Back to Journals » Neuropsychiatric Disease and Treatment » Volume 19
Gene Polymorphisms and Expression of NRG1, DAOA, and DISC1 Genes in a Chinese Han Population with an Ultra-High Risk for Psychosis
Authors He XY, Huang ZH, Wang F, Chen ZL, Wang SB, Jia FJ, Hou CL
Received 10 August 2023
Accepted for publication 5 November 2023
Published 21 November 2023 Volume 2023:19 Pages 2521—2533
DOI https://doi.org/10.2147/NDT.S434856
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Xiao-Yan He,1,2 Zhuo-Hui Huang,1 Fei Wang,1 Zi-Lang Chen,3 Shi-Bin Wang,1 Fu-Jun Jia,1 Cai-Lan Hou1
1Psychological Department, Guangdong Mental Health Center, Guangdong Provincial People’s Hospital (Guangdong Academy of Medical Sciences), Southern Medical University, Guangzhou, People’s Republic of China; 2Psychiatric Rehabilitation Section, The Affiliated Mental Health Center of Jiangnan University, Wuxi Central Rehabilitation Hospital, Wuxi Central Rehabilitation Hospital, Wuxi, People’s Republic of China; 3Psychiatry Department, Luoding Mental Health Center, Yunfu, People’s Republic of China
Correspondence: Cai-Lan Hou, Psychological Department, Guangdong Mental Health Center, Guangdong Provincial People’s Hospital (Guangdong Academy of Medical Sciences), Southern Medical University, Room 709, No. 123, Huifuxi Road, Yuexiu District of Guangzhou, Guangdong, People’s Republic of China, Email [email protected]
Purpose: Although there is previous evidence supporting that ultra-high risk (UHR) for psychosis transformation is associated with NRG1, DAOA, and DISC1 genes, there have been no relevant studies in the Chinese population. The objective of the current study was to explore the gene polymorphism and expression of NRG1, DAOA, and DISC1 genes in a Han population with UHR for psychosis in China.
Methods: Eighteen UHR individuals, 61 first-degree relatives of patients with schizophrenia (FDR), 55 first-episode psychosis individuals (FEP), and 61 healthy controls (HC) were enrolled in the study. The genotypes at four loci of the NRG1 gene, four loci of the DAOA gene, and two loci of the DISC1 gene were tested for all subjects, and mRNAs of NRG1 and DISC1 were examined and analyzed in a pairwise comparison among the four groups. Statistical analysis of genetics was performed using snpStats software. For the case-control association analysis, a single site association study, epistatic effect analysis, and haplotype analysis were used to explore the association of the above genes.
Results: This study found that rs3918341 in the DAOA gene was associated with susceptibility to UHR by single site association analysis. Epistatic effect analysis results showed that the NRG1 gene interacted with the DAOA gene and DISC1 gene in the susceptibility to UHR. Haplotype association analysis showed that all haplotypes were not significantly associated with UHR. NRG1 mRNA was significantly downregulated in the UHR group compared with the HC group as well as the FEP group.
Conclusion: Our preliminary results show that NRG1, DAOA, and DISC1 genes may play a role in psychosis onset, opening the way to the identification of prognostic biomarkers.
Keywords: UHR, genetic susceptibility, NRG1, DAOA, DISC1
Introduction
Ultra-high risk (UHR) for psychosis is a special condition existing before the full-blown onset of psychosis, such as schizophrenia.1 Identification of the UHR population and early intervention are extremely important to prevent or even delay the onset of disorders such as schizophrenia.2,3 A recent meta-study showed a conversion rate of UHR to psychosis was 20% in 2 years.4 However, 80% of the UHR population remains unconverted, so how to more accurately identify and predict the conversion of UHR has become a research focus. However, current diagnosis of UHR often depends on self-report by interview-based subjective assessment, and thus there is an urgent need for biomarkers to determine the diagnosis and prognosis of UHR.
Previous studies have shown that both genome-wide association studies and DNA methylation studies indicate an important role for genetics in the development of schizophrenia.5,6 Hypofunction of NMDA receptors may lead to decreased dopamine activity in the mesocortical pathway, which may manifest as negative symptoms and cognitive dysfunction in schizophrenia.7 Glutamate NMDA receptor hypofunction, generated early in brain development, is regulated by genetic factors acting together with environmental factors to create abnormal fixed loop circuits in the development of the brain’s nervous system, causing very early-onset schizophrenia with minimal or no trigger in adulthood. Reduced NMDA receptor function in schizophrenia may be partly explained by increased d-aminoacid oxidase (DAO) activity regulated by the d-aminoacid oxidase activator (DAOA) leading to a decrease in D-serine, a co-agonist of NMDA receptors.7 Neuregulin 1 (NRG1) function is mediated by binding to the receptor tyrosine kinases ErbB (ErbB3 and ErbB4), and altered NRG1/ErbB4 signaling is thought to contribute to NMDA receptor hypofunction.8,9 Namba et al10 found that DISC1 is a downstream molecule of the NMDA receptor signaling pathway, and DISC1 can be exogenously expressed by lentiviral mediation, which can then improve the transitional migration of newborn neurons triggered by the use of NMDA receptor antagonists. On the other hand, DISC1 plays a key role in dopamine (DA) neurotransmission.11 Previous studies have also shown that the addition of drugs that selectively block DA receptors can improve cognitive performance and psychiatric symptoms in clozapine-resistant treatment-refractory schizophrenia.12
The NRG1, DAOA, and DISC1 genes are therefore all-important candidates for the development of schizophrenia, and similarly it has been shown that polymorphisms or expression characteristics of these genes change at the UHR stage. NRG1 polymorphisms were also shown to predict UHR conversion to schizophrenia.13,14 Two other studies have found that the level of NRG1 mRNA was also able to predict UHR conversion as well as to influence the functional outcome of UHR.15,16 Moreover, it has also been found that polymorphisms in DAOA predict UHR conversion.17 Another study has shown differential expression of the DISC1 gene in different stages of schizophrenia.18 As mentioned above, NRG1, DAOA, and DISC1 may also be promising predictive biomarkers of UHR and conversion to psychosis.
Therefore, the present study aims to examine and compare the polymorphisms and expression in NRG1, DAOA, and DISC1 genes in subjects at different stages of schizophrenia, including healthy controls (HC), first-degree relatives (FDR) of patients with schizophrenia, UHR individuals, and first episode patients with experiences of schizophrenia (FEP).
Methods
Ethical Considerations
The study was conducted by Guangdong Mental Health Center of Guangdong Provincial Peoples’ Hospital, Xinhui Third People’s Hospital and the Luoding Psychiatric Hospital in China. The protocol was approved by the Clinical Research Ethics Committee of Guangdong Provincial Peoples’ Hospital. All participants and their legal guardians (participants less than 18 years old) gave written informed consent. The study was conducted in accordance with the revised Helsinki declaration criteria.
Study Participants
A total of 195 individuals were enrolled in the study, including the UHR group (n=18), FDR group (n=61), FEP group (n=55), and HC group (n=61). The specific operation process is as follows. Patients with schizophrenia were identified in outpatient clinics, wards, or in the community according to ICD-10 criteria, and if any first-degree relatives (parents, children, siblings) of the patient were between the ages of 15 and 45, they were contacted by the investigator, and if the family members of these schizophrenics agreed to participate in the study, the investigator interviewed these family members. All enrollees were evaluated at the first meeting using the Structured Inventory of Prodromal Syndrome (SIPS)19 to determine if they met the criteria for prodromal syndrome (COPS). Those who met COPS as assessed by the SIPS were defined as having a psychiatric risk syndrome for inclusion in the study. In short, 38 subjects satisfied the SIPS criterion for attenuated positive symptom states (APS), two subjects met the SIPS Genetic Risk and Exacerbation Status (GRDS) requirements, and no subjects met the Brief Intermittent Psychotic States (BIPS) criteria. All UHR group patients were medication-naive. Participants in the FDR group were also recruited from first-degree relatives (siblings, parents, or children) of patients with schizophrenia who did not meet the diagnostic criteria for the UHR.
FEP group were patients who met the ICD-1020 diagnostic criteria for schizophrenia with a first episode, and disease duration within 1 year. The HC group was recruited by posting advertisements in the community, and the diagnosis by the psychiatrist ruled out a history of diagnosis or treatment history of psychiatric disorders and had no positive psychiatric family history.
All participants were aged 15–45 years. They were able to sign an informed consent form and were compliant with the study arrangements. They could understand the content of the questionnaire and cooperate to complete all the test examinations. Participants were excluded from this study if they had severe or unstable physical diseases, patients with epilepsy, cerebrovascular disease, or other central nervous system organic diseases, and they had drug dependence or abuse.
Assessments
Data on socio-demographic variables, such as gender, age, marital status, educational level, and employment status were assessed using a form designed for this study by two psychiatrists with at least 3 years of clinical and research experience.
Symptomatology was determined using the Chinese version of positive and negative syndrome scale (PANSS).21 Depressive symptoms were assessed using the Chinese version of the Montgomery-Åsberg Depression Rating Scale (MADRS).22,23 Overall function was evaluated using the global assessment of functioning scale (GAF).24
Single-Nucleotide Polymorphism Selection
SNPs (single nucleotide polymorphisms) of the NRG1, DAOA, and DISC1 genes were determined through the NCBI (www.ncbi.nlm.nlh.gov/SNP) database on the basis of references to relevant literature. Inclusion criteria for SNPs were: (1) positive loci previously reported in the literature to be associated with UHR transformation; (2) selected functional SNPs that are in exon regions (part of the 5’UTR region, 3’UTR region, sites of nonsynonymous mutations in introns); (3) minimum allele frequency ≥5%; (4) trying to select tag SNPs as much as possible; and tag SNPs were determined using the following methods including locate the genotype data of the East Asian population in the 1000 Genomes browser database, applying the solid spine module in haploview software (http://www.Haploview.com) to determine the linkage disequilibrium (LD) regions of genes and selecting one SNPs with a minimum minor allele frequency (MAF) ≥5% in the linkage disequilibrium (LD) region as tag SNPs. Finally, 10 loci were included in this study (NRG1:rs6994992, rs4281084, rs12155594, rs3924999; DAOA:rs778294, rs1341402, rs2391191, rs3918341, DISC1:rs821616, rs3738402).
Genotyping and Gene Expression
Genomic DNA was extracted from whole blood samples of all participants using a standard phenol/chloroform method. Gene expression was performed in four steps, the first being extraction of total RNA, according to the hipure liquid RNA/miRNA kit Pdf instructions operation, the second is to perform RT-PCR, the third is qPCR system and procedure, and the fourth is to run the gum validation.
Statistical Analyses
The SPSS21.0 package was used to analyze the data. The demographic and clinical characteristics of the FEP, UHR, FDR, and HC groups were analyzed using Pearson chi-square analyses or one-way analysis of variance (ANOVA).
SnpStats (online software) was used for analysis of allele frequency, genotype frequency, and Hardy-Weinberg test.25 SNPassoc was employed for correlation analysis on SNP obtained from the different samples. Hardy-Weinberg equilibrium at SNP sites was evaluated with Chi-squared test to assess the potential association between the polymorphism and sample by designing the contingency table. Logistic regression method was used to measure the correlation between genotype and the distinct groups, and the correlation was quantified by calculating OR (odds ratio) of every genotype compared to the reference genome. The optimized SNP model was selected by comparing Akaike’s Information Criterion (AIC). A lower P-value suggests a lower AIC value with higher level of confidence. Haplovdew was employed to calculate haplotype on those SNP loci satisfying Hardy-Weinberg equilibrium and located on the same chromosome. The haplotype frequency was investigated by using the Chi-squared test to measure the differences between groups. In this study, genetic polymorphism loci were compared using two-by-two way, and three comparisons were conducted, namely UHR group vs FEP group, UHR group vs FDR group, and UHR group vs HC group.
The gene expression was quantified with the 2−ΔΔct method to measure the group mean value and statistically examined by T test to get the P-value. All tests were two-sided and considered statistically significant at P<0.05.
Results
Demographic and Clinical Characteristics of the Four Groups
There were no statistically significant differences among the four groups of FEP, UHR, FDR, and HC, regarding gender, but statistically significant differences regarding age, marital status, employment status, and educational level. In the FEP, UHR, and FDR groups, these three groups showed a gradual decline in the PANSS total score, PANSS positive subscale score, PANSS negative subscale score, as well as the PANSS general psychopathology subscale. Additionally, the UHR group was significantly higher than the FDR and HC groups on the SIPS total score as well as on the various subscales. GAF total score increased gradually in the FEP, UHR, FDR, and HC groups, while the MADRS total score decreased gradually (Supplement S1).
Testing for Hardy Weinberg Equilibrium at SNP Loci
All eight polymorphic loci were subjected to Hardy-Weinberg equilibrium test, except Locus rs12155594 of NRG1 and Locus rs1341402 of DAOA, where only one genotype was present and hence it was not appropriate for Hardy-Weinberg test. The sampling results showed that Locus rs3918341 of DAOA did not fit H-W equilibrium on the FEP group and UHR group, and Locus rs3738402 of DISC1 did not fit H-W equilibrium on the FDR group. All other groups could fit H-W equilibrium on all the loci (Supplement S2).
UHR Group vs FEP Group
Analysis of SNP Loci Genotypes between FEP and UHR Groups
SNP rs12155594, SNP rs1341402, and SNP rs778294 were eliminated as they contained only one or two genotypes and could not be investigated by association analysis. The results demonstrated that Locus rs3918341 located in DAOA gene included the genotypes GG, AG, and AA, and distribution in the UHR and FEP groups was not identical. The frequencies showed by these genotypes in UHR group were 22.2%, 77.8%, and 0%, respectively, and in FEP group were 36.4%, 34.5%, and 29.1%, respectively. Hence, the frequency of genotype AG in the UHR group was significantly higher in comparison to that in the FEP group. Under the co-dominant model, genotype frequency distribution of SNP rs3918341 showed a significant difference between the groups (OR=3.68, 95% CI=1.03–13.02, P<0.001). These results demonstrated that genotype AG at Locus rs3918341 of the DAOA gene was a susceptibility gene of UHR. Interestingly, SNP rs2391191 of DAOA was found with a missing value, thereby showing no significant difference between the groups despite a low P-value (0.0088). However, other SNPs showed no distinct differences (Table 1).
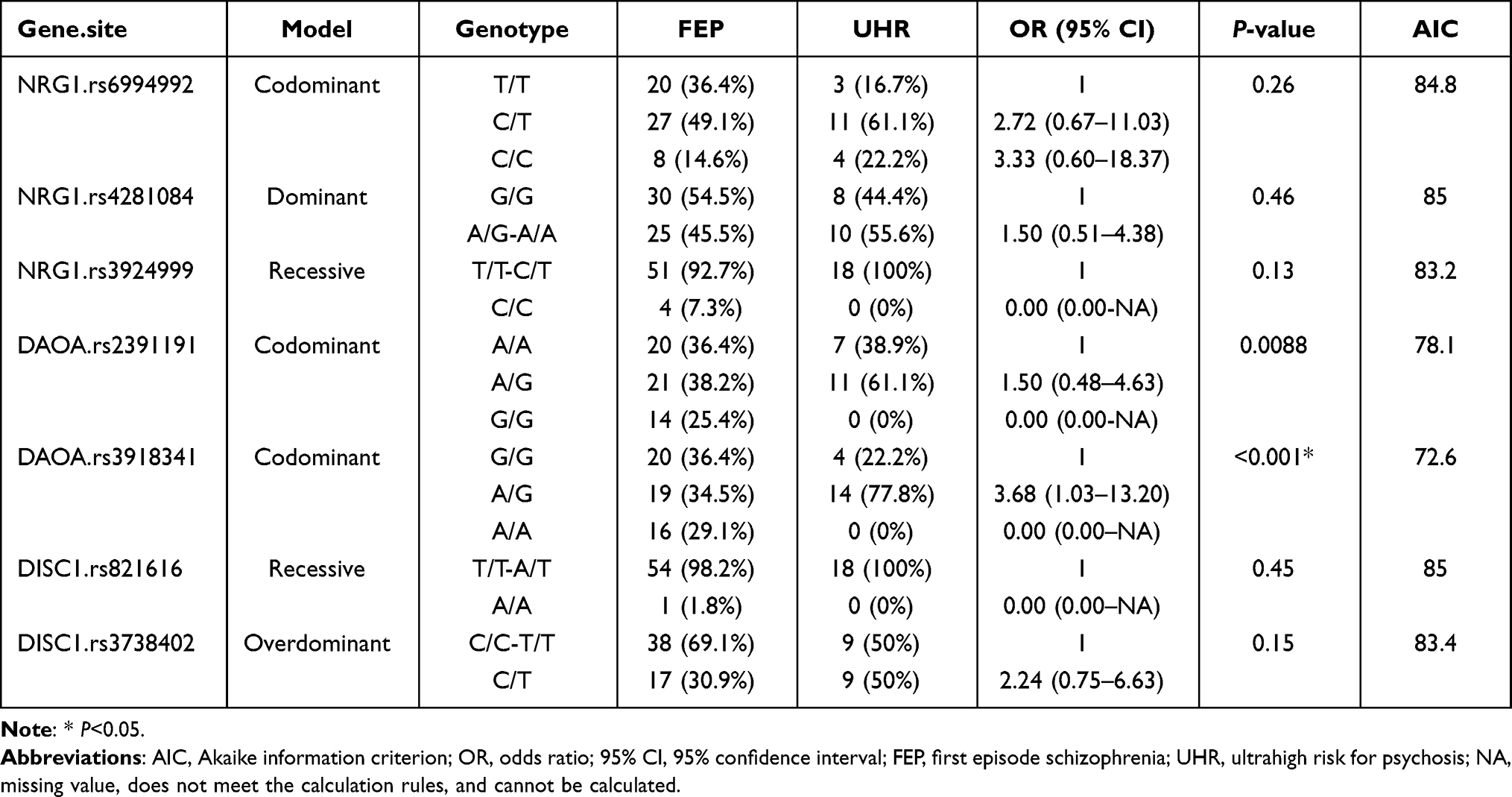 |
Table 1 Genotype for SNPs of NRG1, DAOA, and DISC1 Genes Family in the UHR and FEP Groups |
Interactions between SNPs of the NRG1, DAOA, and DISC1 Genes Family and UHR Risk
In this study, the log-likelihood ratio test was employed to clarify epistatic interaction among the SNPs susceptible to UHR. As indicated, Locus rs2391191 of DAOA (P=0.0177) exhibited the major effect on the UHR risk (recessive model). In addition, log-likelihood ratio test analysis confirmed epistatic interaction of Locus rs6994992 of NRG1 and Locus rs778294 of DAOA (dominant model Pinteraction=0.045), and another epistatic interaction between Locus rs6994991 of NRG1 and Locus rs821616 of DISC1 (over-dominant model, Pinteraction=0.04), which suggested that interactions between the various candidate genes may be involved in UHR (Table 2).
 |
Table 2 Epistatic Interactions between NRG1, DAOA, and DISC1 Genes Family in the UHR and FEP Groups |
Analysis of Haplotypes between SNPs
Haploid analysis using the EM algorithm displayed no significant difference between the FEP group and UHR group on haplotype (Supplement S3).
UHR Group vs FDR Group
Analysis of SNP Loci Genotypes between UHR and FDR Groups
SNP rs12155594, SNP rs1341402, and SNP rs778294 were not entered into the association analysis because they contained only one or two genotypes. The results demonstrated that Locus rs3918341 located in the DAOA gene included the genotypes GG, AG, and AA, and distribution in the UHR and FDR groups was not identical. The frequencies shown by these genotypes in the UHR group were 22.2%, 77.8%, and 0%, respectively, and in the FEP group were 18%, 55.7%, and 44.3%, respectively. Hence, the frequency of genotype AG in the UHR group was significantly higher in comparison to that in the FDR group. Under an over-dominant model, genotype frequency distribution of SNP rs3918341 showed a significant difference between the groups (OR=4.41, 95% CI=1.30–14.94, P=0.01). These results demonstrated that genotype AG at Locus rs3918341 of DAOA gene was a susceptibility gene of UHR. Interestingly, SNP rs2391191 of DAOA was found to have a missing value, thereby showing no significant difference between the groups despite low P-value (0.013). However, other SNPs showed no distinct differences (Table 3).
 |
Table 3 Genotype for SNPs of NRG1, DAOA, and DISC1 Genes Family in the UHR and FDR Groups |
Interactions between SNPs of NRG1, DAOA, and DISC1 Genes Family and UHR Risk
Log-likelihood ratio test analysis confirmed epistatic interaction of Locus rs6994992 of NRG1 and Locus rs2391191 of DAOA (over-dominant model Pinteraction=0.0169), which suggested that interactions between the various candidate genes may be involved in UHR (Table 4).
 |
Table 4 Epistatic Interactions between NRG1, DAOA, and DISC1 Genes Family in the UHR and FDR Groups |
Analysis of Haplotypes between SNPs
Haplotype analysis showed no significant difference between the UHR group and FDR group (Supplement S4).
UHR Group vs HC Group
Analysis of SNP Loci Genotypes between UHR and HC Groups
For the same reason as above, SNP rs12155594, SNP rs1341402, and SNP rs778294 were excluded in the association analysis. The results did not reveal which locus had a genotype that differed between the two groups in the UHR group versus the HC group (Table 5).
 |
Table 5 Genotype for SNPs of NRG1, DAOA, and DISC1 Genes Family in the UHR and HC Groups |
Interactions between SNPs of NRG1, DAOA, and DISC1 Genes Family and UHR Risk
Log-likelihood ratio test analysis confirmed epistatic interaction of Locus rs6994992 of NRG1 and Locus rs2391191 of DAOA (over-dominant model Pinteraction=0.011), and another epistatic interaction between Locus rs4281084 of NRG1 and Locus rs2391191 of DAOA (over-dominant model, Pinteraction=0.033) which suggested that interactions between the various candidate genes may be involved in UHR (Table 6).
 |
Table 6 Epistatic Interactions between NRG1, DAOA, and DISC1 Genes Family in the UHR and HC Groups |
Analysis of Haplotypes and Linkage Disequilibrium between SNPs
Haploid analysis displayed no significant difference between UHR group and HC group on haplotype (Supplement S5).
Results of Gene Expression of NRG1 and DISC1, in Four Groups
The expression content of the DAOA gene could not be measured because the expression level of the DAOA gene in tissues and blood was very low in the gene expression experiments and could not be amplified by qPCR. Only gene expression studies of the NRG1 gene as well as the DISC1 gene were performed. It can be seen from the table as well as the figure that, compared with the UHR group versus the HC group as well as the FEP group, NRG1 mRNA was significantly down regulated with statistical significance. There was no statistically significant difference in the expression amount of DISC1 gene among the four groups (Figure 1 and Supplement S6).
 |
Figure 1 Analysis of the relative expression of NRG1 gene and DISC1 gene; * represents less than 0.05, *** represents less than 0.001. The expression amounts of genes were obtained by the 2−ΔΔct method using the T test for P-values in pairwise comparisons between the UHR group and the other groups. |
Discussion
In this study, four loci in the NRG1 gene (rs699492, rs4281084, rs12155594, rs3924999), four loci in the DAOA gene (rs778294, rs1341402, rs2391191, rs3918341), and two loci in the DISC1 gene (rs821616, rs3738402) were compared between the four groups for differences in genotype distribution in the above 10 loci to understand the information of UHR genetics. At the same time, we measured the mRNA expression of these three genes, but, in the course of the experiment, DAOA gene expression could not be measured, so only the NRG1 gene and DISC1 gene expression were analyzed in the four groups.
The sampling results showed that Locus rs3918341 of DAOA did not fit H-W equilibrium on the FEP group and UHR group, and Locus rs3738402 of DISC1 did not fit H-W equilibrium on the FDR group. The reasons may be as follows, first, the H-W test could be vulnerable to the sample size, which implied that fewer samples or uneven sampling could lead to uneven distribution of the genotypes. This locus is primarily distributed in Asians with a G allele gene frequency of G = 0.6 and an A allele gene frequency = 0.4, which corresponds with the gene frequency in the UHR-FEP group in the present analysis. Thus, the results can be affected if genotype AA in the UHR group was zero during the sampling. On the other hand, previous studies have also shown that the case group may not conform to the H-W equilibrium because sites that are functionally subject to selective pressure (disease also counts as a selective pressure) and H-W disequilibrium is normal.26
The results of the gene polymorphism showed that locus rs3918341 of the DAOA genotype AG was a potential risk factor of the UHR group. The study of epistatic effects found that epistatic interaction of rs6994992 of NRG1 and rs778294 of DAOA; rs6994992 of NRG1 and rs821616 of DISC1; rs6994992 of NRG1 and rs2391191 of DAOA, as well as the epistatic interaction of rs4281084 of NRG1 and rs2391191 of DAOA caused susceptibility of UHR. Haploid analysis displayed no significant difference between the UHR group and other groups on haplotype. This is somewhat inconsistent with previous studies. Primary analysis between the genetic polymorphism and UHR was conducted in 2006 by Hall et al,13 where all 12 high-risk patients with genotype TT of Locus rs6994992 of NRG1 converted to the psychotic disorder, significantly more than those with genotype TC and CC. Similarly, Kéri et al27 found that all individuals with genotype TT of Locus rs6994992 of NRG1 developed psychotic disorder after 1 year of follow-up analysis in a population with ultra-high risk of psychosis. In 2013, Bousman et al28 conducted polymorphism analyses on 225 UHR individuals, but no UHR transformation was found to be predicted by polymorphism of Locus rs6994992. However, rs12155594 and rs4281084 of NRG1 were able to predict UHR transformation, and haploids of them were also functional. Mössner et al17 followed the UHR population for 2 years and reported that all individuals with the DAOA gene rs1341402 genotype CC exhibited psychiatric disorders, with a relative risk ratio 4.58 times higher than that of genotype TT or CT and the conversion rate of individuals with genotype AA at locus rs778294 was 50%. These studies showed that rs6994992, rs12155594, and rs4281084 of the NRG1 gene and rs1341402 and rs778294 of the DAOA gene were associated with the transformation or symptoms of UHR. However, the above loci were not found to be associated with the status of UHR in the analysis of gene polymorphisms in this study. There may be the following reasons for the inconsistent results. First, Locus rs12155594 of NRG1 and Locus rs1341402 of DAOA were found to be linked with one genotype only and eliminated in following analysis. This phenomenon demonstrated large variation of genotype distribution among the different races, thus paving way for more analyses among the different races. In addition, the results of this study are consistent with the view that differences between SNPs identified across studies that confer risk are on expected lines rather than the exception. Indeed, meta-analysis indicated that the specific NRG risk allele may vary among the different populations,29,30 and some researchers have pointed out that risk increased as a result of NRG1 sequence which may not be driven by single SNP but accumulated by diversification of the nucleotide polymorphism.31
In the study we found that genotype AG of Locus rs3918341 of DAOA showed significant higher frequency in UHR population than the FDR group and FEP group, concomitant with a higher frequency than the HC group (not statistically significant). It has been reported that Locus rs3918341 of DAOA displayed different genotype distribution in patients with major depression or bipolar disorder compared with the HC group.32,33 In addition, two meta-analyses have also reported that loci in DAOA is involved in schizophrenia,34,35 which is demonstrated by over-expression of DAOA in the dorsolateral prefrontal cortex in patients with schizophrenia.36 Additionally, the results of this study revealed that the distribution of genotypes varies considerably among the different ethnic groups, and genotype AG of Locus rs3918341 of DAOA is a potential risk genotype in the Chinese UHR population.
Epistatic interaction of Locus rs6994992 of NRG1 and Locus rs2391191 of DAOA were detected with effect on the UHR group upon comparison with the FEP group and HC group. Previous studies have demonstrated that risk allele, Locus rs6994992, constitutes a major part of primitive haplotype, altering three TF-binding loci in the NRG1 promotor region, and has been related to over-expression of IV type transcripts in autopsy tissues.37,38 In addition, another study demonstrated that patients with schizophrenia (with risk genotype T/T on Locus rs6994992) can express more unusual ideas when in conflicts, compared to those with type C carriers (C/T or C/C). Overall, significant mutual genotype-environment interaction of rs6994992 of NRG1 and social psychological stress has been observed.14
Locus rs2391191 of DAOA is a variant coding for DAOA protein, whose mutation can lead to lysine/arginine alteration. Recent analysis has demonstrated that this locus (rs2391191) is related to impaired verbal memory and reduced cortical thickness in patients with schizophrenia, whereas another research showed that polymorphism at Locus rs2391191 of DAOA in the NMDA pathway was most closely related to schizophrenia.39,40 It has been demonstrated that Locus rs2391191 (A) is a risk allele involved in the age of first onset of schizophrenia, especially during early-onset schizophrenia.41 Overall, loci rs6994992 and rs2391191 were potentially found to be related to schizophrenia, and this study indicated that epistatic interaction between them is a possible risk factor in UHR group. More future research is required to understand the mechanisms through which this mutation can increase the risk of schizophrenia.
In this study, we found a significant downregulation of NRG1 mRNA in the UHR group relative to the HC group as well as the FEP group. Previous conclusions regarding the influence of the expression content of the NRG1 gene on UHR are controversial. In 2012, Kiss et al15 examined the expression of NRG1 gene in 97 patients with UHR as well as 50 normal controls, 31 patients with UHR transformation after 1 year, and the expression level of NRG1 gene in peripheral blood was significantly lower in the UHR transformed group than in the UHR untransformed and normal groups, and there was no difference between the UHR untransformed and normal groups. Whereas a 2018 study of 185 high-risk as well as ultrahigh risk individuals with 12 months of follow-up tested for NRG1 gene expression levels at baseline, and distinguished their functional outcomes at 12 months, dividing them into a good functional outcome (GFO) group and a poor functional outcome (PFO) group, which showed that patients with PFO had significantly higher NRG1 mRNA levels than those with GFO. The results suggest that NRG1 may serve as a predictor of functional outcome in individuals at high risk for psychosis.16 Also with somewhat negative results, a 2017 study by Jagannath et al found no association between gene expression of NRG1 and transformation by UHR.42 Larger sample sizes and long-range follow-up studies may be needed in the future to explore the effect of the NRG1 gene on UHR recognition and transformation.
In this study, the amount of DISC1 gene expression in the UHR group was not found to be different from that in the other groups. The upregulation of the DISC1 gene was shown in a 2015 Santoro et al43 study comparing the UHR group with the FEP group, but statistical significance was lost when controlling for age as a covariate.
Limitations and Strengths
Limitations should be considered when interpreting these findings. First, this was a cross-sectional analysis, and as such we were not able to reveal the rate of UHR individuals’ conversion to schizophrenia, which prevented us from investigating the potential predictive value of polymorphisms and expression in NRG1, DAOA, and DISC1 genes with respect to FEP onset. Second, we investigated a limited number of SNPs of NRG1, DAOA, and DISC1 genes. Besides, such as the complex disease etiology of psychotic disorders, which is more suitable to do genome-wide association studies, the results of individual genes are limited. Finally, the sample size of this study was relatively small, especially in the UHR group, due to the small sample size, several groups differed in demographic comparisons, thereby limiting the generalizability of the findings. Therefore, additional sample sizes as well as longitudinal study methods need to be included in future studies to track the role of genes in transformation to UHR.
This study has some worthy strengths, including the few to explore the biological basis of UHR, expecting that the identification of UHR will be more precise. The second we examined whether the genetic basis of UHR differed between ethnicities by verifying previous positive genetic results regarding UHR transformation in the Chinese population. A further strength is the understanding of the NRG1, DAOA, and DISC1 genotypes and gene expression changes in the context of the different stages of schizophrenia, and a better understanding of the role that the NRG1, DAOA, and DISC1 genes play in the development of schizophrenia disorders.
Conclusion
Overall, this study preliminarily explored the roles of the NRG1 gene, DAOA gene, and DISC1 gene in the identification of UHR in a Chinese Han population. The results showed that these genes may be a little helpful in recognizing UHR. Future larger sample and longitudinal studies will be required to fully understand the roles of the above several genes in UHR recognition as well as transformation.
Acknowledgments
This work was supported by Guangdong Provincial Foundation for Basic and Applied Basic Research Natural Science Foundation (2022A1515010619) and Liuzhou Science and Technology Plan Project (2022CAC0113).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Modinos G, McGuire P. The prodromal phase of psychosis. Curr Opin Neurobiol. 2015;30:100–105. doi:10.1016/j.conb.2014.11.003
2. Fusar-Poli P. Why ultra high risk criteria for psychosis prediction do not work well outside clinical samples and what to do about it. World Psychiatry. 2017;16(2):212–213. doi:10.1002/wps.20405
3. Petersen L, Jeppesen P, Thorup A, et al. A randomised multicentre trial of integrated versus standard treatment for patients with a first episode of psychotic illness. BMJ. 2005;331(7517):602. doi:10.1136/bmj.38565.415000.E01
4. De Pablo GS, Radua J, Pereira J, et al. Probability of transition to psychosis in individuals at clinical high risk an updated meta-analysis. JAMA psychiatry. 2021;78(9):970–978. doi:10.1001/jamapsychiatry.2021.0830
5. Need AC, Goldstein DB. Schizophrenia genetics comes of age. Neuron. 2014;83(4):760–763. doi:10.1016/j.neuron.2014.08.015
6. Ni P, Zhou C, Liang S, et al. YBX1‐mediated DNA methylation‐dependent SHANK3 expression in PBMCs and developing cortical interneurons in schizophrenia. Adv Sci. 2023;10(20). doi:10.1002/advs.202300455
7. Stahl SM. Beyond the dopamine hypothesis to the NMDA glutamate receptor hypofunction hypothesis of schizophrenia. CNS Spectr. 2007;12(04):265–268. doi:10.1017/S1092852900021015
8. Li B, Woo RS, Mei L, Malinow R. The neuregulin-1 receptor erbB4 controls glutamatergic synapse maturation and plasticity. Neuron. 2007;54(4):583–597. doi:10.1016/j.neuron.2007.03.028
9. Mei L, Nave KA. Neuregulin-ERBB signaling in the nervous system and neuropsychiatric diseases. Neuron. 2014;83(1):27–49.
10. Namba T, Ming GL, Song H, Waga C, Uchino S. NMDA receptor regulates migration of newly generated neurons in the adult hippocampus via Disrupted-In-Schizophrenia 1 (DISC1). J Neurochem. 2011;71(1):e241.
11. Dahoun T, Trossbach S, Brandon N, Korth C, Howes O. The impact of Disrupted-in-Schizophrenia 1 (DISC1) on the dopaminergic system: a systematic review. Transl Psychiatry. 2017;7(1):e1015. doi:10.1038/tp.2016.282
12. Zhu MH, Liu ZJ, Hu QY, et al. Amisulpride augmentation therapy improves cognitive performance and psychopathology in clozapine-resistant treatment-refractory schizophrenia: a 12-week randomized, double-blind, placebo-controlled trial. Mil Med Res. 2022;9(1):1–13. doi:10.1186/s40779-022-00420-0
13. Hall J, Whalley HC, Job DE, et al. A neuregulin 1 variant associated with abnormal cortical function and psychotic symptoms. Nat Neurosci. 2006;9(12):1477–1478. doi:10.1038/nn1795
14. Kéri S, Kiss I, Seres I, Kelemen O. A polymorphism of the neuregulin 1 gene (SNP8NRG243177/rs6994992) affects reactivity to expressed emotion in schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2009;150b(3):418–420. doi:10.1002/ajmg.b.30812
15. Kiss I, Kelemen O, Kéri S. Decreased peripheral expression of neuregulin 1 in high-risk individuals who later converted to psychosis. Schizophr Res. 2012;135(1–3):198–199. doi:10.1016/j.schres.2011.12.012
16. Jagannath V, Gerstenberg M, Walitza S, et al. Neuregulin 1 (NRG1) gene expression predicts functional outcomes in individuals at clinical high-risk for psychosis. Psychiatry Res. 2018;266:143–146. doi:10.1016/j.psychres.2018.05.025
17. Mössner R, Schuhmacher A, Wagner M, et al. DAOA/G72 predicts the progression of prodromal syndromes to first episode psychosis. Eur Arch Psychiatry Clin Neurosci. 2010;260(3):209–215. doi:10.1007/s00406-009-0044-y
18. Ota VK, Moretti PN, Santoro ML, et al. Gene expression over the course of schizophrenia: from clinical high-risk for psychosis to chronic stages. NPJ Schizophr. 2019;5(1). doi:10.1038/s41537-019-0073-0
19. Rajan C. The psychosis-risk syndrome. handbook for diagnosis and follow-up. Psychiatrist. 2011;35(5):161–163. doi:10.1192/pb.bp.110.031831
20. Sheehan DV, Lecrubier Y, Sheehan KH, et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry. 1998;59(Suppl 20):22–33.
21. Kay SR, Opler LA, Lindenmayer JP. Reliability and validity of the positive and negative syndrome scale for schizophrenics. Psychiatry Res. 1988;23(1):99–110. doi:10.1016/0165-1781(88)90038-8
22. Montgomery SA, Åsberg M, Montgomery SA, Asberg M. A new depression scale designed to be sensitive to change. Br J Psychiatry. 1979;134(4):382. doi:10.1192/bjp.134.4.382
23. Liu H, Zhang H, Xiao W, et al. Scales for evaluating depressive symptoms in Chinese patients with schizophrenia. J Nerv Ment Dis. 2009;197(2):140. doi:10.1097/NMD.0b013e31819636a5
24. Hall RCW. Global assessment of functioning. A modified scale. Psychosomatics. 1995;36(3):267–275. doi:10.1016/S0033-3182(95)71666-8
25. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. 2006;22(15):1928–1929. doi:10.1093/bioinformatics/btl268
26. Trikalinos TA, Salanti G, Khoury MJ, Ioannidis JP. Impact of violations and deviations in Hardy-Weinberg equilibrium on postulated gene-disease associations. Am J Epidemiol. 2006;163(4):300–309. doi:10.1093/aje/kwj046
27. Kéri S, Kiss I, Kelemen O. Effects of a neuregulin 1 variant on conversion to schizophrenia and schizophreniform disorder in people at high risk for psychosis. Mol Psychiatry. 2009;14(2):118–119. doi:10.1038/mp.2008.1
28. Bousman CA, Yung AR, Pantelis C, et al. Effects of NRG1 and DAOA genetic variation on transition to psychosis in individuals at ultra-high risk for psychosis. Transl Psychiatry. 2013;3(4):e251. doi:10.1038/tp.2013.23
29. Gong YG, Wu CN, Xing QH, Zhao XZ, Zhu J, He L. A two-method meta-analysis of Neuregulin 1(NRG1) association and heterogeneity in schizophrenia. Schizophr Res. 2009;111(1–3):109–114. doi:10.1016/j.schres.2009.03.017
30. Li D, Collier DA, He L. Meta-analysis shows strong positive association of the neuregulin 1 (NRG1) gene with schizophrenia. Hum Mol Genet. 2006;15(12):1995–2002. doi:10.1093/hmg/ddl122
31. Weickert CS, Tiwari Y, Schofield PR, Mowry BJ, Fullerton JM. Schizophrenia-associated HapICE haplotype is associated with increased NRG1 type III expression and high nucleotide diversity. Transl Psychiatry. 2012;2(4):e104. doi:10.1038/tp.2012.25
32. Chen J, Xu Y, Zhang J, et al. Genotypic association of the DAOA gene with resting-state brain activity in major depression. Mol Neurobiol. 2012;46(2):361–373. doi:10.1007/s12035-012-8294-5
33. Gomez L, Wigg K, Feng Y, et al. G72/G30 (DAOA) and juvenile-onset mood disorders. Am J Med Genet B Neuropsychiatr Genet. 2009;150b(7):1007–1012. doi:10.1002/ajmg.b.30904
34. Detera-Wadleigh SD, McMahon FJ. G72/G30 in schizophrenia and bipolar disorder: review and meta-analysis. Biol Psychiatry. 2006;60(2):106–114. doi:10.1016/j.biopsych.2006.01.019
35. Shi J, Badner JA, Gershon ES, Liu C. Allelic association of G72/G30 with schizophrenia and bipolar disorder: a comprehensive meta-analysis. Schizophr Res. 2008;98(1–3):89–97. doi:10.1016/j.schres.2007.10.004
36. Korostishevsky M, Kaganovich M, Cholostoy A, et al. Is the G72/G30 locus associated with schizophrenia? Single nucleotide polymorphisms, haplotypes, and gene expression analysis. Biol Psychiatry. 2004;56(3):169–176. doi:10.1016/j.biopsych.2004.04.006
37. Law AJ, Lipska BK, Weickert CS, et al. Neuregulin 1 transcripts are differentially expressed in schizophrenia and regulated by 5’ SNPs associated with the disease. Proc Natl Acad Sci U S A. 2006;103(17):6747–6752. doi:10.1073/pnas.0602002103
38. Tan W, Wang Y, Gold B, et al. Molecular cloning of a brain-specific, developmentally regulated neuregulin 1 (NRG1) isoform and identification of a functional promoter variant associated with schizophrenia. J Biol Chem. 2007;282(33):24343–24351. doi:10.1074/jbc.M702953200
39. Hartz SM, Ho BC, Andreasen NC, Librant A, Wassink TH. G72 influences longitudinal change in frontal lobe volume in schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2010;153B(2):640–647. doi:10.1002/ajmg.b.31033
40. Schultz CC, Koch K, Wagner G, et al. Psychopathological correlates of the entorhinal cortical shape in schizophrenia. Eur Arch Psychiatry Clin Neurosci. 2010;260(4):351–358. doi:10.1007/s00406-009-0083-4
41. Hu GQ, Yang CQ, Lv QY, Zhao J, Yi ZH. Correlation between the polymorphism of locus rs2391191 of DAOA gene and the schizophrenia of Chinese Han patients. J Shanghai Jiaotong Univ. 2015;35(10):1443.
42. Jagannath V, Theodoridou A, Gerstenberg M, et al. Prediction analysis for transition to Schizophrenia in individuals at clinical high risk for psychosis: the relationship of DAO, DAOA, and NRG1 variants with negative symptoms and cognitive deficits. Front Psychiatry. 2017;8:292. doi:10.3389/fpsyt.2017.00292
43. Santoro ML, Gadelha A, Ota VK, et al. Gene expression analysis in blood of ultra-high risk subjects compared to first-episode of psychosis patients and controls. World J Biol Psychiatry. 2015;16(6):441–446. doi:10.3109/15622975.2015.1048724
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.