was diluted with 1 mL of sterile saline), as previously reported.27 The mice were treated with 500 μL/d of fecal bacterial solution by gavage.
Therapeutic Protocols for Skin Graft Model Mice
The mice were randomly divided into four groups as follows: (1) Ctrl group: skin grafting was performed on day 0, and no treatment was performed after skin grafting; (2) Tac group: skin grafting was performed on day 0 and tacrolimus (10 mg/kg/day) gavage treatment was performed on day 1 after grafting for 7 consecutive days; (3) FMT group: The mice were treated with fecal bacterial solution from those gavaged with saline one week before skin grafting. Skin grafting was performed on day 0. FMT continued on day 1 for 7 consecutive days; (4) Tac FMT group: The mice were treated with fecal bacterial solution from those gavaged with tacrolimus solution one week prior to skin grafting. Skin grafting was performed on day 0. FMT continued on day 1 for 7 consecutive days. Each group comprised five mice. The experiments were independently repeated 3 times to confirm result consistency.
Detection of Immune Cells by Flow Cytometry
The flow cytometry was performed as previously reported in the literature.32,33 Mouse peripheral blood, spleen, and lymph node cells from the drainage area were isolated, and 1×106 cells were taken. Single-cell suspensions were prepared by resuspending the cells in PBS containing 2% FBS. For the staining of B cells, CD4+ T cells, and CD8+ T cells, PerCP-Cy5.5-labeled anti-mouse CD19 antibody (Clone No. S-R532, Cat. S0B1764-500T, BD Biosciences), FITC-labeled anti-mouse CD4 antibody (Clone No. GK1.5, Cat. abs182365-25T, BD Biosciences), or PE-labeled anti-mouse CD8 antibody (Clone No. HB129/116-13.1, Cat. abs182402-25T, BD Biosciences) were added to the single-cell suspensions. The cells were incubated for 30 minutes on ice, protected from light. After incubation, the cells were washed and resuspended in PBS containing 2% FBS. The cell suspension was then filtered through a 300-mesh filter, and the proportions of CD19+ B cells, CD4+ T cells, and CD8+ T cells were detected by flow cytometry. For the staining of Th17 and Treg cells, antibodies labeling cell surface molecules were added to single-cell suspensions and incubated. For Th17 cells, a FITC-labeled anti-mouse CD4 antibody was used; for Treg cells, both a FITC-labeled anti-mouse CD4 antibody and an APC-labeled anti-mouse CD25 antibody were employed. Cells were then fixed with 4% paraformaldehyde and permeabilized with 0.1% saponin. Following this, antibodies against intracellular molecules were injected into the cells and incubated: an APC-labeled anti-mouse IL-17 antibody for Th17 cells and a PE-labeled anti-mouse Foxp3 antibody for Treg cells. The proportions of Th17 and Treg cells were detected by flow cytometry. The experiments were independently repeated 3 times to confirm result consistency.
Histopathology
Skin grafts were harvested on the 10th day post-transplantation. The samples were fixed in 4% paraformaldehyde, processed in graded alcohols, embedded in paraffin, cut into 5 µm sections, and then stained with hematoxylin and eosin (HE), as has been reported in previous literature.34 Under light microscopy, the structure and infiltration of inflammatory cells in skin allografts were examined and photographed using a scale from 0 to 5: 0 = absent, 1 = discrete, 2 = moderately discrete, 3 = moderate, 4 = moderately severe, and 5 = severe.
Statistical Analysis
Statistical Analysis of Demographic Characteristics and Clinical Indicators
For continuous variables, comparative analyses were performed using the Mann–Whitney U-test (for non-normally distributed data) or the t-test (for normally distributed data). For categorical data, comparative analyses were performed using the chi-square test. All data were analyzed using SPSS (Version 21.0) software, and a P-value of < 0.05 was considered statistically significant.
Intestinal Flora Analysis
α-diversity metrics were computed using R software, and the Mann–Whitney U-test was used to compare differences in α-diversity between groups. β-diversity was assessed based on principal coordinate analysis (PCoA) using weighted and unweighted UniFrac distance matrices. Permutational multivariate analysis of variance (PERMANOVA) was employed to test the significance of differences between groups. β-Diversity was also assessed and visualized using R software. Additionally, this study utilized the linear discriminant analysis effect size (LEfSe) method to perform linear discriminant analysis (LDA) to identify species with significant differences between groups. Species with LDA values greater than 2 and P < 0.05 were considered to be significantly different between groups.
Analysis of Intestinal Metabolites
Pattern recognition was performed using SIMCA-P 14.1, and the data were pre-processed by Pareto scaling and then subjected to multivariate analyses, including unsupervised principal component analysis (PCA) and supervised orthogonal partial least squares-discriminant analysis (OPLS-DA). Correlation analysis was performed using the Spearman test. Significantly enriched pathways were analyzed using the KEGG database (http://genome.jp/kegg/). The diagnostic value of potential biomarkers was assessed by plotting ROC curves and calculating the area under the ROC curve (AUC). The above statistical analyses and the plotting of volcano plots, bubble plots, Sankey plots, and ROC curves were conducted using R language (V3.4.4).
Research Results
Effect of Tacrolimus on the Intestinal Microbiota in Mice
To investigate the effect of tacrolimus on gut microbiota in mice, we administered a tacrolimus solution to the Tac group and a saline solution to the control group via gavage. After 7 days of consecutive gavage, fecal samples from both groups were collected for 16S rRNA gene sequencing. There was no significant difference in the number of observed species in the fecal samples between the two groups (P=0.77, Figure 1A). Alpha diversity analysis indicated no significant differences between the two groups in terms of Chao1 index (P = 0.77), Faith’s PD index (P = 1), Simpson index (P = 0.39), and Shannon index (P = 0.39) (Figure 1A). These results indicate that tacrolimus did not significantly affect the richness and evenness of the gut microbiota in mice. Furthermore, we evaluated the β-diversity of the gut microbiota in both groups using PCoA and NMDS analyses, and performed statistical analysis of the differences using the PERMANOVA test. The results indicate that the two sample groups can be distinctly separated based on PCoA and NMDS analyses (Figure 1B). The PERMANOVA test indicates a significant difference in beta diversity between the two sample groups (P = 0.025). These results suggest that tacrolimus can influence the composition of the gut microbiota in mice.
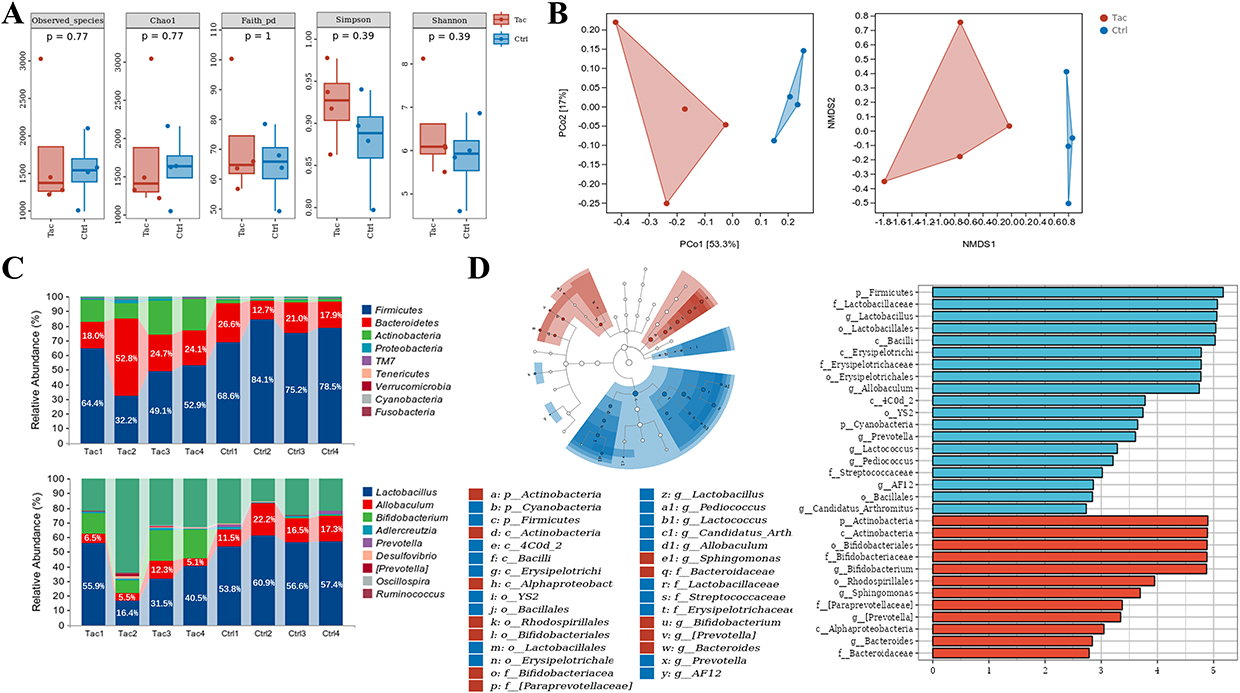 |
Figure 1 Effect of tacrolimus on intestinal flora in mice. (A) α diversity of gut microbiota in the Tac and Ctrl groups. (B) β diversity of gut microbiota in the Tac and Ctrl groups. The PCoA plot (left panel); the NMDS plot (right panel). (C) Compositions of gut microbiota in the Tac and Ctrl groups. (D) Identification of differential gut microbiota between the Tac and Ctrl groups. The differential gut microbiota between the two groups were identified based on data from 16s rRNA gene sequencing and LEfSe analysis (LDA > 2, P < 0.05). Each group comprised four mice. Abbreviations: Ctrl, control group; Tac, tacrolimus-treated group. |
To demonstrate the effect of tacrolimus on the gut microbiota composition of mice more specifically, we used cumulative histograms to illustrate the gut microbiota composition at the phylum and genus levels in the two groups. The results revealed significant differences in the gut microbiota compositions of the two groups at both the phylum and genus levels (Figure 1C and Figure S1). At the phylum level, the dominant microorganisms were Firmicutes (Tac group: 49.7%, control group: 76.6%), Bacteroidetes (Tac group: 29.9%, control group: 19.6%), and Actinobacteria (Tac group: 17.6%, control group: 1.7%). At the genus level, the dominant microorganisms in the Tac group were Lactobacillus (37.8%), Bifidobacterium (36.1%), and Allobaculum (15.6%), whereas the dominant microorganisms in the control group were Lactobacillus (57.2%), Allobaculum (16.9%), and Adlercreutzia (1.1%). To identify differences in gut microbiota between the two groups, we conducted LEfSe analysis and set the cutoff value for LDA scores at 2. The relative abundance of 19 microbial taxa, including Firmicutes, Lactobacillus, Lactobacillales, and Bacilli, showed a significant increase in the Tac group (Figure 1D). In contrast, the relative abundances of 12 microbial groups, including Actinobacteria, Bifidobacteriaceae, and Bifidobacterium, were significantly reduced in the Tac group (Figure 1D).
Effect of Tacrolimus on Intestinal Metabolites in Mice
We further analyzed the metabolic products in the fecal samples of the Tac group and the control group to evaluate the impact of tacrolimus-induced changes in gut microbiota on intestinal metabolic products. The PCA analysis showed partial overlap in the sample points of the two groups in both positive and negative ion modes, indicating differences in intestinal metabolic products between the two groups, albeit with low discrimination ability (Figure 2A). We further conducted the OPLS-DA analysis to elucidate the differences in intestinal metabolic products between the two groups. The results showed that in both positive and negative ion modes, the sample points of the Tac group and the control group were completely separated in the plot, further indicating differences in intestinal metabolic products between the two groups. Additionally, the explanatory ability evaluation parameter R²Y for both groups’ models exceeded 0.9, and the predictive ability evaluation parameter Q² exceeded 0.5. (Figure 2B). At the same time, permutation tests were conducted on two sets of OPLS-DA models in positive and negative ion modes. The results revealed that the slopes of the fitting lines for R² and Q² in both sets of OPLS-DA models were greater than 0, while the intercepts of Q² were less than 0, indicating no overfitting in the models.
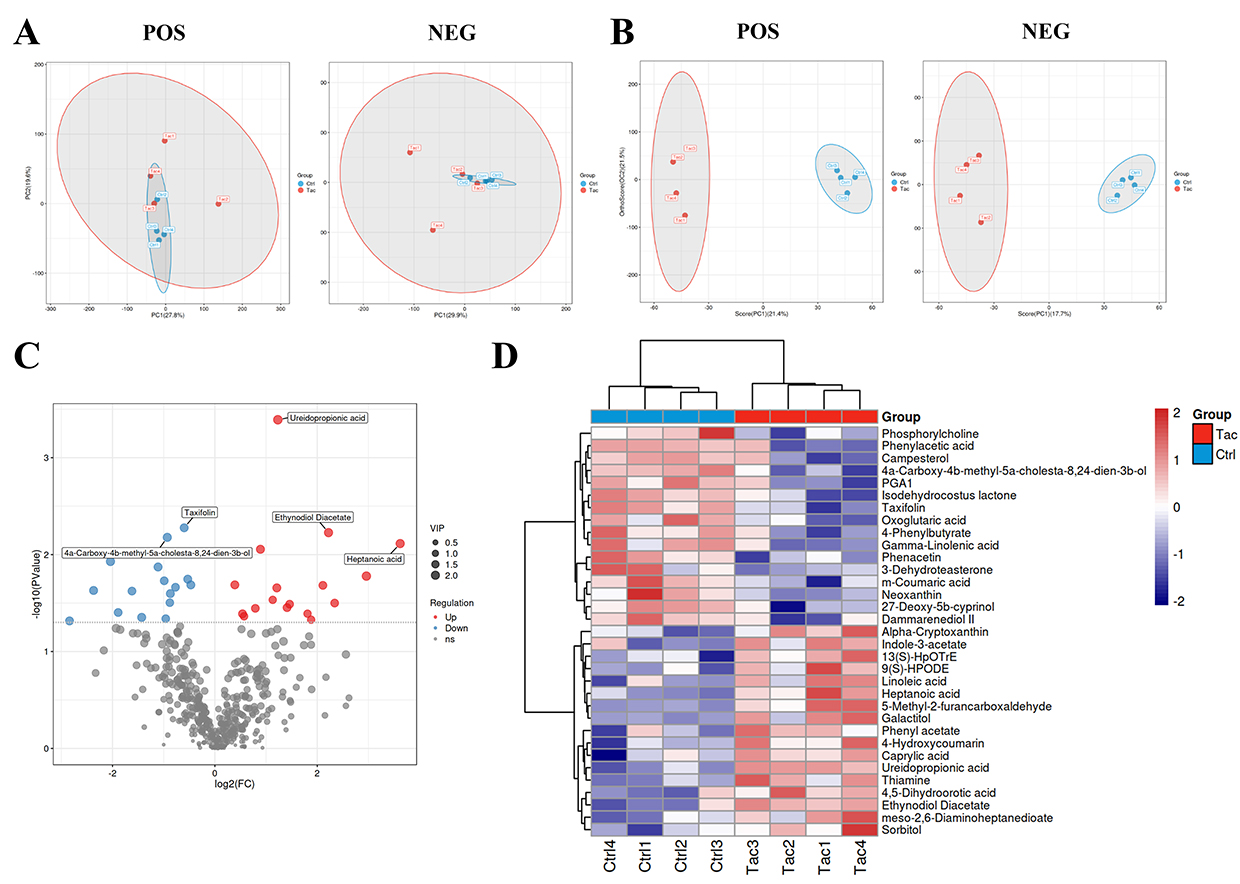 |
Figure 2 Effect of tacrolimus on intestinal metabolites in mice. (A) PCA analysis of intestinal metabolites in Tac and Ctrl groups. (B) OPLS-DA analysis of intestinal metabolites in Tac and Ctrl groups. (C) Screening of differential intestinal metabolites between Tac and Ctrl groups. The volcano plots were used to show the differential intestinal metabolites between the Tac and Ctrl groups (VIP > 1; P < 0.05). The abscissa represents log2(FC) and the ordinate represents -log10(P value). Blue and red denotes down-regulated and up-regulated intestinal metabolites in the Tac group, respectively, and gray represents non-significant features. (D) Identification of differential intestinal metabolites between Tac and Ctrl groups. Blue and red denotes down-regulated and up-regulated intestinal metabolites in the Tac group, respectively. Each group comprised four mice. Abbreviations: POS, positive ion mode; NEG, negative ion mode; Ctrl, control group; Tac, tacrolimus-treated group. |
Later, we combined the results of the single analysis and multivariate analysis to screen for differential metabolites in the intestines of mice from the Tac group and the control group, using screening criteria that met both ① OPLS-DA analysis VIP > 1 and ② univariate analysis P < 0.05. The volcano plot showed that a total of 489 metabolites were detected, of which 17 were upregulated and 16 were downregulated in the Tac group (Figure 2C). The heat map was used to display the differential metabolites, revealing that heptanoic acid, 5-methyl-2-furfural, D-sorbitol, and ethyl acetate were upregulated in the Tac group, while oxalic acid, xanthotoxin, abietic acid, and prostaglandin A1 were downregulated in the same group (Figure 2D).
Protective Effect of the Changes in the Gut Microbiota Induced by Tacrolimus on Skin Grafts in Mice
The mice were randomly divided into four groups as follows: (1) Ctrl group (mice with no treatment); (2) Tac group (mice were treated with tacrolimus); (3) FMT group (mice were treated with fecal bacterial solution from those treated with saline); (4) Tac FMT group (mice were treated with fecal bacterial solution from those treated with tacrolimus). The treatment program is shown in Figure 3A. Results revealed that on the 10th day post-transplantation, skin grafts in both the Ctrl and FMT groups exhibited visible signs of hardening, scabbing, shriveling, and even sloughing off, whereas skin grafts in the Tac and Tac FMT groups remained relatively soft with signs of hair regrowth (Figure 3B). Histopathologically, on the 10th day post-transplantation, the skin grafts from the Ctrl group and FMT group mice exhibited more pronounced rejection characteristics, including follicular destruction, surface ulceration, degenerated squamous epithelium, and vesicular epidermis (Figure 3C). By contrast, the Tac and Tac FMT group mice demonstrated better epidermal tissue integrity and improved squamous epithelial growth status (Figure 3C). Compared to the mice in the Ctrl and the FMT groups, those in the Tac and Tac FMT groups displayed a significant decrease in inflammatory cell infiltration in the skin grafts (Figure 3D). Skin allografts in the Tac and Tac FMT groups survived significantly longer than those in the Ctrl group (P = 0.0101; P = 0.0263, respectively). However, there was no significant increase in survival time in the FMT group (P = 0.1969) (Figure 3E).
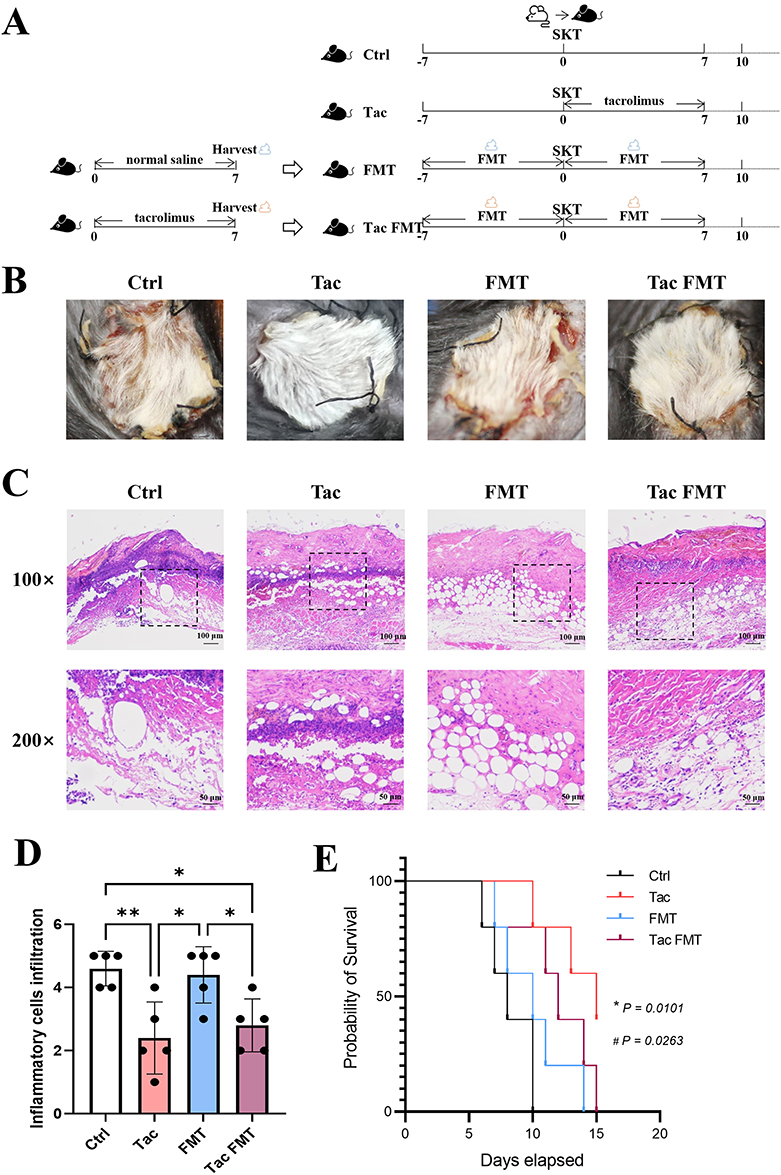 |
Figure 3 Protective effect of tacrolimus-induced changes in intestinal flora on skin grafts in mice. (A) Experimental design of mouse skin allograft and drug administration. (B) General view of mouse skin allograft. On day 10 after transplantation, skin allografts from the mice in each group were observed and photographed. (C) Histological tissue staining of mouse skin allograft. On day 10 after transplantation, skin allografts from the mice in each group were harvested for HE staining, and the sections were observed and photographed under a light microscope at a magnification of 100× or 200×. The 200× magnification images are derived from the dotted squares in the corresponding 100× magnification images. (D) Assessment of inflammatory cell infiltration. (E) Survival time of mouse skin allograft. Survival curves were generated using Kaplan-Meier estimates; between-group *Tac group vs Ctrl group P = 0.0101; #Tac FMT group vs Ctrl group P = 0.0263; differences in survival duration were compared using the Log rank test. Each group comprised five mice. Abbreviations: Ctrl, control group; Tac, tacrolimus-treated group; FMT, the group in which the mice were administrated fecal microbiota transplantation with feces from normal saline-gavaged mice; Tac FMT, the group in which the mice were administrated fecal microbiota transplantation with feces from tacrolimus-gavaged mice; SKT, skin transplantation. |
Impact of Gut Microbiota Changes Induced by Tacrolimus on the Immune Function in vivo
To further investigate whether the change in gut microbiota induced by tacrolimus protects mouse skin grafts by regulating the immune function of mice, we analyzed immune cells in the peripheral blood, spleen, and draining lymph nodes of the Ctrl group, Tac group, FMT group, and Tac FMT group mice on day 7 after skin transplantation. The results showed that no significant differences were observed in the proportions of CD19+ B cells, CD4+ T cells, CD8+ T cells, Th17 cells, and Treg cells among the spleens and peripheral blood of the mice in each group (Figure 4A and B). In the draining lymph nodes, the proportion of Treg cells in the Tac FMT group was significantly higher than that in the Ctrl group (P = 0.009). It is worth noting that, although not statistically significant, the average proportion of Th17 cells in the draining lymph nodes of the Tac group and Tac FMT group was higher compared to the Ctrl group and FMT group (Figure 4C). The proportions of CD19+ B cells, CD4+ T cells, and CD8+ T cells in the draining lymph nodes from the four groups showed no significant differences (Figure 4C and Figure S2). Our study further investigated the changes in immune cells of mesenteric lymph nodes, and the results revealed that the proportion of Treg cells in the Tac FMT group was significantly higher than that in the Ctrl group (P = 0.0425). In contrast, the proportion of Th17 cells in the Tac group and Tac FMT group was significantly lower than that of the Ctrl group (P = 0.0299). There was no significant difference in the proportions of CD19+ B cells, CD4+ T cells, and CD8+ T cells among the four groups (Figure 4D, Figures S3 and S4). Additionally, we observed no statistically significant differences in the proportions of immune cells in the spleen, peripheral blood, draining lymph nodes, or mesenteric lymph nodes between mice in the Tac group and Tac FMT group (Figure 4A–D). The results above suggest that the protective effect of gut microbiota changes induced by tacrolimus on skin grafts may be mediated through the regulation of the balance between Th17 cells and Treg cells in the lymph nodes.
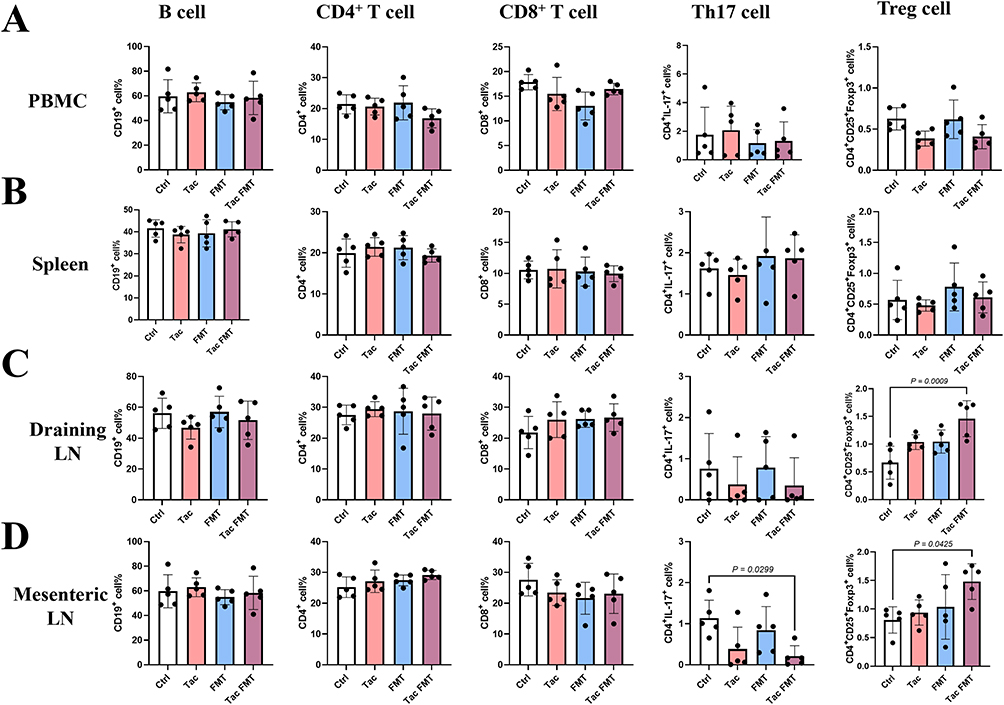 |
Figure 4 Analysis of the immune cells in the mice with skin transplantation. The proportion of B cell, CD4+ T cell, CD8+ T cell, Th17 cell, and Treg cell were analyzed by flow cytometry. (A) Analysis of the immune cells in peripheral blood; (B) Analysis of the immune cells in spleen; (C) Analysis of the immune cells in draining lymph nodes; (D) Analysis of the immune cells in mesenteric lymph nodes. Absence of symbols indicates no statistical difference between groups. Each group comprised five mice. Abbreviations: Ctrl, control group; Tac, tacrolimus-treated group; FMT, the group in which the mice were administrated fecal microbiota transplantation with feces from normal saline-gavaged mice; Tac FMT, the group in which the mice were administrated fecal microbiota transplantation with feces from tacrolimus-gavaged mice. |
Discussion
In general, our study revealed that tacrolimus can induce changes in the composition of gut microbiota and gut metabolic products in mice. Fecal microbiota transplantation using feces from tacrolimus-treated mice significantly prolonged the survival time of skin grafts and attenuated graft rejection. Compared to the Ctrl group, the proportion of Treg cells in the mesenteric lymph nodes was significantly elevated in the Tac FMT group, whereas the proportion of Th17 cells was significantly decreased, suggesting that the tacrolimus-induced gut microbiome may exert its protective effect by influencing the balance between Treg cells and Th17 cells.
In 2017, Tourret et al were the first to use 16S rRNA gene sequencing to explore the impact of tacrolimus and other immunosuppressants on the gut microbiota. The findings indicated that after treatment with tacrolimus, mice exhibited significant changes in their gut microbiota at the family level.28 Subsequently, Zhang et al further confirmed the effects of tacrolimus on the gut microbiota of mice using a skin transplantation model. They demonstrated that the combination of low-dose tacrolimus and fecal microbiota transplantation from mice treated with high-dose tacrolimus had a protective effect on skin grafts.27 Compared to these two studies, our research aims to elucidate the potential mechanisms of tacrolimus treatment for the transplant rejection through the lenses of gut microbiota and metabolism. Consistent with the findings of Zhang’s study, we discovered that although the gavage treatment with tacrolimus in mice did not alter the alpha diversity of the gut microbiota, it significantly affected the beta diversity, suggesting that its impact on the gut microbiota of mice is primarily reflected in composition rather than diversity. Additionally, we identified a total of 19 microbial groups that were upregulated and 12 that were downregulated in the feces of mice treated with tacrolimus. Given the close relationship and cross-talk between the gut microbiota and the immune system,35 we hypothesize that tacrolimus-induced changes in gut microbiota depend on two pathways. The first is the direct action of tacrolimus on the gut microbiota, and the second might be linked to the systemic immunosuppressive effects of tacrolimus, which in turn affect the gut microbiota.
To explore whether the alterations in gut microbiota induced by tacrolimus can provide protection for allogeneic skin grafts, we performed FMT using feces from tacrolimus-treated mice on mice undergoing skin transplantation. The specific protocol involved administering FMT to mice one week prior to skin transplantation, performing the skin graft on day 0, and continuing FMT for an additional 7 days starting on the day after transplantation. The results show that, compared to the control group, the survival time of skin grafts in model mice treated with this method was significantly prolonged. Unlike our study, the research conducted by Zhang et al found that FMT using feces from mice treated with tacrolimus alone did not show a protective effect on the allogeneic skin grafts. This discrepancy may be attributed to the different timing of the FMT procedures that we used. In our study, we performed FMT 7 days before and 7 days after skin transplantation, whereas in the study by Zhang et al, FMT was only conducted 14 days prior to skin transplantation. In studies utilizing FMT to treat Clostridium difficile infection, although the diversity of the gut microbiota in patients did not change significantly after FMT, the composition of the gut microbiota did change as the duration following FMT increased.36 We speculate that the absence of FMT following skin transplantation in the study by Zhang et al may have reduced the protective effect of FMT on the grafts in the model mice. According to our results, only the mice in the tacrolimus treatment group (Tac group) and the group receiving FMT from tacrolimus-gavaged mice (Tac FMT group) showed a prolonged survival time for skin grafts. In contrast, the group receiving FMT from saline-gavaged mice (FMT group) did not exhibit a similar protective effect. These results suggest that the protective effect of tacrolimus-induced alterations in the gut microbiota on skin grafts may be achieved by modifying the composition of the gut microbiota rather than its diversity.
Additionally, we observed a significant increase in the proportion of Treg cells in the mesenteric lymph nodes and the draining lymph nodes of the Tac FMT group. Treg cells, characterized by the expression of CD4, CD25, and the transcription factor Foxp3, exert immunosuppressive effects through various mechanisms. These include the expression of the inhibitory receptor CTLA-4, the production of anti-inflammatory cytokines such as IL-10 and TGF-β1, and the modulation of the effects of IL-2 and ATP.37 Treg cells not only directly suppress the activation of CD4+ and CD8+ T cells and inhibit B cell responses and antibody production, but they also regulate the function of macrophages and natural killer cells.38 In recent years, studies have shown that Treg cells play a crucial role in inducing transplant tolerance. After infusing TGF-β-induced Treg cells into kidney transplant mice experiencing acute transplant rejection, the levels of circulating donor-specific antibodies can be reduced, leading to decreased infiltration of B cells, plasma cells, and Th17 cells, thereby alleviating transplant rejection.39 In contrast, depleting Treg cells in the kidney transplant mice can promote transplant rejection.40 Similar to our research findings, Zhang’s study also revealed that treatment with tacrolimus and fecal microbiota transplantation from tacrolimus-gavaged mice promoted a significant increase in the proportion of Treg cells in the colonic mucosa and the circulation of the mice.27 In addition, we noted a significant decrease in the ratio of Th17 cells to Treg cells. Previous studies have shown that there is a balance between Treg cells and Th17 cells, with Treg cells regulating the proportion and function of Th17 cells. It appears that factors promoting the development of Treg cells typically inhibit the development of Th17 cells.41,42
Unlike Treg cells, Th17 cells can produce the hallmark cytokines IL-17A, IL-17F, and IL-22, which induce antimicrobial peptides in epithelial cells. This plays a crucial role in mucosal defense against extracellular bacteria and fungi, as well as in inducing chemokines to recruit neutrophils and promote inflammatory responses.43 Additionally, research has shown that Th17 cells play a significant role in transplant rejection. In a mouse model of acute rejection following heart transplantation, there is an increase in Th17 cells and an imbalance between Th17 and Treg cells.44 An increase in Th17 cells was observed in the kidneys of patients with failed kidney transplants, and the number of Th17 cells was positively correlated with the course of chronic graft rejection.45 Treatment of individuals experiencing rejection with anti-IL-17 antibodies reduces rejection-induced lesions.46 This evidence further supports the idea that tacrolimus-induced alterations in the intestinal flora of mice may exert immunosuppressive effects by regulating the balance between Treg cells and Th17 cells.
In our study, tacrolimus treatment was also associated with a increased abundance of probiotic genus, Bifidobacterium. Multiple strains of Bifidobacterium are frequently employed as biotherapeutic agents and demonstrate favorable immunomodulatory and anti-inflammatory attributes, which include increasing the expression of inhibitory Tregs, enhancing intestinal barrier integrity, and suppressing intestinal Th2 and Th17 immune responses.47–49 Latest research findings demonstrate that the abundance of Bifidobacterium pseudolongum can be significantly upregulated by IAA, and Bifidobacterium pseudolongum ATCC25526 can alleviate DSS-induced colitis by increasing the proportion of Tregs in colonic tissue.50 Based on this, we hypothesize that the alterations in the Th17/Treg balance observed in this study may be also closely associated with Bifidobacterium. Moreover, we observed that tacrolimus treatment significantly downregulated the abundance of Lactobacillus in the intestinal microbiota of mice. Lactobacillus intestinalis was reported not to directly modulate T cell differentiation, but suppress the C/EBPα-driven production of SAA1 and SAA2 in intestinal epithelial cells. This reduction in SAA1/SAA2 expression, in turn, inhibits the differentiation of Th17 cells.51 Interestingly, another study has shown that during SIV infection in rhesus macaques, specific depletion of gut-resident Lactobacillus is associated with the loss of Th17 cells, suggesting that it thereby contribute to preserving Th17 cells during pathogenic SIV infection.52 Furthermore, we also found that another ligand of AHR, linoleic acid, was elevated in the intestines of tacrolimus-treated mice. Linoleic acid is a key target metabolite that modulates the extent of inflammation. It was reported to inhibit Th17 cell differentiation via phosphorylation of Ser727 in STAT1, while concurrently promoting the differentiation of Tregs.53 Therapeutic restoration of linoleic acid levels has been shown to reduce the Th17/Treg cell ratio in an AHR-dependent manner, thereby conferring anti-colitis effects.53 In contrast, our observations revealed a decreased level of phosphocholine in the intestines of mice following tacrolimus treatment. Studies have demonstrated that IgM against phosphocholine enhanced Treg cell polarization in derived from SLE patients and atherosclerotic plaques while reducing the production of IL-17 and TNF-α in cell cultures from these two populations.54
Metabolites are key substances in the interaction between intestinal flora and the host. Therefore, we examined and analyzed intestinal metabolites in tacrolimus-gavaged and saline-gavaged mice using an untargeted metabolomics approach. The results showed that, compared with the saline-gavaged mice, there were significant differences in the intestinal metabolites of the tacrolimus-gavaged mice. Seventeen metabolites were upregulated in the feces of the tacrolimus-gavaged group, while sixteen metabolites were downregulated in the same group. Indole-3-acetic acid is an important indole derivative produced by the gut microbiota from the catabolism of dietary tryptophan. It acts as a ligand for the aromatic hydrocarbon receptor and can regulate intestinal homeostasis and mucosal immunity through activation of the aromatic hydrocarbon receptor pathway.55 However, whether the tacrolimus-induced alterations in the intestinal flora of mice exert a protective effect on skin grafts by modulating indole-3-acetic acid needs to be verified through in vivo experiments involving the supplementation of indole-3-acetic acid. Similarly, in a mouse model of ankylosing spondylitis, activation of the aromatic hydrocarbon receptor has been reported to inhibit the production of pro-inflammatory cytokines (TNF-α, IL-6, IL-17A, and IL-23) and promote the production of the anti-inflammatory cytokine IL-10. This process increases Treg cell levels by upregulating the transcription factor Foxp3 and decreases Th17 levels by downregulating the transcription factor RORγt.56
Furthermore, we also found that another ligand of AHR, linoleic acid, was elevated in the intestines of tacrolimus-treated mice. Linoleic acid is a key target metabolite that modulates the extent of inflammation. It was reported to inhibit Th17 cell differentiation via phosphorylation of Ser727 in STAT1, while concurrently promoting the differentiation of Tregs.53 Therapeutic restoration of linoleic acid levels has been shown to reduce the Th17/Treg cell ratio in an AHR-dependent manner, thereby conferring anti-colitis effects.53 In contrast, our observations revealed a decreased level of phosphocholine in the intestines of mice following tacrolimus treatment. Studies have demonstrated that IgM against phosphocholine enhanced Treg cell polarization in derived from SLE patients and atherosclerotic plaques while reducing the production of IL-17 and TNF-α in cell cultures from these two populations.54
The present research has some constraints and points that should be addressed. In the present study, we investigated the effect of fecal bacterial solutions derived from tacrolimus-treated mice on skin grafts. We found that these solutions exerted a protective effect, which suggests a direct association between the protective action of tacrolimus and the gut microbiota. However, we did not explore whether antibiotics can abrogate tacrolimus-induced protection of skin grafts. In future research, supplementary experiments are warranted to validate, from multiple perspectives, that the protective effect of tacrolimus is dependent on the gut microbiota. While the results suggest that tacrolimus-induced gut microbiome may influence the balance between Treg cells and Th17 cells, the causal relationship between specific microbiota or metabolites and Treg/Th17 modulation remains unclear. More in-depth experiments, such as metabolite supplementation, germ-free models, or co-culture assays, are needed to clarify the direct relationship. It has been reported that there is a close relationship and cross-talk among organ transplantation, the gut microbiota, and the immune system.35 In order to isolate the effect of skin transplantation and the immune system on the gut microbiota, mice that underwent skin transplantation and received no drug treatment were used as the control group. Our current results cannot explain whether the changes in gut microbiota are due to the direct effects of tacrolimus or are secondary to its overall immune modulation. Control experiments in which tacrolimus is administered systemically but without direct effects on the gut are needed in the future. In this study, we revealed changes in Th17 and Treg cell populations in the mesenteric lymph nodes without investigating the levels of inflammatory cytokines. It is another shortcoming of the present study that we did not investigate the effects of tacrolimus-induced alterations in gut microbiota and their metabolites on the absolute numbers of immune cells. Our future study would explore strategies to reduce the side effects of tacrolimus, such as combining it with probiotics or specific microbial metabolites. In addition, we used a fully mismatched allogeneic transplant model to elicit robust transplant rejection. This model enabled clear observation of how tacrolimus-induced gut microbiota and metabolite alterations influence immune cells and transplant rejection, and facilitated exploration of the effects and mechanisms of myeloid cell-specific nanobiologics and LFA-1 blockade on allografts.57,58 However, clinical HLA phenotyping ensures partial allele sharing between donors and recipients. Future studies will further explore how tacrolimus-induced gut microbiota and metabolite alterations affect skin allografts using a semi-mismatched model.
Conclusions
In summary, our research revealed that tacrolimus had no marked effect on gut microbiota α-diversity but changed β-diversity and composition, and altered 33 fecal metabolites. FMT from tacrolimus-treated mice prolonged skin allograft survival, ameliorated graft pathology, linked to adjusted Treg/Th17 proportions in lymph nodes. These data may provide a basis for establishing gut microbiota-based therapies for allograft rejection, as well as the development of strategies for reshaping the microbiota in a targeted manner. Nevertheless, the use of a murine model may not fully reflect the complexity of human allograft rejection, and there may be potential challenges in translating our findings to clinical practice.
Data Sharing Statement
The raw and processed data generated in this study are available from the corresponding author, Xin Li, upon reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the Natural Science Foundation of Henan Province (No.232300421292), the National Natural Science Foundation of China (No.82270781) and the Henan Provincial Science and Technology Research Project (No.242102311219). The sponsors had no role in study design, data collection, data analysis, data interpretation, or writing of the report.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Halloran P. Immunosuppressive drugs for kidney transplantation. New Engl J Med. 2005;351:2715–2729. doi:10.1056/NEJMra033540
2. Baran DA, Galin I, Sandler D, et al. Tacrolimus in cardiac transplantation: efficacy and safety of a novel dosing protocol. Transplantation. 2002;74(8):1136–1141. doi:10.1097/00007890-200210270-00014
3. Hart A, Smith JM, Skeans MA, et al. OPTN/SRTR 2015 annual data report: kidney. Am J Transplant. 2017;17(Suppl 1):21–116. doi:10.1111/ajt.14124
4. Schreiber SL. Chemistry and biology of the immunophilins and their immunosuppressive ligands. Science. 1991;251(4991):283–287. doi:10.1126/science.1702904
5. Venkataramanan R, Swaminathan A, Prasad T, et al. Clinical pharmacokinetics of tacrolimus. Clin Pharmacokinet. 1995;29(6):404–430. doi:10.2165/00003088-199529060-00003
6. Rath T. Tacrolimus in transplant rejection. Expert Opinion Pharmacother. 2013;14(1):115–122. doi:10.1517/14656566.2013.751374
7. Barbarino JM, Staatz CE, Venkataramanan R, Klein TE, Altman RB. PharmGKB summary: cyclosporine and tacrolimus pathways. Pharmacogen Genom. 2013;23(10):563–585. doi:10.1097/FPC.0b013e328364db84
8. Werk AN, Lefeldt S, Bruckmueller H, et al. Identification and characterization of a defective CYP3A4 genotype in a kidney transplant patient with severely diminished tacrolimus clearance. Clin Pharmacol Ther. 2014;95(4):416–422. doi:10.1038/clpt.2013.210
9. Birdwell KA, Decker B, Barbarino JM, et al. Clinical Pharmacogenetics Implementation Consortium (CPIC) guidelines for CYP3A5 genotype and tacrolimus dosing. Clin Pharmacol Ther. 2015;98(1):19–24. doi:10.1002/cpt.113
10. Van Driest SL, Webber SA. Pharmacogenomics: personalizing pediatric heart transplantation. Circulation. 2015;131(5):503–512. doi:10.1161/CIRCULATIONAHA.114.001382
11. Hong E, Carmanov E, Shi A, et al. Application of physiologically based pharmacokinetic modeling to predict drug-drug interactions between elexacaftor/tezacaftor/ivacaftor and tacrolimus in lung transplant recipients. Pharmaceutics. 2023;15(5):1438. doi:10.3390/pharmaceutics15051438
12. Eriguchi Y, Takashima S, Oka H, et al. Graft-versus-host disease disrupts intestinal microbial ecology by inhibiting Paneth cell production of α-defensins. Blood. 2012;120(1):223–231. doi:10.1182/blood-2011-12-401166
13. Lee JR, Muthukumar T, Dadhania D, et al. Gut microbial community structure and complications after kidney transplantation: a pilot study. Transplantation. 2014;98(7):697–705. doi:10.1097/TP.0000000000000370
14. Oh PL, Martínez I, Sun Y, Walter J, Peterson DA, Mercer DF. Characterization of the ileal microbiota in rejecting and nonrejecting recipients of small bowel transplants. Am J Transplant. 2012;12(3):753–762. doi:10.1111/j.1600-6143.2011.03860.x
15. Ren Z, Jiang J, Lu H, et al. Intestinal microbial variation may predict early acute rejection after liver transplantation in rats. Transplantation. 2014;98(8):844–852. doi:10.1097/TP.0000000000000334
16. Fricke WF, Maddox C, Song Y, Bromberg JS. Human microbiota characterization in the course of renal transplantation. Am J Transplant. 2014;14(2):416–427. doi:10.1111/ajt.12588
17. Vieira AT, Fukumori C, Ferreira CM. New insights into therapeutic strategies for gut microbiota modulation in inflammatory diseases. Clin Transl Immunol. 2016;5(6):e87. doi:10.1038/cti.2016.38
18. Mariño E. The gut microbiota and immune-regulation: the fate of health and disease. Clin Transl Immunol. 2016;5(11):e107. doi:10.1038/cti.2016.61
19. Lei YM, Sepulveda M, Chen L, et al. Skin-restricted commensal colonization accelerates skin graft rejection. JCI Insight. 2019;5(15). doi:10.1172/jci.insight.127569
20. Bromberg JS, Hittle L, Xiong Y, et al. Gut microbiota-dependent modulation of innate immunity and lymph node remodeling affects cardiac allograft outcomes. JCI Insight. 2018;3(19). doi:10.1172/jci.insight.121045
21. Bai YZ, Roberts SH, Kreisel D, Nava RG. Microbiota in heart and lung transplantation: implications for innate-adaptive immune interface. Curr Opin Organ Transplant. 2021;26(6):609–614. doi:10.1097/MOT.0000000000000923
22. Walker A, Pfitzner B, Harir M, et al. Sulfonolipids as novel metabolite markers of Alistipes and Odoribacter affected by high-fat diets. Sci Rep. 2017;7(1):11047. doi:10.1038/s41598-017-10369-z
23. McIntosh CM, Chen L, Shaiber A, Eren AM, Alegre ML. Gut microbes contribute to variation in solid organ transplant outcomes in mice. Microbiome. 2018;6(1):96. doi:10.1186/s40168-018-0474-8
24. Smith PM, Howitt MR, Panikov N, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. 2013;341(6145):569–573. doi:10.1126/science.1241165
25. Wu H, Singer J, Kwan TK, et al. Gut microbial metabolites induce donor-specific tolerance of kidney allografts through induction of T regulatory cells by short-chain fatty acids. J Am Soc Nephrol. 2020;31(7):1445–1461. doi:10.1681/ASN.2019080852
26. Campbell PM, Humphreys GJ, Summers AM, et al. Does the microbiome affect the outcome of renal transplantation? Front Cell Infect Microbiol. 2020;10.
27. Zhang Z, Liu L, Tang H, et al. Immunosuppressive effect of the gut microbiome altered by high-dose tacrolimus in mice. Am J Transplant. 2018;18(7):1646–1656.
28. Tourret J, Willing BP, Dion S, MacPherson J, Denamur E, Finlay BB. Immunosuppressive treatment alters secretion of ileal antimicrobial peptides and gut microbiota, and favors subsequent colonization by uropathogenic Escherichia coli. Transplantation. 2017;101(1):74–82.
29. Cheng CH, Lee CF, Fryer M, et al. Murine full-thickness skin transplantation. J Visualized Exp. 2017;(119).
30. Guo R, Hu F, Weng Q, et al. Guiding T lymphopoiesis from pluripotent stem cells by defined transcription factors. Cell Res. 2020;30(1):21–33. doi:10.1038/s41422-019-0251-7
31. Chong AS, Alegre ML, Miller ML, Fairchild RL. Lessons and limits of mouse models. Cold Spring Harbor Perspect Med. 2013;3(12):a015495. doi:10.1101/cshperspect.a015495
32. Lu Z, Priya Rajan SA, Song Q, et al. 3D scaffold-free microlivers with drug metabolic function generated by lineage-reprogrammed hepatocytes from human fibroblasts. Biomaterials. 2021;269:120668.
33. Li J, Wang X, Chen Y, et al. Exosome-mediated lectin pathway and resistin-MIF-AA metabolism axis drive immune dysfunction in immune thrombocytopenia. Adv Sci. 2025;12(10):e2412378. doi:10.1002/advs.202412378
34. Pullikuth AK, Routh ED, Zimmerman KD, et al. Bulk and single-cell profiling of breast tumors identifies TREM-1 as a dominant immune suppressive marker associated with poor outcomes. Front Oncol. 2021;11:734959. doi:10.3389/fonc.2021.734959
35. Wang W, Xu S, Ren Z, Jiang J, Zheng S. Gut microbiota and allogeneic transplantation. J Transl Med. 2015;13:275. doi:10.1186/s12967-015-0640-8
36. Shankar V, Hamilton MJ, Khoruts A, et al. Species and genus level resolution analysis of gut microbiota in Clostridium difficile patients following fecal microbiota transplantation. Microbiome. 2014;2:13. doi:10.1186/2049-2618-2-13
37. Whibley N, Tucci A, Powrie F. Regulatory T cell adaptation in the intestine and skin. Nat Immunol. 2019;20(4):386–396. doi:10.1038/s41590-019-0351-z
38. Ohkura N, Sakaguchi S. Transcriptional and epigenetic basis of Treg cell development and function: its genetic anomalies or variations in autoimmune diseases. Cell Res. 2020;30(6):465–474. doi:10.1038/s41422-020-0324-7
39. Liao T, Xue Y, Zhao D, et al. In vivo attenuation of antibody-mediated acute renal allograft rejection by ex vivo TGF-β-induced CD4(+)Foxp3(+) regulatory T cells. Front Immunol. 2017;8:1334. doi:10.3389/fimmu.2017.01334
40. Hall BM. CD4+CD25+ T regulatory cells in transplantation tolerance: 25 years on. Transplantation. 2016;100(12):2533–2547. doi:10.1097/TP.0000000000001436
41. Yang J, Chu Y, Yang X, et al. Th17 and natural Treg cell population dynamics in systemic lupus erythematosus. Arthritis Rheum. 2009;60(5):1472–1483. doi:10.1002/art.24499
42. Seki M, Oomizu S, Sakata KM, et al. Galectin-9 suppresses the generation of Th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis. Clin Immun. 2008;127(1):78–88. doi:10.1016/j.clim.2008.01.006
43. Yasuda K, Takeuchi Y, Hirota K. The pathogenicity of Th17 cells in autoimmune diseases. Semin Immunopathol. 2019;41(3):283–297. doi:10.1007/s00281-019-00733-8
44. Jankowska Gan E, Agashe VV, Lema DA, et al. Donor HLA-DR drives the development of De Novo autoimmunity following lung and heart transplantation. Transplant Direct. 2020;6(10):e607. doi:10.1097/TXD.0000000000001062
45. Rodriguez RM, Hernández-Fuentes MP, Corte-Iglesias V, et al. Defining a methylation signature associated with operational tolerance in kidney transplant recipients. Front Immunol. 2021;12:709164. doi:10.3389/fimmu.2021.709164
46. Zhang M, Xu M, Wang K, Li L, Zhao J. Effect of Inhibition of the JAK2/STAT3 signaling pathway on the Th17/IL-17 axis in acute cellular rejection after heart transplantation in mice. J Cardiovasc Pharmacol. 2021;77(5):614–620. doi:10.1097/FJC.0000000000001007
47. Sanders ME, Merenstein DJ, Reid G, Gibson GR, Rastall RA. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat Rev Gastroenterol Hepatol. 2019;16(10):605–616. doi:10.1038/s41575-019-0173-3
48. Verma R, Lee C, Jeun EJ, et al. Cell surface polysaccharides of Bifidobacterium bifidum induce the generation of Foxp3(+) regulatory T cells. Sci Immunol. 2018;3(28). doi:10.1126/sciimmunol.aat6975
49. Henrick BM, Rodriguez L, Lakshmikanth T, et al. Bifidobacteria-mediated immune system imprinting early in life. Cell. 2021;184(15):3884–3898.e3811. doi:10.1016/j.cell.2021.05.030
50. Li M, Han X, Sun L, Liu X, Zhang W, Hao J. Indole-3-acetic acid alleviates DSS-induced colitis by promoting the production of R-equol from Bifidobacterium pseudolongum. Gut Microbes. 2024;16(1):2329147. doi:10.1080/19490976.2024.2329147
51. Wang QW, Jia DJ, He JM, et al. Lactobacillus intestinalis primes epithelial cells to suppress colitis-related Th17 response by host-microbe retinoic acid biosynthesis. Adv Sci. 2023;10(36):e2303457. doi:10.1002/advs.202303457
52. Vujkovic-Cvijin I, Swainson LA, Chu SN, et al. Gut-resident lactobacillus abundance associates with IDO1 inhibition and Th17 dynamics in SIV-infected macaques. Cell Rep. 2015;13(8):1589–1597. doi:10.1016/j.celrep.2015.10.026
53. Jia L, Jiang Y, Wu L, et al. Porphyromonas gingivalis aggravates colitis via a gut microbiota-linoleic acid metabolism-Th17/Treg cell balance axis. Nat Commun. 2024;15(1):1617. doi:10.1038/s41467-024-45473-y
54. Sun J, Lundström SL, Zhang B, et al. IgM antibodies against phosphorylcholine promote polarization of T regulatory cells from patients with atherosclerotic plaques, systemic lupus erythematosus and healthy donors. Atherosclerosis. 2018;268:36–48. doi:10.1016/j.atherosclerosis.2017.11.010
55. Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9(1):3294.
56. Shen J, Yang L, You K, et al. Indole-3-acetic acid alters intestinal microbiota and alleviates ankylosing spondylitis in mice. Front Immunol. 2022;13:762580. doi:10.3389/fimmu.2022.762580
57. Braza MS, Van leent MMT, Lameijer M, et al. Inhibiting inflammation with myeloid cell-specific nanobiologics promotes organ transplant acceptance. Immunity. 2018;49(5):819–828.e816. doi:10.1016/j.immuni.2018.09.008
58. Reisman NM, Floyd TL, Wagener ME, Kirk AD, Larsen CP, Ford ML. LFA-1 blockade induces effector and regulatory T-cell enrichment in lymph nodes and synergizes with CTLA-4Ig to inhibit effector function. Blood. 2011;118(22):5851–5861. doi:10.1182/blood-2011-04-347252
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Tacrolimus Trough Level Variation and Its Correlation to Clinical Outcomes and Consequences in Solid Organ Transplantation
Albilal S, Shawaqfeh MS, Albusaysi S, Fetyani L, Alnashmi F, Alshehri SD, Albekairy NA, Akhulaif A, Alzahrani L, Alwuhayde M, Obaidat AA, Al Bekairy AM
Transplant Research and Risk Management 2023, 15:1-11
Published Date: 8 August 2023