")
Back to Journals » OncoTargets and Therapy » Volume 13
Genetic Variability in the microRNA Binding Sites of BMPR1B, TGFBR1, IQGAP1, KRAS, SETD8 and RYR3 and Risk of Breast Cancer in Colombian Women
Authors Garcia Mesa K, Bermejo JL , Torres D , Gilbert M, Plass C, Hamann U
Received 28 July 2020
Accepted for publication 6 October 2020
Published 30 November 2020 Volume 2020:13 Pages 12281—12287
DOI https://doi.org/10.2147/OTT.S274431
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Takuya Aoki
Karen Garcia Mesa,1,2 Justo Lorenzo Bermejo,1,2 Diana Torres,1,3 Michael Gilbert,1 Christoph Plass,4 Ute Hamann1
1Molecular Genetics of Breast Cancer, German Cancer Research Center (DKFZ), Heidelberg, Germany; 2Institute of Medical Biometry and Informatics, University of Heidelberg, Heidelberg, Germany; 3Institute of Human Genetics, Pontificia Universidad Javeriana, Bogota, Colombia; 4Cancer Epigenomics, German Cancer Research Center (DKFZ), Heidelberg, Germany
Correspondence: Ute Hamann
Molecular Genetics of Breast Cancer, German Cancer Research Center (DKFZ), Im Neuenheimer Feld 580, Heidelberg 69120, Germany
Tel +49 6221 422347
Fax +49 6221 424721
Email [email protected]
Background: Genetic variants in microRNA (miR) binding sites affect the regulation of miR-dependent gene expression and have been linked to the risk of a variety of cancers including breast cancer (BC). Most BC risk variants had been identified in women of European and Asian ancestry, but genetic data for Hispanic women are scarce. Here, we investigate the association between six variants in miR binding sites and BC risk in Colombian women.
Methods: We genotyped miR binding site variants in the BMPR1B, TGFBR1, IQGAP1, KRAS, SETD8 and RYR3 genes in 1022 BC cases and 1023 controls from the Colombian breast cancer case–control (Col-BCCC) study using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Multiple logistic regression and permutation techniques were applied to assess the association between genetic variants and BC risk.
Results: We found no evidence of association between any of the six miR binding site variants and overall or estrogen receptor subtype-specific BC risk in Colombian women.
Conclusion: Our findings may point to ethnic differences in the association between genetic variability in miR binding sites and breast cancer risk.
Keywords: breast cancer, Hispanics, risk factors, miR binding site SNPs, Colombia
Introduction
Breast cancer (BC) is a major health burden in Colombia. It is the leading cancer and the second leading cause of cancer-related death in women with age-standardized incidence and mortality rates of 44.1 and 11.9 per 100,000 women, respectively.1 Identification and improved clinical management of patients at high risk of BC is important to reduce morbidity and mortality caused by this disease.2
The genetic component of BC is reflected on a tendency to cluster in families. Approximately 15–20% of all BCs are familial and 5–10% are hereditary.3 About 45% of the genetic component of BC risk is explained by rare mutations in known high (ie, BRCA1, BRCA2, PALB2) and moderate penetrance susceptibility genes (ie, ATM, CHEK2) and common variants in low-penetrance genes.4–7 The unexplained risk may be in part due to other common BC susceptibility variants. Attractive candidates are genetic variants in regulatory gene regions including microRNA (miR) binding sites.
miRs are a class of highly conserved short (21–23 nucleotides) noncoding RNAs, which play an important role in the regulation of gene expression at the post-transcriptional level. They bind to the complementary sequences of the 3ʹ untranslated region (3´ UTR) of target messenger RNAs, leading to repression of translation and consequently down-regulation of the expression of the proteins.8,9 Multiple miR binding site variants dysregulating important target genes involved in various biological pathways, such as DNA damage repair, signal transduction, and others as well variants in oncogenes and tumor suppressor genes have been identified as biomarkers for cancer susceptibility and prognosis.10–12
Most association studies of miR binding sites variants with risk of various cancer types including BC have been conducted in European and Asian populations.10 For BC, variants in genes involved in the TGF-ß signalling pathway (rs1434536 in bone morphogenetic protein receptor type-1B, BMPR1B and rs334348 in transforming growth factor beta receptor 1, TGFBR1) and the mitogen-activated protein kinase pathway (rs1042538 in IQ motif-containing GTPase-activating protein 1, IQGAP1 and rs61764370 in Kirsten rat sarcoma viral oncogene homolog, KRAS) were associated with disease risk, with the latter, however, yielding conflicting results.13–19 Further, associations were observed for variant rs16917496 in the SET domain containing protein 8 (SETD8) gene, which encodes a lysine methyltransferase that methylates various proteins, such as TP53, the androgen receptor and the estrogen receptor (ER) and variant rs1044129 in the calcium channel ryanodine receptor 3 (RYR3).20,21
Given the scarcity of this type of data for Hispanic women, we report for the first time on the frequency and association effects of six known European/Asian miR binding site risk variants in the BMPR1B, TGFBR1, IQGAP1, KRAS, SETD8 and RYR3 genes for female participants in the Colombian breast cancer case–control study Col-BCCC.
Materials and Methods
Study Population
The study population comprised 1022 female BC patients unselected for family history and age at BC diagnosis and 1023 female healthy controls who participated in the Colombian breast cancer case–control (Col-BCCC) study. Cases were mainly enrolled in the study at hospitals in Bogota, Neiva, and Villavicencio, which are located in the geographic center of Colombia, from 03/2007 to 02/2011. Cases were diagnosed with BC after January 1, 2004. Controls were healthy and unrelated women, who reported no family history of breast or any other type of cancer in two generations and who participated in the Colombian National Pap Smear Program.22 Controls were recruited from 06/2007 to 06/2011 and were matched to cases according to age (± 2 years). Cases and controls were only eligible if they were of Colombian origin and resided in the study region.23,24
Data on established and potential BC risk factors were collected from all study participants including age at diagnosis (cases)/age at interview (controls), proportion of European, Native American and African genetic ancestry, family history of BC in first-degree female relatives (yes, no), oral contraceptive (OC) use (yes, no), menopausal status and postmenopausal hormone therapy (HT) use (premenopausal and postmenopausal women who never used HT, postmenopausal women who ever used HT), body mass index (BMI) (<25, 25–29.9, ≥30 kg/m2), smoking status (never, former, current), parity (0, 1–2, ≥3), age at first full-term pregnancy (<20, 20–29, ≥30 years), age at menarche (≥14, 12–13, <12 years), and breastfeeding (never, ≤12, >12 months). Clinical and histopathological parameters were also collected from all cases (Table 1).
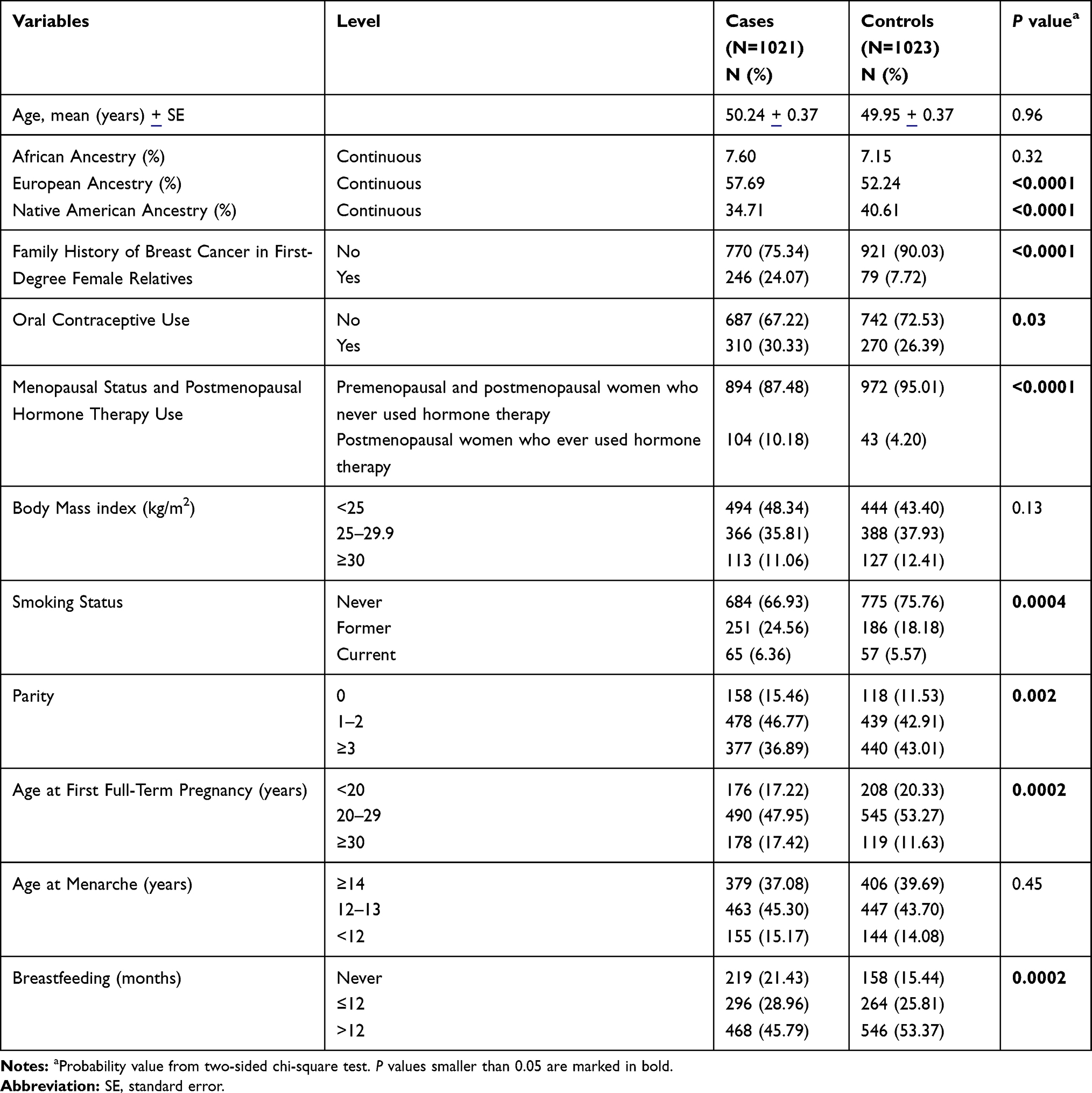 |
Table 1 Baseline Characteristics of the Study Population |
The study was approved by the Institutional Review Board of the Pontificia Universidad Javeriana (2006/75). All participants provided informed consent in writing previous to recruitment.
DNA Isolation and Genotyping Analyses
Genomic DNA was extracted from nine millilitre of peripheral blood collected into an EDTA tube using the salting out extraction method.25 We investigated six miR binding-site SNPs: rs1434536 (A>G in BMPR1B), rs334348 (A>G in TGFBR1), rs1042538 (A>T in IQGAP1), rs61764370 (T>G in KRAS), rs16917496 (T>C in SETD8), and rs1044129 (G>A in RYR3); the minor alleles in Colombians are underlined. The investigated SNPs were selected because i. they have a functional impact in modulating the expression of the target gene and ii. have been previously associated with BC risk in European or Asian populations. A description of the miR binding site SNPs under investigation is provided in Table 2.
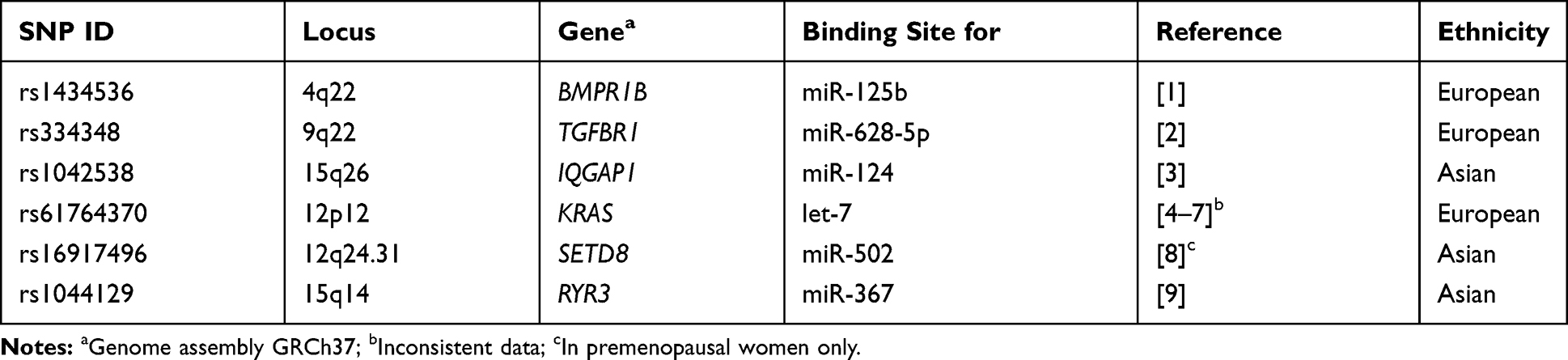 |
Table 2 Description of the SNPs Under Investigation |
Genotyping was performed in 1022 BC patients and 1023 healthy controls using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (Sequenom, San Diego, CA, USA) as previously described by us.24 All primers were synthesized by Sigma-Aldrich Chemie GmbH, Steinheim, Germany. Primer sequences are shown in Supplementary Table 1. Cluster plots and genotypes were inspected with the Typer 4.0.20 (TyperAnalyzer) software (Sequenom). The overall call rates for the six SNPs were ≥98% after removal of samples that consistently failed for >2 SNPs. The concordance rate of 135 duplicates (6.6%) was 99%, and the distribution of all genotypes met Hardy-Weinberg equilibrium (HWE) in the control group.
Thirty Ancestry Informative Markers (AIMs) were also genotyped to estimate the individual proportions of European, Native American, and African ancestry.26 Genotyping of 1757 native DNA samples and 288 Whole Genome Amplified (WGA) DNA samples was performed using the KASP allelic discrimination method (LGC Genomics, Teddington TW11 0LY, UK) as previously described.24 Cluster plots and genotypes were inspected with the SNPviewer2 software version 4.0.0 (http://www.lgcgroup.com/products/genotyping-software/snpviewer/#.-V1aqCEYppXg, LGC Genomics, Teddington TW11 0LY, UK). The genotype concordance between WGA amplified DNA and native DNA was >99%. AIM call rates were ≥98.9% after removal of samples that consistently failed for more than 20% of the SNPs. The concordance rate of 122 DNA duplicates (6.0%) was 100%. Eight AIMs deviated from HWE in controls but corresponding intensity plots revealed clear genotype clusters.
Statistical Analyses
The individual proportions of European, Native American, and African ancestry were estimated based on the 30 genotyped AIMs using the ADMIXTURE software as described previously.24 The association between the investigated genetic variants and BC risk was tested using logistic regression. An additive model was assumed for individual genotypes and the regression analyses were adjusted for the individual proportions of genetic ancestry, family history of BC in first-degree female relatives, OC use, menopausal status and postmenopausal use of HT, BMI, smoking status, parity, age at first full-term pregnancy, age at menarche, and breastfeeding (Table 1). Genotype-specific risks were quantified as per-allele odds ratios (ORs) with their corresponding 95% confidence intervals (CIs). Separate statistical analyses were conducted for any type of BC, ER-positive and ER-negative BC. Probability values were corrected for multiplicity using the permutation-based Westfall–Young method (10,000 permutations).27 Considering the average minor allele frequency of the six investigated variants (MAF=0.29), a Bonferroni-corrected significance level (α=0.0083) and an additive penetrance model, our study had 80% statistical power to detect an OR higher than 1.58. Statistical analyses were performed using the R software environment for statistical computing and SAS version 9.4. Phenotype and genotype data of all study participants are provided in Supplementary Table 2.
Results
Description of the Study Participants
The distribution of selected baseline characteristics between BC patients and controls is shown in Table 1. BC patients and controls showed similar age, BMI, age at menarche, and African ancestry proportions, while differences were noticed in the European and Native American ancestry proportions, family history of BC in first-degree female relatives, OC use, menopausal status and postmenopausal HT use, smoking status, parity, age at first full-term pregnancy, and breastfeeding.
Associations Between Recently Identified Common Risk Variants and Breast Cancer Risk in Colombian Women
No association between the six investigated miR binding site variants and overall BC risk was observed in Colombian women after correction for potential confounders and multiple testing (Table 3). In the stratified analyses by ER status of the diagnosed breast tumours, rs334348 (TGFBR1) and rs61764370 (KRAS) were associated with ER-positive BC risk in multiple logistic regression analyses (OR=1.24, 95% CI: 1.02–1.52; P=0.03 and OR=1.45, 95% CI: 1.02–2.07; P=0.04, respectively). The associations, however, did not reach statistical significance after correction for multiple testing.
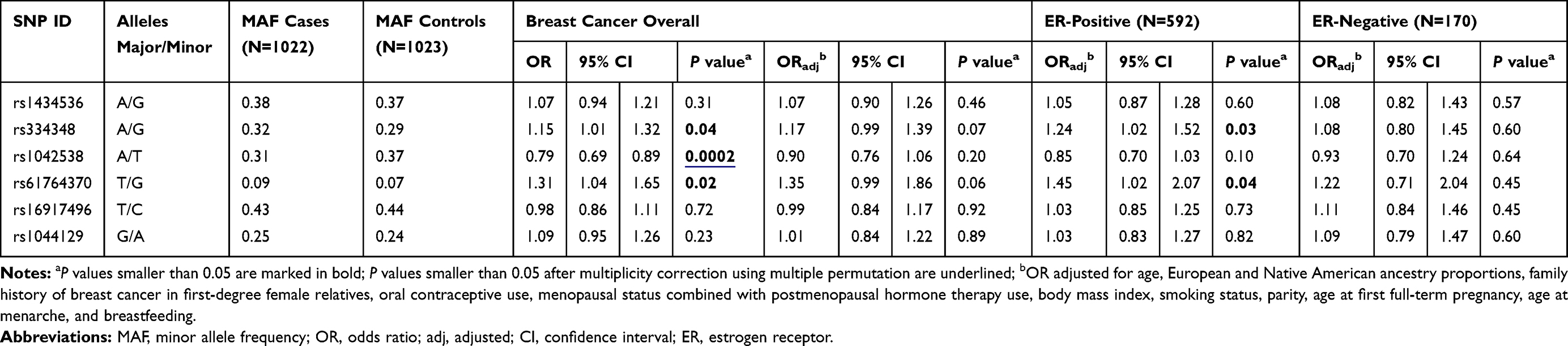 |
Table 3 Association Between miR Binding Site Variants and Breast Cancer Risk in Colombian Women |
Discussion
In this study, we investigated for the first time whether genetic variants located in miR binding sites at the 3´ UTR of the BMPR1B, TGFBR1, IQGAP1, KRAS, SETD8 and RYR3 genes, known to be associated with BC risk in women of European or Asian ancestry, affect BC risk in Colombian women. None of the miR-binding site variants was associated with the risk of overall or ER subtype-specific BC after adjustment for potential confounders and multiple testing.
Associations of two of the variants investigated in the present study, rs334348 (TGFBR1) and rs61764370 (KRAS), have been previously reported in women of European descent. One case–control study showed that women carrying the AG genotype of rs334348 (TGFBR1) had an increased risk of BC, and particularly familial BC.14 Several studies have investigated the association between rs61764370 (KRAS) and BC, which yielded inconsistent results.15–18
Our study has several strengths and limitations. Despite the reasonable sample size with more than 1000 BC cases and controls, the study was only able to detect OR >1.58 with 80% statistical power. A strength was the correction of BC risk estimates for established BC risk factors, genetic ancestry, and multiple testing.
Conclusion
Our data show no evidence of association between genetic variants in miR binding sites of the genes BMPR1B, TGFBR1, IQGAP1, KRAS, SETD8 and RYR3 and the risk of BC in Colombian women. Larger studies are needed to confirm the potential ethnic differences in the association between genetic variability in miR binding sites and breast cancer risk.
Abbreviations
AIMs, ancestry informative markers; BC, breast cancer; BMI, body mass index; BMPR1B, bone morphogenetic protein receptor type-1B; CI, confidence interval; Col-BCCC, Colombian breast cancer case–control study; ER, estrogen receptor; HT, postmenopausal hormone therapy; IQGAP1, IQ motif-containing GTPase-activating protein 1; KRAS, Kirsten rat sarcoma viral oncogene homolog; MAF, minor allele frequency; MALDI-TOF MS, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry; miR, microRNA; SETD8, SET domain containing (lysine methyltransferase) 8; SNP, single nucleotide polymorphism; TGFBR1, transforming growth factor beta receptor 1; RYR3, ryanodine receptor 3; OC, oral contraceptive; OR, odds ratio; ORadj, OR adjusted; SE, standard error; UTR, untranslated region; WGA, whole genome amplified.
Data Sharing Statement
Data beyond the information in this manuscript and supplementary tables cannot be publicly shared because they could be potentially used to re-identify study participants. Original data are available for request to the Institutional Review Board of the Pontificia Universidad Javeriana, Bogota, Colombia (contact via Melisa Diaz Puentes, [email protected], phone: +57 1 5946161 Ext 2470).
Ethics Approval and Informed Consent
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee (Institutional Review Board of the Pontificia Universidad Javeriana; ethics vote number 2006/75) and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Written informed consent was obtained from all study participants.
Acknowledgments
We thank the Colombian Breast Cancer Study Group (COLBCS) for its contribution. COLBCS: researchers: DT, I. Briceno (Pontificia Universidad Javeriana, Bogota); F. Gil (Unit of Clinical Epidemiology and Biostatistics, Pontificia Universidad Javeriana, Bogota); clinicians: J. Caicedo, C. Ramirez, E. Quintero, S. Quintero, J. Robledo (Country Clinic, Bogota); M. Tawil, L. Torregrosa (Pontificia Universidad Javeriana, Bogota); J. German Olaya (Hospital Universitario Hernando Moncaleano Perdomo, Neiva); study coordinator: UH, DKFZ, Heidelberg. We thank all women who participated in this study. We thank David Scherf, Rainer Claus, and Oliver Mücke for their support in MALDI-TOF genotyping analysis.
Author Contributions
UH conceived and designed the study. All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article is submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
We thank the DKFZ for support of the study.
Disclosure
All authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Weitzel JN, Blazer KR, MacDonald DJ, Culver JO, Offit K. Genetics, genomics, and cancer risk assessment: state of the art and future directions in the era of personalized medicine. CA Cancer J Clin. 2011;61(5):327–359.
3. Lynch HT, Lynch JF. Breast cancer genetics in an oncology clinic: 328 consecutive patients. Cancer Genet Cytogenet. 1986;22(4):369–371. doi:10.1016/0165-4608(86)90032-4
4. Ghoussaini M, Pharoah PD. Polygenic susceptibility to breast cancer: current state-of-the-art. Future Oncol. 2009;5(5):689–701. doi:10.2217/fon.09.29
5. Michailidou K, Lindstrom S, Dennis J, et al. Association analysis identifies 65 new breast cancer risk loci. Nature. 2017;551(7678):92–94.
6. Stratton MR, Rahman N. The emerging landscape of breast cancer susceptibility. Nat Genet. 2008;40(1):17–22. doi:10.1038/ng.2007.53
7. Wendt C, Margolin S. Identifying breast cancer susceptibility genes - a review of the genetic background in familial breast cancer. Acta Oncol. 2019;58(2):135–146. doi:10.1080/0284186X.2018.1529428
8. Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–355. doi:10.1038/nature02871
9. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
10. Ryan BM, Robles AI, Harris CC. Genetic variation in microRNA networks: the implications for cancer research. Nat Rev Cancer. 2010;10(6):389–402. doi:10.1038/nrc2867
11. Cipollini M, Landi S, Gemignani F. MicroRNA binding site polymorphisms as biomarkers in cancer management and research. Pharmgenomics Pers Med. 2014;7:173–191.
12. Malhotra P, Read GH, Weidhaas JB. Breast Cancer and miR-SNPs: the Importance of miR Germ-Line Genetics. Noncoding RNA. 2019;5(1).
13. Saetrom P, Biesinger J, Li SM, et al. A risk variant in an miR-125b binding site in BMPR1B is associated with breast cancer pathogenesis. Cancer Res. 2009;69(18):7459–7465. doi:10.1158/0008-5472.CAN-09-1201
14. Nicoloso MS, Sun H, Spizzo R, et al. Single-nucleotide polymorphisms inside microRNA target sites influence tumor susceptibility. Cancer Res. 2010;70(7):2789–2798. doi:10.1158/0008-5472.CAN-09-3541
15. Hollestelle A, Pelletier C, Hooning M, et al. Prevalence of the variant allele rs61764370 T>G in the 3ʹUTR of KRAS among Dutch BRCA1, BRCA2 and non-BRCA1/BRCA2 breast cancer families. Breast Cancer Res Treat. 2011;128(1):79–84. doi:10.1007/s10549-010-1080-z
16. Ovarian Cancer Association Consortium BCAC, Consortium of Modifiers of B, Brca, et al. No clinical utility of KRAS variant rs61764370 for ovarian or breast cancer. Gynecol Oncol. 2016;141(2):386–401.
17. Paranjape T, Heneghan H, Lindner R, et al. A 3ʹ-untranslated region KRAS variant and triple-negative breast cancer: a case-control and genetic analysis. Lancet Oncol. 2011;12(4):377–386. doi:10.1016/S1470-2045(11)70044-4
18. Pilarski R, Patel DA, Weitzel J, et al. The KRAS-variant is associated with risk of developing double primary breast and ovarian cancer. PLoS One. 2012;7(5):e37891. doi:10.1371/journal.pone.0037891
19. Zheng H, Song F, Zhang L, et al. Genetic variants at the miR-124 binding site on the cytoskeleton-organizing IQGAP1 gene confer differential predisposition to breast cancer. Int J Oncol. 2011;38(4):1153–1161.
20. Song F, Zheng H, Liu B, et al. An miR-502-binding site single-nucleotide polymorphism in the 3ʹ-untranslated region of the SET8 gene is associated with early age of breast cancer onset. Clin Cancer Res. 2009;15(19):6292–6300. doi:10.1158/1078-0432.CCR-09-0826
21. Zhang L, Liu Y, Song F, et al. Functional SNP in the microRNA-367 binding site in the 3ʹUTR of the calcium channel ryanodine receptor gene 3 (RYR3) affects breast cancer risk and calcification. Proc Natl Acad Sci U S A. 2011;108(33):13653–13658.
22. Pineros M, Cendales R, Murillo R, Wiesner C, Tovar S. [Pap test coverage and related factors in Colombia, 2005]. Rev Salud Publica (Bogota). 2007;9(3):327–341. Spanish.
23. Torres D, Bermejo JL, Rashid MU, et al. Prevalence and penetrance of BRCA1 and BRCA2 germline mutations in colombian breast cancer patients. Sci Rep. 2017;7(1):4713. doi:10.1038/s41598-017-05056-y
24. Torres D, Lorenzo Bermejo J, Garcia Mesa K, et al. Interaction between genetic ancestry and common breast cancer susceptibility variants in Colombian women. Int J Cancer. 2019;144(9):2181–2191. doi:10.1002/ijc.32023
25. Laitinen J, Samarut J, Holtta E. A nontoxic and versatile protein salting-out method for isolation of DNA. Biotechniques. 1994;17(2):316,318, 320–322.
26. Ruiz-Linares A, Adhikari K, Acuna-Alonzo V, et al. Admixture in Latin America: geographic structure, phenotypic diversity and self-perception of ancestry based on 7342 individuals. PLoS Genet. 2014;10(9):e1004572. doi:10.1371/journal.pgen.1004572
27. Westfall PH, Zaykin DV, Young SS. Multiple tests for genetic effects in association studies. Methods Mol Biol. 2002;184:143–168.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.