Back to Journals » Cancer Management and Research » Volume 13
Further Understanding of the Immune Microenvironment in Head and Neck Squamous Cell Carcinoma: Implications for Prognosis
Authors Denaro N, Merlano MC ![]() , Lo Nigro C
, Lo Nigro C
Received 28 January 2021
Accepted for publication 15 April 2021
Published 17 May 2021 Volume 2021:13 Pages 3973—3980
DOI https://doi.org/10.2147/CMAR.S277907
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eileen O'Reilly
Nerina Denaro,1 Marco Carlo Merlano,1 Cristiana Lo Nigro1,2
1Medical Oncology, S. Croce & Carle Teaching Hospital, Cuneo, Italy; 2Central Laboratory, Galliera Hospital, Genoa, Italy
Correspondence: Cristiana Lo Nigro
Central Laboratory, Galliera Hospital, Via Mura Delle Cappuccine 14, Genoa, 16128, Italy
Tel +39 3176219417
Email [email protected]
Purpose: We aimed to review the literature on the tumor microenvironment as a key player in tumor growth and anti-cancer treatment responses in head and neck cancer.
Patients and Methods: We reviewed the recent literature on this topic, using the following research words: “tumor microenvironment” and “head and neck cancer or neoplasm or head and neck squamous cell carcinoma” and “immune cells” and “stromal cells”. A search was conducted on the PubMed website and reports from international meetings, presentations and abstracts.
Results: The tumor microenvironment is a complex network in which myeloid cells, tumoral cells, growth factors and cytokines are involved in angiogenesis, the extracellular matrix and epithelial-to-mesenchymal transition.
Conclusion: Immune resistance and rapid tumor growth depend on immunosuppressive and pro-tumoral environments. Further investigations to classify and adequately treat patients with head and neck cancer are required.
Keywords: head and neck squamous cell carcinoma, immune microenvironment, prognosis, treatment
Introduction
Head and neck squamous cell carcinoma (HNSCC) is a heterogeneous disease characterized by different anatomic origins and clinical behavior. In the past few decades, the group of human papilloma virus (HPV)-related cancer has been distinguished from non-virally induced cancer. However, among these groups, there are not yet any reliable molecular biomarkers for distinguishing different subgroups of patients at risk.1
The tumor microenvironment (TME) is increasingly being appreciated as an important determinant of cancer outcome, including in HNSCC.
There is no exhaustive definition of TME, as it involves lymphocytes, fibroblasts, neutrophils, eosinophils, macrophages and myeloid-derived suppressor cells, as well as stromal cells and tumoral cells. These cells interact tightly and dynamically, and the balance of proangiogenic factors, tissue pH, growth factors and cytokine production changes over time.2
The TME is the result of factors associated with cancer (such as hypoxia, necrosis, mutational load and tumor mutation burden [TMB]) associated with the patient’s characteristics (age, viral infection, eg cytomegalovirus [CMV], HPV, Epstein–Barr virus [EBV], smoking, lifestyle, sex, previous or concurrent medication) and immune system/inflammation (eg chronic, acute or no inflammation; CD4+/CD8+ ratio; T-cell exhaustion) and factors related to geographic origin (ie microbioma, human leukocyte antigen [HLA] polymorphisms). Moreover, the TME may be influenced by anti-cancer treatment (ie chemotherapy: gemcitabine reduces myeloid cells, but has no effect on cytotoxic cells; cyclophosphamide at low doses reduces T-regulatory cells [T-reg], taxanes reduce myeloid cells, interleukin-1 [IL-1] and tumor necrosis factor-β [TNF-β]; radiotherapy).2
To date, most studies on the predictive markers for response to immunotherapy have focused on protein programmed death ligand-1 (PD-L1) and the TMB, rather than the environment in which the tumor proliferates. However, even though there is a correlation between the high expression of PD-L1 and response, as well as high TMB and response, several other mechanisms can impair the immune response.3
Recent studies have shown that the TME has a more profound influence on the growth and metastatic spread of HNSCC than was previously appreciated.
In HNSCC, tumor–stromal cross-talk is a key step in the pathogenesis of HNSCC. Cancer-associated fibroblasts (CAFs), chemokines, cytokines, and proliferative and inflammatory signal pathways are the major players in this complex network. An active tumor–stromal cross-talk is essential to promote cancer growth and metastization. Pro-tumoral cytokines (eg IL-10 and IL-1) induce interferon-γ (IFN-γ) downregulation, stimulate matrix metalloproteinases [MMPs] (increasing metastatic tumor-cell escape) and participate in angiogenesis.4
The immune microenvironment is of special interest for its possible implications. Little is known about the characteristics of patients who are more likely to experience a long-term benefit from immune checkpoint inhibitor (ICI) therapy.
Figure 1 summarizes the actors involved in TME regulation.
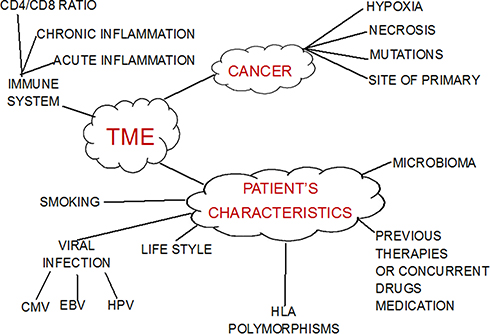 |
Figure 1 Actors in the TME. |
In this review, we address the role of the TME in HNSCC, evaluating, in particular:
- TME cells: myeloid cells, tumor-associated macrophages (TAMs), CAFs;
- neoangiogenesis and fibrosis/stroma and the extracellular matrix (ECM);
- immune signature;
- prognosis and treatment.
TME Cells: Myeloid Cells
Myeloid-derived suppressor cells (MDSCs) are innate immune cells, with the ability to suppress immune anti-tumoral responses (Figure 2). They represent a heterogeneous cell population of different origins. In the first steps of carcinogenesis, the presence of myeloid cells is associated with increased tumor-infiltrating lymphocytes (TILs), lower rate of metastases, less recurrence and improved survival in patients with HNSCC. Conversely, MDSCs in the TME of HNSCC and higher disease stage are associated with a worse prognosis.5
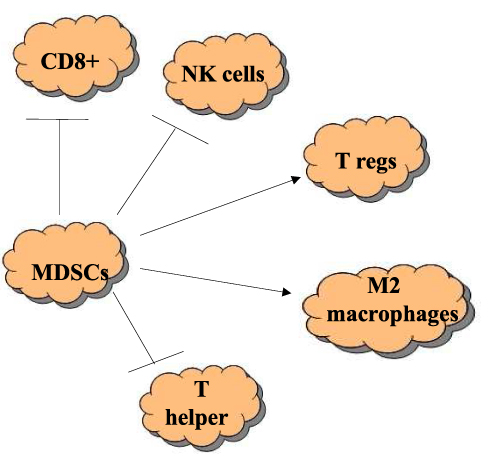 |
Figure 2 Function of MDSCs. Suppression of CD8+, NK cells, inhibition of humoral cells, differentiation into M2 macrophages, induction of differentiation and expansion of T-regs. |
MDSCs promote the formation of T-regs, induce the maturation of TAMs and promote the differentiation of fibroblasts into CAFs.5
T-regs are T cells which have a role in regulating or suppressing other cells in the immune system. They suppress the activation, proliferation and cytokine production of CD4+ T cells and CD8+ T cells, and are thought to suppress B cells and dendritic cells. T-regs can produce soluble messengers which have a suppressive function, including transforming growth factor-β (TGF-β), IL-10 and adenosine.
MDSCs are associated with chronic inflammation. In the inflamed microenvironment, several cytokines (TNF, IL-1, TGF-β and IL-10) subvert the patient’s immune system, inducing the proliferation and differentiation of myeloid precursors.6,7
Moreover, an efficient MDSC stimulates T cells but at the same time induces the secretion of cytokines IL-1 and IL-10. Several studies have reported high levels of MDSCs in the TME independently of HPV status and high levels of PD-L1.8–10
MDSCs are important for the innate immune response and express proteins involved in pathogen recognition called Toll-like receptors (TLRs). TME condition TLR7 but no significant alterations in the expression of other TLRs seem to be related to the TME.11
High TLR5 and low TLR7 expression were associated with poor disease-specific survival in HPV-positive oropharyngeal squamous cell carcinoma (OPSCC) patients.12
Myeloid-derived suppressor cells are immature cells that acquire new competences during their maturation, such as dendritic cells, which are abundant in the immunosuppressive TME. Among MDSCs, three subsets are recognized: monocytic (M-MDSC), granulocytic polymorphonuclear (PMN-MDSC) and early stage (e-MDSC), which lacks the myeloid lineage markers of the first two subsets.13
PMN-MDSCs and M-MDSCs are associated with T-cell suppression, not only in preclinical studies but also in human cancer patients.
An abundance of PMN-MDSCs is associated with poor survival and T-cell exhaustion. In vitro and in vivo assays have demonstrated they are the most potent suppressors of T-cell proliferation and reduce IFN-γ production. Arginase production by PMN-MDSC correlates with their maturation and immunosuppressive activity.13
TME Cells: TAMs
The role of TAMs in HNSCC is controversial. Macrophages possess a huge plasticity and diversity, and the binary classification anti-tumoral M1 and pro-tumoral M2 is an outdated concept.14
On the one hand, Mantovani et al confirmed that TAMs produce growth factors and cytokines promoting cancer cell proliferation but, on the other hand, preclinical evaluations showed that a reduction in TAMs did not affect tumor cell proliferation.15,16
TAMs in the TME cooperate with cancer stem cells (CSCs) to maintain stemness. In HNSCC, the activation of PI3K-4EBP1-SOX2 regulates some CSC properties and it may increase fibrosis/stroma/ECM escape. Gomez et al assessed the correlations among CSC markers and this proliferative pathway in HNSCC. They established a mechanistic link between TAMs and CSCs in response to extracellular signals.17
Targeting the reprogramming of TAMs toward M1-like macrophages has been investigated as a strategy to reverse the immunosuppressive TME.18
TME Cells: CAFs
CAFs are activated fibroblasts found in the TME. Specific markers include fibroblast activation protein-α (FAP-α) and α-smooth muscle actin (α-SMA), which separate them from the larger pool of fibroblasts present in the body.
CAFs are derived from mesenchymal stem cells. Resident fibroblasts are recruited by chemokines/cytokines and growth factors to specific sites, and ECM components at these sites turn these cells into CAFs.19
CAFs represent the fibroblasts’ response to TME stimulation. These activated fibroblasts found in association with cancer are termed CAFs, but also tumor-associated fibroblasts, peritumoral fibroblasts, myofibroblasts or reactive stromal fibroblasts. Activation of these CAFs coevolves with tumor progression as they act to survive in and populate the complex TME and contribute to tumor evolution.20
CAFs organize collagen fibrils in the ECM, providing the way for invasion and metastization, dependent on either CAFs or epithelial-to-mesenchymal transition (EMT).
The increased fibrinogenic and hyaluronic metabolism of the TME instructs carcinoma initiation and dissemination by performing multiple functions.
CAFs can be detected in the peripheral blood of patients with solid tumors (breast, colon and prostate) but not in healthy volunteers.21 They move with circulating tumor cells and are strongly associated with these cells.
CAFs express angiotensin-II (Ang-II) receptor ATR1.22 Ang-II stimulates CAF proliferation and the production of immunosuppressive cytokines, and strongly contributes toward promoting a desmoplastic TME.23
Ang-II contributes to resistance to immunotherapy through the recruitment of inflammatory cells and CAFs. Xie et al demonstrated, in two cancer mouse models, that Ang-II signaling blockade reversed the immunosuppressive TME, and inhibition of the expression of angiotensinogen (AGT), a precursor of Ang-II, strongly triggered an immune-activating cytokine profile in hypoxic mouse cancer cells. In addition, AGT silencing combined with a checkpoint blockade generated an abscopal effect in resistant tumors, stimulating the proliferation of dendritic cells and T cells.24
A combination of inhibitors of Ang-II and PD-1 axis overcame resistance to ICIs in experimental models.25
Studies are ongoing with Ang-II receptor blockers in combination with immunotherapy.
Neoangiogenesis and Fibrosis/Stroma and the ECM in HNSCC
Angiogenesis is a main step in tumor proliferation and metastization. Without new vessels, neoplasms are unable to spread. As in other solid tumors, intratumoral hypoxia is associated with a high proliferative growth index and accumulation of CSCs in the TME.
Up to 90% of HNSCCs highly express angiogenesis factors such as vascular endothelial growth factor (VEGF).26
Moreover, high levels of pro-inflammatory cytokines induce STAT3 activation, which, in turn, regulates the anti-inflammatory response through increased release of IL-10, TGF-β, VEGF, hypoxia inducible factor-1α (HIF-1α) and cyclin D1/D2.27
In the late 1990s, Gallo et al showed that tumor vascularization is dependent on increased endothelial nitric oxide synthase (NOS) activity and cyclic guanosine monophosphate (cGMP) levels. Microvessel density and NOS activity correlate with lymph-node metastases.27
Additional hypoxia and nitric oxide (NO) influence inhibitor of apoptosis protein (IAP), as it has been demonstrated that NO reduces survival. The expression of IAPs increases in response to NO and cytokines. In contrast, X-chromosome-linked inhibitor of apoptosis (XIAP) mRNA levels remained unaffected by cytokines and NO. Activated HIF-1 transcription factor binds to the hypoxia response elements (HREs) to induce the transcription of genes involved in cancer proliferation/survival, angiogenesis, glucose metabolism and invasion/metastasis.
Hypoxia stimulates IL-6, the migration of MDSCs and the recruitment of pro-tumoral macrophages. Both innate and adaptive immune responses are impaired under hypoxic conditions: on the one hand, interferon transcription is reduced and, on the other hand, HLA affinity to tumoral antigens is attenuated (eg viral peptides). Numerous chemokines (CCL-17, CCL-28, TGF-β1) secreted by hypoxic tumors also attract T-regs to the TME.28
Hypoxia plays a critical role in the formation of a cold TME, especially by increasing the infiltration of CAFs and TAMs in hypoxic tumor areas.24
Several HIF prolyl hydroxylase inhibitors are under investigation not only as anti-cancer therapies but also for hematological and chronic vascular diseases. Bommi et al reported that damaged DNA binding protein-2 (DDB2), a nucleotide excision repair protein upregulated by hypoxia, inhibits HIF-1α in HNSCC cells. It inhibits HIF-1α in both normoxia and hypoxia by reducing mRNA expression. Preclinical models with knockdown of DDB2 showed enhanced angiogenesis and tumor growth.29
DDB2 expression is downregulated in advanced HNSCCs, and loss of DDB2 expression coincides with reduced survival. This may depend both on angiogenesis/tumor proliferation and on EMT.
DDB2 constitutively represses mRNA expression of the EMT-regulatory transcription factors SNAIL and ZEB1, and the angiogenic factor VEGF in HNSCC cells.30
Several other ways to target angiogenesis are under investigation: ligand-directed antibodies (against angiotensin), receptor-directed antibodies (anti-VEGF), small molecule inhibitors (multi-target kinase inhibitor) and others.
The first drug studied was the anti-VEGF inhibitor bevacizumab. Argiris et al showed in a phase III randomized trial (E1305) that adding bevacizumab to chemotherapy increased the response rate and prolonged progression-free survival, but did not significantly improve survival.31
Multi-target kinase inhibitors (against vascular endothelial growth factor receptors [VEGFRs], epidermal growth factor receptors [EGFRs], fibroblast growth factor receptors [FGFRs] and platelet-derived growth factor receptors [PDGFRs]) were tested in HNSCC, but with limited benefit.26
Immune Signature in HNSCC
Gene expression signatures for survival stratification in HNSCC patients have been proposed in various studies. In Table 1, we report the most remarkable studies on this topic. The first data set to provide a comprehensive landscape of somatic genomic alterations in HNSCC was The Cancer Genome Atlas (TCGA HNSCC), which showed HPV-associated mutations, smoking-related alterations and other distinct subgroups (with therapeutic candidate targets). Somatic mutations such as TP53, CDKN2A, PIK3CA, HRAS, FBXW7 and NOTCH1 genes are observed in HNSCC tumors.32
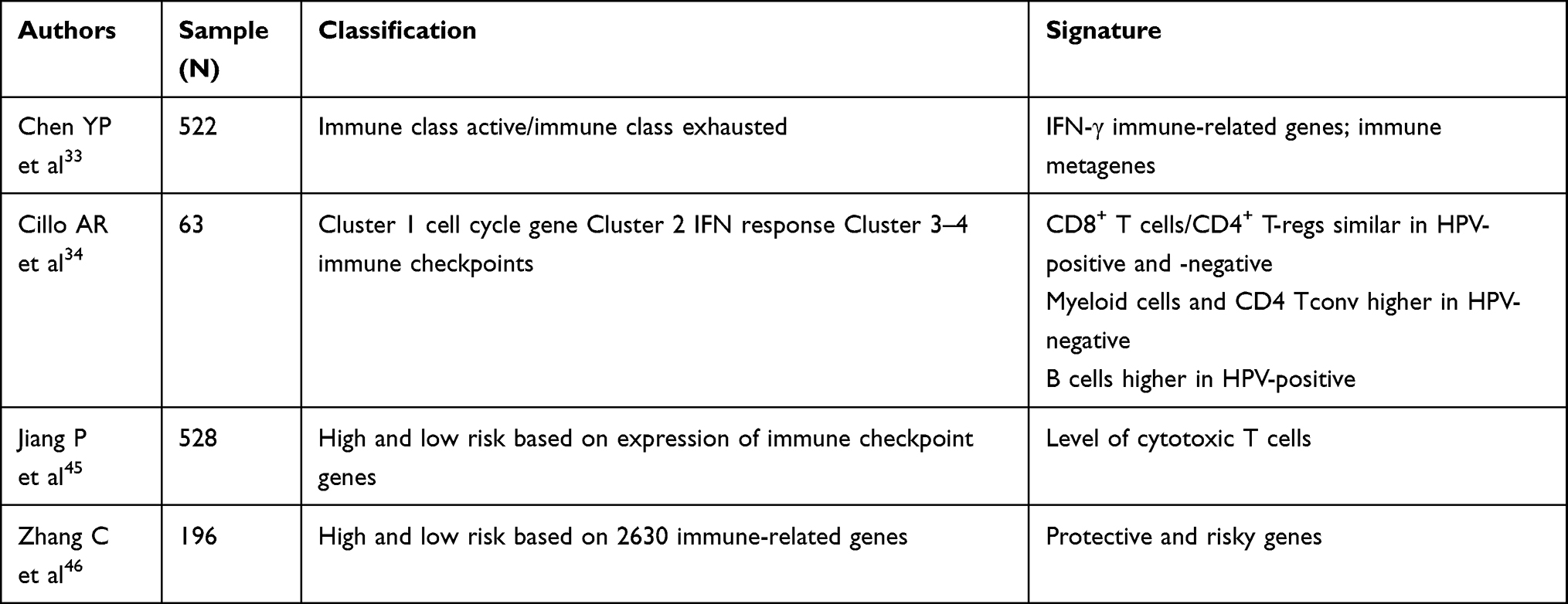 |
Table 1 Gene Expression Signatures for Survival Stratification in HNSCC Patients |
Chen et al sequenced the RNA of 522 HNSCC samples from TCGA. They found that 40% of patients had a new molecular immune phenotype called Immune Class. Patients with Immune Class showed significant enrichment of signatures identifying immune cells and enrichment of the six-gene IFN signature.33
It was found that 27% of Immune Class (56/211) was characterized by the previously reported activated stromal gene signature that captures the activated inflammatory stromal, low expression of B-cells cluster, significantly associated with M2 (anti-inflammatory) macrophages and other immunosuppressive components, eg the WNT/β catenin. This subgroup of Immune Class exhaustion is immunoresistant. Conversely, in patients lacking the activated stroma signature, high expression of the pro-inflammatory M1 macrophage signature was reported by Chen et al.33
Unfortunately, signatures relating to the immune enrichment score and IFN signaling did not differ between the subgroups, and further investigations are required to individualize the right approach in these patients.
Among immune profiles, immune cells within tumors of HPV-negative and HPV-positive HNSCC displayed a spectrum of transcriptional signatures, with helper CD4+ T cells and B cells being relatively divergent and CD8+ T cells and CD4+ regulatory T cells being relatively similar.34
However, the enthusiasm for immunotherapy strategies in HNSCC has led researchers to look for predictors in the TME and to obtain an immune signature. A construction and validation of an individualized prognostic signature based on immune-related genes was proposed by She et al.35 In this immune signature, 27 genes were included, such as UL16-binding protein 1 (ULBP1), chemokine receptors 6 (CCR6), C-C motif chemokine ligand 22 (CCL22), roundabout guidance receptor 1 (ROBO1), dickkopf WNT signaling pathway inhibitor 1 (DKK1) and platelet-derived growth factor subunit A (PDGFA), all of which have previously been shown to be correlated with the pathogenesis and progression in HNSCC.
Moreover, as already known, pathways involved in hypoxia, EMT, proliferation and escape are important in HNSCC (eg JAK/STAT pathway, JAK/PI3K/AKT and SHP2/MAPK signaling pathways).
Several studies are ongoing to assess the predictive role of cytokines (in particular, IFN-α, IFN-γ, IL-2, IL-6 and TGF-β) and chemokines, as well as genes involved in proliferation and immune pathways.
Among HNSCC patients treated with immunotherapies, only 20% responded and about one in ten had a response longer than one year. The goal of adequate classification of the immune microenvironment is to predict those patients who require more aggressive therapies, such as combination immune and chemotherapies.2
A randomized phase III trial, KEYNOTE048, demonstrated that pembrolizumab plus platinum and 5-fluorouracil (5-FU) is an appropriate first line therapy, but in those patients with PD-L1 combined prognostic score (CPS) >20, pembrolizumab alone is also an option.36
Wang et al validated the stromal and immune scores as predictors. A six-gene signature was selected and displayed a robust predictive effect. The expressions of key genes were associated with immune infiltration. Genetic validation is ongoing. The dataset GSE65858 confirmed that gene expression patterns and TP53 mutations are associated with HPV RNA status, lymph-node metastasis and survival.37
Immune Metabolism
Immune metabolism critically influences the TME (Figure 3). How a dysregulated metabolism in immune cells contributes toward suppressing effector functions is an area of active investigation.38 Both tumor cells and viral infection can cause depletion of glucose levels in the microenvironment. The main feature of the TME is the enhanced glycolytic flux that increases lactate production, inducing both M2 polarization of TAMs and suppression of natural killer (NK) cell function.38
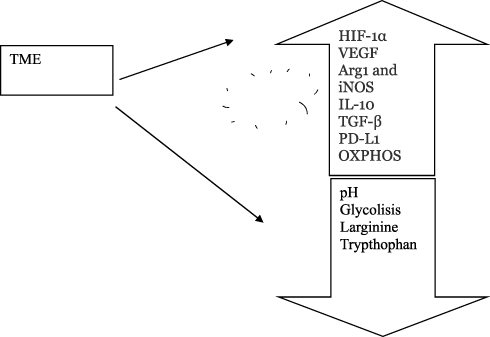 |
Figure 3 Immune metabolism and tumor microenvironment. Markers of the TME include hypoxia, low pH, oxidative stress, inhibition of apoptosis and immune mediators. Hypoxia induces upregulation of arginase 1, iNOS, IL-10, TGF-β and PD-L1. In the TME, oxidative phosphorylation (OXPHOS) is increased. |
Effector T cells are highly glycolytic, whereas memory T cells have an oxidative metabolism. Glutamine is also important, and effector T-cell differentiation is impaired when the supply of glutamine is disrupted. Moreover, the release into the TME of enzymes and cytokines such as arginase and indoleamine-2,3-dioxygenase further impairs T-cell functions. Mature myeloid cells tend to be non-proliferative and so have substantially different metabolic requirements, while pro-tumoral macrophages inactivate oxidative phosphorylation in an NOS-dependent manner.38 Persistent T-cell receptor stimulation during chronic infection results in less glycolysis and more fatty acid oxidation and tricarboxylic acid (TCA or Krebs) cycle usage.28
Amino acids are critical for cellular proliferation and protein biosynthesis, and competition between cancer cells and T cells can be a prime source of immunosuppression. In particular, glutamine and arginine have been shown to correlate with T-cell activation and survival.28
HPV-positive tumors undergo pronounced oxidative phosphorylation, with sequestration of lactic acid from the TME and conversion of the same to pyruvate. This pyruvate is utilized in the TCA, followed by oxidative phosphorylation, leading to higher generation of ATP molecules. Conversely, HPV-negative tumors have higher lactate generation by lactate dehydrogenase A, with higher levels in the microenvironment. The presence of lactate in the environment results in enhanced tumor aggressiveness.39
Prognosis and Treatment
The prognosis is influenced by the TME; indeed, the immunosuppressive TME is considered the first cause of a lack of response to immunotherapy.2
A deeper knowledge of the TME is recommended to understand who and how to treat, ie the selection of the best therapeutic approach. Several studies have demonstrated that the TME evolves during cancer progression and it also changes with cancer treatments. Platinum-based induction chemotherapy or chemoradiotherapy increases PD-L1 expression on tumor and immune cells.40
Although the predictive role of the TME is widely accepted, to date we do not have a way to individually treat patients based on the TME. Moreover, in clinical practice, only the checkpoint inhibitors pembrolizumab (for first and second line) and nivolumab (second line only) can be prescribed. In the first line, a combination of immunotherapy and chemotherapy is recommended when the CPS is low, the disease is rapidly growing and prognostic factors are unfavorable.36
Immunotherapy as monotherapy achieves a response rate of around 20%, with less than 20% long responders at 2 years.2 Therefore, the results of immunotherapy need to be improved.
Many trials have explored the role of innate immunity with drugs targeting TLRs. However, TLR agonists have failed to improve outcomes.41
A hypothesis to explain these results is the low intratumoral dose obtained with subcutaneous or intravenous injection.
Among new TME targets, IAPs appear of great interest.42
Combinations of immunotherapy and chemotherapy or target therapies are being widely investigated to overcome resistance to immunotherapy.
Anti-EGFR in combination with immunotherapy is interesting for the biological background. Trials are ongoing investigating this approach, including a combination of avelumab, cetuximab and palbociclib.42
A promising approach is to target the angiogenesis with multi-target kinase inhibitors and pembrolizumab (study LEAP-0.10; NCT04199104), or with angiopoietin inhibitors and immunotherapy. The rationale of this approach derives from the effect of anti-angiogenic therapies, which increase T-cell infiltration into tumors, reducing the immunosuppressive response and thereby overcoming resistance to checkpoint inhibitors.43
Otherwise, tumors with a favorable TME may benefit from immunotherapy as monotherapy (alone in both first and second line treatment, or with nivolumab in the second line setting).
However, the role of immunotherapy in locally advanced settings is controversial following negative results reported in the JAVELIN 100 trial, which failed to demonstrate an advantage of chemoradiotherapy plus avelumab and avelumab maintenance. Studies are ongoing with pembrolizumab (KEYNOTE-412, KEYNOTE-689) and nivolumab (NIVOPOSTOP/NICO) both as concurrent/maintenance or neoadjuvant therapy and in combination with standard-of-care adjuvant therapy for resectable HNSCC.
A mechanism under investigation aims to elicit the maximum benefit from radiotherapy. It has been hypothesized that hypofractionation may be useful in combination with checkpoint inhibitors. Trial NCT03085719 is investigating low versus high radiotherapy doses in this setting.44
Conclusions
The heterogeneity of the TME is highly correlated with tumor behavior and therapeutic response. Neoangiogenesis, fibrosis/stromal components and myeloid cells are the main components of the TME.
The aim of future personalized therapies should be to target the TME, in particular the factors associated with immunosuppression that reduce the therapeutic efficacy.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Merlano MC, Denaro N, Galizia D, Garrone O. How I treat squamous ENT cancer. ESMO Open. 2019;4(Suppl 2):e000542. doi:10.1136/esmoopen-2019-000542
2. Economopoulou P, Kotsantis I, Psyrri A. Tumor microenvironment and immunotherapy response in head and neck cancer. Cancers. 2020;12(11):3377. doi:10.3390/cancers12113377
3. Chalmers ZR, Connelly CF, Fabrizio D, et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med. 2017;9(1):34. doi:10.1186/s13073-017-0424-2
4. Pries R, Wollenberg B. Cytokines in head and neck cancer. Cytokine Growth Factor Rev. 2006;17(3):141–146. doi:10.1016/j.cytogfr.2006.02.001
5. Veglia F, Perego M, Gabrilovich D. Myeloid-derived suppressor cells coming of age. Nat Immunol. 2018;19(2):108–119. doi:10.1038/s41590-017-0022-x
6. Brocks CP, Pries R, Frenzel H, Ernst M, Schlenke P, Wollenberg B. Functional alteration of myeloid dendritic cells through head and neck cancer. Anticancer Res. 2007;27:817–824.
7. Tesi RJ. MDSC; the Most Important Cell You Have Never Heard Of. Trends Pharmacol Sci. 2019;40(1):1. doi:10.1016/j.tips.2018.10.008
8. Yu GT, Bu LL, Huang CF. PD-1 Blockade Attenuates Immunosuppressive Myeloid Cells Due to Inhibition of CD47/SIRPα Axis in HPV Negative Head and Neck Squamous Cell Carcinoma. Oncotarget. 2015;6:42067–42080. doi:10.18632/oncotarget.5955
9. Russell S, Angell T, Lechner M, et al. Immune Cell Infiltration Patterns and Survival in Head and Neck Squamous Cell Carcinoma. Head Neck Oncol. 2013;5:24.
10. Partlová S, Bouček J, Kloudová K, et al. Distinct Patterns of Intratumoral Immune Cell Infiltrates in Patients with HPV-Associated Compared to Non-Virally Induced Head and Neck Squamous Cell Carcinoma. Oncoimmunology. 2015;4(1):e965570. doi:10.4161/21624011.2014.965570
11. Frenzel H, Hoffmann B, Brocks C, Schlenke P, Pries R, Wollenberg B. Toll-like receptor interference in myeloid dendritic cells through head and neck cancer. Anticancer Res. 2006;26(6B):4409–4413.
12. Jouhi L, Mohamed H, Mäkitie A, et al. Toll-like receptor 5 and 7 expression may impact prognosis of HPV-positive oropharyngeal squamous cell carcinoma patients. Cancer Immunol Immunother. 2017;66(12):1619–1629. doi:10.1007/s00262-017-2054-3
13. Lang S, Bruderek K, Kaspar C, et al. Clinical Relevance and Suppressive Capacity of Human Myeloid-Derived Suppressor Cell Subsets. Clin Cancer Res. 2018;24:4834–4844. doi:10.1158/1078-0432.CCR-17-3726
14. Chávez-Galán L, Olleros ML, Vesin D, Garcia I. Much More than M1 and M2 Macrophages, There are also CD169(+) and TCR(+) Macrophages. Front Immunol. 2015;6:263. doi:10.3389/fimmu.2015.00263
15. Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P. Tumour-associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol. 2017;14:399–416. doi:10.1038/nrclinonc.2016.217
16. Wu S, Dai X, Xie D. Identification and Validation of an Immune-Related RNA Signature to Predict Survival of Patients With Head and Neck Squamous Cell Carcinoma. Front Genet. 2019;10:1252. doi:10.3389/fgene.2019.01252
17. Gomez KE, Wu FL, Keysar SB, et al. Cancer Cell CD44 Mediates Macrophage/Monocyte-Driven Regulation of Head and Neck Cancer Stem Cells. Tumor Biol Immunol. 2020;80(19):54.
18. Wondergem NE, Nauta IH, Muijlwijk T, Leemans CR, van de Ven R. The immune microenvironment in head and neck squamous cell carcinoma: on subsets and subsites. Curr Oncol Rep. 2020;22(8):81. doi:10.1007/s11912-020-00938-3
19. Mishra PJ, Mishra PJ, Humeniuk R, et al. Carcinoma-associated fibroblast-like differentiation of human mesenchymal stem cells. Cancer Res. 2008;68:4331–4339. doi:10.1158/0008-5472.CAN-08-0943
20. Chen X, Song E. Turning foes to friends: targeting cancer-associated fibroblasts. Nat Rev Drug Discov. 2019;18(2):99–115. doi:10.1038/s41573-018-0004-1
21. Ortiz-Otero N, Clinch AB, Hope J, Wang W, Reinhart-King CA, King MR. Cancer associated fibroblasts confer shear resistance to circulating tumor cells during prostate cancer metastatic progression. Oncotarget. 2020;11(12):1037–1050. doi:10.18632/oncotarget.27510
22. Hinsley EE, de Oliveira CE, Hunt S, Coletta RD, Lambert DW. Angiotensin 1-7 inhibits angiotensin II-stimulated head and neck cancer progression. Eur J Oral Sci. 2017;125(4):247–257. doi:10.1111/eos.12356
23. Diop-Frimpong B, Chauhan VP, Krane S, Boucher Y, Jain RK. Losartan inhibits collagen I synthesis and improves the distribution and efficacy of nanotherapeutics in tumors. Proc Natl Acad Sci U S A. 2011;108(7):2909–2914. doi:10.1073/pnas.1018892108
24. Xie G, Cheng T, Lin J, et al. Local angiotensin II contributes to tumor resistance to checkpoint immunotherapy. J Immunotherapy Cancer. 2018;6(1):88. doi:10.1186/s40425-018-0401-3
25. Nakamura K, Yaguchi T, Ohmura G, et al. Involvement of local renin-angiotensin system in immunosuppression of tumor microenvironment. Cancer Sci. 2018;109(1):54–64. doi:10.1111/cas.13423
26. Denaro N, Russi EG, Colantonio I, Adamo V, Merlano MC. The role of antiangiogenic agents in the treatment of head and neck cancer. Oncology. 2012;83(2):108–116. doi:10.1159/000339542
27. Gallo O, Fini-Storchi I, Vergari WA, et al. Role of nitric oxide in angiogenesis and tumor progression in head and neck cancer. JNCI: J Nat Cancer Inst. 1998;90(8):587–596. doi:10.1093/jnci/90.8.587
28. Augustin RC, Delgoffe GM, Najjar YG. Characteristics of the tumor microenvironment that influence immune cell functions: hypoxia, oxidative stress, metabolic alterations. Cancers. 2020;12(12):3802. doi:10.3390/cancers12123802
29. Bommi PV, Chand V, Mukhopadhyay NK, Raychaudhuri P, Bagchi S. NER-factor DDB2 regulates HIF1α and hypoxia-response genes in HNSCC. Oncogene. 2020;39(8):1784–1796. doi:10.1038/s41388-019-1105-y
30. Bommi PV, Ravindran S, Raychaudhuri P, Bagchi S. DDB2 regulates Epithelial-to-Mesenchymal Transition (EMT) in Oral/Head and Neck Squamous Cell Carcinoma. Oncotarget. 2018;9(78):34708–34718. doi:10.18632/oncotarget.26168
31. Argiris A, Li S, Savvides P, et al. Phase III randomized trial of chemotherapy with or without bevacizumab in patients with recurrent or metastatic head and neck cancer. J Clin Oncol. 2019;37(34):3266–3274. doi:10.1200/JCO.19.00555
32. The Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature. 2015;517(7536):576–582. doi:10.1038/nature14129
33. Chen YP, Wang YQ, Lv JW, et al. Identification and validation of novel microenvironment-based immune molecular subgroups of head and neck squamous cell carcinoma: implications for immunotherapy. Ann Oncol. 2019;30:68–75. doi:10.1093/annonc/mdy470
34. Cillo AR, Kürten CHL, Tabib T, et al. Immune Landscape of Viral- and Carcinogen-Driven Head and Neck Cancer. Immunity. 2020;52:183–199. doi:10.1016/j.immuni.2019.11.014
35. She Y, Kong X, Ge Y, et al. Immune-related gene signature for predicting the prognosis of head and neck squamous cell carcinoma. Cancer Cell Int. 2020;20(1):22. doi:10.1186/s12935-020-1104-7
36. Burtness B, Harrington KJ, Greil R, et al. Pembrolizumab alone or with chemotherapy versus cetuximab with chemotherapy for recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-048): a randomised, open-label, Phase 3 study. Lancet. 2019;394(10212):1915–1928. doi:10.1016/S0140-6736(19)32591-7
37. Wang H, Wu X, Chen Y. Stromal-immune score-based gene signature: a prognosis stratification tool in gastric cancer. Front Oncol. 2019;9:1212. doi:10.3389/fonc.2019.01212
38. Kouidhi S, Ben Ayed F, Benammar Elgaaied A. Targeting Tumor Metabolism: a New Challenge to Improve Immunotherapy. Front Immunol. 2018;9:353. doi:10.3389/fimmu.2018.00353
39. Chaudhary S, Ganguly K, Muniyan S, et al. Imunometabolic Alterations by HPV Infection: new Dimensions to Head and Neck Cancer Disparity. J Natl Cancer Inst. 2019;111:233–244. doi:10.1093/jnci/djy207
40. Leduc C, Adam J, Louvet E, et al. TPF induction chemotherapy increases PD-L1 expression in tumour cells and immune cells in head and neck squamous cell carcinoma. ESMO Open. 2018;3:e000257. doi:10.1136/esmoopen-2017-000257
41. Ferris RL, Saba NF, Gitlitz BJ, et al. Effect of Adding motolimod to standard combination chemotherapy and cetuximab treatment of patients with squamous cell carcinoma of the head and neck: the active8 randomized clinical trial. JAMA Oncol. 2018;4(11):1583–1588. doi:10.1001/jamaoncol.2018.1888
42. Kwon M, Jung H, Nam GH, Kim IS. The right Timing, right combination, right sequence, and right delivery for Cancer immunotherapy. J Control Release. 2021;331:321–334. doi:10.1016/j.jconrel.2021.01.009
43. Rini BI, Powles T, Atkins MB, et al. Atezolizumab plus bevacizumab versus sunitinib in patients with previously untreated metastatic renal cell carcinoma (IMmotion151): a multicentre, open-label, phase 3, randomised controlled trial. Lancet. 2019;393(10189):2404–2415. doi:10.1016/S0140-6736(19)30723-8
44. Donlon NE, Power R, Hayes C, Reynolds JV, Lysaght J. Radiotherapy, immunotherapy, and the tumour microenvironment: turning an immunosuppressive milieu into a therapeutic opportunity. Cancer Lett. 2021;502:84–96. doi:10.1016/j.canlet.2020.12.045
45. Jiang P, Gu S, Pan D, et al. Signatures of T cell dysfunction and exclusion predict cancer immunotherapy response. Nat Med. 2018;24(10):1550–1558. doi:10.1038/s41591-018-0136-1
46. Zhang C, Luo Y, Zhang Z, et al. Identification of a prognostic immune signature for esophageal squamous cell carcinoma to predict survival and inflammatory landscapes. Front Cell Dev Biol. 2020;8. doi:10.3389/fcell.2020.580005
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.