")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
Genetic Effects of DISC1 and G72 (DAOA) on Visual Learning of Patients with Schizophrenia
Authors Chang JP, Huang KH , Lin CH , Lane HY
Received 24 October 2019
Accepted for publication 26 February 2020
Published 20 March 2020 Volume 2020:16 Pages 771—780
DOI https://doi.org/10.2147/NDT.S235675
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jun Chen
Jane Pei-Chen Chang,1,* Kuo-Hao Huang,1,* Chieh-Hsin Lin,2,3 Hsien-Yuan Lane1,3,4
1Department of Psychiatry & Brain Disease Research Center, China Medical University Hospital, Taichung, Taiwan; 2Kaohsiung Chang Gung Memorial Hospital, Chang Gung University College of Medicine, Kaohsiung, Taiwan; 3Graduate Institute of Biomedical Sciences, China Medical University, Taichung, Taiwan; 4Department of Psychology, College of Medical and Health Sciences, Asia University, Taichung, Taiwan
*These authors contributed equally to this work
Correspondence: Chieh-Hsin Lin
Department of Psychiatry, Kaohsiung Chang Gung Memorial Hospital, No. 123, Dapi Road, Niaosong District, Kaohsiung 833, Taiwan
Tel +886-7-7317123 ext. 8753
Fax +886-7-7326817
Email [email protected]
Hsien-Yuan Lane
Department of Psychiatry, China Medical University Hospital, No. 2, Yuh-Der Road, Taichung 404, Taiwan
Tel +886-4-22062121 ext. 1074
Fax +886-4-2236-1230
Email [email protected]
Background: Visual learning plays an important role in general populations and patients with schizophrenia. Genetic influences on visual learning remain unknown. Two functional single nucleotide polymorphisms (SNPs), Ser704Cys of the DISC1 gene and M24 (rs1421292) of the G72 gene, are strongly associated with pathogenesis and pathophysiology of schizophrenia. This study examined these two SNPs’ effects on visual learning in schizophrenia patients.
Methods: Two hundred seventy-one patients (mean age, 37.0 years [SD = 9.3]; 159 men) with chronic schizophrenia were genotyped for the DISC1 Ser704Cys and G72 M24 SNPs and assessed for visual learning with Visual Reproduction II (delayed reproduction) of Wechsler Memory Scale – III (WMS-III). For comparison, verbal learning (using Word list II of WMS-III) and attention (by Continuous Performance Test) were also measured.
Results: The DISC1 Ser carriers excelled DISC1 Cys/Cys homozygotes in visual learning (p=0.004, effect size: 0.43), but not in other cognitive functions. G72 M24 A-allele carriers and G72 M24 T/T homozygotes performed similarly (effect size: 0.07). In SNP-SNP interaction analysis, the patients with Ser carrier_T/T had better visual learning than those with Cys/Cys_T/T (p=0.004, effect size: 0.70) and those with Cys/Cys_A-allele carrier (p=0.003, effect size: 0.65). Education had a positive effect (p=0.007), while negative symptoms had a negative effect (p< 0.001) on visual learning.
Conclusion: The findings suggest that genetic variations in DISC1 Ser704Cys and G72 M24 affect visual learning in schizophrenia patients. The effect sizes of SNP-SNP interaction surpassed the sum (0.50) of effect sizes from two individual genes, suggesting synergistic DISC1-G72 interaction.
Keywords: attention, DISC1, G72, visual and verbal learning, schizophrenia
Introduction
Visual learning plays an important role in daily life,1 while schizophrenia is a debilitating disorder with significant visual learning dysfunction.2 Visual learning, one of the seven cognitive domains (speed of processing, attention/vigilance, working memory, verbal learning, visual learning, reasoning and problem solving, and social cognition) of NIMH-recommended MATRICS (Measurement and Treatment Research to Improve Cognition in Schizophrenia),3 has gained growing attention for search of endophenotypes of schizophrenia.4–10 Three stages of visual learning/memory include encoding of visual perception, visual memory consolidation, and visual memory retrieval. Episodic visual memory encoding, involving visual attention and elaboration, relies heavily on the frontal lobes and visual association cortices.11 Visual memory consolidation depends mainly on the right medial-temporal lobe (MTL), particularly the hippocampus.11 The right MTL recapitulates the visual learning associated activation pattern and strengthens connections across the relevant lateral cortices.11 MTLs and lateral cortices are necessary for retrieving unconsolidated and consolidated visual memories, respectively.11 Visual memory impairments, mediated by organizational deficits,5 reflect prefrontal cortex mediated executive dysfunction in patients with schizophrenia;12 while deficits of visual perception are related with their premorbid social functioning,13 illness severity and chronicity.2 Whereas visual memory deficits reflect organization skill deficits,5 memory encoding and consolidation act as prognostic predictors of the illness.2,5 Of note, in patients with schizophrenia, visual memory declines with duration of illness.5 In addition, both verbal and visual memory abnormalities are well-established features of the schizophrenia spectrum, such as schizotypal personality disorder.6 Interestingly, visual memory and verbal memory show distinct generational trajectories and visual memory may offer a better potential than verbal memory to predict future risk of developing the disease.9 Further, visual memory deficits are related with anhedonia in schizophrenia patients, but not with that in psychotic bipolar patients or healthy controls.10 However, studies exploring visual learning/memory have been far less extensive and consistent than the studies on verbal memory.14–16
The disrupted-in-schizophrenia (DISC1) gene, disrupted by the chromosome 1 breakpoint of a balanced t(1;11) translocation,17,18 and G72 (also known as D-amino acid oxidase activator, DAOA) gene15,19,20 have both been strongly associated with schizophrenia and hippocampal dysfunction. DISC1 is expressed in neurons and glial cells, and greatly impacts the neurodevelopmental processes including neuritic outgrowth, neuronal migration, synaptogenesis, and glutamatergic transmission.18 Glutamate synapse activation leads to N-methyl-D-aspartic acid receptor (NMDAR) triggered long-term potentiation. Reduced expression of DISC1 binding patterns in postmortem frontal and parietal cortices and hippocampus of schizophrenia patients further implicates the role of DISC1 in schizophrenia pathogenesis.21 Altered DISC1 expression in orbitofrontal cortex, linking to dorsolateral prefrontal cortex (DLPFC) and verbal working memory, has been reported in schizophrenia.14 DISC1 genetic variance was associated with neurocognitive dysfunctions, such as deficits in the P300 event-related potential, verbal working memory, verbal attention,4 and verbal memory,14 but not yet visual learning/memory in schizophrenia. One study reported the association between DISC1 haplotype, HEP3 (containing two SNPs, T-A allele at rs751229 and rs3738401), and poor short-term visual attention and visual working memory.4 A common DISC1 nonsynonymous SNP at amino acid 704 (Ser704Cys, rs821616) leading to a serine-to-cystine substitution is associated with altered hippocampal structure and function, impaired verbal working memory, and increased risk for schizophrenia.22 DISC1-Cys allele was also associated with reduced hippocampal grey matter volume23 and impaired cognitive ability in women.24 Since visual memory consolidation depends mainly on the right MTL, particularly the hippocampus,11 the potential influence of the DISC1 SNP at amino acid 704 on visual memory deserves study too.
Another susceptibility gene, G72, modulates NMDAR neurotransmission via activating D-amino acid oxidase (DAAO) to oxidize D-serine, a potent NMDAR agonist,25 to hydroxy-pyruvate.26 Hence, G72 expression with DAAO activation results in decreased NMDAR neurotransmission and schizophrenia phenotype presentation.25 However, G72 regulation for DAAO activity requires further elucidation.26 Schizophrenia is strongly associated with markers in the 3ʹ region of G72, especially marker M24 (rs1421292).19 G72 genetic abnormalities lead to altered D-serine metabolism and NMDAR neurotransmission.26 G72 regulates prefrontal synaptic dopamine and is upregulated in postmortem DLPFC of schizophrenia patients.21 T-allele of M24 compared to A-allele predicts cognitive impairments and altered cortical activity.19 T/T genotype of G72 M24 genetic variance has been associated with decreased hippocampal activation, increased frontal lobe activation during episodic memory encoding and retrieval,19 verbal fluency tasks16 and verbal sentence completion tasks15 in both healthy and high-risk individuals. G72 M24 genetic variances also affect hippocampal complex and prefrontal cortex function in high-risk individuals.15 As aforementioned, since visual memory consolidation depends mainly on the hippocampus,11 the potential influence of G72 M24 genetic variance on visual memory deserves study too.
Structural and functional studies have converged on the potential regulatory effects of DISC1 and G72 on verbal learning/memory.14,19 DISC1 genetic variances, not including the Ser704Cys SNP, have been associated with visual attention,4 while there is no study on G72 and visual learning/memory. Hence, whether DISC1 and G72 influence visual learning/memory in schizophrenia patients deserves further study. To our knowledge, this is the first genetic study on visual learning in humans, aiming to investigate the role of Ser704Cys SNP (rs821616) of DISC1 and M24 SNP (rs1421292) of G72 in neurocognition, with emphasis on visual learning/memory, of patients with schizophrenia.
Methods
Subjects
The study was approved by institutional review board of China Medical University Hospital, Taiwan and conducted in accordance with the current revision of the Declaration of Helsinki.
Han Chinese schizophrenia patients were recruited with the inclusion criteria of 1) having sufficient education to communicate effectively and complete the assessments of the study; 2) being interviewed by experienced research psychiatrist with the Structured Clinical Interview for DSM-IV27 to confirm the diagnosis of schizophrenia and exclude other psychiatric disorders, including personality disorders and mental retardation; 3) aged 18–65; 4) physically healthy determined by physical examination and laboratory tests (normal blood routine and biochemical tests); and 5) keeping stable dosages of antipsychotic treatment for at least 2 months before enrollment. Patients were excluded when they currently 1) presented with other comorbid psychiatric disorders, substance use disorder or mental retardation; 2) had other existing physical or neurological illnesses; and 3) failed to cooperate with the study.
The participants were 271 unrelated patients with chronically stable schizophrenia, 159 men and 112 women, with a mean age of 37.0 years (SD = 9.3), a mean education level of 11.2 years (SD = 2.4), and a mean duration of illness of 162.0 months (SD = 102.2). After complete description, all participants provided written informed consent.
Clinical Assessments
The Positive and Negative Syndrome Scale (PANSS)28 was used to assess the clinical symptoms. Clinical ratings were performed by trained and experienced research psychiatrists. Inter-rater reliability was analyzed with the ANOVA test. Only raters reaching intra-class correlation coefficients of ≥0.90 during pre-study training were allowed for rating. Raters met at least once a month for training and reliability re-testing to maintain high interrater reliability and prevent rater drift.
Measurement of Neurocognitive Function
Patients’ cognitive functions, including verbal learning, visual learning, and attention were measured. Word listing II of Wechsler Memory Scale – III (WMS-III)29 was applied to measure “verbal learning”; Visual Reproduction II of the WMS-III was used for “visual learning”; and the d’ value of Continuous Performance Test (CPT)30 was employed to assess “sustained attention.”
In “Word list I” of WMS-III, the subjects recalled immediately after a word list of 12 items was read to them each time for four times, and the Word list I score was the number of items correctly recalled after four repeats of the 12 items. A second “interfering” word list without any of the previous 12 items was introduced to the subjects and subjects needed to recall the words from the first word list. In “Word list II,” the subjects recalled as much of the first word list after a 30-min interval since the first list had been initially introduced, and the Word list II score was the number of items correctly recalled after the 30-min interval.
In “Visual Reproduction I,” subjects drew a figure 10 s after the figure was shown to them. Subjects were shown a total of 7 figures. The first 3 figures were shown with a one figure per page display and the remaining 4 figures with a 2 figures per page display. In the “Visual Reproduction II or delayed visual reproduction,” the subjects recalled and drew the figures 30 mins later.
Genotyping
For determining the genotype of DISC1-Ser704Cys SNP (rs821616) from venous blood samples, polymerase chain reaction (PCR)-restriction fragment length polymorphism (RFLP) analysis procedures17 were applied with DISC1-F (AGGCCATGTGAAAAGGACAG) and DISC1-R (GTCTCAGCTGCAAGTGTCCA). PCR conditions were: 95ºC for 4 min initial heating, followed by 35 cycles of 95ºC for 30 sec, 61ºC for 30 sec, and 72ºC for 30 sec; and finally, 72ºC for 7 min. Subjects with Cys/Cys-DISC1 allele gave rise bands of 294 base pairs (bp); Ser/Ser allele, 225 bp and 69 bp; and Ser/Cys allele, 294 bp, 225 bp and 69 bp.
The G72 M24 SNP genotyping was performed using the Taqman SNP genotyping assay (ABI: Applied Biosystems Inc., Foster City, CA, USA). The primers and probes of SNPs were from ABI Company. The PCR reaction was conducted in 15μL reaction volume, containing 0.4 μL DNA sample (50 ng), 7.5 μL Master mix (Roche), and 0.4 μL 40x primer pairs and probes. A pre-incubation at 95°C for 10 min was employed to activate the Hot-Start DNA polymerase and denature DNA and was followed by 40 amplification cycles of 92°C denaturation for 15 sec, and 60°C for 60 sec. The probe fluorescence signal detection was performed using the ABI Prism 7500 Real-Time PCR System. The results were analyzed with SDS software 2.0 using the allelic discrimination assay program.
Data Analysis
All statistical analysis was carried out with Statistical Package for the Social Science (SPSS), version 22.0 for windows. Deviation of the genotype counts from the Hardy–Weinberg equilibrium was tested using a Chi-square goodness-of-fit test. Demographic and clinical characteristics and cognitive function of patients among genotypes were compared by Chi-square test, Fisher’s exact test, independent sample t test or one-way ANOVA where appropriate. Here, since the number of subjects homozygous for Ser allele was small (n= 5), we grouped Ser carriers together; that is, DISC1-Ser/Ser and DISC1-Ser/Cys patients were combined as DISC1-Ser carriers to compare with DISC1-Cys/Cys homozygotes.24 Meanwhile, A-allele carriers of G72 M24 (A/A and A/T) were combined and compared to G72 M24-T/T homozygotes.16
Multiple regressions were used to compare neurocognitive functions between DISC1-Ser carrier and DISC1 Cys/Cy homozygotes, between G72 M24-A carriers and G72 M24-T/T homozygotes, and among four diplotype groups (1. DISC-Ser carrier with G72-M24 T/T, 2. DISC1-Ser carrier with G72 M24-A carrier, 3. DISC1-Cys/Cys with G72 M24-T/T, and 4. DISC1-Cys/Cys with G72 M24-A carrier) while controlling confounding factors, such as age, onset age, typical/atypical antipsychotics, education years, PANSS-Positive subscale score and PANSS-Negative subscale score. Age,31 onset age,32 illness duration,33 typical/atypical antipsychotic,34 education years35 and active symptoms2 have been associated with effects on cognitive function and visual learning/memory.
Results
Genotype Distribution
The genotype distribution of the DISC1-Ser704Cys SNP was Cys/Cys in 212 patients and Ser carrier in 59 patients (including 54 Ser/Cys and 5 Ser/Ser 59 patients). The distribution was in accordance with Hardy–Weinberg equilibrium (χ2 = 0.66, d.f.= 1, p > 0.05). The frequency (98%) of Cys allele was similar to those of other Han Chinese studies,17,36 but higher than that (69.2%) in Caucasian populations.22
Regarding the genotype distribution of G72 M24 SNP, there were 161 A-allele carriers (48 AA and 113 TA) and 110 TT homozygotes. The A allele frequency (59%) was lower than that of the Japanese population (75%),37 but higher than that (44%) in Caucasian populations.38 This is the first study exploring G72 M24 SNP on Han Chinese patients with schizophrenia.
Cognitive Functions Among Genotype Groups
As shown in Figure 1, the three DISC1 genotypes showed an allele-dose effect on Visual Reproduction II performance, while the three G72 M24 genotypes had no significant influence. Univariate regression was used for comparison among the three genotype groups: compared to the performance (4.7±2.6) of the patients who carried no Ser allele (Cys/Cys homozygotes), those who carried 1 Ser allele (Ser/Cys heterozygotes) had better visual learning ability (5.8±2.7, t=2.593, P=0.010) and those who carried 2 ser alleles (Ser/Ser homozygotes) also tended to perform better (6.8±3.9, t=1.735, P=0.084), albeit insignificantly perhaps due to the very small sample size of the Ser/Ser group (n=5). We, therefore, grouped Ser carriers (DISC1 Ser/Ser and DISC1 Ser/Cys patients) as a group to compare with DISC1 Cys/Cys homozygotes;24 meanwhile, A-allele carriers of G72 M24 (A/A and A/T) were combined and compared to G72 M24 T/T homozygotes (Table 1).16
Demographic characteristics, clinical manifestations, CPT, and Word list II between DISC1 genotypes (Cys homozygotes vs Ser carriers) and between G72 M24 genotypes (TT homozygotes vs A carriers) were similar (Table 1). Of note, the Ser carriers excelled Cys homozygotes in Visual Reproduction II performance (t= 2.912, p=0.004), while TT homozygotes and A carriers performed similarly (Table 1).
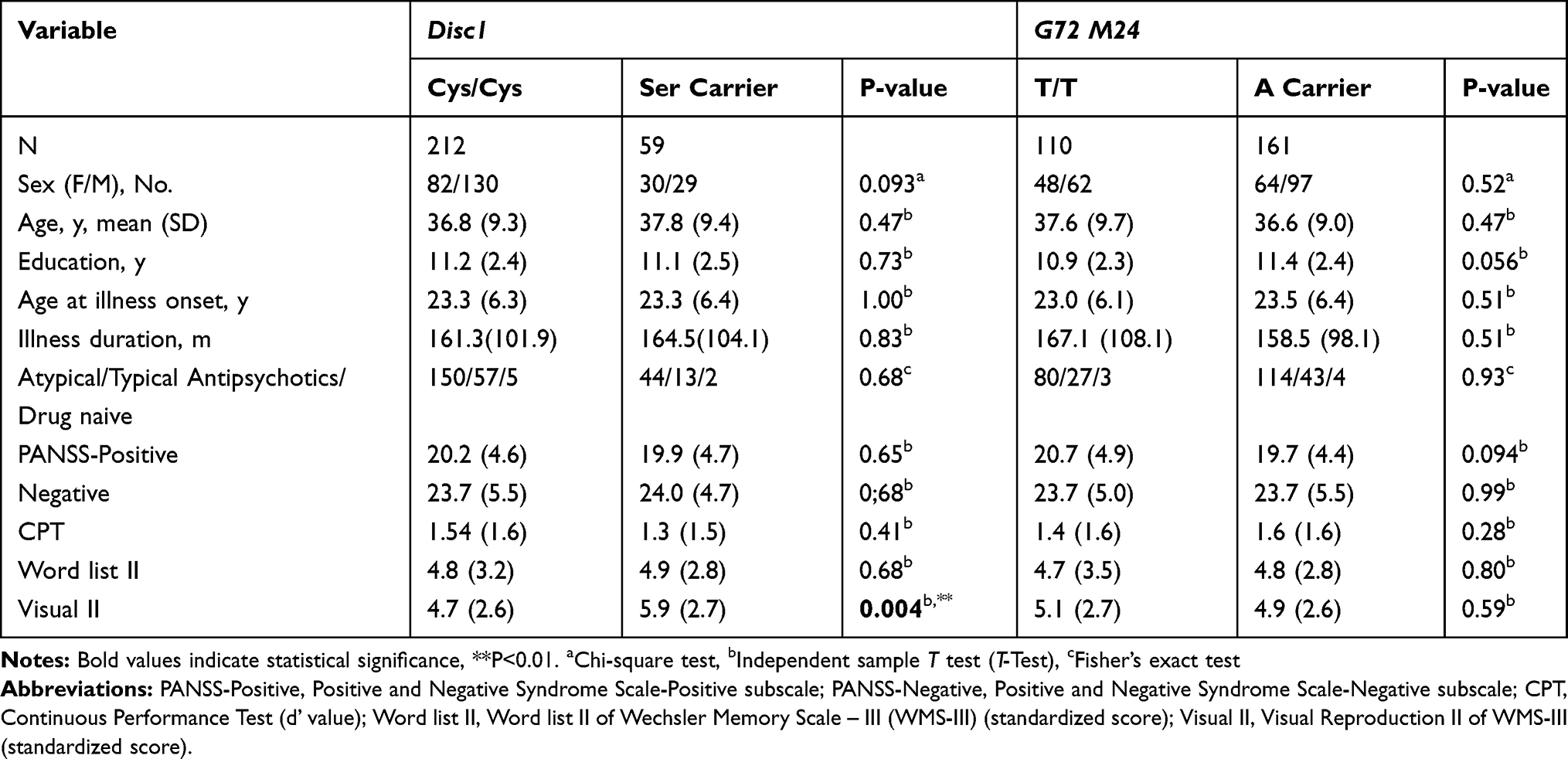 |
Table 1 Demographic Characteristics, Clinical Features, and Cognitive Functions of Schizophrenia Patients with DISC1 Ser704Cys and G72 M24 Genotypes |
Cognitive Functions Among Diplotype Groups
Demographic characteristics, clinical manifestations, CPT, and Word list II among the 4 diplotype groups (Ser carrier_TT, Ser carrier_A carrier, Cys/Cys_TT, Cys/Cys_A carrier) were similar (Table 2). However, the four diplotype groups differed significantly in Visual Reproduction II performance (F= 3.621, p=0.014) (Table 2).
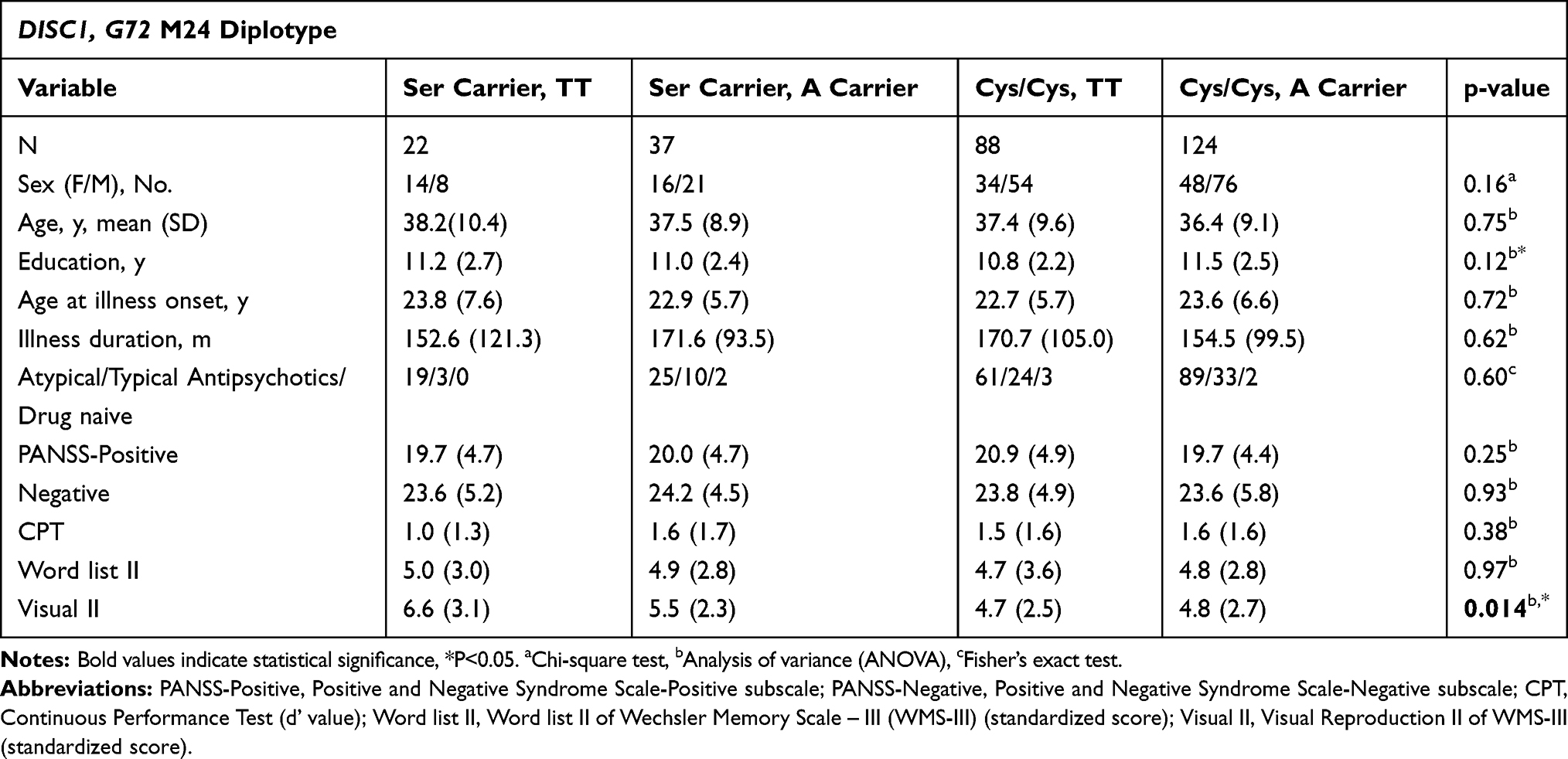 |
Table 2 Demographic Characteristics, Clinical Features, and Cognitive Functions of Schizophrenia Patients with Different DISC1 Ser704Cys and G72 M24 Diplotypes |
Moreover, Cys/Cys_T/T group (4.7±2.5, p=0.004) and Cys/Cys_A carrier group (4.8±2.7, p=0.003), yet not Ser carrier_A carrier group (5.5±2.3, p=0.125), had poorer Visual Reproduction II scores when compared to Ser carrier_T/T group (6.6±3.1) by univariate regression (Figure 1). Furthermore, after controlling demographic and clinical variables (Table 3) using multiple regression, education duration had positive effect (p=0.007), while negative symptoms (p< 0.001), Cys/Cys_T/T (p=0.010) diplotype, and Cys/Cys_A carrier (p=0.002) diplotype had negative effects on Visual Reproduction II scores (Table 3).
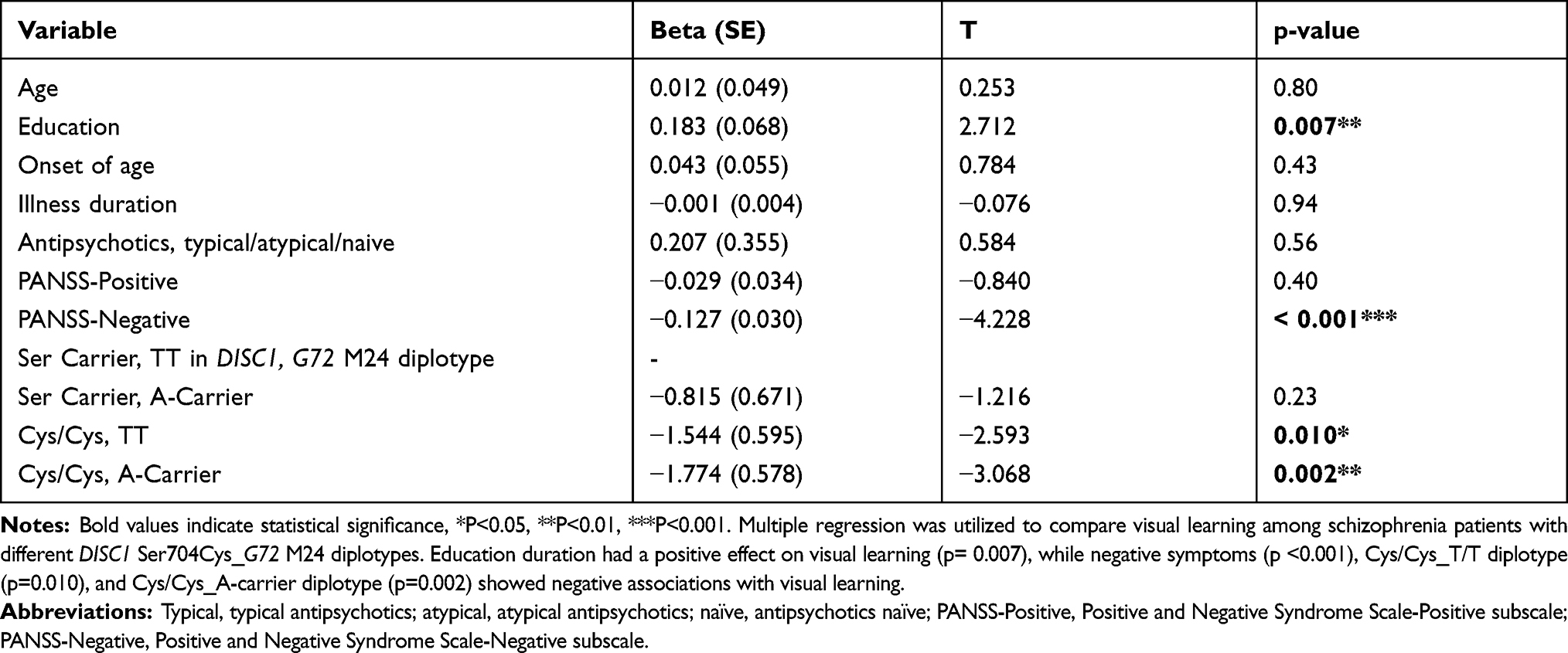 |
Table 3 Clinical and Genetic Variables Affecting Visual Learning, Represented by Standardized Score of Delayed Visual Reproduction of Wechsler Memory Scale – III |
Synergistic Effect of DISC1Ser704Cys and G72 M24 SNP on Visual Learning
We then evaluated genetic effects on delayed visual memory by the effect size of the two SNPs and their combination on delayed visual memory. The effect size on delayed visual memory for DISC1 Ser704Cys SNP was 0.43 and for G72 M24 SNP was 0.07 (Figure 2). The additive effect size of both SNPs was 0.50 (= 0.43 + 0.07). Effect sizes of Cys/Cys_T/T (0.70) and Cys/Cys_A-allele (0.65) on delayed visual memory were greater than the additive effect size (0.50) (Figure 2). This implicated that the effect of DISC1Ser704Cys and G72 on visual learning appeared to be synergistic.
Discussion
The first major finding of this study is that DISC1-Ser carriers performed better than DISC1-Cys/Cys homozygotes in visual learning (as represented by delayed visual reproduction), but not in verbal learning or attention. Secondly, the patients with the diplotype of DISC1-Ser carrier_G72 M24-T/T surpassed DISC1-Cys/Cys_G72 M24-T/T or DISC1-Cys/Cys_G72 M24-A carrier in visual learning, with a larger effect size than the sum of the effect sizes of DISC1 and G72 M24 individually, implying that the interactive gene effect of DISC1-G72 was additive (Figure 2). The finding, therefore, lends support to the notion that both DISC1 and G72 are involved in visual learning and suggest that the two genes interact synergistically.
Visual memory/learning has not yet been widely investigated in schizophrenia patients. Hennah et al are the first to report the significance of visual attention and visual working memory in the pathogenesis of schizophrenia,4 while others emphasized on verbal memory/learning.14,15,19 Visual learning involves the ability to store and retrieve previously experienced visual sensations and perceptions when the stimuli that originally evoked them are no longer present.11 And one, with intact memory encoding and consolidation, should be able to place in and retrieve memory information that resembles objects, places, animals or people in sort of a mental image from “the mind’s eye.”11 Moreover, visual perception and memory often perceive image as a whole and not in pieces, and group images according to six features: proximity, similarity, closure, symmetry, common fate and continuity.39 It is sensitive in ensembling characteristics of complex objects such as face,39 and its impairments are associated with poor social functioning13 and illness severity2 in schizophrenia.
Furthermore, visual learning deficits, mediated by organization skill deficits5 and poor memory encoding,5 contribute to poor planning skills5 and executive dysfunction.12 Patients with schizophrenia often use a piecemeal approach and have difficultly processing the gestalt of a complex visual stimulus in visual learning evaluation.7 Hence, impaired processing of global features in a local-global paradigm in these patients40 often results in overemphasis on non-critical features and depletion of available attention resources before they can process the critical features or interpret wholes as meaningful gestalts;40 which further implies their inefficient organizational strategy.7 Deficits in visual perception, visual memory encoding and consolidation all contribute to visual learning deficits. Li et al also found that Cys-allele carriers have less efficient information transfer than Ser homozygotes.41 Cys-allele carriers had a significantly lower global efficiency of their brain networks and decreased white matter integrity.41
Animal genetic study has demonstrated a close association of DISC1, cognitive functions, and hippocampal function/structure.18 Human studies have also shown an association of several DISC1 haplotypes, such as HEP1 (rs6675821, rs1000731, rs3890280), HEP2/HEP3 segment (rs1615409, rs766288, rs751229, rs3738401), with altered cognitive function.4,14,24 Cys allele has been associated with impaired verbal learning/long-term memory in schizophrenia and normal controls;14,24 while a few DISC1 haplotypes, but not including the SNP of the current study, for example, HEP2/HEP3 segment (rs1615409, rs766288, rs751229, rs3738401), HEP3 (rs751229, rs3738401) and hCV1650649 were associated with visual attention/search, visual working memory, and visuospatial memory.4,14 On the other hand, G72 M24-TT homozygotes were associated with decreased hippocampal activation and increased prefrontal activation in fMRI during episodic memory encoding and retrieval, working memory and verbal completion tasks.15,19,20 We are the first group to report the association of genetic effects between DISC1 Ser704Cys SNP and G72 M24 SNP and delayed visual memory/learning impairment. Interaction of the SNPs on visual memory was supported by a greater effect size of SNP combination (0.70, 0.65) than the sum of each SNP (0.50) (Figure 2). Unlike previous studies,22,24 no association was shown between DISC1 Ser704Cys SNP and G72 M24 SNP, and verbal learning/memory. This may be due to different neuropsychological assessment tools used between ours and other studies.4,14 In addition, verbal memory rather than visual memory might be more susceptible to antipsychotic (dopamine) effects.42
Finding of poorer visual learning and delayed visual memory in Cys allele carriers are consistent with previous frontal-parietal network (insular cortex) disruption studies in schizophrenia patients,43 where the insular cortex of the temporal cortex is implied for general consolidation of visual recognition memories.43 Association of DISC1 Ser704Cys SNP and G72 M24 SNP and visual memory/learning impairment is further supported by the impact of the DISC1 Ser704Cys SNP on visual memory associated brain structural changes44 and the impact of the G72 M24 SNP on memory task associated brain activations.19 Cys allele has been associated with reduced supramarginal gyrus volume, which is part of inferior parietal lobe responsible for enactment effect specific for visual memory encoding.45
Visual memory recall predominantly activates the left superior frontal gyrus and the intraparietal cortex, whereas recall of visually learned locations activates the bilateral superior parietal cortices.11 DISC1 and G72 genetic variances are closely associated with these visual memory-associated brain areas including insular cortex, medial superior frontal and prefrontal gyri, and hippocampus.14,19,23 Ser/Ser homozygote with schizophrenia had similar or larger medial superior frontal gyrus and insular cortex volumes when compared with Cys carriers.44 Moreover, DISC1 Ser allele homozygotes compared to Cys allele carriers had greater hippocampal gray matter volume,23 which further supported the possible concept that schizophrenia patients with Cys allele had poorer visual memory (Figure 1).
The study also found that longer education duration benefited visual learning (Table 3). This finding is consistent with previous studies implying that higher Intelligent Quotient (IQ) favors better delayed visual memory performance.35 This study also found that negative symptoms had detrimental effects on visual learning. Negative symptoms have been associated with visual memory impairment in patients with schizophrenia.2
Limitations
This study has several limitations. Firstly, the distribution of Cys and Ser allele is different from the previous study.22 Frequency of Cys homozygotes is higher, while Ser homozygotes is lower in our study. This may be explained by the Han Chinese ethnicity of the subjects. However, the distribution of the DISC1 genotype is similar to the report of NCBI.36 Moreover, there is no control group for this study. The reported effects of the studied SNPs on visual memory may be general effects for any subject, which would mean that they are not specific to schizophrenia. Inclusion of a control group in future replication study is needed. Thirdly, we used Word List of WMS-III rather than California Verbal Test to assess verbal memory due to the limited availability of the Chinese version of the California Verbal Test. However, the measures from the two tests were highly correlated.46 Fourthly, the use of a single neuropsychological test to assess visual learning may be considered a shortcoming of the paper.
Conclusions
The present study is the first genetic study of visual memory in humans. It is also the first to investigate G72 M24 SNP in Han Chinese patients with schizophrenia and the first to explore the relationship between DISC1 Ser 704Cys and G72 M24 genetic effects and visual memory in schizophrenia patients. Study findings indicated that schizophrenia patients with the Cys/Cys_T/T or Cys/Cys_A carrier had poorer visual learning. The combination of the two SNPs also supported an additive effect of the individual SNPs, suggesting that DISC1 and G72 might interact synergistically and thereby accounting for variance in hippocampal structure, function, and activation.19,23 Callicot also suggested that the risk of developing schizophrenia and associated cognitive deficits may be due to haplotypes monitored by Ser704Cys rather than the SNP itself.22 Thus, investigating the roles of other genetic variants of DISC4,17,22 and their interactions with G72 genetic variances15,19,20 may further help search for the endophenotype and subtyping of schizophrenia patients and development of targeted treatments.
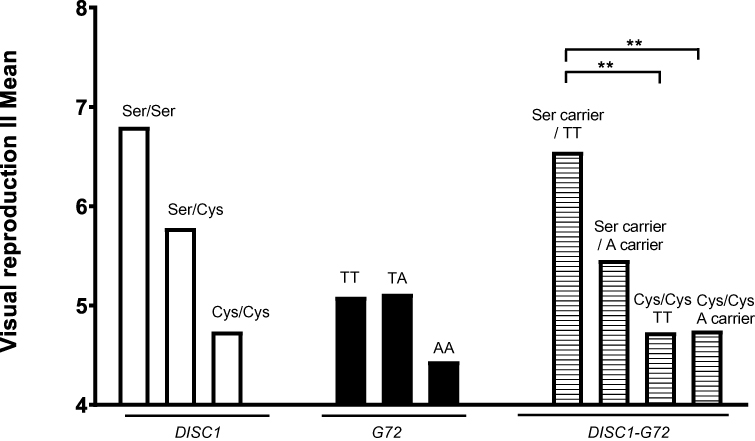 |
Figure 1 Influences of DISC1 Ser704Cy genotypes, G72 M24 (rs1421292) genotypes, and DISC1 Ser704Cys_G72 M24 interactions on visual learning, represented by means of Visual Reproduction II of Wechsler Memory Scale – III. **p< 0.01. |
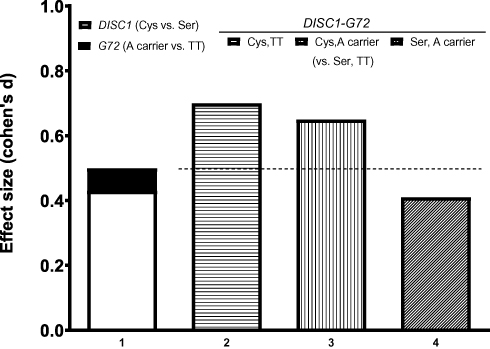 |
Figure 2 Effect sizes of the DISC1 genotype, G72 genotype, and DISC1_G72 interactions on visual learning, represented by Visual Reproduction II Wechsler Memory Scale – III. The dotted lines represent the sum of the effect sizes of the DISC1 and G72 individually. The DISC1 Ser carriers excelled DISC1 Cys/Cys homozygotes in visual learning (p=0.004, effect size: 0.43). Meanwhile, G72 M24 A-allele carriers and G72 M24 T/T homozygotes performed similarly (effect size: 0.07). In interaction analyses, the patients with Ser carrier_T/T had better visual learning than those with Cys/Cys_T/T (p= 0.004, effect size: 0.70) and those with Cys/Cys _A-allele carrier (p= 0.003, effect size: 0.65). Therefore, the effect sizes (0.70 and 0.65) of the DISC1_G72 interactions were larger than the sum (0.50 = 0.43 + 0.07) of the effect sizes of DISC1 and G72 individually, suggesting that the interactive gene effect of DISC1-G72 was more than addition. |
Acknowledgments
This work was funded by Ministry of Science and Technology, Taiwan (MOST 108-2314-B-039-002), National Health Research Institutes (NHRI-EX108-1073NI), China Medical University Hospital, Taiwan (DMR-106-102), and Taiwan Ministry of Health and Welfare Clinical Trial and Research Center of Excellence (MOHW108-TDU-B-212-133004).
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
The aforementioned institutes had no further role in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the paper for publication.
Disclosure
All authors declare that they have no conflicts of interest in this work.
References
1. Ko PC, Ally BA. Visual cognition in Alzheimer’s disease and its functional implications. In: The Clinical Spectrum of Alzheimer’s Disease-The Charge Toward Comprehensive Diagnostic and Therapeutic Strategies. IntechOpen; 2011.
2. O’leary DS, Flaum M, Kesler ML, Flashman LA, Arndt S, Andreasen NC. Cognitive correlates of the negative, disorganized, and psychotic symptom dimensions of schizophrenia. J Neuropsychiatry Clin Neurosci. 2000;12:4–15. doi:10.1176/jnp.12.1.4
3. Lo CH, Tsai GE, Liao CH, et al. Emotional management and 5-HT2A receptor gene variance in patients with schizophrenia. Biol Psychol. 2010;83:79–83. doi:10.1016/j.biopsycho.2009.11.002
4. Hennah W, Tuulio-henriksson A, Paunio T, et al. A haplotype within the DISC1 gene is associated with visual memory functions in families with a high density of schizophrenia. Mol Psychiatry. 2005;10:1097–1103. doi:10.1038/sj.mp.4001731
5. Seidman LJ, Lanca M, Kremen WS, Faraone SV, Tsuang MT. Organizational and visual memory deficits in schizophrenia and bipolar psychoses using the Rey-Osterrieth complex figure: effects of duration of illness. J Clin Exp Neuropsychol. 2003;25:949–964. doi:10.1076/jcen.25.7.949.16482
6. McClure MM, Romero MJ, Bowie CR, Reichenberg A, Harvey PD, Siever LJ. Visual-spatial learning and memory in schizotypal personality disorder: continued evidence for the importance of working memory in the schizophrenia spectrum. Arch Clin Neuropsychol. 2007;22:109–116. doi:10.1016/j.acn.2006.11.004
7. Kim MS, Namgoong Y, Youn T. Effect of organizational strategy on visual memory in patients with schizophrenia. Psychiatry Clin Neurosci. 2008;62:427–434. doi:10.1111/j.1440-1819.2008.01821.x
8. Girard TA, Christensen BK, DeGroote MG, Rizvi S. Visual-spatial episodic memory in schizophrenia: a multiple systems framework. Neuropsychology. 2010;24:368–378. doi:10.1037/a0018313
9. Maziade M, Rouleau N, Merette C, et al. Verbal and visual memory impairments among young offspring and healthy adult relatives of patients with schizophrenia and bipolar disorder: selective generational patterns indicate different developmental trajectories. Schizophr Bull. 2011;37:1218–1228. doi:10.1093/schbul/sbq026
10. Bodapati AS, Jenkins LM, Sharma RP, Rosen C. Visual memory uniquely predicts anhedonia in schizophrenia but not bipolar disorder. J Neuropsychol. 2019;13:136–146. doi:10.1111/jnp.2019.13.issue-1
11. Brewer JB, Zhao Z, Desmond JE, Glover GH, Gabrieli JD. Making memories: brain activity that predicts how well visual experience will be remembered. Science. 1998;281:1185–1187. doi:10.1126/science.281.5380.1185
12. Kim JJ, Kwon JS, Park HJ, et al. Functional disconnection between the prefrontal and parietal cortices during working memory processing in schizophrenia: a[15(O)]H2O PET study. Am J Psychiatry. 2003;160:919–923. doi:10.1176/appi.ajp.160.5.919
13. Silverstein SM, Knight RA, Schwarzkopf SB, West LL, Osborn LM, Kamin D. Stimulus configuration and context effects in perceptual organization in schizophrenia. J Abnorm Psychol. 1996;105:410–420. doi:10.1037/0021-843X.105.3.410
14. Cannon TD, Hennah W, van Erp TG, et al. Association of DISC1/TRAX haplotypes with schizophrenia, reduced prefrontal gray matter, and impaired short- and long-term memory. Arch Gen Psychiatry. 2005;62:1205–1213. doi:10.1001/archpsyc.62.11.1205
15. Hall J, Whalley HC, Moorhead TW, et al. Genetic variation in the DAOA (G72) gene modulates hippocampal function in subjects at high risk of schizophrenia. Biol Psychiatry. 2008;64:428–433. doi:10.1016/j.biopsych.2008.03.009
16. Krug A, Markov V, Krach S, et al. Genetic variation in G72 correlates with brain activation in the right middle temporal gyrus in a verbal fluency task in healthy individuals. Hum Brain Mapp. 2011;32:118–126. doi:10.1002/hbm.v32.1
17. Hwu HG, Liu CM, Fann CS, Ou-yang WC, Lee SF. Linkage of schizophrenia with chromosome 1q loci in Taiwanese families. Mol Psychiatry. 2003;8:445–452. doi:10.1038/sj.mp.4001235
18. Johnstone M, Thomson PA, Hall J, McIntosh AM, Lawrie SM, Porteous DJ. DISC1 in schizophrenia: genetic mouse models and human genomic imaging. Schizophr Bull. 2011;37:14–20. doi:10.1093/schbul/sbq135
19. Goldberg TE, Straub RE, Callicott JH, et al. The G72/G30 gene complex and cognitive abnormalities in schizophrenia. Neuropsychopharmacology. 2006;31:2022–2032. doi:10.1038/sj.npp.1301049
20. Jansen A, Krach S, Krug A, et al. Effect of the G72 (DAOA) putative risk haplotype on cognitive functions in healthy subjects. BMC Psychiatry. 2009;9:60. doi:10.1186/1471-244X-9-60
21. Rastogi A, Zai C, Likhodi O, Kennedy JL, Wong AH. Genetic association and post-mortem brain mRNA analysis of DISC1 and related genes in schizophrenia. Schizophr Res. 2009;114:39–49. doi:10.1016/j.schres.2009.06.019
22. Callicott JH, Straub RE, Pezawas L, et al. Variation in DISC1 affects hippocampal structure and function and increases risk for schizophrenia. Proc Natl Acad Sci U S A. 2005;102:8627–8632. doi:10.1073/pnas.0500515102
23. Di Giorgio A, Blasi G, Sambataro F, et al. Association of the SerCys DISC1 polymorphism with human hippocampal formation gray matter and function during memory encoding. Eur J Neurosci. 2008;28:2129–2136. doi:10.1111/j.1460-9568.2008.06482.x
24. Thomson PA, Harris SE, Starr JM, Whalley LJ, Porteous DJ, Deary IJ. Association between genotype at an exonic SNP in DISC1 and normal cognitive aging. Neurosci Lett. 2005;389:41–45. doi:10.1016/j.neulet.2005.07.004
25. Lane HY, Lin CH, Huang YJ, Liao CH, Chang YC, Tsai GE. A randomized, double-blind, placebo-controlled comparison study of sarcosine (N-methylglycine) and D-serine add-on treatment for schizophrenia. Int J Neuropsychopharmacol. 2010;13:451–460. doi:10.1017/S1461145709990939
26. Sacchi S, Bernasconi M, Martineau M, et al. pLG72 modulates intracellular D-serine levels through its interaction with D-amino acid oxidase: effect on schizophrenia susceptibility. J Biol Chem. 2008;283:22244–22256. doi:10.1074/jbc.M709153200
27. Association AP. Diagnostic and Statistical Manual of Mental Disorders (DSM-IV). American Psychiatric Pub; 1994.
28. Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13:261–276. doi:10.1093/schbul/13.2.261
29. Lo AH, Humphreys M, Byrne GJ, Pachana NA. Test-retest reliability and practice effects of the Wechsler memory scale-III. J Neuropsychol. 2012;6:212–231. doi:10.1111/j.1748-6653.2011.02023.x
30. Chen WJ, Liu SK, Chang CJ, Lien YJ, Chang YH, Hwu HG. Sustained attention deficit and schizotypal personality features in nonpsychotic relatives of schizophrenic patients. Am J Psychiatry. 1998;155:1214–1220. doi:10.1176/ajp.155.9.1214
31. Kennedy KM, Rodrigue KM, Head D, Gunning-dixon F, Raz N. Neuroanatomical and cognitive mediators of age-related differences in perceptual priming and learning. Neuropsychology. 2009;23:475–491. doi:10.1037/a0015377
32. Bellino S, Rocca P, Patria L, et al. Relationships of age at onset with clinical features and cognitive functions in a sample of schizophrenia patients. J Clin Psychiatry. 2004;65:908–914. doi:10.4088/JCP.v65n0705
33. Cuesta MJ, Peralta V, Zarzuela A. Illness duration and neuropsychological impairments in schizophrenia. Schizophr Res. 1998;33:141–150. doi:10.1016/s0920-9964(98)00068-1
34. Kertzman S, Reznik I, Grinspan H, Weizman A, Kotler M. Antipsychotic treatment in schizophrenia: the role of computerized neuropsychological assessment. Isr J Psychiatry Relat Sci. 2008;45:114–120.
35. Skelley SL, Goldberg TE, Egan MF, Weinberger DR, Gold JM. Verbal and visual memory: characterizing the clinical and intermediate phenotype in schizophrenia. Schizophr Res. 2008;105:78–85. doi:10.1016/j.schres.2008.05.027
36. NCBI. Reference SNP (rs) Report: rs821616. 2019.
37. Ohi K, Hashimoto R, Yasuda Y, et al. Association study of the G72 gene with schizophrenia in a Japanese population: a multicenter study. Schizophr Res. 2009;109:80–85. doi:10.1016/j.schres.2009.01.019
38. Schumacher J, Jamra RA, Freudenberg J, et al. Examination of G72 and D-amino-acid oxidase as genetic risk factors for schizophrenia and bipolar affective disorder. Mol Psychiatry. 2004;9:203–207. doi:10.1038/sj.mp.4001421
39. Bruce V, Green P, Georgeson M. Visual Perception: Physiology, Psychology, and Ecology. East Sussex, UK: Psychology Press; 1996.
40. Johnson SC, Lowery N, Kohler C, Turetsky BI. Global-local visual processing in schizophrenia: evidence for an early visual processing deficit. Biol Psychiatry. 2005;58:937–946. doi:10.1016/j.biopsych.2005.04.053
41. Li Y, Liu B, Hou B, et al. Less efficient information transfer in Cys-allele carriers of DISC1: a brain network study based on diffusion MRI. Cereb Cortex. 2013;23:1715–1723. doi:10.1093/cercor/bhs167
42. Chen PS, Yang YK, Lee YS, et al. Correlation between different memory systems and striatal dopamine D2/D3 receptor density: a single photon emission computed tomography study. Psychol Med. 2005;35:197–204. doi:10.1017/S0033291704003101
43. Nagai M, Kishi K, Kato S. Insular cortex and neuropsychiatric disorders: a review of recent literature. Eur Psychiatry. 2007;22:387–394. doi:10.1016/j.eurpsy.2007.02.006
44. Takahashi T, Suzuki M, Tsunoda M, et al. The disrupted-in-Schizophrenia-1 Ser704Cys polymorphism and brain morphology in schizophrenia. Psychiatry Res. 2009;172:128–135. doi:10.1016/j.pscychresns.2009.01.005
45. Russ MO, Mack W, Grama CR, Lanfermann H, Knopf M. Enactment effect in memory: evidence concerning the function of the supramarginal gyrus. Exp Brain Res. 2003;149:497–504. doi:10.1007/s00221-003-1398-4
46. McDowell BD, Bayless JD, Moser DJ, Meyers JE, Paulsen JS. Concordance between the CVLT and the WMS-III word lists test. Arch Clin Neuropsychol. 2004;19:319–324. doi:10.1016/S0887-6177(03)00023-4
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.