Back to Journals » Infection and Drug Resistance » Volume 15
Genomic and Phenotypic Characterization of a Colistin-Resistant Escherichia coli Isolate Co-Harboring blaNDM-5, blaOXA-1, and blaCTX-M-55 Isolated from Urine
Authors Hao J, Zeng Z, Xiao X, Ding Y, Deng J, Wei Y ![]() , Liu J
, Liu J
Received 31 December 2021
Accepted for publication 24 February 2022
Published 29 March 2022 Volume 2022:15 Pages 1329—1343
DOI https://doi.org/10.2147/IDR.S355010
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Jingchen Hao,1,* Zhangrui Zeng,1,* Xue Xiao,2 Yinhuan Ding,1 Jiamin Deng,3 Yueshuai Wei,1 Jinbo Liu1
1Department of Laboratory Medicine, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, Sichuan, People’s Republic of China; 2Department of Laboratory Medicine, Southwest Medical University, Luzhou, 646000, Sichuan, People’s Republic of China; 3Department of Respiratory and Critical Care Medicine, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, Sichuan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jinbo Liu, Department of Laboratory Medicine, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China, Tel/Fax +86 830 3165730, Email [email protected]
Background: Colistin is one of the few options for treating carbapenem-resistant Enterobacterales (CREs). There is little available information about the epidemic status of colistin-resistant CREs in Southern Sichuan, China. This study mainly investigated the genomic and phenotypic characteristics of an extensively drug resistant E. coli LZ00114 isolated from Luzhou, China.
Materials and Methods: In 2020, LZ00114 was isolated from the urine of a patient with hydronephrosis and urinary tract infection in Luzhou, China. We assessed the resistance profile of LZ00114 in the presence and absence of the protonophore carbonyl cyano-m-chlorophenylhydrazine (CCCP) and 1-(1-naphthylmethyl)-piperazine (NMP) by antimicrobial susceptibility testing. The growth kinetics, motility, and pathogenicity of LZ00114 were determined to evaluate its microbial characteristics. In combination with whole genome sequencing (WGS) and real-time reverse transcription PCR (RT-PCR), we comprehensively analyzed the resistance mechanisms of LZ00114.
Results: LZ00114 was resistant to various antimicrobial agents, including meropenem, tetracycline, ciprofloxacin, gentamicin, fosfomycin, and polymyxin B. Notably, CCCP reversed the resistance of LZ00114 to polymyxin B. LZ00114 displayed high pathogenicity in the infection model (P< 0.01) compared with the Lab-WT strain, and its growth rate and motility were not significantly different from the Lab-WT strain. WGS and conjugation revealed that LZ00114 belonged to ST410 and carried a blaNDM-5-harboring self-transmissible IncX3 plasmid and a multi-replicon IncFII/FIA/FIB plasmid carrying blaOXA-1-blaCTX-M-55-tet(B)-aac(6’)-Ib-cr-dfrA17-sul1-fosA3. Comparative genomics revealed genetic relatedness between LZ00114 and strains isolated from other regions. Furthermore, there were point mutations in pmrA (S29G, G144S), pmrB (D283G, Y358N), marR (G103S, Y137H), emrA (I219V), and emrD (G323D) of LZ00114. RT-PCR confirmed the overexpression of efflux pumps and PmrABC in LZ00114.
Conclusion: This study provides valuable information for the surveillance of antimicrobial resistance and a theoretical basis for the prevention and control of colistin-resistant E. coli. There is still a need to be vigilant about the clone spread of the high-risk clone group E. coli ST410.
Keywords: E. coli, antimicrobial resistance, whole genome sequencing, colistin, carbonyl cyanide m-chlorophenylhydrazine, CCCP, ST410
Introduction
Escherichia coli is a common Gram-negative opportunistic pathogen that causes invasive host infections through virulence factors such as flagella, toxin secretion, and adhesins. According to the source of the infection, pathogenic E. coli can be classified as intestinal (diarrheagenic) and extraintestinal (ExPEC). Uropathogenic E. coli (UPEC) is the most common lineage of ExPEC. In hospitals, the likelihood of opportunistic infections is high in immunocompromised patients as well as those needing catheters.1 These pathogens adhere to the uroepithelium through the synergistic actions of various virulence factors and cause ascending infections of the bladder, kidneys, and blood by regulating the movement of bacterial pili.2,3 However, the emergence of antimicrobial resistance makes treating these bacterial infections a thorny problem.4 ST131 was previously considered the central lineage of antimicrobial resistance transmission in extraintestinal E. coli, with ST167 and ST410 gradually emerging as new high-risk clones.5–7 For these strains, carbapenemase production is the primary mechanism of carbapenem resistance in Enterobacterales. The worldwide epidemic of blaNDM-producing Enterobacterales has caused widespread concern. Based on NDM-1, several NDM variants with higher levels of carbapenem resistance have been identified.8 IncX and IncF plasmids are usually the primary mediators of blaNDM dissemination. They can carry various acquired antimicrobial resistance genes to promote the formation of multidrug-resistant bacteria and expand the host range. Although fitness cost usually accompanies acquired resistance, bacteria can reduce this effect through additional chromosomal mutation, epistatic effects, and environmental factors.9 Growth, motility, and pathogenicity are often important factors in evaluating the resistance cost of resistant bacteria. Therefore, it is necessary to study the microbial characteristics of these pathogens to assess the alterations in their fitness.
Avibactam, a new β-lactamase inhibitor, does not inhibit the activity of metallo-beta-lactamases (MBLs), and colistin is one of the primary drugs for treating MBLs-producing MDR and XDR resistant Enterobacterales. Therefore, once the MBL-producing CRE develops colistin resistance, it will further limit antimicrobial therapy. In E. coli, colistin resistance is usually associated with the two-component system, namely, the PmrAB, PhoPQ-mediated lipopolysaccharide (LPS) modification, and the acquisition of exogenous mcr genes.10 For two-component mediated colistin resistance, other factors besides colistin can also select mutations, that is, colistin-resistant strains can be isolated from patients without colistin treatment.10 Furthermore, the efflux pump was associated with the formation of heteroresistance to colistin.11 In recent years, it has been found that the efflux pump inhibitor CCCP can reverse the colistin resistance of bacteria, but the exact mechanism needs to be further clarified.12 Therefore, more valuable information is needed to elucidate the colistin resistance mechanisms in Enterobacterales. In our study, we described the isolation of a colistin-resistant extensively drug-resistant E. coli isolate LZ00114 from urine, which to our knowledge is also the first report of colistin-resistant E. coli isolated from Southern Sichuan, China. In addition, we focus on its molecular genetics and microbial characteristics. The results of this study provide a theoretical basis for the prevention and control of colistin-resistant E. coli worldwide.
Materials and Methods
Bacterial Isolate
In April 2020, LZ00114 was isolated from a female patient with hydronephrosis and urinary tract infection in the Affiliated Hospital of Southwest Medical University (Luzhou, China). This isolate was identified by matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS) (Bruker Daltonik GmbH, Bremen, Germany) as E. coli. At that time, the clinical microbiology laboratory reported that LZ00114 was resistant to beta-lactams, carbapenems, fluoroquinolones, tetracyclines, chloramphenicol, and gentamicin. The patient was treated with tigecycline and showed improved symptoms of the infection. During hospitalization, the patient was not treated with colistin. To explore whether LZ00114 is also resistant to colistin and fosfomycin and to define its resistance mechanisms, as well as the microbial characteristics of uropathogenic E. coli, we performed a follow-up study.
Antimicrobial Susceptibility Testing
We tested the minimum inhibitory concentration (MIC) breakpoints of 14 antimicrobial agents according to the definition of extensive drug resistance.13 The MIC breakpoints of ampicillin, aztreonam, cefepime, ciprofloxacin, tetracycline, meropenem, imipenem, chloramphenicol, and tigecycline were determined using the MicroScan WalkAway System (Siemens, Germany). The microbroth dilution method based on the 96-well plate was used to determine the MIC value of polymyxin B. The MIC of fosfomycin was measured using the agar dilution method. The antimicrobial susceptibility results were interpreted according to CLSI 2020-M100.14 The MIC of tigecycline was interpreted using the guidelines of the European Committee on Antimicrobial Susceptibility Testing (2021, https://www.eucast.org/). The E. coli ATCC25922 strain served as the quality control.
Detection of Carbapenem Genes, Chromosome, and Plasmid-Mediated Colistin Resistance Genes
Total DNA was extracted from bacteria using the boiling method.15 In brief, blaKPC, blaNDM, blaVIM, blaIMP, blaOXA-48, pmrAB, phoPQ and pmrD genes were detected by polymerase chain reactions (PCR). All PCR primers used in this study were listed in Table S1. Positively amplified products were sequenced by the Sanger sequencing method and compared with the NCBI BLAST database (http://www.ncbi.nlm.nih.gov/BLAST/).
Efflux Pump Inhibition Assay
To assess the role played by efflux pumps in antimicrobial resistance, we evaluated the changes in MICs in the presence of efflux pump inhibitors, namely, CCCP (Sigma, USA) and NMP (Alfa Aesar, CN). The final concentrations of CCCP and NMP were 10 mg/L and 100 mg/L, respectively. A 4-fold or more reduction in the MICs after adding the efflux pump inhibitors was interpreted as a high expression of the efflux pump, and an 8-fold or more reduction in the MIC of colistin after adding CCCP was interpreted as colistin resistance reversal.16,17
Galleria mellonella Infection Model
To assess pathogenicity, the virulence of bacteria was modeled as previously described using Galleria mellonella larvae, with minor modifications.18 The concentrations of the bacterial suspensions were adjusted to 1×107 CFU/mL. Hamilton precision syringes were used to inject 10 µL of the bacterial suspension (the actual inoculation volume was 105 CFU) into 15 healthy larvae, followed by incubation at 37°C for 72 h in the dark. The number of nonviable larvae was recorded every 24 h. The Lab-WT E. coli ATCC25922 strain served as the negative control. All assays were conducted in triplicate.
Biofilm Formation Capacity, Growth Kinetics, and Motility Assays
The biofilm formation capacity of the isolate was measured by staining with 0.1% crystal violet as previously described with minor modifications.19 The plates were incubated at 37°C for 24 h, washed, fixed, stained, and solubilized with 95% ethanol. Subsequently, the absorbance was measured at 570 nm with a microplate reader (PerkinElmer, USA). The A. baumannii ATCC19606 and sterile LB broth served as positive and negative controls, respectively. The data were expressed as mean ± SD. All assays were conducted in triplicate.
For growth kinetics, briefly, 10 µL of 0.5 McFarland turbidity standard suspension was added to each well of a 96-well plate, followed by 190 µL of LB broth per well. Three replicate wells were set up for each strain. The absorbance was measured at 600 nm at 2 h intervals at 37°C.20 Motility tests were performed as previously described with minor modifications.21 In brief, 1.5 µL of 0.5 McFarland turbidity standard suspension was added to 0.25% tryptone agar plates. The plates were incubated for 16 h at 35°C. All assays were conducted in triplicate. The experimental results were compared with Lab-WT (Wild Type) E. coli ATCC25922.
Conjugation
E. coli J53 was used as the recipient, and LZ00114 E. coli was used as the donor. The two strains were added to antibiotic-free LB broth at a ratio of 2:1 and incubated for 24 h. MH plates containing sodium azide (180 µg/mL) and meropenem (4 µg/mL) were used to screen the transconjugants.
Whole Genome Sequencing and Analysis
Total genomic DNA was extracted using a magnetic-bead–based kit (Qiagen, Germany). The purified DNA was used to construct the library according to the NEBNext Ultra DNA Library preparation kits and sequenced using Illumina NovaSeq 6000 PE150 and Oxford nanopore platforms. Read sequences were de novo assembled using continuous long reads following Canu software (version 1.7). Prokka software (version 1.10) was engaged to predict the coding genes, tRNAs, and rRNAs in the assembled genome. The resistance genes, serotypes, and plasmid types were analyzed by the Center for Genomic Epidemiology (http://www.genomicepidemiology.org/), with the threshold for identity using the default settings for web pages. Virulence genes were identified by VirulenceFinder 2.0 (https://cge.cbs.dtu.dk/services/VirulenceFinder/) and virulence factor database (VFDB, http://www.mgc.ac.cn/VFs/). The BLAST (http://blast.ncbi.nlm.nih.gov), OAT software (v 0.931), and BRIG (BLAST Ring Image Generator) v0.95 software were used for comparative genomic analysis.22 The analysis of average nucleotide identity (ANI) used chromosome sequences of E. coli ST410 strains (WCHEC025943, PT109, and AMA1167) previously deposited into the NCBI database.23–25A phylogenetic tree based on single nucleotide polymorphisms (SNPs) was constructed by using the CSI Phylogeny 1.4 (https://cge.cbs.dtu.dk/services/CSIPhylogeny/). IslandPath-DIOMB (v 0.2) was used for genomic island prediction analysis.26 E. coli str. K-12 substr. MG1655 (GenBank accession number: NC_000913.3) served as a reference strain for chromosome point mutation analysis. All genomic data (LZ00114) were stored in the NCBI database under accession numbers: CP087570, CP087571, and CP087572.
Transcriptional Expression of Efflux Pumps and PmrAB Two-Component System by Quantitative Real-Time PCR
The expression levels of resistance-nodulation-cell division (RND) efflux pump genes (acrA, acrB, and acrD), major facilitator superfamily (MFS) efflux pump genes (emrA, emrD, and emrY), and the two-component regulatory system PmrAB were detected by RT-PCR. The selected purified colonies were inoculated in 5 mL of LB broth and cultured to the mid-exponential phase. Subsequently, total RNA was extracted using the RNA kit according to the reagent manufacturer’s recommendations. The design of the RT-PCR primers is provided in Table S1, with danE and gapA serving as internal reference genes. E. coli ATCC25922 served as the control strain. The relative expression of the indicated genes was calculated by the 2−ΔΔCt method. The data were expressed as mean ± SD, and statistical analysis was performed using Student’s t-test. All assays were conducted in triplicate.
Results
General Characteristics of LZ00114
LZ00114 was resistant to β-lactams, fluoroquinolones, aminoglycosides, polymyxin B (8 µg/mL), and other common antibiotics except for tigecycline (Table 1). PCR showed that LZ00114 carried the blaNDM-5 gene, but the mcr-1 gene was not identified. Using E. coli ATCC25922 as the Lab-WT reference strain, we tested the microbial characteristics of LZ00114. Growth kinetics showed that LZ00114 grew faster than the Lab-WT strain over 24 h, and the OD600 values of LZ00114 and the Lab-WT strain reached 0.6 at approximately 14 h and 16 h, respectively. As an indicator of the pathogenicity of uropathogenic E. coli, the motility of LZ00114 did not show significant differences compared with the Lab-WT strain. In the G. mellonella infection model, larvae inoculated with 105 CFU of LZ00114 showed a 46.7% mortality rate within 4 days, while those inoculated with 105 CFU of the Lab-WT strain showed a 6.7% mortality rate within 4 days (Figure 1). For biofilm formation capacity, the OD570 value of the negative control was 0.12±0.01, LZ00114, and A. baumannii ATCC19606 (positive control) exhibited moderately adherent (0.26±0.03, 2+) and strongly adhesion (0.57±0.05, 3+), respectively.
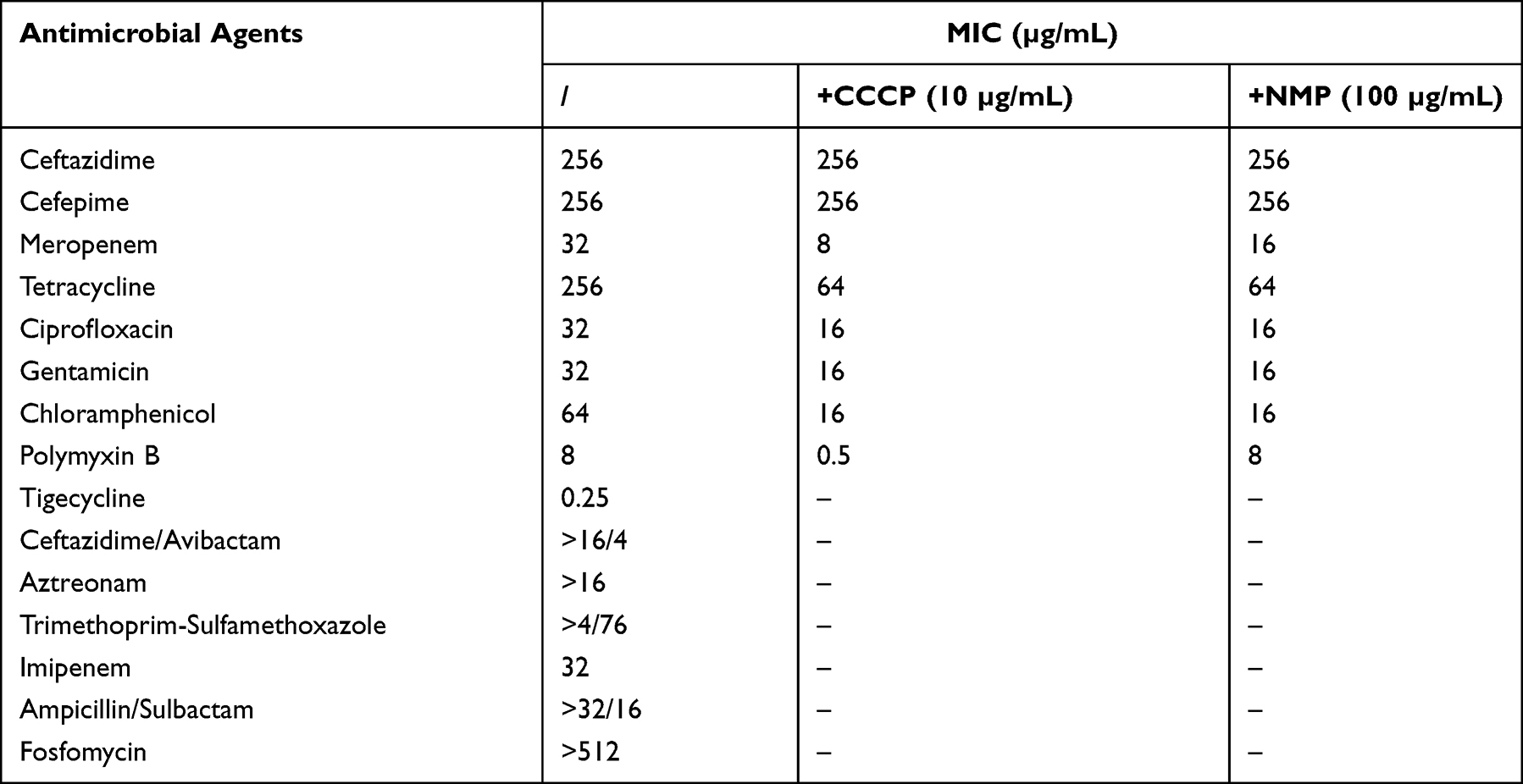 |
Table 1 Antimicrobial Susceptibility and Efflux Pump Inhibition Tests of LZ00114 |
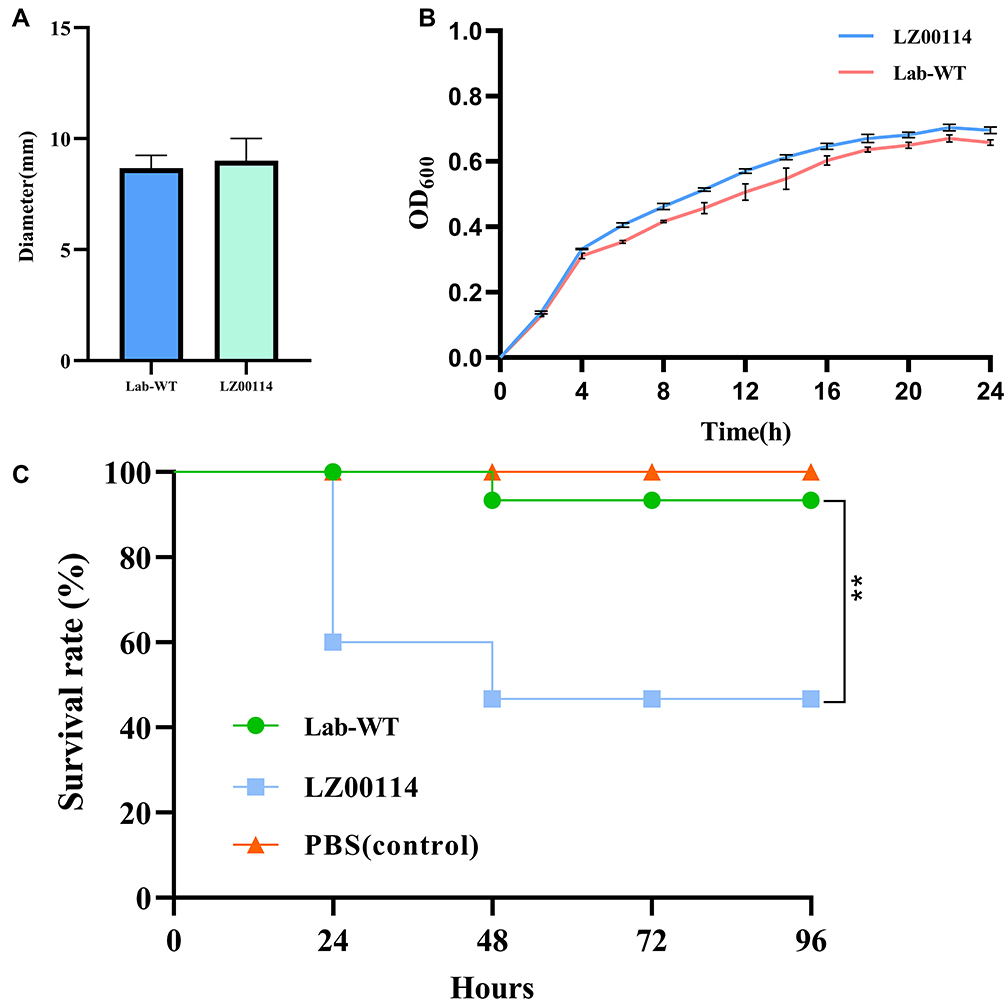 |
Figure 1 Microbial characteristics of LZ00114 were drawn by GraphPad Prism 9.2.0. (A) Motility test results of LZ00114. The data represents the average diameter of the swimming. (B) Growth dynamic curve of LZ00114 and Lab-WT E. coli ATCC25922 for 24h. (C) A total of three groups were allocated 15 G. mellonella larvae each. Two groups were infected with 105 CFU of LZ00114 and Lab-WT, respectively. The remaining group was injected with 10 µL PBS, which served as blank control. The error bar represents the standard deviation of the mean. **P < 0.01 by Log rank test. |
The LZ00114 genome consisted of three contigs of 4,953,723 bp containing a chromosome backbone (4,755,648 bp) and two circular plasmids. Furthermore, blaOXA-1 and blaNDM-5 were located on IncF (151,915 bp) and IncX3 (46,160 bp) plasmids, respectively. According to silico MLST analysis, the sequence type of LZ00114 belonged to ST410. Comparison of wzt, wzy, and flic genes by SerotypeFinder showed that the serotype was O8:H9 (identity>99.5%). Moreover, LZ00114 carried many virulence factors for iron uptake and adhesion. The Fim/fimH/fumC types, virulence factors, and acquired antimicrobial resistance genes are shown in Table 2. In particular, the chromosomal genome of LZ00114 showed highly similar ANIs (99.98%, 99.88%, and 99.99%, respectively) with E. coli ST410 isolate WCHEC025943 (China, GenBank accession number: CP027205.2), PT109 (Portugal, GenBank accession number: CP041031.1), and AMA1167 (Denmark, GenBank accession number: CP024801.1). Additionally, LZ00114 showed very high ANI with two E. coli strains from KBN10P04869 (Korea, GenBank accession number: CP026473.1) and RUT3575 (Spain, GenBank accession number: CP048010.1) and H10407 is an enterotoxigenic E. coli isolate (GenBank accession number: NC_017633.1), while the complete genome of LZ00114 and WCHEC025943 had 99.97% of ANI (Figure 2A). The phylogenetic tree showed that LZ00114 was evolutionarily closer to WCHEC025943 and AMA1167, as shown in Figure 2B.
 |
Table 2 General Characteristics of the LZ00114 Genomes |
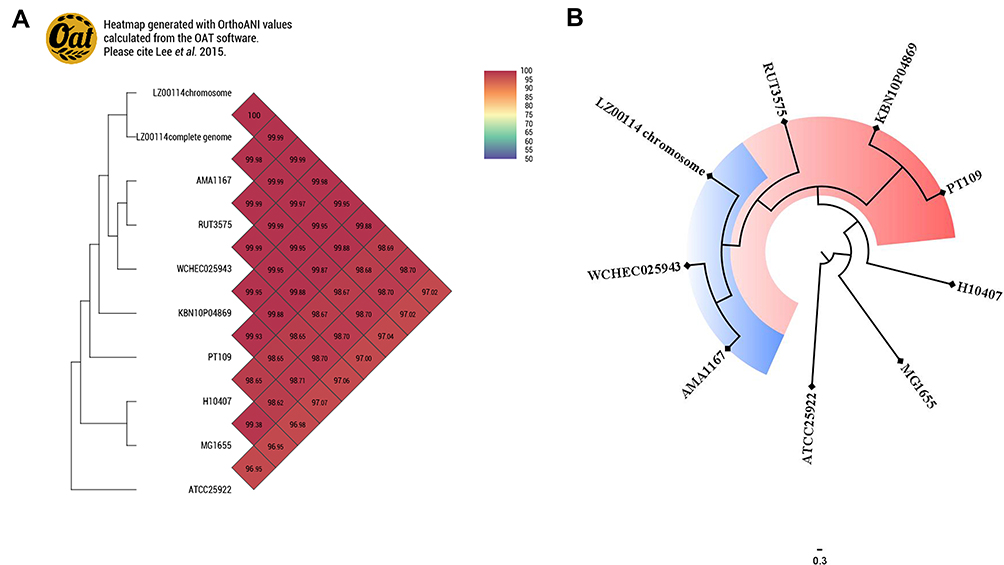 |
Figure 2 (A) Orthologous average nucleotide identity values of LZ00114, E. coli str. K-12 substr. MG1655 and other E. coli strains. E. coli ATCC25922 (GenBank accession number: CP009072.1). (B) The SNPs-based phylogenetic tree was constructed using LZ00114 and 6 E. coli genomes and visualized by FigTree v1.4.4 software. E. coli str.k-12 substr. MG1655 served as the reference genome. |
Chromosomal Genome Analysis of LZ00114
We selected the E. coli K12 MG1655 as the reference strain to identify the chromosomal point mutations. There were amino acid substitutions in gyrA (S83L, D87N), parE (S458A), and parC (S80I) within the fluoroquinolone resistance-determining region of LZ00114. As the mcr genes were not identified in the genome, we speculated that chromosomal mutations mediate polymyxin B resistance. By Sanger sequencing, we observed that the PCR products of two-component regulatory genes, namely, pmrAB and phoPQ had point mutations as follows: pmrA (S29G, G144S) and pmrB (D283G, Y358N). Whole genome sequencing subsequently confirmed these results, and amino acid substitutions were found in the eptA (pmrC) gene (D348G, T413S). Furthermore, the nfsA gene, which encodes NADPH-dependent nitroreductase also had a point mutation (R203H). We then identified point mutations in marR (G103S, Y137H), acrD (Q656K, A740D), ermA (I219V), emrD (G323D). Subsequently, we verified the efflux pump phenotype and confirmed the presence of efflux pump overexpression by RT-PCR.
In addition, we identified a genomic island (GI) containing blaCMY-2 with a length of 15,343 bp on the chromosome backbone of LZ00114 (4,213,994–4,229,337bp). The GC ratio of this GI was 48.87% lower than that of the other chromosomal regions. This region may have been integrated into the chromosome by mobile elements, and further analysis suggested that integration may have been accomplished by IS600 and IS66-like elements at either end of the GI (Figure S2). We compared the genetic environment and found that blaCMY-2 integrated into the chromosome lacking blc and yggR in the surrounding environment compared with plasmid-derived blaCMY-2 (Figure S1). The blaCMY-2 genetic environment of LZ00114 was similar to the E. coli strain RUT3575 (GenBank accession number: CP048010.1), with the same blaCMY-2 located on the chromosome (Figure S1).
Characteristics of an IncX3 blaNDM-5-Harboring Plasmid p25NDM-5 and an IncFII/FIA/FIB blaOXA-1-Harboring Plasmid p1OXA77
blaNDM-5 was located on a 46,160 bp IncX3 plasmid p25NDM-5 with a GC ratio of 46.7%. By BLAST-based sequence alignment using the GenBank nucleic acid sequence database, we found that two plasmids carrying blaNDM-5 isolated from E. coli from Shandong, China (pVH1, accession number: CP028705.1) and Korea (pCREC-591_4, GenBank accession number: CP024825.1) were highly similar to p25NDM-5. The identities of p25NDM-5 with pVH1 (query coverage: 100%), pCREC-591_4 (query coverage: 100%) and p744T-NDM5 (query coverage: 98%) were ≥ 99.9%. p744T-NDM5 (GenBank accession number: MF547511.1) was an IncX3 plasmid isolated from food animal-derived E. coli. The comparative genome circle revealed that p25NDM5 was almost identical to the plasmid structures of pCREC-591_4 and pVH1, and ISAba125 of p744T-NDM5 was truncated (Figure 3). Further analysis showed that the surrounding genetic environment of blaNDM-5 of p25NDM-5 consisted of a highly conserved region (blaNDM-5-bleMBL(bleomycin binding protein)-trpF (phosphoribosylanthranilate isomerase)-tat (signal sequence domain protein), and dct (divalent-cation tolerance protein), umuD (mutagenesis and repair protein), IS26, IS5, ISAba125 and IS3000 (Figure 3).
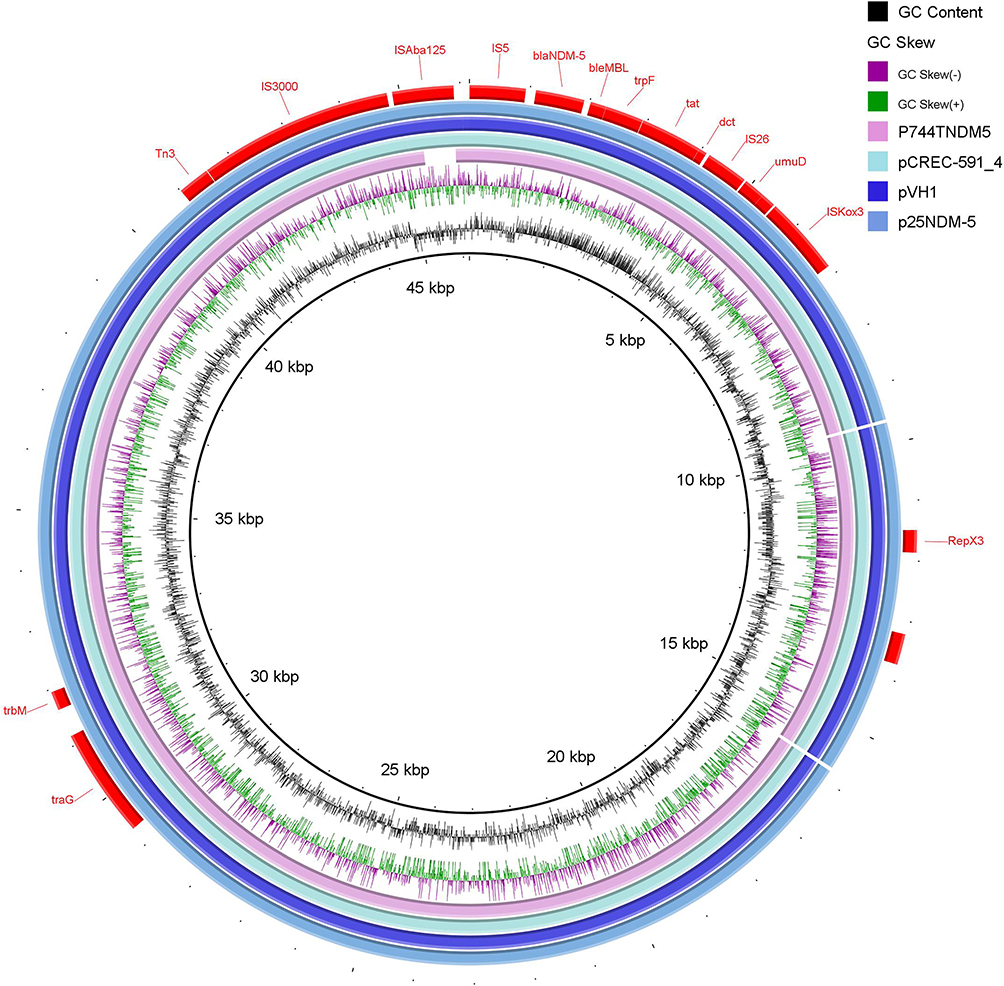 |
Figure 3 Circular genomic comparison of p25NDM-5 with three other plasmids drawn by BRIG software. Different colors represent plasmids from different isolates. Red bars indicate genetic annotations, and blank represents the missing region compared with the reference genome. |
The GC ratio of p1OXA77 was 51.4%, and it contained many type IV secretion system (T4SS) encoding genes and plasmid addiction systems (pemKI, ccdAB, and vapBC). We did not identify plasmid sequences with high similarity to p1OXA77 in the NCBI database. Furthermore, p1OXA77 carried blaOXA-1, blaCTX-M-55, an incomplete blaTEM, tetracycline, aminoglycoside, trimethoprim, sulfonamide, fosfomycin and chloramphenicol resistance genes (tet(B)-aac(6’)-Ib-cr-dfrA17-sul1-fosA3-ΔcatB). The replicon typing of p1OXA77 belonged to IncFII/FIA/FIB, which contained the multiple replication regulatory protein RepA. We selected five plasmid sequences isolated from E. coli in the NCBI database through BLAST for alignment as follows: p38_A-OXA140 (GenBank accession number: CP048377.1), p92944-mph (GenBank accession number: MG838205.1), blaNDM-5-carrying plasmid unnamed 3 (GenBank accession number: CP083875.1), pBJ114-141 (GenBank accession number: MF679146.1), and pEC71-IncHI2 (GenBank accession number: CP085623.1). Genome alignment revealed that p1OXA77 was similar to p92944-mph (query coverage: 61%; identity: 99.95%) and pBJ114-141 (query coverage: 60%; identity: 99.89%), but with many regional deletions (Figure 4). In addition, there are many insertion sequences in p1OXA77, especially IS26.
 |
Figure 4 Circular genomic comparison of p1OXA77 with five other plasmids drawn by BRIG software. Different colors represent plasmids from different isolates. Black bars indicate genetic annotations, and blank represents the missing region compared with the reference genome. |
Furthermore, we confirmed that p25NDM-5 could be transferred horizontally into the E. coli J53 strain by conjugation experiments, which significantly reduced the susceptibility of E. coli J53 to cephalosporins and meropenem (Table S2). However, we performed repeated conjugation experiments and did not observe the horizontal transfer of p1OXA77 to J53.
Efflux Pump Phenotype Test
To evaluate the role of efflux pumps in antimicrobial resistance, we used efflux pump inhibitors to determine the efflux pump phenotype. The MICs of chloramphenicol, tetracycline, meropenem, and ciprofloxacin decreased 4-fold, and the MIC of gentamicin decreased 2-fold after adding CCCP at 10 µg/mL. In marked contrast to the addition of NMP, CCCP restored the MIC of polymyxin B to 0.5 µg/mL and reversed polymyxin B resistance (Table 1).
Quantitative Reverse Transcription-PCR
The expression levels of PmrAB, RND, and MFS efflux pumps of LZ00114 were detected by RT-PCR. Compared with E. coli ATCC25922, acrA (8.39±0.38) and acrB (2.99 ± 0.27) genes of LZ00114 were upregulated approximately 8-fold and 3-fold, respectively, which may be related to the marR gene mutation (G103S, T137S). The transcript levels of acrD (2.18 ± 0.15), emrA (2.04 ± 0.32), emrD (1.90 ± 0.14), and emrY (17.51 ±0.23) genes were generally upregulated. For colistin resistance, the expression of pmrA (16.37 ± 1.12), pmrB (1.67 ± 0.12), and eptA (2.45 ± 0.13) genes in a colistin-resistant mutant (LZ00114) were significantly upregulated compared with the Lab-WT strain (Figure 5). These results indicated that the overexpression of these genes may have a synergistic role with the acquired antimicrobial resistance genes, thereby promoting high levels of antimicrobial resistance of LZ00114.
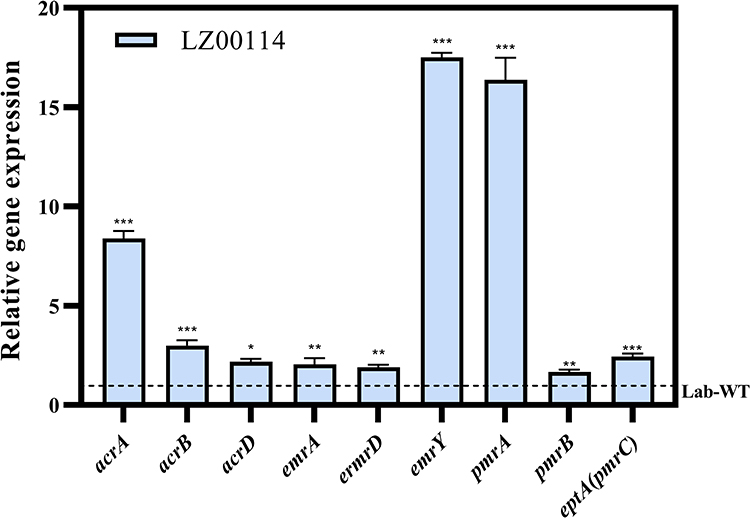 |
Figure 5 The gene transcriptional levels of pmrA, pmrB, eptA(pmrC), acrA, acrB, acrD, emrA, emrD and emrY were drawn by GraphPad Prism 9.2.0. *P < 0.05, **P < 0.01, ***P < 0.001 by Student’s t-test. The error bar represents the standard deviation of the mean. |
Discussion
In 2016, a study provided preliminary evidence for clonal and interspecies transmission of the newly emerged high-risk clone of E. coli ST410.27 A subsequent study in 2018 demonstrated the clonal spread of E. coli ST410 in countries such as the United Kingdom, Japan, Denmark, Brazil, the United States, and Singapore, which can also carry carbapenem and colistin resistance determinants.7 The average nucleotide identify is one of the reliable indexes to analyze the genetic relationship among strains. We reconfirmed the clone transmission of ST410 by comparing the ANI values of LZ00114 and E. coli ST410 strains isolated from different regions. ANI range > 95–96% is generally defined as the species circumscription, and the higher the value, the closer the genetic relationship between strains.28,29 ST410 is a common ExPEC lineage belonging to the ST23 complex, with only one allele difference compared with ST23. Fitness is usually an important indicator to assess whether these resistant microorganisms can stably spread in the environment, whereas there is little information on the fitness of the resistant E. coli ST410 clonal group. It is generally believed that the acquisition of antimicrobial resistance could be an additional metabolic stress for bacteria in the absence of antibiotics, the so-called fitness cost.30 Considering that horizontal plasmid transfer may occur when LZ00114 is co-cultured with non-resistant antibiotic strains, we did not conduct competitive growth experiments to evaluate the fitness costs of LZ00114 in the current study. However, growth kinetics and mobility showed that LZ00114 may have more substantial survival and competitiveness than WT susceptible strains even in an environment without antibiotics (Figure 1). It is still necessary to further study the relationship between this microbial characteristic and sequence type ST410. E. coli ST410 is widely spread in natural and clinical settings and may have a more complex genetic background.27 In some specific environments, it helps to select lower-cost antimicrobial resistance.31 For multi-drug resistant strains, the epistatic effects of genetic elements and compensatory mutations cannot be ignored in alleviating the resistance costs.32 Although the serotype of LZ00114 is O8:H9 (identity>99.5%), we did not find any virulence plasmids in LZ00114 (Table 2). Moreover, the motility, biofilm formation ability, and virulence factors of LZ00114 are consistent with the general characteristics that UPEC can colonize the infection site by expressing fimbriae and biofilms, and siderophores can contribute to the survival of UPEC in the urinary tract environment.33,34
Carbapenem-resistant Enterobacterales can develop into extensively drug-resistant (XDR), and pandrug-resistant (PDR) strains by mobile elements and chromosomal mutations. In E. coli, the high plasticity of the genome is the main reason for driving the development of antimicrobial resistance.35 In the present study, the ANI of the LZ00114 genome, the reference strain E. coli ATCC25992, and MG1655 also suggested that LZ00114 carried many exogenous genes. WGS revealed that p25NDM-5 and p1OXA77 of LZ00114 had genes encoding β-lactamases. Of these, blaOXA belonged to class D β-lactamases and was initially identified in nonfermenters. The horizontal transfer of blaOXA to Enterobacterales via mobile elements has become an emerging threat. Although we did not observe the horizontal transfer of p1OXA77 to J53 in the present study, we cannot conclude that p1OXA77 lacks the ability of horizontal transfer. p1OXA77 carried the multiple replication regulatory protein RepA (Figure 4). We speculated that the fitness cost of the plasmid restricts the transmission of the p1OXA77.36 However, the successful transfer of p25NDM-5 reduced the susceptibility of the recipient strains to β-lactamase antibiotics, which still indicated the potential risk of carbapenem resistance transmission. IncX3 is the primary incompatibility group spreading blaNDM-5 in China, and it carries blaNDM-5 isolated from different regions, which also display similar sizes.37,38 We confirmed the correlation of p25NDM-5 with previously isolated blaNDM-5-harboring plasmids by sequence alignment and re-emphasized the clonal spread of such plasmids (Figure 3). And the chromosome genome of LZ00114 is also highly similar to that of an E. coli isolate WCHEC025943 isolated from sewage, suggesting the transfer of pathogens may occur from the natural environment. In vitro environments, especially those containing sublethal levels of antibiotics, show a strong facilitative effect on the selection and spread of antimicrobial resistance in these pathogens.39 We speculated that environmental factors promote the integration of a series of mobile elements in LZ00114. Meanwhile, a case of E. coli ST410 with a resistance pattern similar to that of LZ00114 was previously reported in Hangzhou, China.40 Environmental factors and population migration are often crucial in the spread of antimicrobial resistance across regions.41 Thus, it is necessary to monitor these pathogens to control the further spread of antimicrobial resistance.
In addition to acquiring numerous antimicrobial resistance genes via mobile elements, there were many chromosomal point mutations in LZ00114. Amino acid substitution of LZ00114 in gyrA, parE, and parC genes has previously been associated with quinolones.42 Previous studies revealed that R203C and R203L substitutions in the nfsA gene are associated with nitrofurantoin resistance in E. coli.43,44 The contribution of the R203H substitution in the nfsA gene in LZ00114 to nitrofurantoin resistance remains to be further clarified. The marR gene is an important transcription factor that regulates multi-drug resistance and stress response. The acrAB genes of LZ00114 were significantly upregulated without sense mutations, which again emphasized the regulatory effects of the marR gene on the AcrAB efflux pump.45 The overexpression of emrA and emrY has been related to β-lactams, tetracycline, and quinolone resistance. In particular, the EmrD efflux pump can regulate the transport of common antibiotics and increase the resistance of bacteria to CCCP,46 while a previous study indicated that emrD mutations could increase the susceptibility of bacterial cells to CCCP.47 In the present study, we found that CCCP (10 µg/mL) could inhibit the efflux pump activity of LZ00114 in the presence of emrD overexpression. We speculated that it may have been related to a point mutation in emrD (G323D), but it is not clear to what extent this point mutation can affect the inhibitory effects of CCCP on the efflux pump. Mutations in the sensor histidine kinase pmrB are an essential mechanism that mediates polymyxins resistance in E. coli, especially in uropathogenic strains,48,49 while amino acid substitutions at S29G and G144S in pmrA, D283G in pmrB, and D348G, T413S in eptA are also present in colistin-susceptible strains as described in the previous study. By contrast, the Y358N substitution mutation in the pmrB gene has been described mainly in colistin-resistant strains.50 Therefore, it is necessary to evaluate the values of these chromosomal point mutations in identifying colistin-resistant strains. In fact, the patient was not treated with colistin during hospitalization, which also suggests that there may be other factors causing colistin resistance mutations. Notably, CCCP completely reversed the resistance of LZ00114 to polymyxin B with an 8-fold decrease in the MIC. Previous studies have reported that CCCP can reverse the resistance to colistin in colistin-resistant Enterobacterales.17,51 Colistin is a cationic polypeptide. The resistance mechanism of bacteria to colistin is usually due to the modification of lipopolysaccharides, the main component of the bacterial outer membrane. These modifications reduced the outer membrane’s negative charge, resulting in weaker binding to colistin.52 CCCP is a proton carrier efflux pump inhibitor, which mainly acts by reducing the transmembrane electrochemical gradient. Ni et al53 suggested that CCCP could restore the negative charges on the outer membrane by disrupting the proton gradient. However, it is still unclear whether efflux pumps are involved in colistin resistance or CCCP directly plays the role in reversing polymyxins resistance, and the exact mechanism needs further study.
There are some limitations in this study. First, we focused on single colistin-resistant E. coli isolate in the present study, while the prevalence of colistin-resistant Enterobacterales in Luzhou, China is still unclear. In the future, we will screen colistin-resistant strains isolated from the same hospital and comprehensively analyze the prevalence of colistin-resistant strains in this area combined with clinical data. Second, we identified several chromosomal point mutations in LZ00114, of which the contribution of the emrD mutation (G323D) to antimicrobial and CCCP resistance in LZ00114 remains to be further determined. Third, although we confirmed that CCCP achieves reversal of colistin resistance in E. coli, further studies are lacking to analyze the exact reversal mechanisms.
Conclusion
In conclusion, we reported for the first time to the best of our knowledge the complete characterization of a colistin-resistant CREC (carbapenem-resistant E. coli) ST410 isolate LZ00114 and an IncX3 plasmid carrying blaNDM-5 in Southern Sichuan. The resistance phenotypes of LZ00114 were closely related to previous isolates from other regions, which were formed under the synergism of plasmids carrying acquired antimicrobial resistance genes and chromosomal point mutations. The spread of the E. coli ST410 clone group and antimicrobial resistance across regions requires close monitoring. Our research found that CCCP reversed the polymyxin B resistance of LZ00114, which provides a basis for further research on the mechanism of colistin resistance and the development of new drugs adjuvants.
Data Sharing Statement
The LZ00114 genome used in the present study has been submitted to the NCBI database under the GenBank accession numbers: chromosome of LZ00114 (GenBank accession number: CP087570), p1OXA77 (GenBank accession number: CP087571), and p25NDM-5 (GenBank accession number: CP087572).
Ethics Statement
The study protocol was approved by the Institutional Review Board of the Affiliated Hospital of Southwest Medical University (Project No. KY2020043). All participants provided written informed consent to participate in this study.
Author Contributions
All authors have made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas, have drafted or written, or substantially revised or critically reviewed the article, have agreed on the journal to which the article will be submitted, reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage, and agree to take responsibility and be accountable for the contents of the article.
Funding
This research was supported by the grants from the Health Commission of Sichuan Province program (19PJ293), Sichuan Science and Technology Department Program (20QYCX0056, 2021YFH001), and Sichuan Education Department Training Program of Innovation and Entrepreneurship for College Students (S202010632250).
Disclosure
The author reports no conflicts of interest in this work.
References
1. Jacobsen SM, Stickler DJ, Mobley HLT, Shirtliff ME. Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin Microbiol Rev. 2008;21(1):26. doi:10.1128/cmr.00019-07
2. Croxen MA, Finlay BB. Molecular mechanisms of Escherichia coli pathogenicity. Nat Rev Microbiol. 2010;8(1):26–38. doi:10.1038/nrmicro2265
3. Toval F, Koehler C-D, Vogel U, et al. Characterization of Escherichia coli isolates from hospital inpatients or outpatients with urinary tract infection. J Clin Microbiol. 2014;52(2):407–418. doi:10.1128/jcm.02069-13
4. Temkin E, Adler A, Lerner A, Carmeli Y. Carbapenem-resistant Enterobacteriaceae: biology, epidemiology, and management. Ann N Y Acad Sci. 2014;1323(1):22–42. doi:10.1111/nyas.12537
5. Petty NK, Ben Zakour NL, Stanton-Cook M, et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc Natl Acad Sci USA. 2014;111(15):5694–5699. doi:10.1073/pnas.1322678111
6. Garcia-Fernandez A, Villa L, Bibbolino G, et al. Novel insights and features of the NDM-5-producing Escherichia coli sequence Type 167 high-risk clone. mSphere. 2020;5(2). doi:10.1128/mSphere.00269-20
7. Roer L, Overballe-Petersen S, Hansen F, et al. Escherichia coli sequence type 410 is causing new international high-risk clones. mSphere. 2018;3(4). doi:10.1128/mSphere.00337-18
8. Liu Z, Li J, Wang X, et al. Novel variant of New Delhi metallo-beta-lactamase, NDM-20, in Escherichia coli. Front Microbiol. 2018;9. doi:10.3389/fmicb.2018.00248
9. Bottery MJ, Wood AJ, Brockhurst MA. Adaptive modulation of antibiotic resistance through intragenomic coevolution. Nat Ecol Evol. 2017;1(9):1364–1369. doi:10.1038/s41559-017-0242-3
10. Jeannot K, Bolard A, Plésiat P. Resistance to polymyxins in Gram-negative organisms. Int J Antimicrob Agents. 2017;49(5):526–535. doi:10.1016/j.ijantimicag.2016.11.029
11. Telke AA, Olaitan AO, Morand S, Rolain JM. soxRS induces colistin hetero-resistance in Enterobacter asburiae and Enterobacter cloacae by regulating the acrAB-tolC efflux pump. J Antimicrob Chemother. 2017;72(10):2715–2721. doi:10.1093/jac/dkx215
12. Liao W, Lin J, Jia H, et al. Resistance and heteroresistance to colistin in Escherichia coli isolates from Wenzhou, China. Infect Drug Resist. 2020;13:3551–3561. doi:10.2147/idr.S273784
13. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi:10.1111/j.1469-0691.2011.03570.x
14. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing.
15. Green MR, Sambrook J. Isolating DNA from gram-negative bacteria. Cold Spring Harb Protoc. 2017;2017(1). doi:10.1101/pdb.prot093369
16. Schumacher A, Steinke P, Bohnert JA, Akova M, Jonas D, Kern WV. Effect of 1-(1-naphthylmethyl)-piperazine, a novel putative efflux pump inhibitor, on antimicrobial drug susceptibility in clinical isolates of Enterobacteriaceae other than Escherichia coli. J Antimicrob Chemother. 2006;57(2):344–348. doi:10.1093/jac/dki446
17. Baron SA, Rolain JM. Efflux pump inhibitor CCCP to rescue colistin susceptibility in mcr-1 plasmid-mediated colistin-resistant strains and Gram-negative bacteria. J Antimicrob Chemother. 2018;73(7):1862–1871. doi:10.1093/jac/dky134
18. Kim M, Park J, Park W. Genomic and phenotypic analyses of multidrug-resistant Acinetobacter baumannii NCCP 16007 isolated from a patient with a urinary tract infection. Virulence. 2021;12(1):150–164. doi:10.1080/21505594.2020.1867421
19. Zhao X, Lv Y, Adam FEA, et al. Comparison of antimicrobial resistance, virulence genes, phylogroups, and biofilm formation of Escherichia coli isolated from intensive farming and free-range sheep. Front Microbiol. 2021;12:699927. doi:10.3389/fmicb.2021.699927
20. Ma T, Fu J, Xie N, et al. Fitness cost of bla(NDM-5)-carrying p3R-IncX3 plasmids in wild-type NDM-free Enterobacteriaceae. Microorganisms. 2020;8(3):377. doi:10.3390/microorganisms8030377
21. Lane MC, Simms AN, Mobley HLT. Complex interplay between type 1 fimbrial expression and flagellum-mediated motility of uropathogenic Escherichia coli. J Bacteriol. 2007;189(15):5523–5533. doi:10.1128/jb.00434-07
22. Lee I, Ouk Kim Y, Park SC, Chun J. OrthoANI: an improved algorithm and software for calculating average nucleotide identity. Int J Syst Evol Microbiol. 2016;66(2):1100–1103. doi:10.1099/ijsem.0.000760
23. Long HY, Feng Y, Ma K, Liu L, McNally A, Zong ZY. The co-transfer of plasmid-borne colistin-resistant genes mcr-1 and mcr-3.5, the carbapenemase gene bla(NDM-5) and the 16S methylase gene rmtB from Escherichia coli. Sci Rep. 2019;9(1):6. doi:10.1038/s41598-018-37125-1
24. Brilhante M, Menezes J, Belas A, et al. OXA-181-producing extraintestinal pathogenic Escherichia coli sequence type 410 isolated from a dog in Portugal. Antimicrob Agents Chemother. 2020;64(4):6. doi:10.1128/aac.02298-19
25. Overballe-Petersen S, Roer L, Ng K, et al. Complete nucleotide sequence of an Escherichia coli sequence Type 410 strain carrying bla(NDM-5) on an IncF multidrug resistance plasmid and bla(OXA-181) on an IncX3 plasmid. Genome Announc. 2018;6(5). doi:10.1128/genomeA.01542-17
26. Hsiao W, Wan I, Jones SJ, Brinkman FS. IslandPath: aiding detection of genomic islands in prokaryotes. Bioinformatics. 2003;19(3):418–420. doi:10.1093/bioinformatics/btg004
27. Schaufler K, Semmler T, Wieler LH, et al. Clonal spread and interspecies transmission of clinically relevant ESBL-producing Escherichia coli of ST410-another successful pandemic clone? FEMS Microbiol Ecol. 2016;92(1):9. doi:10.1093/femsec/fiv155
28. Richter M, Rossello-Mora R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A. 2009;106(45):19126–19131. doi:10.1073/pnas.0906412106
29. Jain C, Rodriguez-R LM, Phillippy AM, Konstantinidis KT, Aluru S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat Commun. 2018;9(1):8. doi:10.1038/s41467-018-07641-9
30. Martinez JL, Rojo F. Metabolic regulation of antibiotic resistance. FEMS Microbiol Rev. 2011;35(5):768–789. doi:10.1111/j.1574-6976.2011.00282.x
31. Gullberg E, Cao S, Berg OG, et al. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011;7(7):e1002158. doi:10.1371/journal.ppat.1002158
32. Durão P, Balbontín R, Gordo I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018;26(8):677–691. doi:10.1016/j.tim.2018.01.005
33. Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2(2):123–140. doi:10.1038/nrmicro818
34. Soto SM, Smithson A, Horcajada JP, Martinez JA, Mensa JP, Vila J. Implication of biofilm formation in the persistence of urinary tract infection caused by uropathogenic Escherichia coli. Clin Microbiol Infect. 2006;12(10):1034–1036. doi:10.1111/j.1469-0691.2006.01543.x
35. Horesh G, Blackwell GA, Tonkin-Hill G, Corander J, Heinz E, Thomson NR. A comprehensive and high-quality collection of Escherichia coli genomes and their genes. Microb Genom. 2021;7(2). doi:10.1099/mgen.0.000499
36. San Millan A, MacLean RC. Fitness costs of plasmids: a limit to plasmid transmission. Microbiol Spectr. 2017;5(5). doi:10.1128/microbiolspec.MTBP-0016-2017
37. Ho PL, Wang Y, Liu MCJ, et al. IncX3 epidemic plasmid carrying bla(NDM-5) in Escherichia coli from Swine in multiple geographic areas in China. Antimicrob Agents Chemother. 2018;62(3):5. doi:10.1128/aac.02295-17
38. Li X, Fu Y, Shen M, et al. Dissemination of bla(NDM-5) gene via an IncX3-type plasmid among non-clonal Escherichia coli in China. Antimicrob Resist Infect Control. 2018;7(1):59. doi:10.1186/s13756-018-0349-6
39. Andersson DI, Hughes D. Microbiological effects of sublethal levels of antibiotics. Nat Rev Microbiol. 2014;12(7):465–478. doi:10.1038/nrmicro3270
40. Huang J, Ma S, Yu Q, et al. Whole genome sequence of an Escherichia coli ST410 isolate co-harbouring bla(NDM-5), bla(OXA-1), bla(CTX-M-15), bla(CMY-2), aac(3)-IIa and aac(6’)-Ib-cr genes isolated from a patient with bloodstream infection in China. J Glob Antimicrob Resist. 2019;19:354–355. doi:10.1016/j.jgar.2019.10.027
41. Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M beta-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–2155. doi:10.1093/jac/dkx146
42. Moon DC, Seol SY, Gurung M, et al. Emergence of a new mutation and its accumulation in the topoisomerase IV gene confers high levels of resistance to fluoroquinolones in Escherichia coli isolates. Int J Antimicrob Agents. 2010;35(1):76–79. doi:10.1016/j.ijantimicag.2009.08.003
43. Sandegren L, Lindqvist A, Kahlmeter G, Andersson DI. Nitrofurantoin resistance mechanism and fitness cost in Escherichia coli. J Antimicrob Chemother. 2008;62(3):495–503. doi:10.1093/jac/dkn222
44. Sorlozano-Puerto A, Lopez-Machado I, Albertuz-Crespo M, Martinez-Gonzalez LJ, Gutierrez-Fernandez J. Characterization of fosfomycin and nitrofurantoin resistance mechanisms in Escherichia coli isolated in clinical urine samples. Antibiotics. 2020;9(9). doi:10.3390/antibiotics9090534
45. Perera IC, Grove A. Molecular mechanisms of ligand-mediated attenuation of DNA binding by MarR family transcriptional regulators. J Mol Cell Biol. 2010;2(5):243–254. doi:10.1093/jmcb/mjq021
46. Yu L, Li W, Xue M, et al. Regulatory role of the two-component system bassr in the expression of the EmrD multidrug efflux in Escherichia coli. Microb Drug Resist. 2020;26(10):1163–1173. doi:10.1089/mdr.2019.0412
47. Naroditskaya V, Schlosser MJ, Fang NY, Lewis K. An E. coli gene emrD is involved in adaptation to low energy shock. Biochem Biophys Res Commun. 1993;196(2):803–809. doi:10.1006/bbrc.1993.2320
48. Phan MD, Nhu NTK, Achard MES, et al. Modifications in the pmrB gene are the primary mechanism for the development of chromosomally encoded resistance to polymyxins in uropathogenic Escherichia coli. J Antimicrob Chemother. 2017;72(10):2729–2736. doi:10.1093/jac/dkx204
49. Janssen AB, van Schaik W. Harder, better, faster, stronger: colistin resistance mechanisms in Escherichia coli. PLoS Genet. 2021;17(1):4. doi:10.1371/journal.pgen.1009262
50. Luo QX, Yu W, Zhou K, et al. Molecular epidemiology and colistin resistant mechanism of mcr-positive and mcr-negative clinical isolated Escherichia coli. Front Microbiol. 2017;8:13. doi:10.3389/fmicb.2017.02262
51. Osei Sekyere J, Amoako DG. Carbonyl Cyanide m-Chlorophenylhydrazine (CCCP) reverses resistance to colistin, but not to carbapenems and tigecycline in multidrug-resistant Enterobacteriaceae. Front Microbiol. 2017;8:228. doi:10.3389/fmicb.2017.00228
52. Trimble MJ, Mlynarcik P, Kolar M, Hancock REW. Polymyxin: alternative mechanisms of action and resistance. Cold Spring Harb Perspect Med. 2016;6(10):22. doi:10.1101/cshperspect.a025288
53. Ni W, Li Y, Guan J, et al. Effects of efflux pump inhibitors on colistin resistance in multidrug-resistant gram-negative bacteria. Antimicrob Agents Chemother. 2016;60(5):3215–3218. doi:10.1128/aac.00248-16
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.