Back to Journals » Neuropsychiatric Disease and Treatment » Volume 11
Genetic association between G protein-coupled receptor kinase 6/β-arrestin 2 and dopamine supersensitivity psychosis in schizophrenia
Authors Oda Y, Kanahara N ![]() , Kimura H, Watanabe H
, Kimura H, Watanabe H ![]() , Hashimoto K
, Hashimoto K ![]() , Iyo M
, Iyo M ![]()
Received 3 April 2015
Accepted for publication 8 June 2015
Published 29 July 2015 Volume 2015:11 Pages 1845—1851
DOI https://doi.org/10.2147/NDT.S86042
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Wai Kwong Tang
Yasunori Oda,1 Nobuhisa Kanahara,2 Hiroshi Kimura,1 Hiroyuki Watanabe,2 Kenji Hashimoto,3 Masaomi Iyo1
1Department of Psychiatry, Chiba University Graduate School of Medicine, Chiba, Japan; 2Division of Medical Treatment and Rehabilitation, 3Division of Clinical Neuroscience, Chiba University Center for Forensic Mental Health, Chiba, Japan
Background/aim: Dopamine supersensitivity psychosis (DSP), clinically characterized by unstable and severe psychosis or tardive dyskinesia and often categorized as treatment-resistant schizophrenia, is promoted by long-term antipsychotic treatment. An upregulation of the dopamine D2 receptor caused by antipsychotic(s) is involved in the development of DSP. The present study explored the potential roles of G protein-coupled receptor kinase 6 (GRK6) and β-arrestin 2 (ARRB2) that are involved in the trafficking of DRD2 in patients with DSP.
Methods: We conducted a genetic association study of GRK6/ARRB2 between the patients with DSP episodes [DSP(+) group: N=108] and the patients without DSP(-) episodes [DSP(-) group: N=169] from the total group of patients (N=333). Based on the patients’ treatment history, a DSP episode was defined as withdrawal psychosis, developed tolerance to antipsychotic effect, and tardive dyskinesia (the remaining 56 patients were excluded due to insufficient information).
Results: The results revealed that none of the allelic or genotyping distributions of five single nucleotide polymorphisms (SNPs) of GRK6 and three SNPs of ARRB2 showed any significant difference between the DSP(+) and DSP(-) groups.
Conclusion: The results suggest that the SNP analyses of these two molecules fail to classify patients into the potential clinical subtype of DSP(+) or DSP(-) group. However, since GRK6 and ARRB2 are surely involved in dopamine D2 receptor metabolism, further studies based on prospective observations of the onset of DSP under specific antipsychotic treatments are needed.
Keywords: antipsychotic, dopamine D2 receptor, tardive dyskinesia, treatment-resistant
Introduction
Schizophrenia, a severe and chronic mental illness that occurs in approximately 1% of the human population, profoundly reduces the quality of life of the affected individuals. Despite the extensive basic and clinical research concerning schizophrenia over the past three decades, the disease pathogenesis remains incompletely understood. In clinical practice, most of the available antipsychotic agents are effective in treating schizophrenia, particularly for positive symptoms, which targets mainly the dopamine D2 receptor (DRD2), although clozapine is the exceptional agent.1 However, some patients with schizophrenia respond less or not at all to these agents regardless of the appropriate pharmacotherapy; this is known as treatment-resistant schizophrenia (TRS).2,3
Drug-induced dopamine supersensitivity psychosis (DSP) has been speculated to be involved in the process of the development of TRS. DSP is characterized by an acute exacerbation of symptoms upon the discontinuation of antipsychotics or even after minor stress (withdrawal psychosis), and by tardive dyskinesia (TD), a typical neurological sign of DSP.4,5 A development of tolerance to antipsychotic therapeutic effects, such that even high doses of antipsychotics no longer control symptoms, is also included in the concept of DSP. It was estimated that approximately 50% of the cases of TRS are due to DSP, based on clinical trials showing relatively high occurrence rates of TD in patients with TRS.6 A number of basic research and neuroimaging studies have demonstrated that a prolonged blockade of DRD2 by antipsychotics leads to a compensatory increase in DRD2 densities, and this phenomenon is presumed to be closely related to the development of dopamine supersensitivity.7–11 We recently reported that patients experiencing DSP episode(s) are a relatively typical subgroup in TRS and that they might differ in symptomatology and clinical course from patients who have never experienced such episodes.12 The elucidation of the biological mechanism underlying the upregulation of DRD2 caused by antipsychotics could thus contribute to an effective treatment strategy for patients with drug-induced DSP and TRS.
DRD2 desensitization is regulated by G protein-coupled receptor kinase 6 (GRK6) and beta-arrestin 2 (ARRB2). Following the activation of DRD2 by dopamine, GRK6 phosphorylates the receptor, and then ARRB2 binds to the phosphorylated receptors and prevents further G-protein activation. After this process, DRD2 is internalized from the cell membrane. Finally, the internalized receptors recycle back to the cell surface or degrade.13 Therefore, GRK6 and ARRB2 have a potential impact on the DRD2 upregulation by antipsychotics, and it is suspected that patients who tend to achieve DRD2 upregulation by antipsychotics are more likely to experience DSP episodes (including withdrawal psychosis and TD) compared with patients who do not. These two molecules have already been reported to be potentially related to schizophrenia,14–16 but in relation to DSP, the potential involvement of ARRB2 in TD is reported in only one study by Liou et al.15 Here, we conducted a genetic association study to investigate the involvement of GRK6 and ARRB2 in the occurrence of DSP episode(s) in patients with schizophrenia. To that end, the present study compared the single nucleotide polymorphism (SNP) distribution of these molecules between the patients with DSP episodes and those without such episodes.
Materials and methods
Subjects
This is a case-control study, and all of the study participants were Japanese. As shown in Table 1, the subjects included 333 patients with schizophrenia:164 males and 169 females, age 46.0±14.7 years (mean ± standard deviation: male 45.2±14.4 years old, female 46.8±15.1 years old, P>0.05), age range 20–81 years, who met the DSM-IV-TR17 criteria for schizophrenia (SZ) and who were outpatients or inpatients at Chiba University Hospital or affiliated hospitals. There were 330 healthy controls; they were medical staff in our hospital and were recruited for several previous research projects by us: 193 males and 137 females, age 37.3±15.7 years, age range 20–85 years. The lack of a personal or family history of psychotic disorders in the controls was confirmed through careful interviews by experienced psychiatrists for each participant. There were significant differences in age and sex between the SZ and control groups.
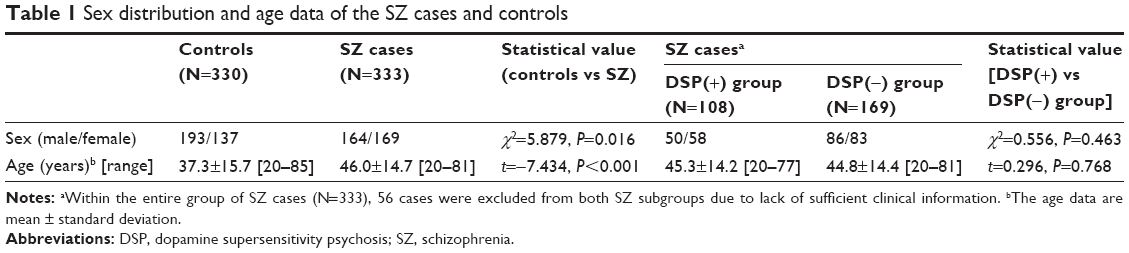 | Table 1 Sex distribution and age data of the SZ cases and controls |
This study was performed after approval was obtained from the Ethics Committee of the Chiba University Graduate School of Medicine, and was conducted in accord with the Helsinki Declaration. All subjects provided written informed consent for the use of their DNA samples as part of this study.
Assessment of dopamine supersensitivity psychosis
We classified the patients with schizophrenia into two groups: the patients with [DSP(+) group: N=108, 32.4%: age 45.3±14.2 years: age range 20–77 years] and without DSP episode(s) [DSP(-) group: N=169, 50.8%: age 44.8±14.4 years: age range 20–81 years], based on our investigation of their complete medical records. For the other 56 patients (16.8%), it was not possible to accurately categorize them due to a lack of sufficient medical information. We thereby evaluated the presence or absence of past DSP episodes for every patient according to the following criteria that were developed by our group, but were slightly modified from the original criteria proposed by Chouinard.18 In particular, we ignored the condition of “the last 5 years” and judged the following three items throughout each patient’s entire treatment history. These items cover the basic concept or criteria of DSP proposed by other researchers:4,5,19 1) Withdrawal psychosis: an acute relapse or exacerbation of psychosis appeared after a dose reduction or discontinuation of antipsychotics within 6 weeks for oral medication, and 3 months for long-acting intramuscular injection; or 2) the development of tolerance to antipsychotic effects: an acute relapse or exacerbation of psychosis that occurred independently of a dose reduction or discontinuation of antipsychotic therapy and that could not be successfully controlled by a 20% increased titration of the drug, or psychotic symptoms that were new to the patient or of greater severity, which occurred immediately after a decrease in drug dosage; or 3) TD: based on the information available from the medical records, if at least one of these three criteria was met, the patient was classified as a member of the DSP(+) group, and if not, the patient was classified as a member of the DSP(-) group. Since DSP is generally considered to be a secondary state that could be relevant to pharmacotherapy for some duration, we excluded patients who were in their first acute episode of psychosis or who had never shown relief from psychotic symptoms regardless of the duration of appropriate pharmacotherapy. We also excluded patients with comorbidities such as substance abuse/dependence, and patients who were clearly observed to refuse treatment and take medication. In addition, DSP could develop with antipsychotic treatment for some duration, but not a very long-term duration,18 and thus, the present study included patients who had undergone at least 1 year of pharmacotherapy. All of the decisions regarding the placement of patients into the DSP(+) or DSP(-) group were made by two of the authors (Yasunori Oda and Nobuhisa Kanahara), based on clinical observations for at least 2 years after the blood-drawing procedure. If their assessment of a patient differed, the two raters discussed the case in depth to reach a consensus for the placement.
Single nucleotide polymorphisms and genotyping
We selected five SNPs (rs883576, rs867755, rs2287694, rs1130857, rs335435) of GRK6 and three SNPs (rs2271167, rs1045280, rs3786047) of ARRB2. All of these SNPs were chosen on the basis of a minor allele frequency (≥0.05) in reference to the database for the Japanese population (the Japanese Single Nucleotide Polymorphisms [JSNP] database, at http://snp.ims.u-tokyo.ac.jp/).20
Blood samples were obtained from all participants. The DNA was isolated from whole blood using a QIAamp DNA Blood Mini kit (250) (Qiagen, Valencia, CA, USA). Genotyping was performed with a TaqMan assay (Applied Biosystems, Foster City, CA, USA) and the ABI PRISM 7000 Sequence Detection System (Applied Biosystems, Foster City, CA, USA) in accord with the manufacturer’s protocols. A polymerase chain reaction was performed under the following conditions: 1 cycle at 95°C for 10 minutes, followed by 40 cycles at 92°C for 15 seconds, and at 58°C (rs335435), or 60°C (rs883576, rs867755, rs2287694, rs1130857, rs2271167, rs1045280, and rs3786047) for 60 seconds.
Statistical analysis
Deviation from the Hardy–Weinberg equilibrium was tested using the Pearson chi-square test, and the differences between groups [ie, SZ patients vs controls and DSP(+) group vs DSP(-) group] in terms of their genotype and allele frequencies were tested using the Pearson chi-square tests for independence (3×2 table for genotype and 2×2 table for allele frequency, respectively). These analytic procedures were conducted with SPSS Version 21.0 for Windows (Chicago, IL, USA). The statistical significance threshold was set at P<0.05 (two-tailed).
Results
The genotype distributions of seven of the eight SNPs examined in the control subjects and patients with schizophrenia were in the Hardy–Weinberg equilibrium, but the SNP rs335435 of GRK6 was not. The analyses revealed no significant differences in allelic or genotyping distributions between the patients and the controls for all SNPs examined in this study (Table 2). These results did not change in the separate analyses of the male and female subject groups. In addition, in our within-cases comparison between the DSP(+) and DSP(-) groups, no significant difference in allelic or genotyping distributions was observed in any SNPs of either GRK6 or ARRB2. Although only the genotyping distribution of rs335435 of GRK6 indicated a tendency to differ between the DSP(+) and DSP(-) groups (P=0.045), this P-value did not survive as a significant difference after Bonferroni correction (Table 3).
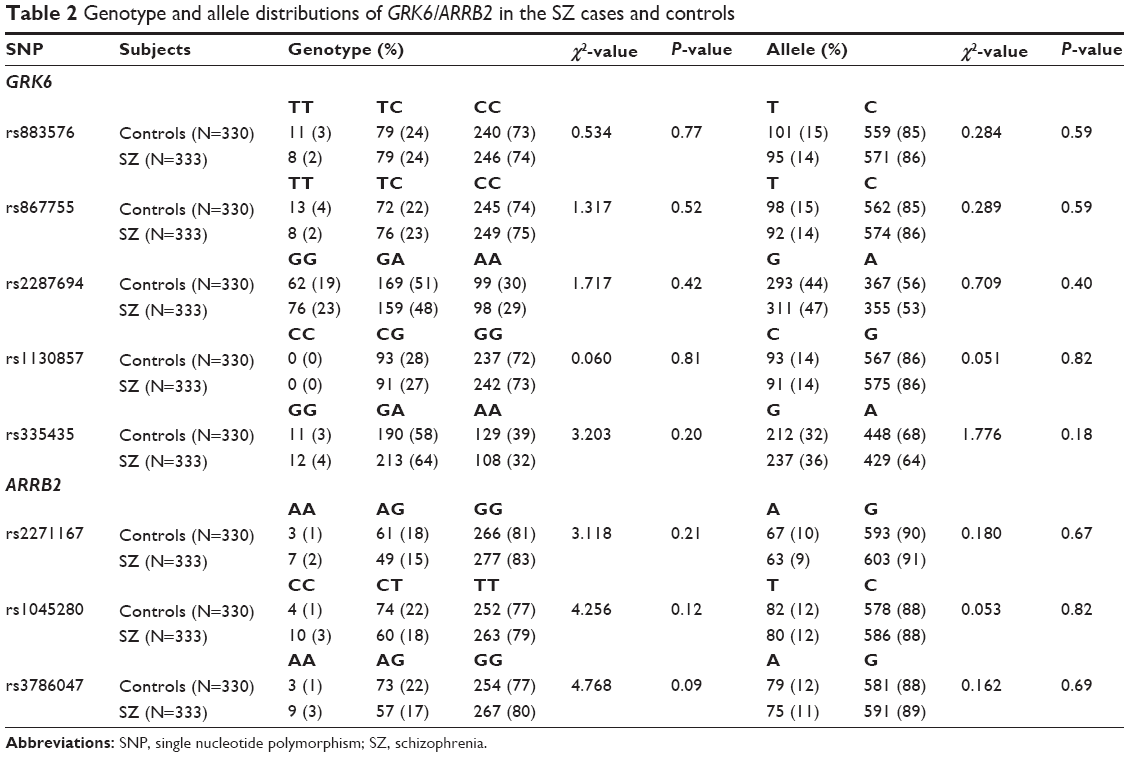 | Table 2 Genotype and allele distributions of GRK6/ARRB2 in the SZ cases and controls |
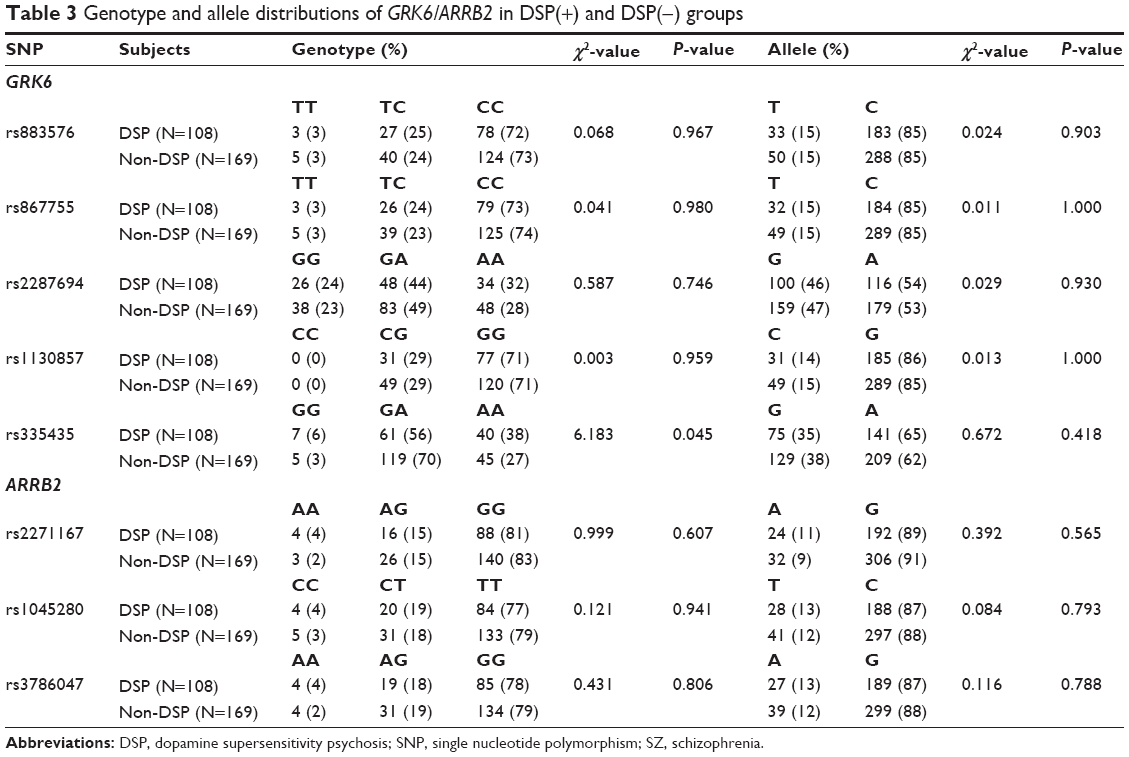 | Table 3 Genotype and allele distributions of GRK6/ARRB2 in DSP(+) and DSP(−) groups |
Discussion
Our results revealed that neither the allelic nor the genotyping distributions of all eight SNPs of GRK6 and ARRB2 showed a significant difference between the patients with schizophrenia and the controls. Our further analyses based on the classification of the clinical phenotype, ie, DSP(+) or DSP(-), also showed no significant difference in the allelic or genotyping distributions of these genes between the groups. We had hypothesized that the GRK6 and/or ARRB2 gene were involved in the development of DSP, which could affect the symptoms or clinical course of patients presenting with such an episode, rather than disease susceptibility. However, the present results did not support this hypothesis.
GRK6 and ARRB2 are profoundly involved in DRD2 trafficking, suggesting that they are key proteins in the DRD2 upregulation caused by a chronic administration of antipsychotics. GRK6 is widely expressed throughout many brain areas, particularly in the caudate/putamen in rat brain.21 Overexpression of GRK6 in rats promoted the internalization of DRD2 and also improved L-DOPA-induced dyskinesia, possibly by preventing an excessive G-protein signal.22 On the other hand, GRK6-knockout mice showed a significantly increased locomotor response to psychostimulants such as cocaine and amphetamine,23 and the mice inhibited the ability of haloperidol to induce catalepsy, possibly due to a lack of suppression of the G-protein signal.24 Similarly, ARRB2 is involved in the internalization of DRD2 following the phosphorylation of DRD2 by GRK6, and it was reported that ARRB2-knockout mice were conversely less sensitive to dopamine.25 These findings indicate that GRK6 and ARRB2 strongly participate in the internalization of DRD2 following the binding to the receptor by endogenous dopamine or antipsychotics.
We conducted a retrospective investigation of the medical records and diagnosed each patient as having DSP or not. Iatrogenic DSP appears with specific clinical features such as withdrawal psychosis and TD. Here, we also focused on the development of tolerance to antipsychotic effects, which indicates extremely severe exacerbation and difficulty in controlling the relapsed psychotic symptoms. These three episodes (ie, withdrawal psychosis, TD, and developed tolerance to antipsychotics) are included in the original criteria of DSP by Chouinard,18 and the diagnoses of DSP(+) vs DSP(-) in our study were made on the basis of these episodes.
Nevertheless, the present results did not support the hypothesis that these two molecules play key roles in developing clinical DSP, and this finding would be explained by several reasons as follows: first, unknown molecules other than GRK6 and ARRB2 may be involved in the occurrence of clinical DSP. Second, each aspect of the DSP phenotype, eg, TD, withdrawal psychosis, and developed tolerance to antipsychotic effects, has a biological background that is different from those of the others, and thus the extent of the effect of their differential distribution in the specific DSP phenotype might be reduced when all aspects of the DSP phenotypes were bracketed together as DSP(+), as done in the present study.
To the best of our knowledge, there are only three studies about the genetic association of GRK6 and ARRB2. Among them, the studies of GRK616 and ARRB214 reported no significant difference in any SNP distributions between patients with schizophrenia and control groups, although neither of these studies focused on DSP episodes in the patient group. Another study of ARRB215 examined the association between rs1045280 and TD induced by antipsychotics in patients with schizophrenia, and the results demonstrated that the T allele in rs1045280 was a risk allele for TD. However, in our study, there was no significant difference in rs1045280 between the patients with episodes of TD and those without. In light of the findings reported in these studies and our present results, it is possible that GRK6 and ARRB2 are not susceptibility genes for schizophrenia itself, but are involved in the clinical expression of DSP as by-products of treatment with antipsychotics. However, further studies considering the symptomatological variance of DSP are necessary to test this idea.
There are several limitations in the present study. First, we conducted a retrospective investigation of medical records. If a patient in the present study had had TD at any treatment period but the dyskinesia was successfully resolved by subsequent appropriate therapy, she/he could have been classified in the DSP(-) group, when there was no accurate registration of the TD in his/her medical records. In other words, the diagnosis of DSP might have been done in a conservative manner, resulting in a slight underestimation of its presence. Our sample size was very small, particularly for the DSP(+) group, leading to a reduction of the statistical power of our analysis. However, previous studies showed that the number of DSP(+) patients is generally small at 4.5%–22% of schizophrenia patients.26,27 Second, even if a patient was considered a member of the DSP(-) group based on his/her clinical history, future switching or increasing of antipsychotics could induce one or more new DSP episodes, indicating that such a patient could latently develop DSP. Third, the development and clinical appearance of DSP are dependent on varying aspects of antipsychotic treatment: its duration, class and dosage, adherence level, and more. In the present study, we did not access such detailed treatment information, and thus further studies using homogeneous treatment series are needed. Fourth, our patients and controls were not age- and sex-matched, and both the patient and control groups showed a wide age range. Thus, some caution is merited regarding the interpretation of our findings. For example, it is possible that some of the young-aged subjects among the controls could show a transition to psychosis in the future.
Conclusion
In conclusion, the present findings suggest that in eight SNPs of the GRK6/ARRB2 gene, there is no significant difference between patients with schizophrenia and healthy controls, nor between DSP(+) and DSP(−) patients with schizophrenia. However, since these molecules play significant roles in DRD2 trafficking following the action of antipsychotic(s), further studies focusing on clinical DSP episodes and specific antipsychotics are needed.
Acknowledgment
This study was supported by Grant-in-Aid for Young Scientists (B) from the Japan Society for the Promotion of Science (JSPS).
Disclosure
Dr Kanahara reported honoraria from Eli Lilly, Otsuka, and Janssen. Dr Kimura reported honoraria from Janssen, Meiji Seika, and Otsuka. Dr Watanabe received a lecturer’s fee from Eli Lilly, Dainippon Sumitomo, Takeda, and Mochida. Dr Hashimoto received the research grant or consultant fee from Abbott, Astellas, Otsuka, Taisho, and Dainippon Sumitomo. Dr Iyo received a consultant fee from Eli Lilly, Dainippon Sumitomo, Pfizer, and Abbott, and reported honoraria from Janssen, Eli Lilly, Otsuka, Meiji Seika, Astellas, Dainippon Sumitomo, Ono, GlaxoSmithKline, Takeda, Mochida, Kyowa Hakko, MSD, Eisai, Daiichi-Sankyo, Novartis, Teijin, Shionogi, Hisamitsu, and Asahi Kasei. The authors report no other conflicts of interest in this work.
References
Seeman P. Targeting the dopamine D2 receptor in schizophrenia. Expert Opin Ther Targets. 2006;10:515–531. | ||
Kane J, Honigfeld G, Singer J, et al. Clozapine for the treatment-resistant schizophrenic. A double-blind comparison with chlorpromazine. Arch Gen Psychiatry. 1988;45:789–796. | ||
Brenner HD, Dencker SJ, Goldstein MJ, et al. Defining treatment refractoriness in schizophrenia. Schizophr Bull. 1990;16:551–561. | ||
Moncrieff J. Does antipsychotic withdrawal provoke psychosis? Review of the literature on rapid onset psychosis (supersensitivity psychosis) and withdrawal-related relapse. Acta Psychiatr Scand. 2006;114:3–13. | ||
Iyo M, Tadokoro S, Kanahara N, et al. Optimal extent of dopamine D2 receptor occupancy by antipsychotics for treatment of dopamine supersensitivity psychosis and late-onset psychosis. J Clin Psychopharmacol. 2013;33:398–404. | ||
Chouinard G, Chouinard VA. A typical antipsychotics: CATIE study, drug-induced movement disorder and resulting iatrogenic psychiatric-like symptoms, supersensitivity rebound psychosis and withdrawal discontinuation syndromes. Psychother Psychosom. 2008;77:69–77. | ||
Köhler U, Schröder H, Augusatin W, Sabel BA. A new animal model of dopamine supersensitivity using s.c. implantation of haloperidol releasing polymers. Neurosci Lett. 1994;170:99–102. | ||
Huang N, Ase AR, Hébert C, van Gelder NM, Reader TA. Effects of chronic neuroleptic treatments on dopamine D1 and D2 receptors: homogenate binding and autoradiographic studies. Neurochem Int. 1997;30:277–290. | ||
Inoue A, Miki S, Seto M, et al. Aripiprazole, a novel antipsychotic drug, inhibits quinpirole-evoked GTPase activity but does not up-regulate dopamine D2 receptor following repeated treatment in the rat striatum. Eur J Pharmacol. 1997;321:105–111. | ||
Silvestri S, Seeman MV, Negrete JC, et al. Increased dopamine D2 receptor binding after long-term treatment with antipsychotics in humans: a clinical PET study. Psychopharmacology (Berl). 2000;152:174–180. | ||
Tadokoro S, Okamura N, Sekine Y, Kanahara N, Hashimoto K, Iyo M. Chronic treatment with aripiprazole prevents development of dopamine supersensitivity and potentially supersensitivity psychosis. Schizophr Bull. 2011;38:1012–1020. | ||
Suzuki T, Kanahara N, Yamanaka H, et al. Dopamine supersensitivity psychosis as a pivotal factor in treatment-resistant schizophrenia. Psychiatr Res. 2015;227:278–282. | ||
Métayé T, Gibelin H, Perdrisot R, Kraimps JL. Pathophysiological roles of G-protein-coupled receptor kinases. Cell Signal. 2005;17:917–928. | ||
Ikeda M, Ozaki N, Suzuki T, et al. Possible association of beta-arrestin 2 gene with methamphetamine use disorder, but not schizophrenia. Genes Brain Behav. 2007;6:107–112. | ||
Liou YJ, Wang YC, Chen JY, et al. The coding-synonymous polymorphism rs1045280 (Ser280Ser) in beta-arrestin 2 (ARRB2) gene is associated with tardive dyskinesia in Chinese patients with schizophrenia. Eur J Neurol. 2008;15:1406–1408. | ||
Tang W, Cai J, Yi Z, Zhang Y, Lu W, Zhang C. Association study of common variants within the G protein-coupled receptor kinase 6 gene and schizophrenia susceptibility in Han Chinese. Hum Psychopharmacol. 2014;29:100–103. | ||
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 4th-TR ed. Washington, DC: American Psychiatric Press; 2000. | ||
Chouinard G. Severe cases of neuroleptic-induced supersensitivity psychosis. Diagnostic criteria for the disorder and its treatment. Schizophr Res. 1990;5:21–33. | ||
Kirkpatrick B, Alphs L, Buchanan RW. The concept of supersensitivity psychosis. J Nerv Ment Dis. 1992;180:265–270. | ||
Hirakawa M, Tanaka T, Hashimoto Y, et al. JSNP: a database of common gene variation in the Japanese population. Nucleic Acids Res. 2002;30:158–162. | ||
Erdtmann-Vourliotis M, Mayer P, Ammon S, Riechert U, Höllt V. Distribution of G-protein-coupled receptor kinase (GRK) isoforms 2, 3, 5 and 6 mRNA in the rat brain. Brain Res Mol Brain Res. 2001;95:129–137. | ||
Ahmed MR, Berthet A, Bychkov E, et al. Lentiviral overexpression of GRK6 alleviates L-dopa-induced dyskinesia in experimental Parkinson’s disease. Sci Transl Med. 2010;2:28ra28. | ||
Gainetdinov RR, Bohn LM, Sotnikova TD, et al. Dopaminergic supersensitivity in G protein-coupled receptor kinase 6-deficient mice. Neuron. 2003;38:291–303. | ||
Managò F, Espinoza S, Salahpour A, et al. The role of GRK6 in animal models of Parkinson’s disease and L-DOPA treatment. Sci Rep. 2012;2:301. | ||
Bohn LM, Gainetdinov RR, Caron MG. G protein-coupled receptor kinase/beta-arrestin systems and drugs of abuse: psychostimulant and opiate studies in knockout mice. Neuromolecular Med. 2004;5:41–50. | ||
Chouinard G, Annable L, Chouinard AR. Supersensitivity psychosis and tardive dyskinesia: a survey in schizophrenic outpatients. Psychopharmacol Bull. 1986;22:891–896. | ||
Hunt JI, Singh H, Simpson GM. Neuroleptic-induced supersensitivity psychosis: retrospective study of schizophrenic inpatients. J Clin Psychiatry. 1988;49:258–261. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.