")
Back to Journals » Infection and Drug Resistance » Volume 17
Genomic Sequencing of Clinical Cupriavidus gilardii Isolates Revealed Their Diverse Antimicrobial Resistance Mechanisms
Authors Zhao W, Yang H, Huang Y, Fan X, Tong Z
Received 2 September 2023
Accepted for publication 26 January 2024
Published 16 February 2024 Volume 2024:17 Pages 655—664
DOI https://doi.org/10.2147/IDR.S438328
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Weichao Zhao,1,2,* Huqin Yang,1,* Yong Huang,3,* Xin Fan,4 Zhaohui Tong1
1Department of Respiratory and Critical Care Medicine, Beijing Institute of Respiratory Medicine, Beijing Chao-Yang Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Department of Respiratory Medicine, Strategic Support Force Medical Center, Beijing, People’s Republic of China; 3State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing, People’s Republic of China; 4Department of Infectious Diseases and Clinical Microbiology, Beijing Institute of Respiratory Medicine, Beijing Chao-Yang Hospital, Capital Medical University, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhaohui Tong; Xin Fan, NO. 8 Gong Ti South Road, Beijing, 10020, People’s Republic of China, Tel +86 1085231297 ; +86 13521438062, Email [email protected]; [email protected]
Purpose: Cupriavidus gilardii is an emerging multidrug-resistant pathogen found in many environments and few clinical samples. The clinical infectiousness, pathogenicity, and resistance mechanisms of C. gilardii are still unclear due to the lack of clinical and sequencing data. We need to obtain insight into the clinical characteristics, virulence, and resistance mechanisms of C. gilardii.
Patients and Methods: We isolated five C. gilardii isolates from hospitalized patients and carried out assay, culture and genome sequencing. We analyzed the genomic features of clinical C. gilardii isolates and took insight into their clinical characteristics, virulence, and resistance mechanisms.
Results: These isolates were resistant to meropenem, gentamicin, and other antimicrobials due to intrinsic resistance genes. Furthermore, the sequencing results revealed the widespread presence of the MCR-5.1 gene in C. gilardii. The virulence magnitude of C. gilardii is closely correlated with the number of virulence factors they carry. Some C. gilardii strains can acquire resistance to levofloxacin through gyrA gene mutation during treatment. The diverse antimicrobial resistance mechanisms challenge the treatment of C. gilardii infections.
Conclusion: We present the genomic characteristics of clinically isolated C. gilardii to improve (i) our understanding of this pathogen and (ii) treatment options.
Keywords: Cupriavidus gilardii, clinical characteristics, antimicrobial resistance, MCR-5.1, gyrA
Introduction
Cupriavidus gilardii is an aerobic, Gram-negative, non-glucose-fermenting bacterium belonging to the family Burkholderia of the phylum Betaproteobacteria, characterized by copper resistance, a rod shape, and motility.1–3 C. gilardii is widespread in many environments, including water and heavy metal-contaminated plants and soil.4–6 As microbial culture and identification technology advance, the increasingly successful detection of C. gilardii has gradually attracted clinical attention.7,8 C. gilardii had been isolated and identified from a variety of clinical samples, including cerebrospinal fluid, bone marrow, wounds, and respiratory secretions.9 There are currently no recommendations for antimicrobial use to treat C. gilardii infections.8 Moreover, the high case fatality rate of C. gilardii infections may be related to the characteristics of acquired drug resistance.8,10 Compared with other clinically well-known pathogens, we know very little about the genome, clinical characteristics, and therapy of C. gilardii. To date, only six clinical infection cases have been reported,10–15 and 11 non-clinical C. gilardii strains have been sequenced. The genomic differences between the clinical and environmental isolates still remain to be explored.
C. gilardii was first thought to be an opportunistic pathogen,10–12 but recent studies have shown that C. gilardii infection also exists in immunocompetent patients.13,14 Predisposing factors for C. gilardii infection remain unclear. C. gilardii is intrinsically resistant to many antimicrobials and could acquire drug resistance with continued antimicrobial therapy.10,13 The acquired drug resistance has been reported to inhibit the efficacy of antimicrobials and even lead to fatal consequences.10 Therefore, it is critical to determine the clinical infection characteristics and drug resistance mechanisms of C. gilardii to reduce the case fatality rate.
We found four cases of C. gilardii infection in our daily work recently, and all four patients were seriously ill. In the present study, we cultured and isolated the microbes from the four patients and analyzed their clinical characteristics. Moreover, the microbes were identified and whole-genome sequencing was performed. Their virulence, and drug resistance were studied and compared with those of other environmental isolates.
Materials and Methods
Clinical Characteristics
During the period from July 2020 to January 2021, four patients from different places in China were hospitalized in the Respiratory Intensive Care Unit (RICU) in Beijing Chao-Yang Hospital. Demographic and clinical data were obtained from electronic medical records, including personal history, comorbidities, symptoms, laboratory tests, length of stay in the RICU, severity scores (SOFA, APACHE II), outcomes, and follow-up.
Isolation and Identification
From all four patients, we collected five body liquid samples (Figure 1A) and cultured the bacteria on blood agar plates under aerobic conditions for 24h at 37°C. Colonies were picked for identification using a VITEK MS Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (database version 3.2, bioMérieux Inc.). The minimum inhibitory concentration (MIC) assay was performed using broth microdilution methods per Clinical and Laboratory Standards Institute last guideline [CLSI M100Ed30].16 The other antibiotics were tested using Disk Diffusion Assay (Kirby Bauer).
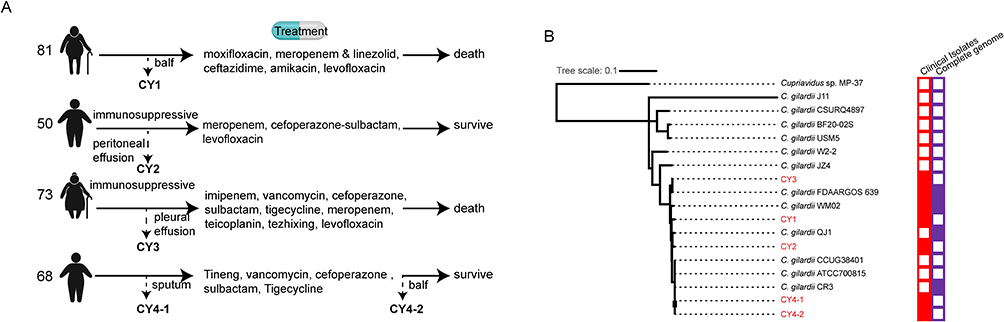 |
Figure 1 Information on the clinical and genetic background of the five sequenced C. gilardii isolates. (A) Schematic diagram of the treatment process of the four patients. Dashed lines with arrows indicate where/when samples were collected and sample names. The used antimicrobials are listed in sequence. (B) Phylogenetic tree of the 16 C. gilardii strains based on the SNPs called using parSNP. Red and purple boxes indicate that the sample is clinically sourced and completely assembled, respectively. The strains sequenced in this study are shown in red font. |
Genome Extraction and Sequencing
Genomic DNA was extracted using Easy-Pure Bacteria Genome DNA Kit (TransGen Biotech) and quantified by a Qubit® 2.0 Fluorometer (Thermo Scientific). The whole genomes of the five C. gilardii isolates were sequenced using an Illumina NovaSeq PE150 platform at Beijing Novogene Bioinformatics Technology Co., Ltd., according to the library construction protocols of Illumina.
Genome Assembly and Analyses
The raw sequencing reads were trimmed with Trimmomatic (v0.32) and then assembled using SPAdes (v3.10.0)17 and SOAPdenovo.18 The assembly results of the two software programs were integrated with CISA. We filtered the fragments below 500 bp. Annotation was performed using Prokka.19 The annotated proteins were used to identify genes involved in antimicrobial resistance based on the CARD database (v3.2.2) using BLAST (v2.9). We only considered the genes with >70% coverage and ≥70% identity. A pathogen–host interactions database was used to analyze pathogenicity. Comparative genomic analysis was performed with a custom script that integrated BLAST (v2.9). The phylogenetic tree was generated using parSNP.20
Results
Clinical Characteristics
The clinical characteristics of the four patients (three males and one female) infected with C. gilardii are listed in Table S1. Their age ranged from 50 to 84 years. Two of them were immunosuppressed (esophageal cancer), and the other two were not immunocompromised or immunosuppressed. Case reports are displayed as a flowchart in Figure 1A. We used advanced antimicrobials such as cefoperazone-sulbactam, carbapenems, and vancomycin at the early stage for all four patients, but treatment was not effective. This may be correlated with the inherent resistance of C. gilardii. The treatment methods were adjusted, and of the four patients, two survived. Three patients got purulent infections, including empyema, mediastinal abscess, and lung abscess. The values of IgG/IgM, CD4+/8+ T cells, and CD19+ B cells were lower in the two patients who died than in the two patients who survived.
Identification and Antimicrobial Susceptibility of C. gilardii Isolates
To characterize the pathogens, we cultured the pathogens from the body fluid (bronchoalveolar lavage fluid [BALF], peritoneal effusion, pleural effusion, and sputum) after the early treatment stage on blood agar plates. The MALDI-TOF MS assay results showed that the bacteria were C. gilardii (named strains CY1, CY2, CY3, and CY4-1). No other organisms were cultured.
Furthermore, we sampled the RICU environment (pool, ventilator, dining table, bed rails, bedside) during treatment, but all C. gilardii tests were negative, excluding the possibility of nosocomial transmission.
Interestingly, CY4-1 was initially sensitive to levofloxacin but acquired resistance during treatment. Thus, we re-cultured the drug-resistant bacterium from the sputum after treatment and named it C. gilardii strain CY4-2 (Figure 1A). For these five C. gilardii strains, the MIC was subsequently assayed using broth microdilution methodology (Table 1). The antimicrobial susceptibility profiles of C. gilardii strains showed a certain degree of resistance to 12 antimicrobials, including imipenem (IMI), ceftazidime (TAZ), cefepime (FEP), ceftazidime/avibactam (CZA), amikacin (AMI), meropenem (MERO), ceftolozane/tazobactam (C/T), aztreonam (AZT), piperacillin/tazobactam (P/T), tobramycin (TOB), colistin (COL) and ampicillin (AMP). All strains are resistant to Imipenem, different from environmental isolates but consist with some clinical isolates.10,12 Both CY4-1 and CY4-2 showed resistance to colistin. CY4-2 was the only strain that was resistant to levofloxacin.
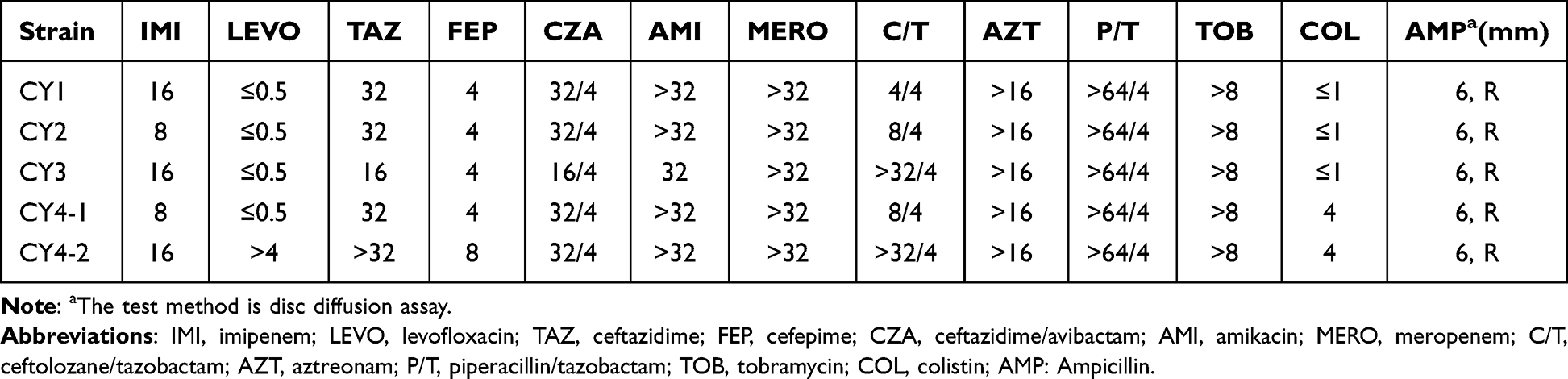 |
Table 1 Antimicrobial Agent Susceptibilities |
Genome Sequencing and Assembly of the Five C. gilardii Strains
We sequenced the genomes of the five C. gilardii strains using a PE150 NovaSeq system and analyzed the data. After assembly and annotation, we characterized the sequencing data and genome features (Table 2). The clean data sizes of all strains that could be used to assemble fragments were similar, yielding an average coverage depth of approximately 100× for each isolate. Due to the existence of repeat regions (especially the ribosomal RNA of C. gilardii) and the absence of long read sequencing, we could not assemble the complete genomes of the five C. gilardii strains with only NovaSeq sequencing data. However, this did not interfere with the analysis of the components and functions of the genomes. The number of coding sequences and GC content differed little between strains.
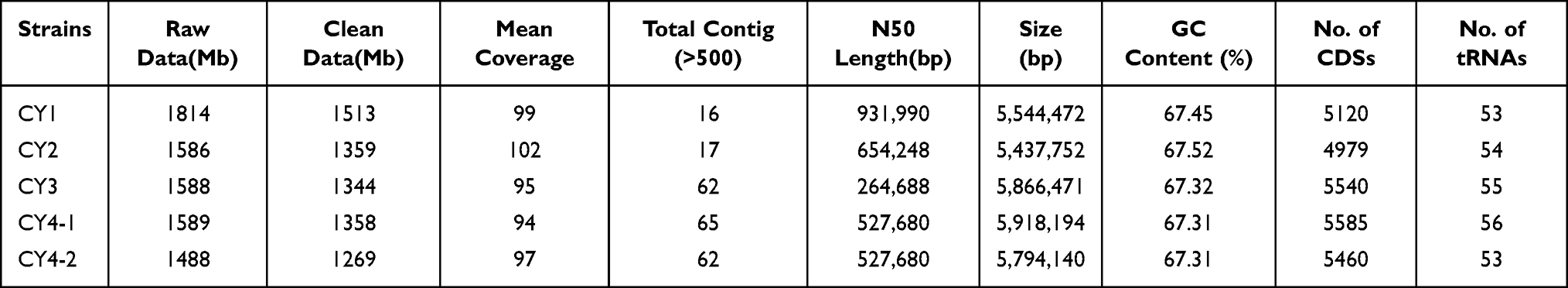 |
Table 2 Sequencing and Assembly Information of the Five C. gilardii |
Whole-Genome Phylogenetic Analysis
To analyze the genome similarity and the evolutionary relationships to other C. gilardii strains, we constructed a single-nucleotide polymorphism (SNP)-based phylogenetic tree using all reported C. gilardii genomes (12 in total) and Cupriavidus sp. MP-37 as outgroup species (Figure 1B). Most of the 12 reported genomes were drafted, and only four were complete genomes. Additionally, C. gilardii FDAARGOS 639 and WM02 were described to be a clinical isolate. All five sequenced strains were grouped into C. gilardii. CY4-1/2 showed a very high similarity (98%) to C. gilardii CR3, which was isolated from a tar pit. CY1, CY2, and CY3 were different from each other. The SNP-based phylogenetic tree showed that the infection sources are different. Since clinical case reports of C. gilardii infection are rare, attentions should be paid to the appearance of four cases in such a short period.
Analysis of Antimicrobial Resistance Genes
C. gilardii, despite being an opportunistic pathogen, has intrinsic resistance to many antimicrobials. Clinical treatment of C. gilardii is challenged by the multidrug resistance. From the assembled genomes and annotations, we identified the antimicrobial resistance genes of the five clinical C. gilardii isolates (Table 3). Genes sharing at least 70% coverage and 70% identity to genes in the CARD antimicrobial resistance gene database were identified using BLASTP. Nine genes were found, including five genes encoding efflux pump systems (MuxC/B, MexB/D, and ceoB), three intrinsic resistance genes (OXA-837, AAC (3)-IVb, and ANT(3”)-Ib), and the colistin resistance gene MCR-5.1. Interestingly, the antimicrobial was changed during treatment. MCR-5.1 was found in many other C. gilardii strains present in a transposon. We compared the neighboring regions of the MCR-5.1 gene in C. gilardii strains (Figure 2). Six out of the 12 available C. gilardii genomes were found to possess MCR-5.1, of which WM02, QJ1 and CR3 were completely assembled. The adjacent regions of MCR-5.1 in CY4-2 are highly homologous to those of ATCC700815 and CCUG38401. Genes mdtH, chrB1, and hin always co-exist with the MCR-5.1 gene. Segments with this combination of genes are present in a variety of E. coli plasmids, such as pSGMCR103 and pEC1897-13, and on the C. gilardii QJ1 plasmid plas1. Furthermore, this segment was integrated twice into the QJ1 genome, adjacent and in opposite directions. Interestingly, we did not identify any apparent presence of plasmids in CY4-1 and CY4-2 based on the sequencing results. The integration site in CSURQ4897 is different from those in other strains, which is consistent with the phylogenetic tree in Figure 1. This suggests that the MCR-5.1 gene may be integrated into bacterial genomes via E. coli-like plasmids. The integration site in the C. gilardii genome varies depending on genetic differences.
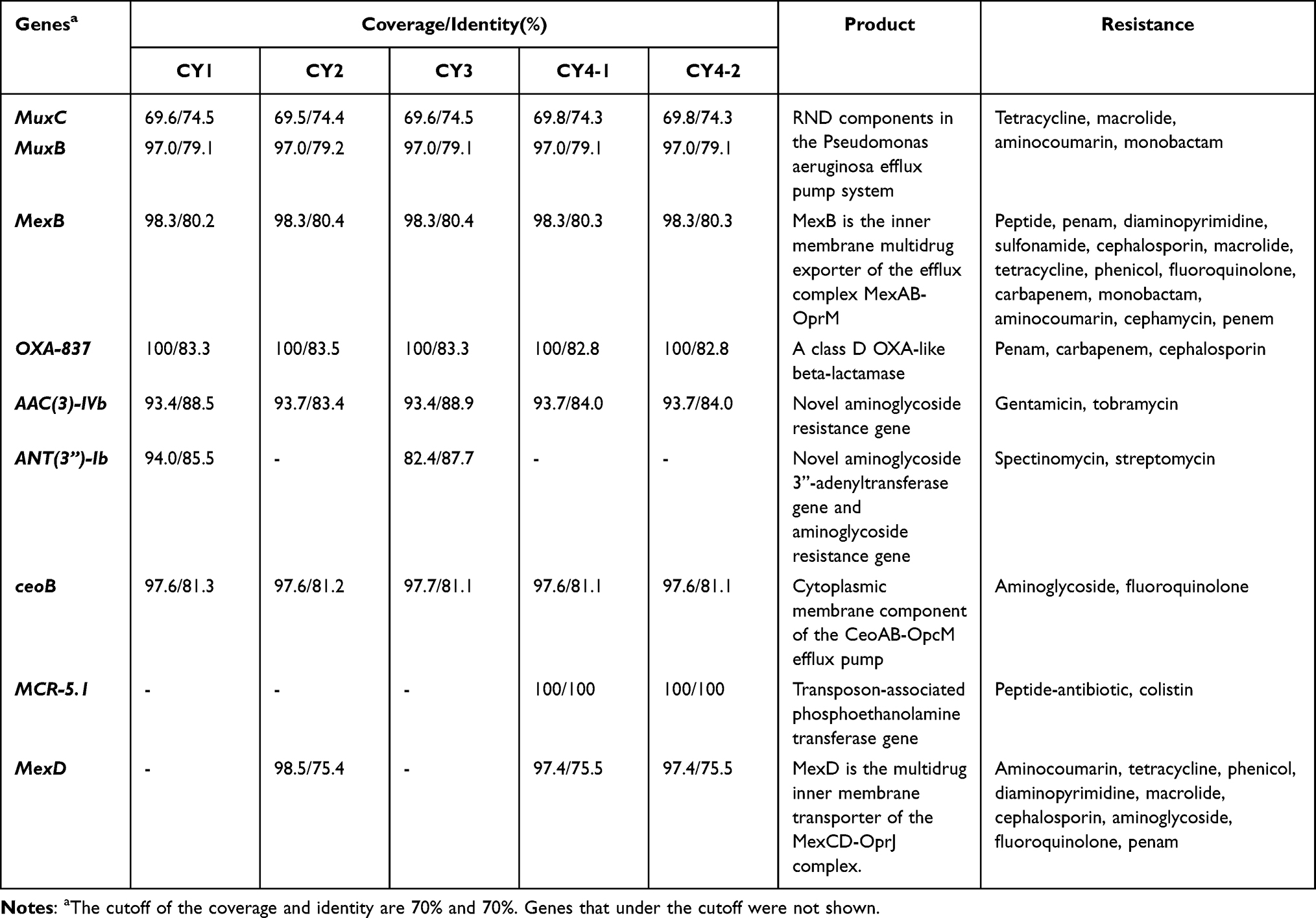 |
Table 3 Antimicrobial Genes of the Five Sequenced C. Gilardii |
 |
Figure 2 Genomic regions near the MCR-5.1 gene. Homologies between different strains are linked by light green quadrilaterals. MCR-5.1 genes are highlighted in red. |
According to the literature,21 gyrA and gyrB are levofloxacin resistance-related genes. We compared the sequences of gyrA and gyrB from all C. gilardii strains (Figure 3A). The results demonstrated that there are mutations I83T and T514S in gyrA during the treatment, however, no mutation in gyrB were observed in gyrB, which may account for the reason for levofloxacin resistance (Table 3). This acquisition of resistance occurred during a 2-week treatment. The analysis of this longitudinal sample implied the emergence of acquired drug resistance of bacteria during the treatment, which may partially interrogate the high mortality rate of C. gilardii infection.
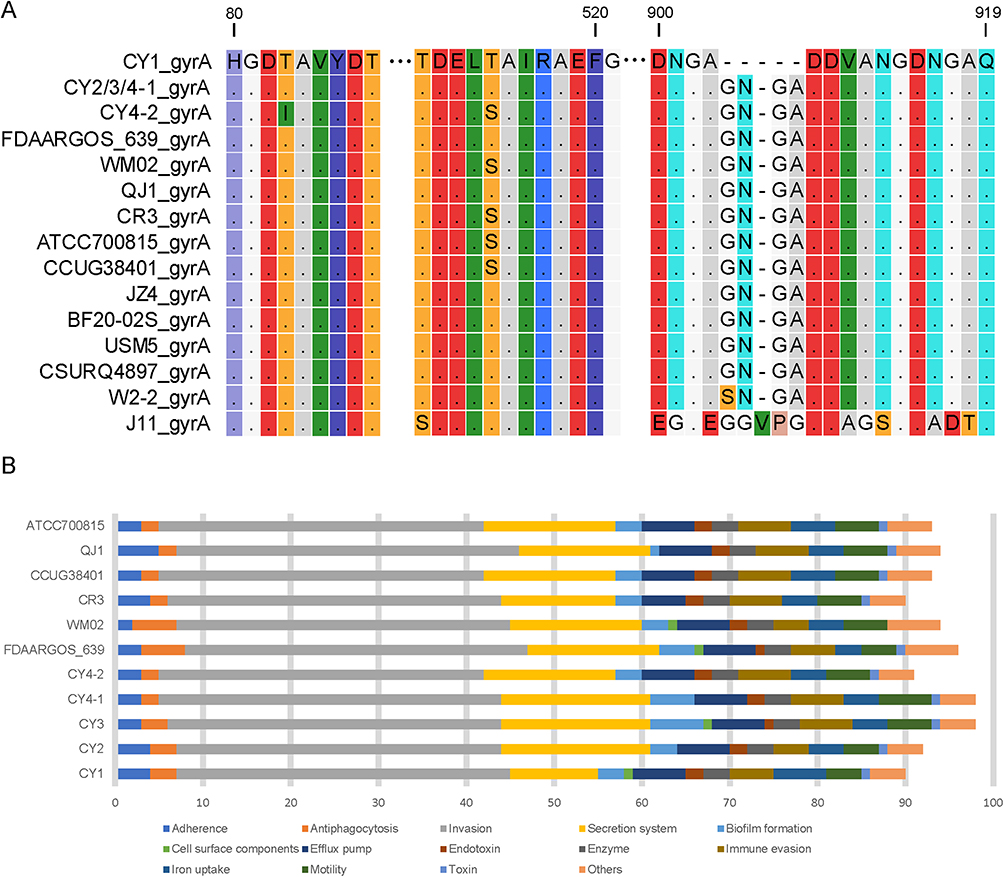 |
Figure 3 The resistant and virulent gene comparison of multiple C. gilardii strains. (A) Gene mutations in resistant gene gyrA. The amino acids in mutated regions are shown, while non-mutated ones are hidden. Different amino acids are distinguished by different colors. (B) the virulence factor counts of C. gilardii strains. The VF categories were marked with different colors. The x-axis represents the number of genes, while the y-axis represents the strains. |
Analysis of Virulence Factors
Virulence factors (VFs) in bacteria, including those involved in adhesion, invasion, toxin production, and immune evasion, are closely related to bacterial toxicity. The five strains isolated in this study showed different virulences, and two of them caused death (CY1 and CY3). We screened for known/potential VFs of 10 strains (five strains in this study and five closest strains in Figure 1A) from the latest virulence factor database22 using VFanalyzer.23 The VFs numbers were counted according to different VF categories (Figure 3B). The total number of VFs ranged from 90 to 98. Both CY4-1 and CY3 had 7 more VFs than Cy4-2, mainly including biofilm formation and secretion system. Cy4-1 and Cy4-2 were isolated from the same patient, among which Cy4-2 was obtained after the patient recovered. Judging from the clinical data (Table S1), CY3 was also highly toxic. Interestingly, both CY3 and CY1, which caused the host patients died, have cell surface components. Though FDAARGOS 639 has cell surface components, there is no information to indicate its toxic in clinic. In general, the virulence magnitude of C. gilardii is closely correlated with the number of virulence factors they carry, with biofilm formation, secretion system, and cell surface components may play an important role.
Discussion
C. gilardii is an opportunistic pathogen that is widely present in the environment and has potential use in the detoxification of heavy metals and other biotechnological applications.24–27 There are increasing case reports of infections caused by C. gilardii, even fatal cases.10 However, the clinical analysis of C. gilardii infection is still relatively lacking. There are two clinical genomes available to date. One is C. gilardii strain FDAARGOS_639 reported by the US Food and Drug Administration in 2020 (currently a representative genome), but with no clinical information. The other one is C. gilardii strain WM02 reported recently from an acute non-lymphocytic leukemia patient from China, also with no clinical information. To our knowledge, the four cases included in this study are the largest sample size so far for this rare infectious disease. We isolated and sequenced five strains from different body liquids, enriching the genetic data of C. gilardii and providing insight into the genetic differences between clinical and environmental strains. The clinical characteristics, treatment and outcome were explored to gain a deeper understanding of this disease.
Interestingly, three of the four patients got abscess (empyema and mediastinal abscess for the esophageal cancer patients and lung abscess for an immunocompetent patient) during the treatment, indicating there is a risk of purulent infection after C. gilardii infection. Abscess caused by C. gilardii infection has also been previously reported.12,13 Tena et al reported a renal transplant recipient developed a muscular abscess caused by C. gilardii. Similarly, C. gilardii were isolated from the wound and blood culture in a case of pacemaker-associated infection. Thus, our study showed a representative clinical characteristic similar to that in other studies.
The mortality rate of our C. gilardii infection was also high. Thus, we compared the different characteristics between survivors and non-survivors. The results demonstrated that the values of IgG/IgM, CD4+/8+ T cells, and CD19+ B cells were lower in the fatal patients, suggesting the possibility of potential immunosuppression in deceased patients, which was first described by our group. Given the relatively small sample size, further studies are needed to determine the role of immunity in the C. gilardii infection.
Since several studies have reported unsatisfied clinical effect of multiple antibiotics,10–14 we discussed the potential drug resistance genes based on the sequencing data. C. gilardii strains CY4-1/2 were genetically highly homologous to environmental isolates CR3 (tar pit, China), CCUG38401 (whirlpool, Sweden), and ATCC700815 (whirlpool, USA). Both carried the colistin resistance gene MCR-5.1, indicating that the source of infection in this patient may be a C. gilardii strain that is present in the environment. Besides, we found that almost half of the C. gilardii strains carried the MCR-5.1 gene, indicating that a high probability of acquiring the resistance gene through gene transfer. C. gilardii is naturally resistant to a variety of antimicrobials,28 and these intrinsic genes were also found in the present study, such as OXA-837, AAC (3)-IVb, and ANT (3)-Ib. In addition, we further confirmed that during the treatment process, the bacteria also acquired resistance to certain antimicrobials through gene mutation. In conclusion, C. gilardii has diverse antimicrobial resistance mechanisms.
We investigate the virulence of C. gilardii. Investigations on C. gilardii virulence factors are limited to the VFs counts and TTSS gene on individual trains. We conducted a screening of known and potential VFs in 10 strains, aiming to establish a correlation between the count of VFs and the clinically observable toxicity, particularly in the case of CY4-1 and CY4-2. However, despite our efforts, we have not yet identified a specific factor that plays a significant role in the virulence of C. gilardii among the numerous VFs assessed. Further assessments are required to unveil the underlying mechanism of virulence in C. gilardii.
At present, there are still difficulties in clinical identification and treatment of C. gilardii. The high-throughput sequencing method used here to study the clinically isolated C. gilardii strains can accurately identify its drug resistance genes and elucidate the mechanisms underlying acquired drug resistance, guiding antimicrobial treatment. Though the sample size in the present study was small and the genome assemblies are drafts, we successfully investigated the pathogenicity and drug resistance mechanisms. For the clinical treatment of C. gilardii infection, tetracyclines, ceftazidime, and cefepime are recommended first. Advanced antimicrobials such as meropenem and imipenem are not recommended.13,14,29 Sequencing methods to identify resistance genes and predict drug resistance can guide antimicrobial treatment, reducing the abuse of antimicrobials and improving therapy.
Conclusion
Collectively, our study represented four clinical cases of C. gilardii infection and conducted a high-throughput sequencing of the isolated C. gilardii strains. By identifying the clinical and genetic characteristics of this rare infectious disease, as well as the possible genes mediated the drug resistance, our work enhances the understanding of the disease and provides some directions for rational antimicrobial treatment. Further researches need to be performed to elucidate the pathogenic mechanism and the significant virulence genes of C. gilardii infection and to explore new therapeutic targets.
Abbreviations
RICU, Respiratory Intensive Care Unit; SOFA, Sequential Organ Failure Assessment; APACHE II Acute Physiology And Chronic Health evaluation II.
Data Sharing Statement
The draft assembled genomes of C. gilardii strains CY1, CY2, CY3, CY4-1, and CY4-2 have been deposited in the National Center for Biotechnology Information (NCBI) GenBank database (GenBank accession numbers JAODVC000000000, JAODVD000000000, JAODVE000000000, JAODVF000000000, and JAODVG000000000). The raw sequencing reads were shared through NCBI-SRA with the bioproject IDs: PRJNA881018.
Ethical Approval
The ethics committee of the affiliated Beijing Chao-Yang Hospital, Capital Medical University approved this study (2021-ke-501). This study was conducted in accordance with the Declaration of Helsinki. Written informed consent was waived due to the retro spective noninterventional design.
Acknowledgments
We thank Dr. Fengshuang Yi and Dr. Song Nan from Beijing Institute of Respiratory Medicine, Capital Medical University for the diagnosis of respiratory pathogens for these patients; Dr. Shengcai Zheng from Capital Medical University for the English revision of this article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Ministry of Science and Technology of the People’s Republic of China [grant number 2020YFC0844600]; National Natural Science Foundation of China [grant number 82200007]; Beijing Hospitals Authority Youth Programme [grant number QML20190301] and Development Program of Beijing Institute of Respiratory Medicine (Grant no. Ggyfz202329).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Coenye T, Falsen E, Vancanneyt M, et al. Classification of Alcaligenes faecalis-like isolates from the environment and human clinical samples as Ralstonia gilardii sp. nov. Int J Syst Bacteriol. 1999;49(Pt 2):405–413. doi:10.1099/00207713-49-2-405
2. Vaneechoutte M, Kämpfer P, De Baere T, Falsen E, Verschraegen G. Wautersia gen. nov., a novel genus accommodating the phylogenetic lineage including Ralstonia eutropha and related species, and proposal of Ralstonia [Pseudomonas] syzygii (Roberts et al. 1990) comb. nov. Int J Syst Evol Micr. 2004;54(Pt 2):317–327. doi:10.1099/ijs.0.02754-0
3. Huang N, Mao J, Zhao Y, Hu M, Wang X. Multiple transcriptional mechanisms collectively mediate copper resistance in Cupriavidus gilardii CR3. Environ Sci Technol. 2019;53(8):4609–4618. doi:10.1021/acs.est.8b06787
4. Harmon DE, Miranda OA, McCarley A, Eshaghian M, Carlson N, Ruiz C. Prevalence and characterization of carbapenem-resistant bacteria in water bodies in the Los Angeles-Southern California area. Microbiologyopen. 2019;8(4):e00692. doi:10.1002/mbo3.692
5. Huang N, Mao J, Hu M, Wang X, Huo M. Responses to copper stress in the metal-resistant bacterium Cupriavidus gilardii CR3: a whole-transcriptome analysis. J Basic Microb. 2019;59(5):446–457. doi:10.1002/jobm.201800693
6. Tanaka R, Nouzaki K, Navarro RR, et al. Activated sludge microbiome in a membrane bioreactor for treating Ramen noodle-soup wastewater. J Gen Appl Microbiol. 2021;66(6):339–343. doi:10.2323/jgam.2020.01.006
7. Kweon OJ, Lim YK, Kim HR, Kim TH, Ha SM, Lee MK. Isolation of a novel species in the genus Cupriavidus from a patient with sepsis using whole genome sequencing. PLoS One. 2020;15(5):e0232850. doi:10.1371/journal.pone.0232850
8. Cherak Z, Loucif L, Ben KM, et al. MCR-5-producing colistin-resistant Cupriavidus gilardii strain from well water in Batna, Algeria. Msphere. 2021;6(5):e0057521. doi:10.1128/mSphere.00575-21
9. Coenye T, Vandamme P, LiPuma JJ. Infection by Ralstonia species in cystic fibrosis patients: identification of R. pickettii and R. mannitolilytica by polymerase chain reaction. Emerg Infect Dis. 2002;8(7):692–696. doi:10.3201/eid0807.010472
10. Karafin M, Romagnoli M, Fink DL, et al. Fatal infection caused by Cupriavidus gilardii in a child with aplastic anemia. J Clin Microbiol. 2010;48(3):1005–1007. doi:10.1128/JCM.01482-09
11. Wauters G, Claeys G, Verschraegen G, et al. Case of catheter sepsis with Ralstonia gilardii in a child with acute lymphoblastic leukemia. J Clin Microbiol. 2001;39(12):4583–4584. doi:10.1128/JCM.39.12.4583-4584.2001
12. Tena D, Losa C, Medina MJ, Sáez-Nieto JA. Muscular abscess caused by Cupriavidus gilardii in a renal transplant recipient. Diagn Microbiol Infec Dis. 2014;79(1):108–110. doi:10.1016/j.diagmicrobio.2014.01.023
13. Kobayashi T, Nakamura I, Fujita H, et al. First case report of infection due to Cupriavidus gilardii in a patient without immunodeficiency: a case report. BMC Infect Dis. 2016;16(1):493. doi:10.1186/s12879-016-1838-y
14. Zhang Z, Deng W, Wang S, Xu L, Yan L, Liao P. First case report of infection caused by Cupriavidus gilardii in a non-immunocompromised Chinese patient. Idcases. 2017;10:127–129. doi:10.1016/j.idcr.2017.10.009
15. Fang X, Lu K, Sun S, Zhang H, Yu X, Han W. Infection caused by Cupriavidus gilardii in a convalescent COVID-19 patient. Int J Infect Dis. 2023;134:287–289. doi:10.1016/j.ijid.2023.07.015
16. Limbago B. M100-S11, Performance standards for antimicrobial susceptibility testing. Clin Microbiol Newsl. 2001;23(6):49. doi:10.1016/S0196-4399(01)88009-0
17. Bankevich A, Nurk S, Antipov D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19(5):455–477. doi:10.1089/cmb.2012.0021
18. Luo R, Liu B, Xie Y, et al. SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience. 2012;1(1):18. doi:10.1186/2047-217X-1-18
19. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. doi:10.1093/bioinformatics/btu153
20. Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15(11):524. doi:10.1186/s13059-014-0524-x
21. Miyachi H, Miki I, Aoyama N, et al. Primary levofloxacin resistance and gyrA/B mutations among Helicobacter pylori in Japan. Helicobacter. 2006;11(4):243–249. doi:10.1111/j.1523-5378.2006.00415.x
22. Liu B, Zheng D, Zhou S, Chen L, Yang J. VFDB 2022: a general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022;50(D1):D912–D917. doi:10.1093/nar/gkab1107
23. Liu B, Zheng D, Jin Q, Chen L, Yang J. VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019;47(D1):D687–D692. doi:10.1093/nar/gky1080
24. Wang X, Chen M, Xiao J, et al. Genome sequence analysis of the naphthenic acid degrading and metal resistant bacterium Cupriavidus gilardii CR3. PLoS One. 2015;10(8):e0132881. doi:10.1371/journal.pone.0132881
25. Lafi FF, Bokhari A, Alam I, Bajic VB, Hirt H, Saad MM. Draft genome sequence of the plant growth-promoting Cupriavidus gilardii strain JZ4 isolated from the desert plant tribulus terrestris. Genome Announc. 2016;4(4). doi:10.1128/genomeA.00678-16
26. Wu X, Wang W, Liu J, et al. Rapid biodegradation of the herbicide 2,4-Dichlorophenoxyacetic acid by Cupriavidus gilardii T-1. J Agr Food Chem. 2017;65(18):3711–3720. doi:10.1021/acs.jafc.7b00544
27. Pan D, Xu Y, Ni Y, Zhang H, Hua R, Wu X. The efficient persistence and migration of Cupriavidus gilardii T1 contribute to the removal of MCPA in laboratory and field soils. Environ Pollut. 2022;304:119220. doi:10.1016/j.envpol.2022.119220
28. Ruiz C, McCarley A, Espejo ML, Cooper KK, Harmon DE, Castanheira M. Comparative genomics reveals a well-conserved intrinsic resistome in the emerging multidrug-resistant pathogen Cupriavidus gilardii. Msphere. 2019;4(5). doi:10.1128/mSphere.00631-19
29. Massip C, Coullaud-Gamel M, Gaudru C, et al. In vitro activity of 20 antibiotics against Cupriavidus clinical strains. J Antimicrob Chemoth. 2020;75(6):1654–1658. doi:10.1093/jac/dkaa066
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.