Back to Journals » Infection and Drug Resistance » Volume 17
Genetic Characteristics of Novel IncpSE5381-aadB Plasmids, Integrative and Mobilizable Elements, and Integrative and Conjugative Elements in Pseudomonas aeruginosa
Authors He J, Lu X, Yuan C, Zheng Y, Chen F, Luo J, Ma K, Yang F, Wang P ![]() , Zhou D, Wang L, Yin Z
, Zhou D, Wang L, Yin Z
Received 4 February 2024
Accepted for publication 11 May 2024
Published 25 May 2024 Volume 2024:17 Pages 2053—2068
DOI https://doi.org/10.2147/IDR.S462670
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Zhi Ruan
Jiaqi He,1,* Xiuhui Lu,2,* Chenchen Yuan,1 Yali Zheng,2 Fangzhou Chen,2 Jing Luo,2 Kejiao Ma,2 Fan Yang,2 Peng Wang,2 Dongsheng Zhou,2 Li Wang,1 Zhe Yin2
1Department of Clinical Laboratory, The First Affiliated Hospital of Henan University, Kaifeng, 475000, People’s Republic of China; 2State Key Laboratory of Pathogen and Biosecurity, Academy of Military Medical Sciences, Beijing, 100071, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li Wang, Department of Clinical Laboratory, The First Affiliated Hospital of Henan University, Kaifeng, 475000, People’s Republic of China, Email [email protected] Zhe Yin, State Key Laboratory of Pathogen and Biosecurity, Academy of Military Medical Sciences, Beijing, 100071, People’s Republic of China, Email [email protected]
Purpose: Pseudomonas aeruginosa is a common causative bacteria in nosocomial infections. This study aims to describe the structure and evolutionary characteristics of mobile genetic elements (MGEs) carrying antibiotic resistance genes (ARGs) from P. aeruginosa and to conduct bioinformatics and comparative genomic analysis to provide a deeper understanding of the genetic characteristics and diversity of MGEs in P. aeruginosa.
Methods: Fifteen clinical isolates of P. aeruginosa from China were collected and sequenced in this study, and 15 novel MGEs were identified. Together with four MGEs from GenBank, a total of 19 MGEs were used to perform detailed modular structure dissection and sequence comparison. Then, the biological experiments were carried out to verify the biological characteristics of these isolates and MEGs.
Results: The novel MGEs identified in this study displayed diversification in modular structures, which showed complex mosaic natures. The seven types of 19 MGEs included in this study were divided into three groups: i) novel MGEs (firstly identified in this study): four IncpSE5381-aadB plasmids and three Tn 7495-related integrative and mobilizable elements (IMEs); ii) newly defined MGEs (firstly designated in this study, but with previously determined sequences): four Tn 7665-related IMEs; iii) novel transposons with reference prototypes identified in this study: two Tn 6417-related integrative and conjugative elements (ICEs), two IS-based transposition units, two Tn 501-related unit transposons, two Tn 1403-related unit transposons. At least 36 ARGs involved in resistance to 11 different classes of antimicrobials and heavy metals were identified. Additionally, three novel blaOXA variants were identified. Antimicrobial susceptibility testing showed that these variants were resistant to some β-lactamase antibiotics and blaOXA-1204 was additionally resistant to cephalosporins.
Conclusion: The continuous evolution of ARG-carrying MGEs during transmission, leading to the emergence of novel MGEs or ARGs, which facilitates the spread of antibiotic resistance in P. aeruginosa and enhances the diversity of transmission modes of bacterial resistance.
Keywords: Pseudomonas aeruginosa, genome sequencing, mobile genetic elements, antimicrobial resistance, OXA
Introduction
Pseudomonas aeruginosa is a common causative Gram-negative bacteria in nosocomial infections, which are often associated with acute and chronic infections in immunocompromised patients (eg post-operative or severe burn patients).1 The increasing incidence and antibiotic treatment of P. aeruginosa infection has made the species resistant to multiple existing antibiotics, so much so that these bacteria have been classified as ESKAPE and pose a major challenge to clinical anti-infection treatment.2
β-lactam antibiotics are antimicrobial agents widely used to treat P. aeruginosa infection,3 the emergence of β-lactamase has become a crucial factor in exacerbating resistance of P. aeruginosa to these antibiotics. Based on amino acid homology, β-lactamases are divided into four classes (A – D). Class B enzymes comprise metal-β-lactamases, the others are serine-dependent β-lactamases.4,5 Class D β-lactamases, also known as OXA enzymes, are characterized by wide expression across bacterial species,6 with OXA-10 being the first class D β-lactamases detected in P. aeruginosa.7 These enzymes were frequently identified in clinical isolates and comprised the largest family of β-lactamases with more than 1200 variants identified. Most are narrow-spectrum, though some (eg OXA-11, OXA-16) show broad-spectrum behavior.8 Genes for these enzymes may be carried by plasmids or present on bacterial chromosomes, most have been identified in the gene cassette of class 1 integrons. P. aeruginosa usually carried class 1 integrons with an OXA-2-like or OXA-10-like gene cassette on their chromosomes.9
The spread of resistance genes is often linked to mobile genetic elements (MGEs), such as plasmids, transposons, and integrons, which are transmitted via horizontal gene transfer (HGT).10 In P. aeruginosa, resistance genes move to recipient bacteria by the HGT of plasmids, integrative and mobilizable elements (IMEs), or integrative and conjugative elements (ICEs).11,12 Conjugative plasmids encode products for facilitating conjugation, including relaxase (induction of DNA strand breaks and rejoining), type IV secretion systems (transmembrane channel linking donor and recipient bacteria and DNA transfer), oriT (origin of transfer), and coupling proteins (intracellular machinery to link the secretion system).13 IMEs typically utilize conjugation mechanisms encoded by other MGEs to achieve transfer and integration14 while ICEs mainly use tyrosine/serine recombinases or DDE transposases for chromosomal integration and excision.15 In previous study, a number of novel MGEs were identified in P. aeruginosa in China, such as the novel Inc groups: Incp60512−IMP and IncpPA7790;16 Tn6852-related IMEs carrying blaPER-1 and blaCARB-2: Tn6853;17 Tn6417-related ICEs carrying blaVIM-1: Tn6953;18 and Multiple Tn1403-related transposons carrying different class 1 integrons: Tn6847, Tn6849, Tn7454, Tn7484, etc.16,17,19 Multiple types of novel MGEs are frequently identified in China, which pose unknown challenges to the containment of bacterial resistance. It is necessary to pay close attention to the emergence of novel MGEs and their genetic evolution characteristics.
This study aims to describe the genetic structure and evolutionary characteristics of novel MEGs to explore the evolution and role of MGEs in the transmission of antibiotic resistance in P. aeruginosa. We sequenced 15 P. aeruginosa isolates from China. Detailed genetic dissection and comparison were applied to 19 MGEs, including 15 identified here and an additional four obtained from GenBank. These 19 MGEs were classified into seven types: IncpSE5381-aadB plasmids; Tn7495-related IMEs; Tn7665-related IMEs; Tn6417-related ICEs; IS-based transposition units; and two unit transposons. In addition, we also identified three novel variants of class D β-lactamases and characterized their antibiotic resistance phenotypes. These findings provide a deeper understanding of the genetic characteristics and diversity of MGEs in P. aeruginosa.
Materials and Methods
Bacterial Strains
In the initial stage of this study, more than 1000 drug-resistant P. aeruginosa bacteria were isolated from clinical samples in China for large-scale next-generation sequencing (NGS). After preliminary prediction and screening, the isolates carrying multiple ARGs or novel MGEs were screened for third-generation sequencing (TGS). Finally, 15 clinical isolates of P. aeruginosa carrying novel MGEs were included in this study. These fifteen P. aeruginosa isolates (Table S1) were isolated from various fluid specimens from patients in five different Chinese public hospitals from 2011 to 2018. Bacterial species were identified using average nucleotide identity analysis based on genome sequencing (http://www.ezbiocloud.net/tools/ani).20 Antimicrobial susceptibility testing was performed using VITEK 2 (Table S2) and interpreted according to the 2022 Clinical and Laboratory Standards Institute (CLSI) guidelines.21
Sequencing and Sequence Assembly
Bacterial genomic DNA was extracted using the UltraClean Microbial Kit (Qiagen, NW, Germany) and sequenced from a sheared DNA library with the average size of 15 kb (range: 10 to 20 kb) on a PacBio RSII sequencer (Pacific Biosciences, CA, USA). A paired-end library with an average insert size of 350 bp (range: 150 to 600 bp) was sequenced on a HiSeq sequencer (Illumina, CA, USA). Paired-end short Illumina reads were used to correct long PacBio reads with proovread,22 corrected reads were assembled using SMARTdenovo (https://github.com/ruanjue/smartdenovo). As mentioned earlier, further sequence data mining was carried out.23,24
Multi‑locus Sequence Typing (MLST)
Sequence types (STs) of P. aeruginosa isolates were identified according to the online P. aeruginosa MLST scheme (https://pubmlst.org/paeruginosa/). This method mainly performs sequence typing by comparing the diversity of the core sequences of multiple housekeeping genes.
Sequence Annotation and Comparison
Open reading frames (ORF) and pseudogenes were predicted using RAST 2.025 combined with BLASTP/BLASTN searches26 against the RefSeq27 and UniProtKB/Swiss-Prot databases.28 Resistance genes, MGEs, and their characteristics were annotated using online databases including ResFinder,29 Tn Number Registry,30 ISfinder,31 INTEGRALL,32 and CARD,33 as well as our published database, DANMEL.34 Multiple and pairwise comparisons were performed using BLASTN/BLASTX. Gene organization diagrams were created using Inkscape 1.0 (https://inkscape.org/en/).
Cloning Experiments
Coding regions of blaOXA-1202/1203/1204 together with 443-bp upstream (promoter) sequence and 160-bp downstream (terminator) sequence from isolates NY7570/SE5381/NY11384, respectively, were subcloned into the vector plasmid pUC57-Kan. In parallel, coding regions for the other blaOXA variants together with the above promoter- and terminator-proximal region were synthesized and subcloned into pUC57-Kan. Resulting recombinant plasmids were transformed into E. coli TOP10 by electroporation to generate the corresponding electroporant. 200mg/mL ampicillin (for blaOXA-1202/1203/1204) was used for electroporant selection and antimicrobial susceptibility testing was performed using the classic broth microdilution method and interpreted according to the 2022 Clinical and Laboratory Standards Institute (CLSI) guidelines.21
Nucleotide Sequence Accession Numbers
The complete chromosome sequences of SE5356, SE5416, NY11084, NY7723, NY5051, NY11352, SE5430, SE5431, NY7570, NY4383, NY3649, NY7583, SE5381, NY11384, and SE5418 and those plasmids of pNY7583-catB3, pSE5381-aadB, pNY11384-aadB, and pSE5418-aacA3 were submitted to GenBank under accession numbers CP120704, CP046404, CP120705, CP120708, CP120707, CP132352, CP054790, CP054791, CP120857, CP120859, CP124612, CP120881, CP054789, CP120883, CP054790, CP120882, CP121188, CP120884, CP123896, respectively.
Results
Identification of P. Aeruginosa Isolates
Identification of Seven STs and T3SS Virulotypes from the 15 Clinical P. Aeruginosa Isolates
Seven STs were identified from the 15 sequenced P. aeruginosa isolates, including ST235, ST244, ST292, ST606, ST697, ST740, and ST1284 (Table S1). Among these, eight belonged to ST235 or ST244, considered among the global top 10 P. aeruginosa high-risk clones. Four isolates, NY7583, SE5381, NY11384, and SE5418 belonged to ST244; four others, NY5051, NY11352, SE5430, and NY4383 belonged to ST235. Three isolates, SE5356, SE5416, and NY11084 belonged to ST697. Isolates NY7723, SE5431, NY7570, and NY3649 belonged to ST606, ST1284, ST740, and ST292, respectively.
The type 3 secretion system (T3SS) is a virulence system used by P. aeruginosa and exhibits widespread presence within the species. T3SS-associated virulence genes (exoT, exoY, exoS, and exoU) were identified through BLAST searches and PCR analysis (Table S1). Among the 15 isolates, exoT was detected in all (n=15), followed by exoY (n=13), exoS (n=9) and exoU (n=5). The most common virulotype was exoT+/exoY+/exoS+/exoU-.
Identification of 15 MGEs from the 15 P. Aeruginosa Isolates
Whole genome sequencing was performed on 15 P. aeruginosa isolates, leading to identification of 15 MGEs that subsequently underwent detailed genetic characteristic analysis: i) pNY7583-catB3, pSE5381-aadB, pNY11384-aadB, and pSE5418-aacA3 from isolates NY7583, SE5381, NY11384, and SE5418, respectively; ii) Tn7495, Tn7496a, and Tn7496b from isolates SE5356, SE5416, and NY11084, respectively; and iii) Tn7666, Tn7667, and T7665RENY11352 from isolates NY7723, NY5051, and NY11352, respectively; iv) Tn7671 from isolates NY7570; v) Tn7669 and Tn7670 from isolates SE5430 and SE5431, respectively; vi) Tn7672, Tn7673 from isolates NY4383, NY3649, respectively (Table 1).
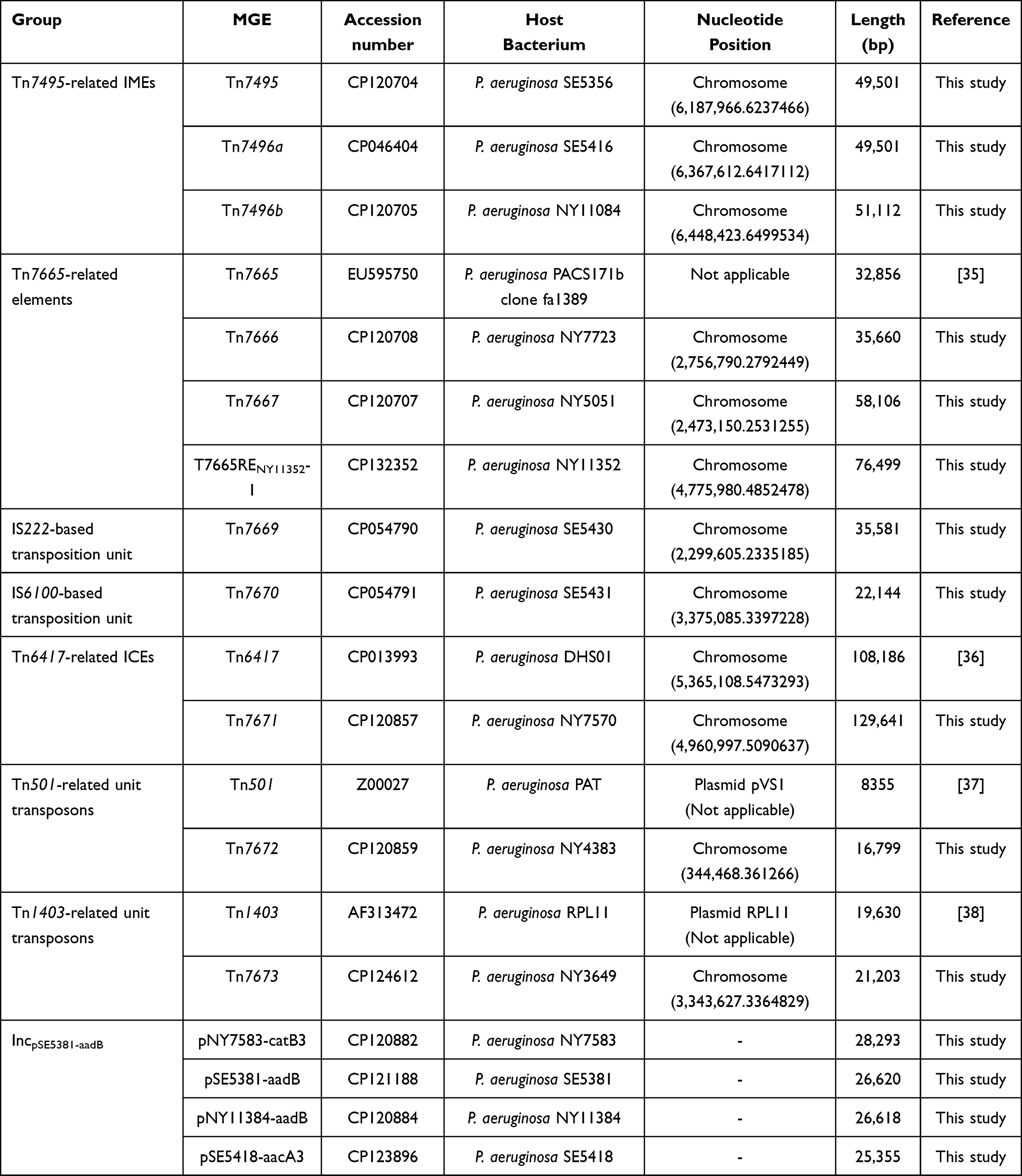 |
Table 1 Major Features of MGEs Characterized in This Work |
Collection of 19 MGEs for Detailed Sequence Comparison
A detailed genome sequence comparison was applied to a total of 19 MGEs (14 chromosome-borne MGEs and four reference/ prototype MGEs from GenBank: Tn7665,35 Tn6417,39 Tn501,40 and Tn140341) that were classified into seven types, which were divided into three groups in this study (Table 1): i) novel families of MGEs (firstly identified in this study): IncpSE5381-aadB plasmids (including pNY7583-catB3, pSE5381-aadB, pNY11384-aadB, pSE5418-aacA3) and Tn7495-related IMEs (including Tn7495, Tn7496a, and Tn7496b); ii) newly defined MGEs (firstly designated in this study, but with previously determined sequences): Tn7665-related IMEs (including Tn7665, Tn7666, Tn7667, and its derivatives T7665RENY11352); iii) novel transposons with reference prototypes identified in this study: Tn6417-related ICEs (including Tn6417 and Tn7671), IS-based transposition units (including Tn7669 and Tn7670), Tn501-related unit transposons (including Tn501 and Tn7672), Tn1403-related unit transposons (including Tn1403 and Tn7673). In addition, 12 novel class 1 integrons were identified in these 19 MGEs. At least 36 antibiotic resistance genes (ARGs), involved in resistance to 11 different categories of antimicrobials and heavy metals, were identified among these 19 MGEs (Figure 1 and Table S3).
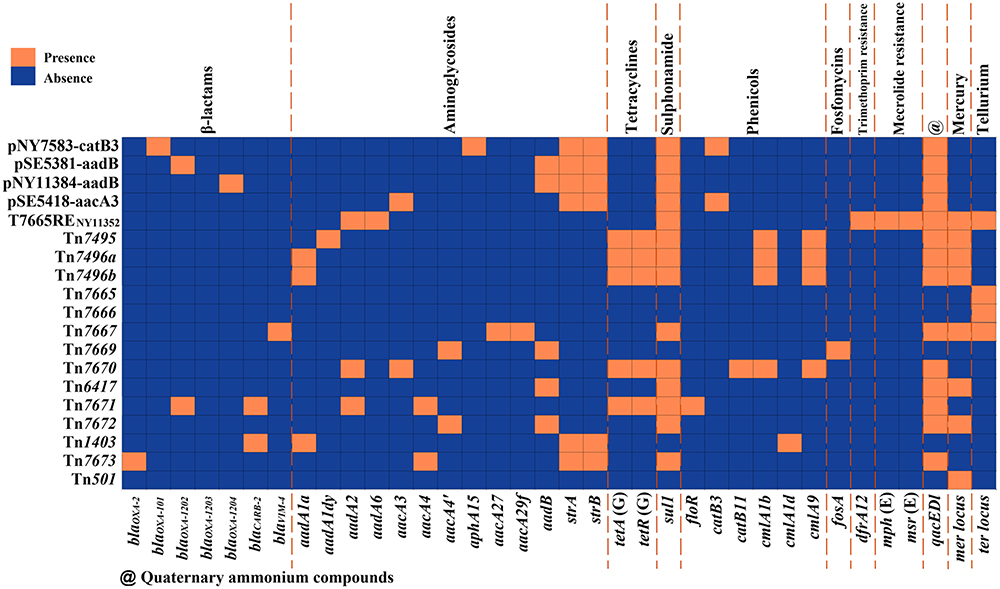 |
Figure 1 Heatmap of prevalence of resistance genes. Note: The original data are shown in Table S3. |
Novel Families of MGEs Identified in This Study
A Novel Group of IncpSE5381-aadB Plasmids
We characterized four novel plasmids: pNY7583-catB3, pSE5381-aadB, pNY11384-aadB, and pSE5418-aacA3. All had a rep (replication initiation protein) gene that could not be assigned to any known Inc groups, so were proposed as a novel Inc group, IncpSE5381-aadB. A detailed sequence comparison of these four plasmids (Figure 2) revealed a highly similar backbone structure (nucleotide identity >90%), including rep, parA/E, tivB6/8/10.
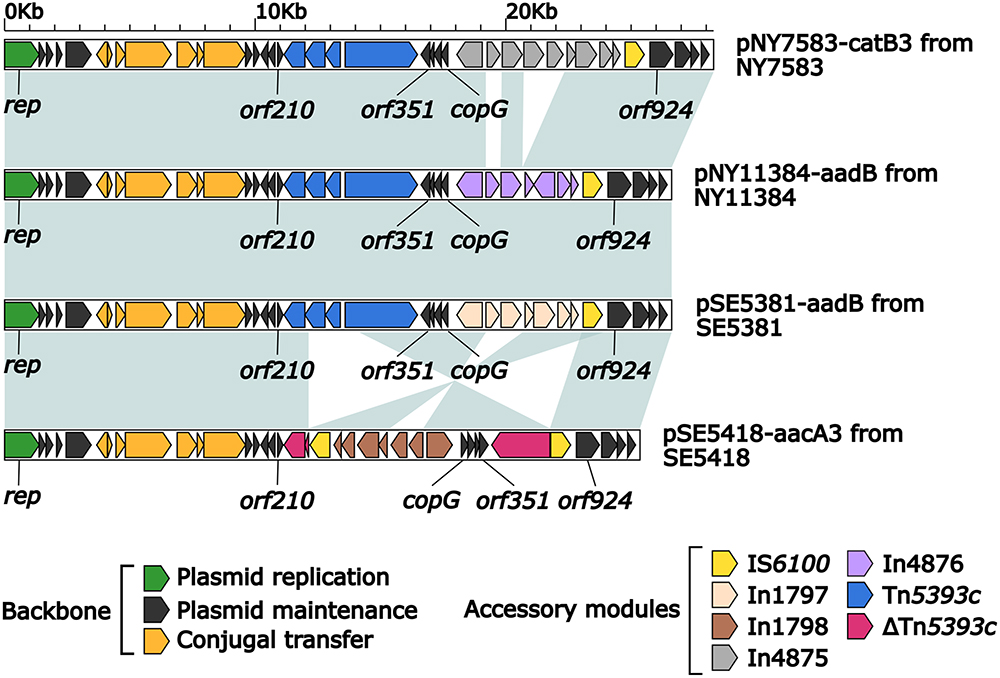 |
Figure 2 Linear comparison of the four IncpSE5381-aadB plasmids. Notes: Genes are denoted by arrows. Genes, MGEs, and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity ≥ 90%). |
Each of pNY7583-catB3, pSE5381-aadB, pNY11384-aadB, and pSE5418-aacA3 contained two accessory modules. First, each of the four plasmids acquired a novel concise class 1 integron In4875 (see below), In1797 (see below), In4876 (see below), and In1798 (see below), respectively, integrated at the same site downstream of copG. Moreover, pNY7583-catB3, pSE5381-aadB, and pNY11384-aadB harbored a complete Tn5393c at the same site downstream of orf210, while pSE5418-aacA3 had a ΔTn5393c interrupted by IS6100. There was also a major modular difference, which occurred only in pSE5418-aacA3, involving a 9.5 kb inversion likely caused by the insertion of IS6100 into Tn5393c.
Three Related Novel IMEs Tn7495, Tn7496a, and Tn7496b
Tn7495 (49.5 kb in length), Tn7496a (49.5 kb in length), and Tn7496b (51.1 kb in length) were integrated at the same site upstream of the P. aeruginosa chromosomal gene tRNASer. These three IMEs had a highly similar backbone, including core backbone markers attL, attR, int1, and int2 (Figure 3a). Tn7495 differed from Tn7496 only in the insertion of integrons: In1796 (see below) and In2094 (see below), integrated at the same site within Tn7495 and Tn7496. In comparison to Tn7496a, Tn7496b inserted an ISCfr1 at the IRt (inverted repeat at the tni end) site of integron In2094.
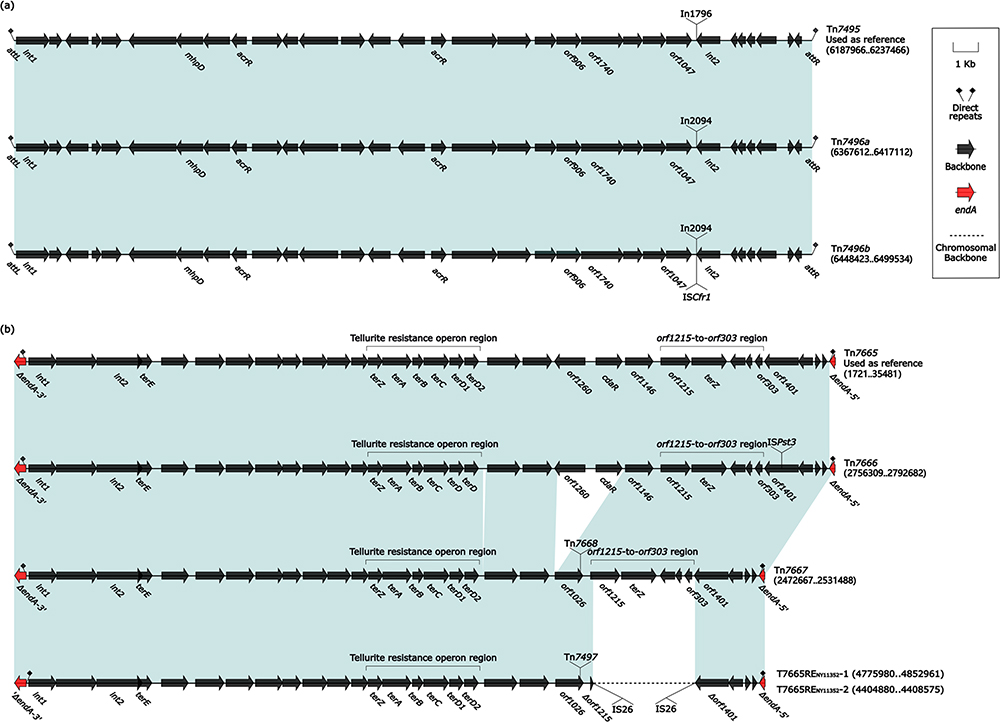 |
Figure 3 Comparison of three Tn7495 related IMEs and five Tn7665 related elements. Notes: (a) Comparison of three Tn7495-related IMEs; (b) Comparison of five Tn7665-related elements. Genes are denoted by arrows. Genes, MGEs, and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity ≥ 90%). Numbers in brackets indicate nucleotide positions within the chromosome of isolates SE5356, SE5416, NY11084, PACS171b, NY7723, NY5051, and NY11352. The accession number of Tn7665 used as reference is EU595750.35 |
Newly Defined MGEs Identified in This Study
Tn7665 and Its Derivatives Tn7666, Tn7667, T7665RENY11352-1, and T7665RENY11352-2
Tn7665(PACS171b GI),35 a prototype IME originally discovered from P. aeruginosa PACS171b, had a 33.0 kb backbone (containing int1, int2, and a tellurite resistance operon region). Tn7665, Tn7666, Tn7667, and T7665RENY11352 were integrated at the same site of the P. aeruginosa chromosomal gene endA (DNA-specific endonuclease I) and producing 2-bp direct repeats (5′-CA) (Figure 2b) Their backbones had at least two main modular differences: i) the orf1260–to–orf303 region was found only in Tn7665 and Tn7666; ii) Tn7667 and T7665RENY11352 had similar backbones; however, T7665RENY11352 was divided into T7665RENY11352-1 and T7665RENY11352-2 (found downstream of T7665RENY11352-1) and the orf1215-to-orf303 region was absent in the backbone structure of T7665RENY11352.
No accessory modules were detected in Tn7665, whereas Tn7666, Tn7667, and T7665RENY11352 exhibited distinct accessory modules (Figure 3b): i) Tn7666 acquired a single accessory module ISPst3, inserted into the site of orf1401; ii) Tn7667 and T7665RENY11352-1 acquired a Tn1403 related unit transposon Tn7668 (see below) and Tn7497 (see below), respectively, integrated into the same site of orf1206; iii) the orf1215 gene of T7665RENY11352-1 and the orf1401 gene of T7665RENY11352-2 each harbored an IS26 element. Notably, we found a Tn6582-related ICE at the upstream site of T7665RENY11352-2, which may have caused the interruption or movement of T7665RENY11352 due to its identification of the IS26 site.
Novel Transposons with Reference Prototypes Identified in This Study
Comparison of Two Related ICEs Tn6417 and Tn7671
Tn6417,36 a 108.2 kb ICE first found in P. aeruginosa DHS01,39 had an 86.0 kb backbone (containing attL/R, int, cpl, and rlx) and an insertion module, Tn6532. Tn7671 (129.6 kb in length) was identified from P. aeruginosa NY7570. Tn6417 and Tn7671 were integrated at the same site downstream of the P. aeruginosa chromosomal gene tRNAGly. There were three main modular differences in their backbones: i) Tn6417 contained its unique backbone region orf1371, smc−to−hepA region, zot–to–orf249 region, and orf378–to–orf1068 region; ii) Tn7671 harbored a unique backbone region including orf828, orf306, piL1–to–orf189 region, orf3336−to−orf1758 region, zot–to–orf219 region, and orf378–to–orf693 region; iii) orf384–to–rlx region and orf909–to–orf2514 region from Tn7671 shared <90% nucleotide identity with the corresponding regions of Tn6417 (Figure 4). Like Tn6417, Tn7671 also carried a single accessory module In4877 (see below), a novel concise class 1 integron, which was integrated into the gene ftsk, which encodes a cell division protein.
 |
Figure 4 Comparison of two related ICEs Tn6417 and Tn7671. Notes: Genes are denoted by arrows. Genes, MGEs and other features are colored based on their functional classification. Shading in light blue denotes regions of homology (nucleotide identity ≥90%), light orange (nucleotide identity ˂90%). Numbers in brackets indicate nucleotide positions within the chromosome of isolate NY7570. The accession number of Tn641739 used as reference is EU696790. |
Four Transposons Tn7669, Tn7670, Tn501, and Tn7672
Tn7669 and Tn7670 (Figure 5a and b) belong to the IS-based transposition units and underwent mobilization as a complete unit, bordered by IRL/IRR. Tn7669 is an IS222-based transposition unit, and harbors an IS222 together with captured fosA genes (fosfomycin resistance) and a novel concise class 1 integron In1800 (see below). Tn7669 is an IS6100-based transposition unit, and harbors IS6100 together with captured 18.1 kb T1403RESE5431-1 (see below) and ISCR1.
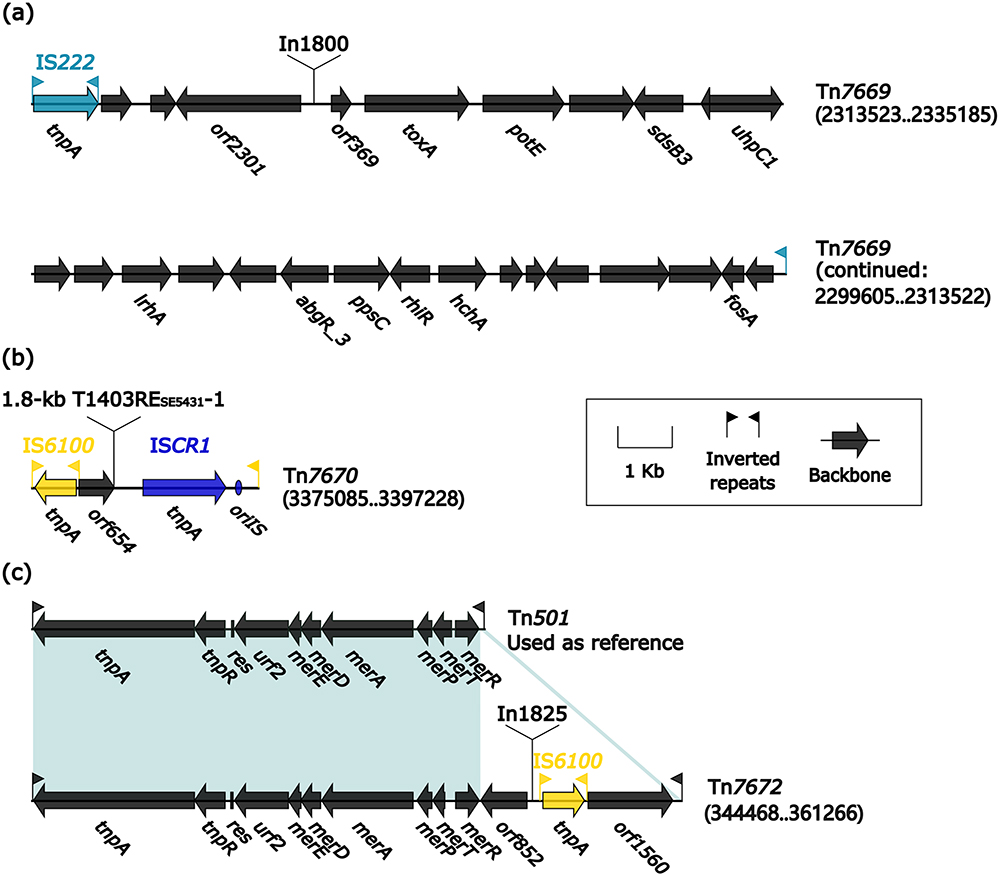 |
Figure 5 Organization of Tn7669, Tn7670, Tn501 and Tn7672. Notes: (a) Organization of Tn7669; (b) Organization of Tn7670; (c) Comparison of Tn501 and Tn7672. Genes are denoted by arrows. Genes, MGEs, and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity ≥ 90%). Numbers in brackets indicate nucleotide positions within the chromosome of isolates SE5630, SE5431, and NY4383. The accession number of Tn50140 used as reference is Z00027. |
Tn501 and Tn7672 belong to the Tn3-family unit transposons. The prototype transposon Tn501 was initially discovered in pVS1, a non-conjugative plasmid carried by P. aeruginosa PAT.37,42 Tn501 has a 12.6 kb backbone with the core backbone structure IRL–tnpA–tnpR–res–mer–IRR. Tn7672 (16.7 kb in length) has almost all Tn501 core backbone structure (Figure 5c), and showed two main modular differences: i) Tn7672 contained its unique backbone region orf825 and orf1560; ii) Tn501 carried no accessory modules, but Tn7672 acquired a novel class 1 integron In1825 (see below) and IS6100.
Tn1403 and Its Derivatives Tn7673, Tn7668, Tn7497, T1403RESE5431-1, and T1403RESE5431-2
A sequence comparison (Figure 6) was conducted on five Tn1403 derivatives, namely Tn7673, Tn7668, Tn7497, T1403RESE5431-1, and T1403RESE5431-2 (Tn7668, Tn7497, T1403RESE5431-1, and T1403RESE5431-2 were identified from the above-mentioned elements Tn7667, Tn6861, and Tn6876, respectively), together with the GenBank-derived reference sequence Tn1403. Tn140338 (19.6 kb in length), a prototypal unit transposons of the Tn3-family, initially identified in plasmid RPL11 from P. aeruginosa, harbored a core backbone structure IRL–tnpA–tnpR–res–sup–uspA–dksA–yjiK–IRR. Tn7673, Tn7668, and Tn7497 shared the same core backbone structure as Tn1403, while Tn7668 and Tn7497 contained a unique backbone region crp–to–orf693 region bracketed by 9-bp direct repeats (5′-CCCATGACG). T1403RESE5431-1, and T1403RESE5431-2 were recognized as two parts of a disrupted MGE.
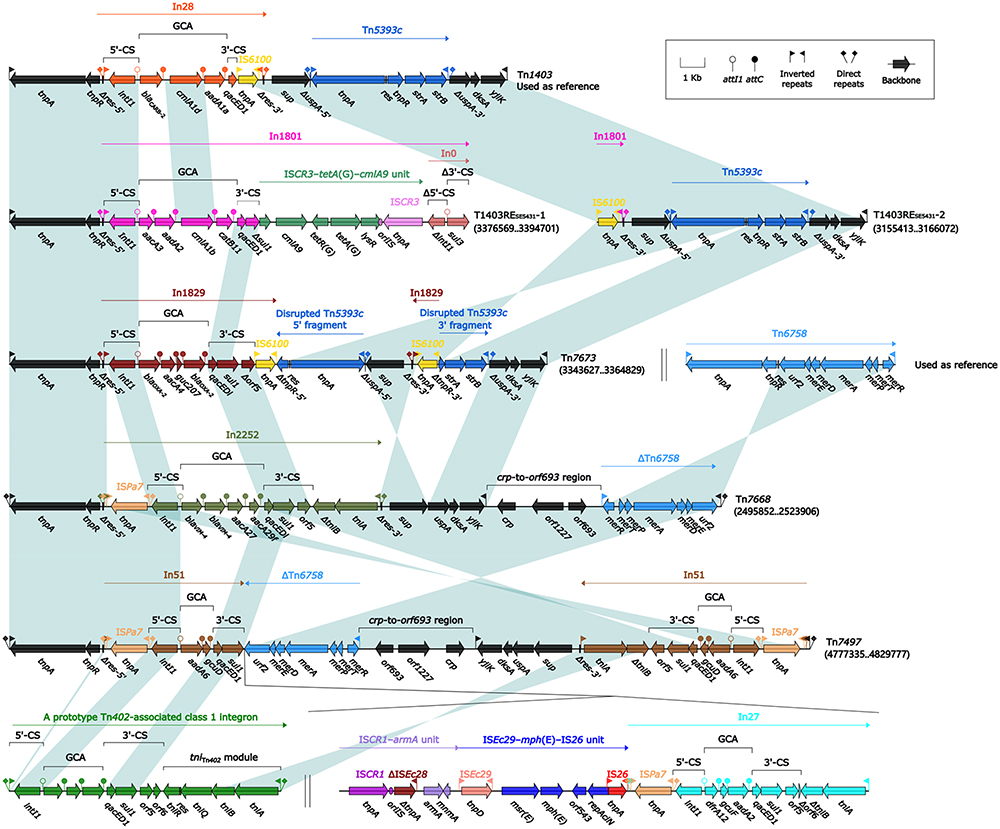 |
Figure 6 Comparison of Tn1403 and five related elements. Notes: Genes are denoted by arrows. Genes, MGEs and other features are colored based on their functional classification. Shading in light blue denotes regions of homology (nucleotide identity ≥90%). Numbers in brackets indicate nucleotide positions within the chromosome of isolates SE5431, NY3649, NY5051, and NY11352. The accession number of Tn140341 used as reference is AF313472. |
These Tn1403 derivatives’ backbones had at least three major modular differences. First, each acquired one or more integrons into the res site, instead of In28 (GCA: blaCARB-2–cmlA1d–aadA1) in Tn1403: class 1 integrons In1801 (see below), In1829 (see below), and In2252 (see below) were inserted in T1403RESE5431-1, Tn7673, and Tn7668, respectively; two In51 were integrated into Tn7497. Second, Tn1403, Tn7673, and T1403RESE5431-2 acquired a Tn5393c at the same site of backbone gene uspA; however, integron In1829 of Tn7673 inserted an IS6100, which might lead to disruption of Tn5393c and inversion of a 6.3 kb region (including IS6100, backbone region Δres-3′–sup–ΔuspA-5′ and disrupted Tn5393c-5′ fragment). Third, Tn7668 and Tn7497 carried a ∆Tn6758 (mer region from Tn6758) at the same site downstream of crp–to–orf693 region, and Tn7497 also harbored a unique 20.8 kb MDR region composed of ISCR1–armA unit, ISEc29–mph(E)–IS26 unit, and In27. Moreover, insertion of a 20.8 kb MDR region might be responsible for a 22.2 kb inversion in Tn7497 (including In51, backbone region Δres-3′–to–orf693, and ΔTn6758).
Comparison of 12 Novel Integrons
Twelve novel integrons were discovered among the 15 newly sequenced MGEs; all were concise class 1 integrons (Figure 7). Except for In1801 (GCA: aacA3–aadA2–cmlA1b–catB11), which is truncated, all remaining 11 integrons exhibited the core integron structure (including IRi/IRt, 5′-CS, GCA, 3′-CS, and tin402 module). Among them, In1800 (GCA: aadB–aacA4’–aadB) contained a complete tniABQR module, enabling self-transfer and facilitating enhanced mobility of mobile components when integrated into elements such as transposons. On the other hand, In2252 (GCA: blaVIM-4–blaVIM-4–aacA27–aacA29f) contained a truncated tniABQR module, potentially resulting in impaired its self-transferability.
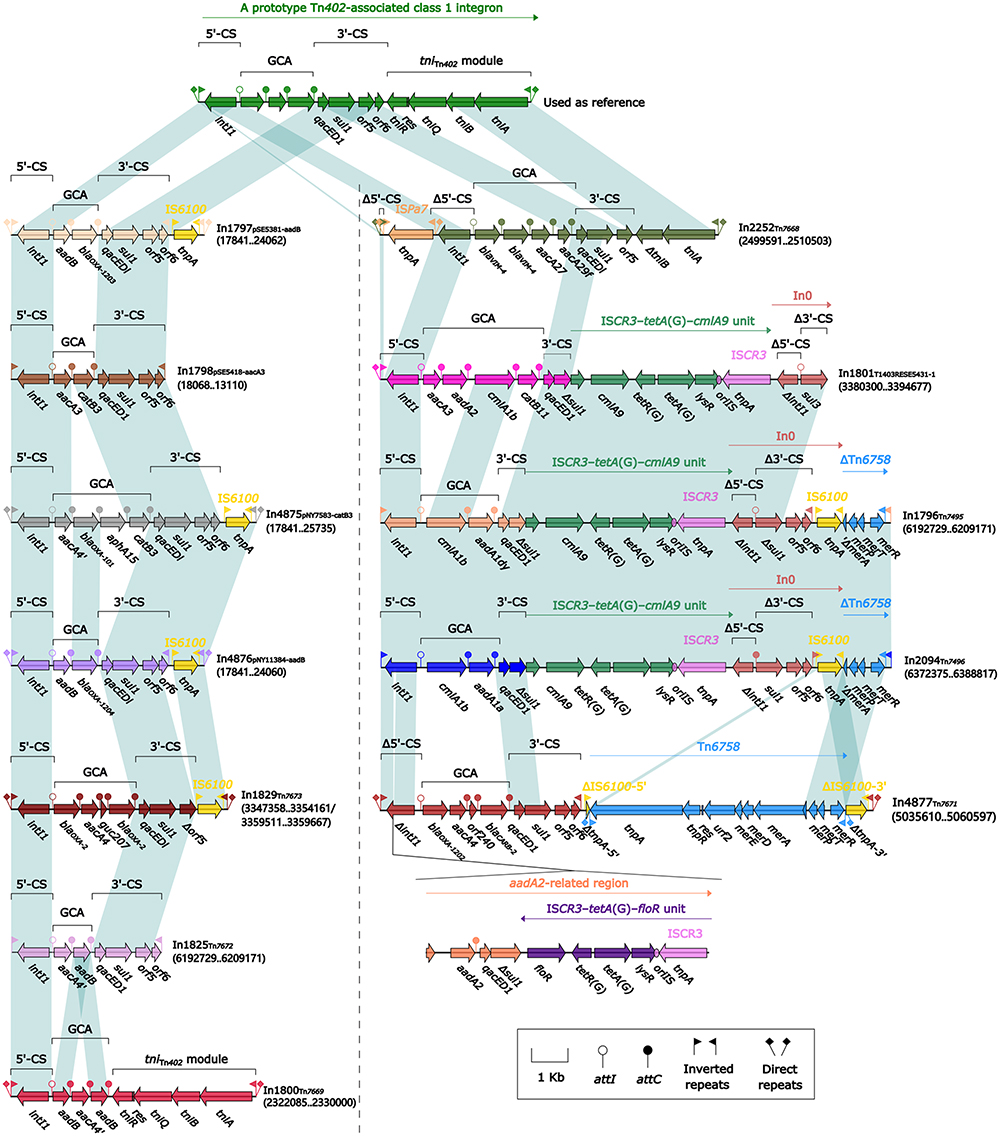 |
Figure 7 Comparison of the 12 novel integrons. Notes: Genes are denoted by arrows. Genes, mobile elements and other features are colored based on their functional classification. Shading denotes regions of homology (nucleotide identity ≥ 90%). Numbers in brackets indicate nucleotide positions within the corresponding plasmids pSE5381-aadB, pSE5418-aacA3, pNY7583-catB3, and pNY11384-aadB or within the chromosomes of isolates NY3649, NY4383, SE5430, NY5051, SE5431, SE5356, SE5416, and NY7570. |
In addition, except for In1798 (GCA: aacA3–catB3), In1825 (GCA: aacA4’–aadB), and In1800, the other nine integrons had at least one insertion module in the 5’-CS or 3’-CS or tniTn402: i) In1797 (GCA: aadB–blaOXA1203), In4875 (GCA: aacA4’–blaOXA-101), In4876 (GCA: aadB–blaOXA-1204–aphA15–catB3) and In1829 (GCA: blaOXA-2–aacA4–guc207–blaOXA-2) exhibited the insertion of one IS6100 at the site downstream of the 3′-CS region; ii) In2252 was inserted by a separate module ISPa7 in the 5’-CS region; iii) In1801 (GCA: aacA3–aadA2–cmlA1b–catB11) had In0 and ISCR3–tetA(G)–cmlA9 unit; (iv) In1796 (GCA: cmlA1b–aadA1dy) and In2094 (GCA: cmlA1b–aadA1a) harbored In0, ISCR3-tetA(G)-cmLA9 unit, and ΔTn6758; (v) In4877 (GCA: blaOXA-1202–aacA4–orf240–blaCARB-2) acquired an aadA2-related region at the site of ΔintI1 and an IS6100 at the site downstream of orf6, which was interrupted by a complete Tn6758. In particular, blaOXA-101 and its three novel variants blaOXA-1202/1203/1204 were identified within the GCA of integrons In4875, In4877, In1797, and In4876, respectively.
Summary of Newly Identified or Designated MGEs
We identified 23 new MGEs, including four IMEs Tn7495, Tn7496, Tn7666, and Tn7667; one ICE Tn7671; four unit transposons Tn7668, Tn7497, Tn7672, and Tn7673; two IS-based transposition units Tn7669 and Tn7670; and 12 integrons In1796, In2094, In2252, In4877, In1825, In1800, In1801, In1829, In4875, In1797, In4876, and In1798. One novel Inc group IncpSE5381-aadB was designated. Additionally, the element PACS171b GI previously designated with determined sequences was renamed as standard Tn designations Tn7665.
Three Novel Variants of Antibiotic Resistance Genes and Their Characteristics
Three novel blaOXA-101 variants encoding OXA-β-lactamase, namely blaOXA-1202, blaOXA-1203, and blaOXA-1204 (Figure S1), were identified from three distinct clinical isolates: P. aeruginosa NY7570, SE5381, and NY11384, respectively. These three novel variants, all of which were categorized within the OXA-10-like OXA enzyme, featured genes that were 801 bp in length and encoded a 266 amino acid-long protein. The deduced OXA-1202 differs from OXA-101 by a single amino acid substitution Val188Ala; OXA-1203 differed from OXA-101 by one amino acid substitution Ala66Val; OXA-1204 differed from the OXA-101 by one amino acid substitution Trp154Gly.
OXA-1202/1203/1203 gene fragments, their point mutants OXA-101, and the prototype OXA variant OXA-10 (as a positive control) were cloned into pUC57K (a kanamycin-resistant plasmid) and transformed into E. coli TOP10 to obtain E. coli electroporants TOP10/pUC57K-OXA-1202/1203/1204/101/10. Classic broth microdilution was applied to test antimicrobial susceptibility of these electroporants and their negative control strains (TOP10, TOP10/pUC57K, and ATCC 25,922). Strains encoding OXA-1202/1203/1204 enzymes were highly resistant to amoxicillin and ampicillin relative to the negative controls, but were still susceptible to piperacillin (Table 2). Furthermore, TOP10/pUC57K-OXA-1204 was also resistant to ceftazidime and cefoxitin, while TOP10/pUC57K-OXA-1202/1203 was susceptible to cephalosporins and exhibited narrow-spectrum antibiotic resistance similar to OXA-101.
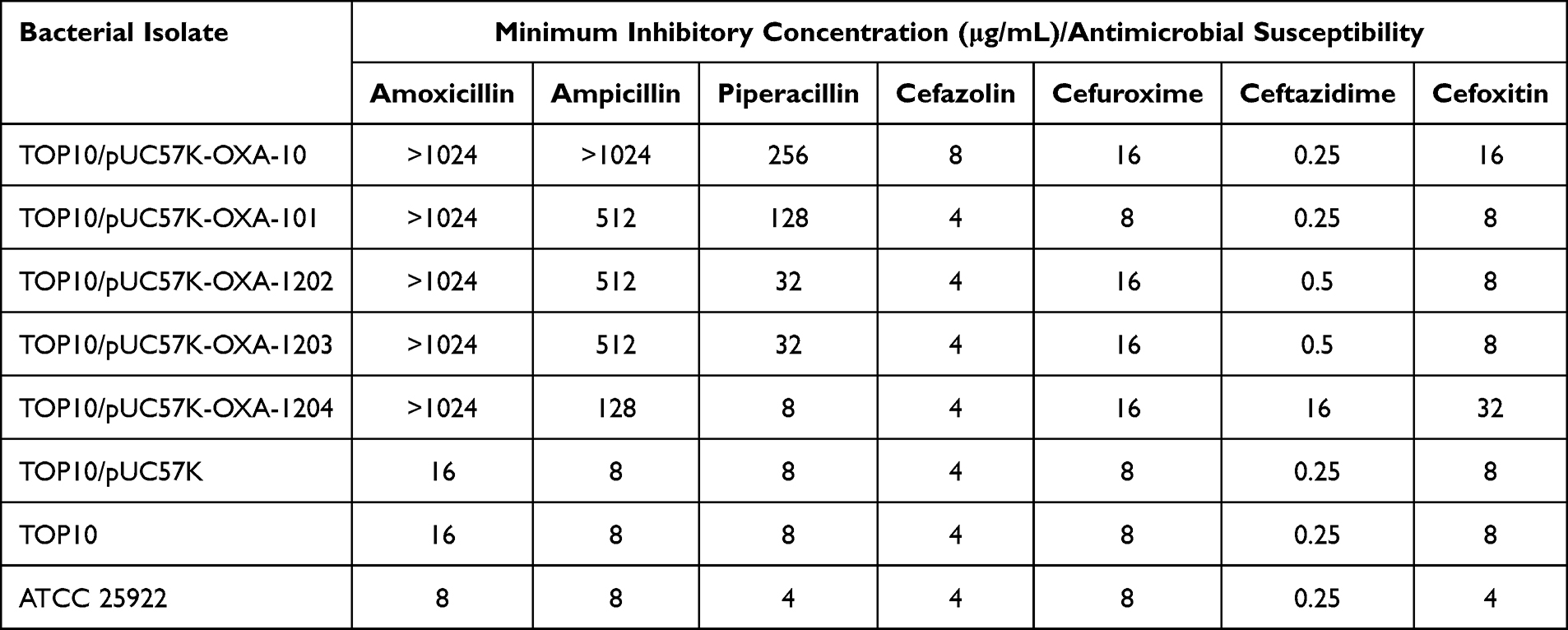 |
Table 2 MICs of Selected Antimicrobial Agents for the Transformants Carrying blaOXA |
Discussion
Fifteen multiple drug resistance (MDR) isolates were isolated from clinical samples and completely sequenced in this study. Among these isolates, ST235 and ST244 are the more common high-risk clones. All ST335 isolates harbored exoU+/exoS- virulotypes, while all ST244 isolates harbored exoU-/exoS+ virulotypes. This finding indicates a potential correlation between the high-risk clones and T3SS virulotypes, consistent with previous work.43,44 In addition, T3SS virulence genes were detected in all 15 MDR isolates collected. Previous studies have shown a correlation between the presence of virulence genes and bacterial resistance, and antibiotic resistance genes has been easily detected in T3SS-positive isolates.45,46 which might jointly contribute to the high prevalence of P. aeruginosa.
A total of 15 novel MGEs were identified through comprehensive genome alignment. In particular, the IncpSE5381-aadB plasmids and Tn7495-related IMEs were identified and sequenced for the first time. By blasting the rep sequence and the backbone sequence among GenBank (identity≥95%, last accessed 20 Feb, 2023), we found that IncpSE5381-aadB plasmid was only harbored in P. aeruginosa. To verify its transfer ability, we performed multiple conjugation experiments, but did not succeed in transferring IncpSE5381-aadB plasmid from their wild isolates to PAO1 or ATCC 27,853, possibly due to incomplete functionality of the plasmid-encoded conjugative transfer protein. Tn7495 family IMEs presented a conserved backbone with the same chromosomal insertion site. Similar to conventional IMEs, Tn7495 family IMEs do not undergo intercellular transfer autonomously. BLAST searches of the integrase sequence of Tn7495-related IMEs among GenBank (identity≥95%, with relatively complete element backbone, last accessed 20 Feb, 2023) indicated that as of yet these IMEs have only been identified in P. aeruginosa. Although the newly discovered elements in this study had only been identified in P. aeruginosa, they might also be widely spread to other species in the process of genetic evolution, and this unknown situation deserved our close attention.
As prototypes or references, Tn7665, Tn6417, Tn501, and Tn1403 were all initially found in P. aeruginosa. Tn7665-related IMEs had been frequently reported in P. aeruginosa and there are documented instances of insertions of this group of elements within the chromosomal gene endA,47 consistent with the findings of this study. Tn6417-related ICEs had been identified not only in P. aeruginosa but also occasionally among other species, such as Klebsiella pneumoniae, Morganella morganii, Bordetella, and Achromobacter xylosoxidans. The presence of Tn501-related elements and Tn1403-related elements in P. aeruginosa has been reported multiple times.17,48,49 Overall, the types of MGEs identified in this study exhibit a high prevalence among clinical P. aeruginosa isolates, revealing that they may play a critical role in facilitating the spread of resistance in P. aeruginosa.
In our study, a total of 36 ARGs were detected from the 19 MGEs analyzed, involving 11 classes of antibiotics. Among them, aminoglycoside resistance genes (n=13) and β-lactam resistance genes (n=7) accounted for the largest proportion. Thirty of these ARGs were found in the gene cassette of class 1 integron; the remaining six belonged to other sub-regions: genes strA and strB in the Tn5393c; gene fosA in the Tn7669 backbone; genes mph(E) and msr(E) in the ISEc29–mph(E)–IS26 unit; gene ter in the Tn7495-related IMEs backbone (Table S3). In addition, we identified three novel β-lactamase genes (blaOXA-1202/1203/1204) located within integrons, showing that integron activity not only aided the spread of ARGs but may also expedite the evolution of antibiotic resistance, as found previously.50 During the transfer of these MGEs, some mutations or the occurrence of the homologous recombinant events accelerated the evolution of the MGEs and promoted the production of novel ARGs. These ARGs-carrying MGEs formed intricate mosaic structures and exhibited intercellular or intracellular mobility, facilitating the widespread dissemination of bacterial resistance.
OXA enzymes are a large family of enzymes with more over a thousand variants that are widely distributed. Three newly identified blaOXA variants (blaOXA-1202/1203/1204) in P. aeruginosa belonged to the OXA-10-like subfamily like their point mutant blaOXA-101. The presence of OXA-10-like β-lactamases has been repeatedly observed in P. aeruginosa,7,51 and has also been occasionally reported in Acinetobacter baumannii52 and Enterobacteriaceae.53 According to antibiotic susceptibility testing, OXA-1202, OXA-1203 presented a similar resistance phenotype as their point mutant OXA-101. Notably, OXA-1204 showed reduced hydrolytic activity against some antibiotics tested (eg ampicillin and piperacillin) compared with OXA-101 and exhibited exclusive resistance to ceftazidime and cefoxitin. The substitution of W154G in OXA-1204 might play a role in increasing resistance against certain cephalosporins in P. aeruginosa. Previous studies have shown that lysine carbamylation had important mechanistic functions in the catalytic activity of OXA-10 from P. aeruginosa.54,55 The carbamyl group of Lys-70 in OXA-10 can be stabilized by interacting with Ser-67, Trp-154 and a crystal water molecule.54 Therefore, the mutation of amino acid 154 in OXA-1204 could potentially influence the carbamylation of Lys-70 and result in a reduction of its hydrolytic activity towards ampicillin and other antibiotics (Figure S2). The diversity of the OXA family continues to expand. While certain inhibitors might impede the activity of these genes’ products,56 the presence of extended-spectrum class D β-lactamases and carbapenem-hydrolysing class D β-lactamases continues to pose a formidable challenge to clinical anti-infective therapy, necessitating utmost attention.
Conclusion
A detailed sequence comparison of a total of 19 novel MGEs from seven types of P. aeruginosa revealed that they carried different combinations of resistance genes or entirely novel resistance gene variants. This study reported the IncpSE5381-aadB plasmids and Tn7495-related IMEs for the first time, and also identified the novel resistance genes blaOXA-1202/1203/1204 carried by MGEs. These MGEs underwent continuous modification during transmission, leading to the evolution of intricate mosaic structures through transposition or homologous recombination. In turn, this facilitated the spread of antibiotic resistance and enhanced the adaptability of P. aeruginosa under the selective pressure of various antibiotics. The present study has elucidated the genetic evolutionary characteristics of novel ARGs-carrying MGEs, thereby enhancing the diversity of transmission modes of antibiotic resistance in P. aeruginosa and providing a deeper understanding of the drug resistance mechanisms of clinical P. aeruginosa. In addition, some of the elements identified in this study were only found in P. aeruginosa, but these elements are constantly evolving during transmission and may be more widely spread to other species, which deserves our continuous attention in future studies.
Ethics Approval and Informed Consent
This study uses the clinical bacterial isolates obtained from the Chinese public hospitals as listed in Table S1. The local legislation did not require the study to be reviewed or approved by an ethics committee, because the bacterial isolates involved in this study was part of the routine laboratory procedures. The research involving biohazards and all related procedures were approved by the Biosafety Committee of the Beijing Institute of Microbiology and Epidemiology.
Acknowledgments
All experiments and data analyses were done in Dr. Dongsheng Zhou’s laboratory.
Funding
This work was supported by the National Key Research and Development Program of China (Grant no: 2022YFC2303900), the Key Project of Medical Science and Technology Research Program of Henan Province (Grant no: SBGJ202302089), and the Research Project from the State Key Laboratory of Pathogen and Biosecurity (Grant no SKLPBS2214).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lynch JP, Zhanel GG. Pseudomonas aeruginosa pneumonia: evolution of antimicrobial resistance and implications for therapy. Semin Resp Crit Care Med. 2022;43(2):191–218. doi:10.1055/s-0041-1740109
2. De Oliveira DMP, Forde BM, Kidd TJ, et al. Antimicrobial Resistance in ESKAPE Pathogens. Clin Microbiol Rev. 2020;33(3). doi:10.1128/CMR.00181-19
3. Glen KA, Lamont IL. β-lactam resistance in pseudomonas aeruginosa: current status, future prospects. Pathogens. 2021;10(12):1638. doi:10.3390/pathogens10121638
4. Ambler RP. The structure of beta-lactamases. Philos Trans R Soc London, Ser B. 1980;289(1036):321–331.
5. Fisher JF, Meroueh SO, Mobashery S. Bacterial resistance to beta-lactam antibiotics: compelling opportunism, compelling opportunity. Chem Rev. 2005;105(2):395–424. doi:10.1021/cr030102i
6. Bush K. Proliferation and significance of clinically relevant β-lactamases. Ann NY Acad Sci. 2013;1277:84–90. doi:10.1111/nyas.12023
7. Matthew M, Sykes RB. Properties of the beta-lactamase specified by the Pseudomonas plasmid RPL11. J Bacteriol. 1977;132(1):341–345. doi:10.1128/jb.132.1.341-345.1977
8. Antunes NT, Lamoureaux TL, Toth M, Stewart NK, Frase H, Vakulenko SB. Class D β-lactamases: are they all carbapenemases? Antimicrob Agents Chemother. 2014;58(4):2119–2125. doi:10.1128/AAC.02522-13
9. Yoon EJ, Jeong SH. Class D β-lactamases. J Antimicrob Chemother. 2021;76(4):836–864. doi:10.1093/jac/dkaa513
10. Haudiquet M, de Sousa JM, Touchon M, Rocha EPC. Selfish, promiscuous and sometimes useful: how mobile genetic elements drive horizontal gene transfer in microbial populations. Philos Trans R Soc London, Ser B. 2022;377(1861):20210234. doi:10.1098/rstb.2021.0234
11. Botelho J, Roberts AP, León-Sampedro R, Grosso F, Peixe L. Carbapenemases on the move: it’s good to be on ICEs. Mobile DNA. 2018;9:37. doi:10.1186/s13100-018-0141-4
12. Bonnin RA, Bogaerts P, Girlich D, et al. Molecular Characterization of OXA-198 carbapenemase-producing pseudomonas aeruginosa clinical isolates. Antimicrob Agents Chemother. 2018;62(6). doi:10.1128/AAC.02496-17
13. Smillie C, Garcillán-Barcia MP, Francia MV, Rocha EP, de la Cruz F. Mobility of plasmids. Microbiol Mol Biol Rev. 2010;74(3):434–452. doi:10.1128/MMBR.00020-10
14. Guédon G, Libante V, Coluzzi C, Payot S, Leblond-Bourget N. The obscure world of integrative and mobilizable elements, highly widespread elements that pirate bacterial conjugative systems. Genes. 2017;8(11). doi:10.3390/genes8110337
15. Johnson CM, Grossman AD. Integrative and conjugative elements (ICEs): what they do and how they work. Ann Rev Genet. 2015;49:577–601. doi:10.1146/annurev-genet-112414-055018
16. Mu X, Li X, Yin Z, et al. Abundant diversity of accessory genetic elements and associated antimicrobial resistance genes in pseudomonas aeruginosa isolates from a single Chinese hospital. Ann Clinic Microbiol Antimicrob. 2023;22(1):51. doi:10.1186/s12941-023-00600-3
17. Yu T, Yang H, Li J, et al. Novel chromosome-borne accessory genetic elements carrying multiple antibiotic resistance genes in pseudomonas aeruginosa. Front Cell Infect Microbiol. 2021;11:638087. doi:10.3389/fcimb.2021.638087
18. Chen F, Wang P, Yin Z, et al. VIM-encoding Inc(pSTY) plasmids and chromosome-borne integrative and mobilizable elements (IMEs) and integrative and conjugative elements (ICEs) in Pseudomonas. Ann Clinic Microbiol Antimicrob. 2022;21(1):10. doi:10.1186/s12941-022-00502-w
19. Li X, Mu X, Chen F, et al. Characterization of Three Novel IMP Metallo-β-Lactamases, IMP-89, IMP-91, and IMP-96, and Diverse bla(IMP)-carrying accessory genetic elements from Chinese clinical isolates. Microbiol Spect. 2023;11(3):1.
20. Richter M, Rosselló-Móra R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci USA. 2009;106(45):19126–19131. doi:10.1073/pnas.0906412106
21. CLSI supplement M100Clinical and Laboratory Stantardeds Institute. CLSI: Performance Standards for Antimicrobial Susceptibility Testing.
22. Hackl T, Hedrich R, Schultz J, Förster F. proovread: large-scale high-accuracy PacBio correction through iterative short read consensus. Bioinformatics. 2014;30(21):3004–3011. doi:10.1093/bioinformatics/btu392
23. Li C, Jiang X, Yang T, et al. Genomic epidemiology of carbapenemase-producing Klebsiella Pneumoniae in China. Genomics Proteomics Bioinf. 2022;20:1154–1167. doi:10.1016/j.gpb.2022.1002.1005
24. Fu J, Zhang J, Yang L, et al. Precision methylome and in vivo methylation kinetics characterization of Klebsiella Pneumoniae. Genomics Proteomics Bioinf. 2022;20(2):418–434. doi:10.1016/j.gpb.2021.04.002
25. Brettin T, Davis JJ, Disz T, et al. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci Rep. 2015;5:8365. doi:10.1038/srep08365
26. Boratyn GM, Camacho C, Cooper PS, et al. BLAST: a more efficient report with usability improvements. Nucleic Acids Res. 2013;41:W29–33. doi:10.1093/nar/gkt282
27. O’Leary NA, Wright MW, Brister JR, et al. Reference sequence (RefSeq) database at NCBI: current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016;44(D1):D733–45. doi:10.1093/nar/gkv1189
28. Boutet E, Lieberherr D, Tognolli M, et al. UniProtKB/Swiss-Prot, the manually annotated section of the UniProt knowledgebase: how to use the entry view. Meth Mol Bio. 2016;1374:23–54.
29. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
30. Roberts AP, Chandler M, Courvalin P, et al. Revised nomenclature for transposable genetic elements. Plasmid. 2008;60(3):167–173. doi:10.1016/j.plasmid.2008.08.001
31. Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34:D32–D36. doi:10.1093/nar/gkj014
32. Moura A, Soares M, Pereira C, Leitão N, Henriques I, Correia A. INTEGRALL: a database and search engine for integrons, integrases and gene cassettes. Bioinformatics. 2009;25(8):1096–1098. doi:10.1093/bioinformatics/btp105
33. Jia B, Raphenya AR, Alcock B, et al. CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017;45(D1):D566–D573. doi:10.1093/nar/gkw1004
34. Wang P, Jiang X, Mu K, et al. DANMEL: a manually curated reference database for analyzing mobile genetic elements associated with bacterial drug resistance. mLife. 2022;1:460–464. doi:10.1002/mlf2.12046
35. Hayden HS, Gillett W, Saenphimmachak C, et al. Large-insert genome analysis technology detects structural variation in Pseudomonas aeruginosa clinical strains from cystic fibrosis patients. Genomics. 2008;91(6):530–537. doi:10.1016/j.ygeno.2008.02.005
36. Zeng L, Zhan Z, Hu L, et al. Genetic Characterization of a bla (VIM-24)-Carrying IncP-7β Plasmid p1160-VIM and a bla (VIM-4)-harboring integrative and conjugative element Tn6413 from clinical pseudomonas aeruginosa. Front Microbiol. 2019;10:213. doi:10.3389/fmicb.2019.00213
37. Stanisich VA. Interaction between an R factor and an element conferring resistance to mercuric ions in Pseudomonas aeruginosa. Mol Gen Genet. 1974;128(3):201–212. doi:10.1007/BF00267109
38. Levesque RC, Jacoby GA. Molecular structure and interrelationships of multiresistance beta-lactamase transposons. Plasmid. 1988;19(1):21–29. doi:10.1016/0147-619X(88)90059-5
39. Valot B, Rohmer L, Jacobs MA, Miller SI, Bertrand X, Hocquet D. Comparative genomic analysis of two multidrug-resistant clinical Isolates of ST395 epidemic strain of pseudomonas aeruginosa obtained 12 years apart. Genome Announcem. 2014;2(3). doi:10.1128/genomeA.00515-14
40. Diver WP, Grinsted J, Fritzinger DC, et al. DNA sequences of and complementation by the tnpR genes of Tn21, Tn501 and Tn1721. Mol Gen Genet. 1983;191(2):189–193. doi:10.1007/BF00334812
41. Stokes HW, Elbourne LD, Hall RM. Tn1403, a multiple-antibiotic resistance transposon made up of three distinct transposons. Antimicrob Agents Chemother. 2007;51(5):1827–1829. doi:10.1128/AAC.01279-06
42. Stanisich VA, Bennett PM, Richmond MH. Characterization of a translocation unit encoding resistance to mercuric ions that occurs on a nonconjugative plasmid in Pseudomonas aeruginosa. J Bacteriol. 1977;129(3):1227–1233. doi:10.1128/jb.129.3.1227-1233.1977
43. Peña C, Cabot G, Gómez-Zorrilla S, et al. Influence of virulence genotype and resistance profile in the mortality of Pseudomonas aeruginosa bloodstream infections. Clin Infect Dis. 2015;60(4):539–548. doi:10.1093/cid/ciu866
44. Weber C, Schultze T, Göttig S, et al. Antimicrobial activity of ceftolozane-tazobactam, ceftazidime-avibactam, and cefiderocol against multidrug-resistant pseudomonas aeruginosa recovered at a German University Hospital. Microbiol Spectr. 2022;10(5):e0169722. doi:10.1128/spectrum.01697-22
45. Jabalameli F, Mirsalehian A, Khoramian B, et al. Evaluation of biofilm production and characterization of genes encoding type III secretion system among Pseudomonas aeruginosa isolated from burn patients. Burns. 2012;38(8):1192–1197. doi:10.1016/j.burns.2012.07.030
46. Sawa T, Shimizu M, Moriyama K, Wiener-Kronish JP. Association between Pseudomonas aeruginosa type III secretion, antibiotic resistance, and clinical outcome: a review. Crit Care. 2014;18(6):668. doi:10.1186/s13054-014-0668-9
47. Di Pilato V, Pollini S, Rossolini GM. Tn6249, a new Tn6162 transposon derivative carrying a double-integron platform and involved with acquisition of the blaVIM-1 metallo-β-lactamase gene in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2015;59(3):1583–1587. doi:10.1128/AAC.04047-14
48. Vézina G, Levesque RC. Molecular characterization of the class II multiresistance transposable element Tn1403 from Pseudomonas aeruginosa. Antimicrob Agents Chemother. 1991;35(2):313–321. doi:10.1128/AAC.35.2.313
49. Perez F, Hujer AM, Marshall SH, et al. Extensively drug-resistant pseudomonas aeruginosa isolates containing blaVIM-2 and elements of Salmonella genomic island 2: a new genetic resistance determinant in Northeast Ohio. Antimicrob Agents Chemother. 2014;58(10):5929–5935. doi:10.1128/AAC.02372-14
50. Souque C, Escudero JA, MacLean RC. Integron activity accelerates the evolution of antibiotic resistance. eLife. 2021;2021:10.
51. Huovinen P, Huovinen S, Jacoby GA. Sequence of PSE-2 beta-lactamase. Antimicrob Agents Chemother. 1988;32(1):134–136. doi:10.1128/AAC.32.1.134
52. Giannouli M, Tomasone F, Agodi A, et al. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii strains in intensive care units of multiple Mediterranean hospitals. J Antimicrob Chemother. 2009;63(4):828–830. doi:10.1093/jac/dkp032
53. Porto A, Ayala J, Gutkind G, Di Conza J. A novel OXA-10-like beta-lactamase is present in different Enterobacteriaceae. Diag Microbiol Infect Dis. 2010;66(2):228–229. doi:10.1016/j.diagmicrobio.2009.09.010
54. Maveyraud L, Golemi D, Kotra LP, et al. Insights into class D beta-lactamases are revealed by the crystal structure of the OXA10 enzyme from Pseudomonas aeruginosa. Structure. 2000;8(12):1289–1298. doi:10.1016/S0969-2126(00)00534-7
55. Li J, Cross JB, Vreven T, Meroueh SO, Mobashery S, Schlegel HB. Lysine carboxylation in proteins: OXA-10 beta-lactamase. Proteins. 2005;61(2):246–257. doi:10.1002/prot.20596
56. Nishimura T, Kawahara T, Kagaya N, et al. JBIR-155, a Specific Class D β-lactamase inhibitor of microbial origin. Organic Letters. 2021;23(11):4415–4419. doi:10.1021/acs.orglett.1c01352
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.