Back to Journals » Infection and Drug Resistance » Volume 12
Functional Synergy Of Antimicrobial Peptides And Chlorhexidine Acetate Against Gram-Negative/Gram-Positive Bacteria And A Fungus In Vitro And In Vivo
Authors Zhu J, Huang Y ![]() , Chen M, Hu C, Chen Y
, Chen M, Hu C, Chen Y ![]()
Received 8 June 2019
Accepted for publication 25 September 2019
Published 15 October 2019 Volume 2019:12 Pages 3227—3239
DOI https://doi.org/10.2147/IDR.S218778
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Jie Zhu,1,2 Yibing Huang,1,2 Mingxia Chen,3 Cuihua Hu,4 Yuxin Chen1–3
1Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, Jilin University, Changchun, People’s Republic of China; 2College of Life Sciences, Jilin University, Changchun, People’s Republic of China; 3Jiangsu ProteLight Pharmaceutical & Biotechnology Co., Ltd., Jiangyin, People’s Republic of China; 4International Research Centre for Nano Handling and Manufacturing, Changchun University of Science and Technology, Changchun, People’s Republic of China
Correspondence: Yuxin Chen
Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, Jilin University, No. 2699, Qianjin St., Changchun 130021, People’s Republic of China
Tel/fax +86 431 8515 5200
Email [email protected]
Background and purpose: To reduce the resistance and allergic reaction to chlorhexidine acetate (CHA) in the current treatment of (Bacterial vaginosis) BV and (vulvovaginal candidiasis) VVC in female vaginitis. In this study, the antimicrobial activities and mechanism of action of the synergistic effects of antimicrobial peptides (AMPs) HPRP-A1 and HPRP-A2, and CHA, against Gram-negative and Gram-positive bacteria, and one fungus Candida albicans (C. albicans) were investigated in vitro and in mouse and rat vaginitis infection models in vivo.
Results: HPRP-A1, HPRP-A2 and CHA showed significant synergistic effects on the antimicrobial activities against different Gram-negative and Gram-positive bacteria and C. albicans. The combined application of HPRP-A2 and CHA exhibited strong synergistic effects in the mouse and rat vaginitis models caused by bacteria or C. albicans.
Conclusion: This study may prompt the development of new drug combinations against vaginitis infections, including mixed bacterial and fungal infections and multi-drug-resistant infections.
Keywords: antimicrobial peptide, chlorhexidine acetate, antibacterial, antifungal, vaginitis
Introduction
Bacterial vaginosis (BV) and vulvovaginal candidiasis (VVC) are common mucosal infections that affect a large percentage of women, some of whom experience recurrent infections that are usually refractory.1–4 BV is a disease characterized by partial loss of the indigenous vaginal lactobacilli and vaginal epithelium anaerobic overgrowth. VVC is caused by the overgrowth of Candida species, mainly Candida albicans.5,6 Clinical prescription drugs for vaginitis are nystatin polyenes.7 There are some studies showing the non-inferiority of chlorhexidine (CHX) biguanide compared with metronidazole or clindamycin.8 These drugs have a short-term therapeutic effect, in addition drug resistance can occur. Thus, alternative treatment approaches and new pharmacological targets are being considered as the main therapeutic strategies to prevent and treat these types of infections.9,10
CHX is a bisbiguanide disinfectant and preservative with a broad spectrum of antibacterial activity against Gram-negative and Gram-positive bacteria. It is used for infection controls, including systemic lavage, oral cleansing and surgical hand washing in intensive care units.11 CHX attaches to the negatively charged bacterial membrane, causing the outer leaflet to break. High concentrations of CHX can severely damage the cytoplasmic membrane and cause cell lysis. At low concentrations, near the minimal inhibitory concentration (MIC), CHX distorts the cell wall of Gram-positive and Gram-negative bacteria, resulting in morphological changes on the cell surface.12,13 CHX is easily available and inexpensive, and its pH is suitable for the human vaginal environment.14,15 In addition, CHX has no tissue-dissolving activity.16 Therefore, CHX is considered a suitable compound for intravaginal treatment. However, high concentrations of CHX can irritate the mucosa.17 In addition, some patients may have allergic reactions.18
HPRP-A1 is an α-helical antimicrobial peptide composed of 15 L-amino acid residues, which was derived from the ribosomal L1 (RpL1) N-terminal sequence of Helicobacter pylori isolated from the gastric mucosa of patients with gastropathy.19 It has wide-spectrum antimicrobial effects against Gram-negative and Gram-positive bacteria, and fungi,20,21 due to its membrane-disrupting ability. Previous studies have shown that HPRP-A1 is a peptide targeting on the bilayer of cell membranes.22,23 The specificity of α-helical AMPs to prokaryotic and eukaryotic cells depends on the composition of membrane phospholipids. For eukaryotes, which are dominated by neutral phospholipids and contain large amounts of cholesterol and sphingomyelin, peptides form pores in the hydrophobic core of the lipid bilayers, leading to the death of eukaryotic cells. For prokaryotic cells characterized by strong transmembrane potential, peptides act as detergent agents to decompose cell membranes as in “carpet model”.24–26 It is reported that the HPRP-A2 peptide that contains all D-amino acids is more stable than the HPRP-A1 peptide that contains all L-amino acids due to resistance against proteolysis in the plasma and tissue fluid.22,27
Although the combination of antimicrobial peptides (AMPs) and traditional antibiotics has been widely reported, the synergistic effect of AMPs and the disinfectant CHA has not been studied to date. In this study, the synergistic effect and mechanism of action of AMPs HPRP-A1/HPRP-A2, and CHA were investigated. Three Gram-negative bacteria Escherichia coli (E. coli), Pseudomonas aeruginosa (P. aeruginosa), Klebsiella pneumonia (K. pneumonia), three Gram-positive bacteria Staphylococcus aureus (S. aureus), Staphylococcus epidermidis (S. epidermidis), Bacillus subtilis (B. subtilis) and one fungus Candida albicans (C. albicans) were used to evaluate the synergistic effects between HPRP-A1, HPRP-A2 and CHA in vitro. The combined application of HPRP-A2 and CHA was also studied on mouse and rat vaginitis models caused by bacteria and a fungus, respectively, in vivo. We believe that the combined application of AMPs and CHA may be a promising approach for the treatment of BV and VVC in clinical practices.
Materials And Methods
Materials
Agar, tryptone and yeast extract were obtained from Oxoid (Ogdensburg, USA). Sabouraud dextrose (SD) broth was obtained from the Beijing Shuangxuan microbial culture medium products factory (Beijing, China). Acetonitrile (HPLC grade) was obtained from Fisher (Beijing, China). BBcell ProbeTM live/dead bacterial staining kit was obtained from BestBio (Shanghai, China). Sytox green dead nucleic acid stain was obtained from Biorab (Beijing, China). Reactive oxygen species (ROS) assay kits were provided by Solarbio Life Science (Beijing, China). DL 2000 DNA Marker was obtained from Takara Biomedical Technology Co., Ltd. (Beijing, China). Lipopolysaccharides (LPS) from E. coli 055: B5 was purchased from Sigma-Aldrich Co. (St. Louis, USA). Chlorhexidine acetate (CHA) was obtained from Aladdin Reagents Co. (Shanghai, China). Peptides were supplied by Jiangsu ProteLight Pharmaceutical and Biotechnology Co. Ltd. (Wuxi, China). Rink amide MBHA was obtained from Tianjin Nankai Hecheng Science & Technology Co., Ltd. (Tianjin, China). All of the N-α-Fmoc protected amino acids and coupling reagents for peptide synthesis were obtained from GL Biochem (Shanghai, China).
Bacterial Strains And Animals
The Gram-negative bacterial strains E. coli ATCC 25922, P. aeruginosa ATCC 27853 and K. pneumonia ATCC 700603, the Gram-positive bacterial strains S. aureus ATCC 25923, S. epidermidis ATCC 12228 and B. subtilis ATCC 6633 were purchased from the American Type Culture Collection (Manassas, VA, USA), and the fungal strain C. albicans JLC 30364 was purchased from the Center of Disease Control of Jilin Province (Jilin, China). Adult female ICR mice (20–25 g) and Wistar rats (180–200 g) were provided by the Laboratory Animal Center of Jilin University (Changchun, China). All procedures were approved and supervised by the Animal Care and Use Committee, School of Life Sciences, Jilin University (Changchun, Jilin, China). The entire procedure was carried out in accordance with related guidelines and regulations.
Bacterial And Fungal Culture Conditions
The bacterial strains (Table 2) were cultured in lysogeny broth (LB) agar plates from stock cultures. LB agar plates were incubated at 37°C for 18–24 hrs to obtain fresh cultures prior to each in vitro bioassay. Dry powder of C. albicans was dissolved in sterile 10 mM phosphate buffer saline (PBS) (pH 7.4), vortexed, and spread on an SD broth agar plate. After 18–24 hrs of incubation at 35 °C, a colony was inoculated into the SD broth medium and incubated overnight at 35 °C with shaking to acquire logarithmic phase growth. Microbial strains were confirmed by a Gram stain and by colonial and growth characteristics.
 |
Table 1 Sequences Of AMPs Used In This Study |
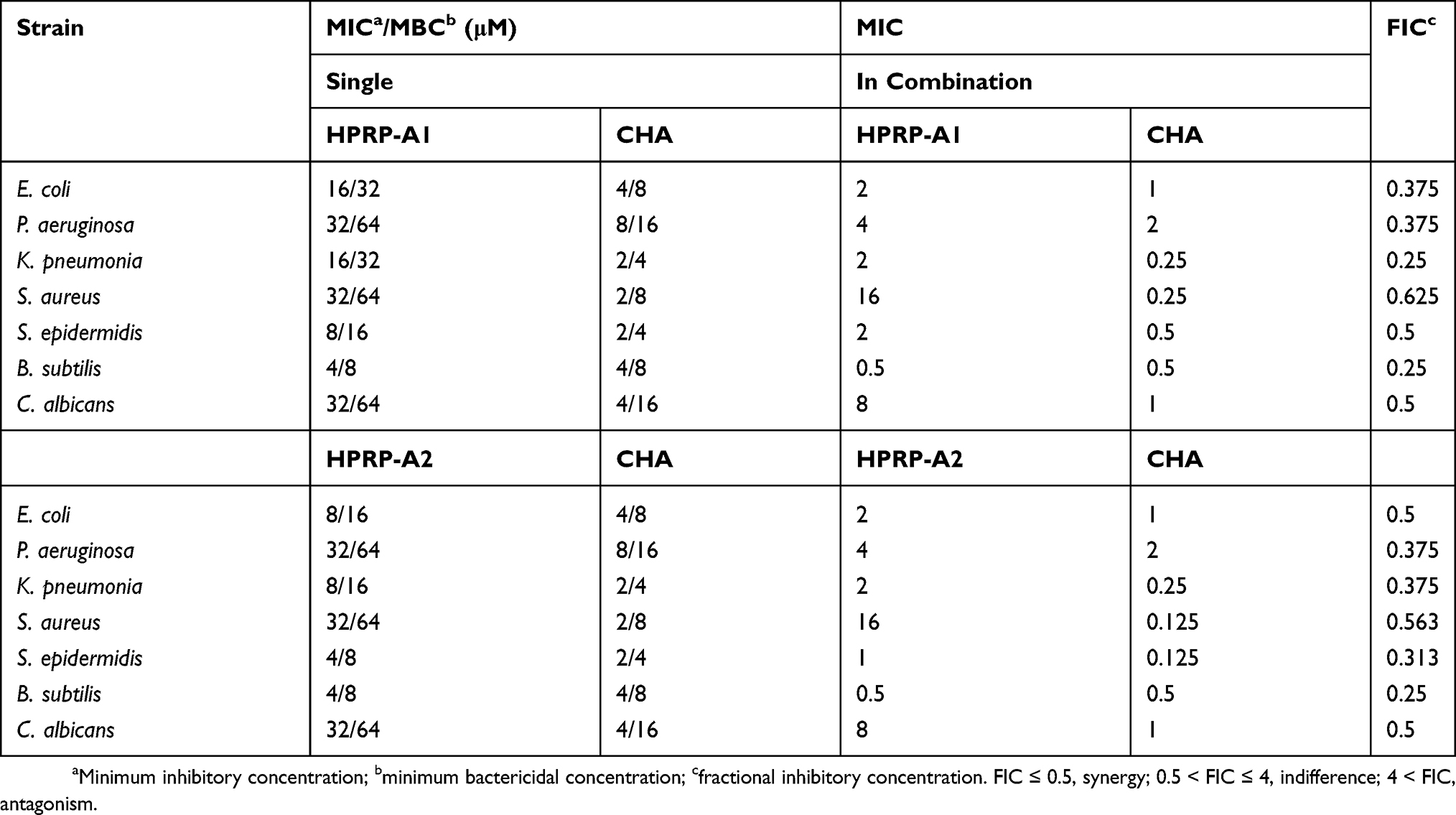 |
Table 2 MIC Values Of HPRP-A1 And HPRP-A2 Single Or In Combination With CHA And The Respective Fractional Inhibitory Concentration Index Values Obtained Using The Checkerboard Method |
Peptide Synthesis And Purification
Basing on 9-fluorenylmethoxycarbonyl (Fmoc) chemistry.24 AMPs were synthesized by Fmoc Solid Phase Peptide Synthesis (SPPS) on an 4-methyl-benzhydrylamine (MBHA) resin, then purified by preparative Shimadzu LC-6A reverse-phase high-performance liquid chromatography (RP-HPLC) using a Zorbax 300 SB-C8 column (250-mm×9.6-mm inner diameter, 6.5-μm particle size, 300-Å pore size). Eluent A was 0.1% aqueous solution of trifluoroacetic acid (TFA), and eluent B was 0.1% TFA in acetonitrile solution. Peptides were characterized by mass spectrometry and amino acid analysis.
Measurement Of Antimicrobial Activity
The minimum inhibitory concentration (MIC) of peptides or agents was used to determine the antimicrobial activity; the minimum bactericidal concentration (MBC) of peptides or agents was used to describe the killing ability against bacteria and fungi. Bacteria and fungi were grown in 96-well microtiter plates. Determination of MICs was carried out by a standard microdilution method. Briefly, bacteria and fungi were grown overnight at 37°C and 35°C, respectively, in LB medium and SD broth medium, respectively, diluted in the same sterile medium. HPRP-A1, HPRP-A2 and CHA were serially diluted in sterile PBS and added to 96-well microtiter plates in a volume of 10 μL; then, 90 μL of bacteria was added to give a final inoculum of 5×105 CFU/mL for the Gram-negative and Gram-positive strains and 2×105 CFU/mL for the fungal strain. Then, plates were incubated at 37°C or 35°C, respectively, for 18–24 hrs.
To determine whether peptides work synergistically with CHA, we used the broth microdilution checkerboard method to measure the efficacy of combinations of HPRP-A1 and HPRP-A2 with CHA against Gram-negative and Gram-positive bacteria and a fungus.28 AMPs HPRP-A1 (5 μL), HPRP-A2 (5 μL) and CHA (5 μL) were serially diluted, and then 90 μL of bacteria was added to the 96-well microtitre plates to produce a final inoculum of 5×105 CFU/mL of bacteria and 2×105 CFU/mL of fungus.1 The plates were incubated for 18–24 hrs at 37°C and 35°C, respectively. The peptides and agent dilution ranges were 0.25–500 μM. The fractional inhibitory concentration (FIC) index for combining two drugs was calculated according to the equation: (FIC = MICA/B/MICA+ MICB/A/MICB), where MICA, MICB is the MIC of drug A, B single, and MICA/B, MICB/A is the MIC of the compound A, B in combination. We classified the results as synergistic activity (FIC ≤ 0.5), indifferent activity (0.5 < FIC < 4) and antagonism (FIC ≥ 4).
PI Uptake Assay
Fluorescence microscopy (FM). The propensity of HPRP-A1, HPRP-A2 and/or CHA to cause loss of membrane integrity was measured by a PI uptake assay using FM. PI and SYTO 9 are derived from BBcell ProbeTM live/dead bacterial staining kit for staining live/dead bacteria and fungus. Briefly, freshly collected logarithmic E. coli, B. subtilis and fungus C. albicans cultures with an inoculum size of 1×107 CFU/mL were incubated with synergistic concentrations of HPRP-A1, HPRP-A2 and/or CHA, or PBS as the negative control. After incubation at 37 °C for 1 hr, PI solution was added to acquire a final concentration of 50 μg/mL, and SYTO 9 was added at 2 µL/mL and a further incubation was conducted for 15 mins protected from the light. Cells were then centrifuged, washed and re-suspended in PBS and analyzed by FM (LSM 710 META; Zeiss, Oberkochen, Germany), Adobe Photoshop CS6 13.0.1 (Photoshop Software, Ireland, UK) was used to analyze the image area. The calculation was made according to the following formula: %Bacteriostatic = Sr/(Sr + Sg)×100 to analyze the image area, Sr is red area, Sg is green area.
Fluorescence-Activated Cell Sorting (FACS)
To quantitatively analyze the loss of fungal membrane integrity after peptide treatment, C. albicans cells (1×107 CFU/mL) were treated with increasing concentrations of HPRP-A1, HPRP-A2 and/or CHA and were incubated for 1 hr at 37 °C with E. coli and B. subtilis, or for 1 hr at 35 °C with C. albicans and then stained with PI (final concentration = 50 μg/mL) for 15 mins at room temperature in the dark environment. Subsequently, the cells were harvested by centrifugation, washed and suspended in PBS and finally analyzed by flow cytometry (FACSCalibur, Becton-Dickinson, San Jose, CA, USA).
LPS Binding Assay
Determination of the binding ability of HPRP-A1, HPRP-A2 and/or CHA to bacterial LPS was performed by mixing 50 μL of exogenous E. coli LPS (0.25 μg/mL) with each group of drugs in 96-well microtiter plates, followed by antibacterial tests of E. coli at 50 μL final concentration of 5×105 CFU/mL for 1 hr at 37 °C. Then, 0.5 μL of Sytox green diluent mixture was added to each well. Fluorescence was measured at an excitation wavelength of 510 nm at room temperature and an emission wavelength of 535 nm using the multifunctional enzyme marker (Infinite F200 pro, TECAN, Männedorf, Switzerland). Sytox green dye was added to each well at a final concentration of 0.5 μM for 15 mins in a dark environment.
Cellular Reactive Oxygen Species Assay
The amounts of reactive oxygen species (ROS) were measured by a fluorometric assay with 2′,7′-dichlorofluorescein diacetate (DCFH-DA).29 Fungal cells (2×106 CFU/mL) were treated with HPRP-A1, HPRP-A2 and/or CHA or left untreated for 1 hr at 35 °C. After incubation, cells were stained with DCFH-DA for 30 mins. The fluorescence intensity of the treated cells was measured with a spectrofluorometer at different intervals with the maximum excitation wavelength of 485 nm and the maximum emission wavelength of 535 nm.
DNA Binding Assay
E. coli genomic DNA was extracted using a bacterial genomic DNA extraction kit (TransGen Biotech, China); then, 10 μL of the purified genomic DNA was incubated with 10 μL of different concentrations of HPRP-A1, HPRP-A2 and/or CHA for 1 hr at room temperature. A 9 μL of mixed solution was added to 1 μL of native 10× loading buffer, 5 μL DL 2000 DNA marker as a measure of the molecular weight was separated by 1.0% agarose gel electrophoresis for analysis of the DNA binding, respectively.
Preparation Of Experimental Animals
In each experimental group, 10 animals were used (ICR mice and Wistar rats). It is necessary to feed for 3 days before the experiment. Animals are fed with standard feed, natural light, and free eating water. Light is under 12 hrs of alternating light and dark environments. Mouse weight is between 20 and 25 g and rat weight is 180–200 g. A final concentration of approximate 1×107 CFU/mL was used to prepare E. coli and S. aureus strains, and a final concentration of approximate 1×108 CFU/mL was used to prepare C. albicans strain for in vivo tests.30
Vaginitis Infection Models
Rat bacterial vaginitis. A 1:1 suspension of E. coli and S. aureus was taken up with a micropipette (0.025 mL/100 g), and the pipette was slowly inserted 1 cm to 1.5 cm into the vagina of each experimental group, with the exception of the blank control, and the bacterial suspension was injected. Each rat was inverted for 2 mins after inoculation to prevent bacterial fluid from overflowing. The blank group was given the same amount of normal saline. The vaginal lesions of rats were observed each day after inoculation. The vaginal mucous membrane was obviously congested. Many infecting bacteria were detected in the vagina of rats by the culture of vaginal secretions, which proved that the model of bacterial vaginitis infection by E. coli and S. aureus had been successfully constructed.
Mouse C. albicans vaginitis. The mice were subcutaneously injected with 0.05 mg/mL of 2 mg/mL estradiol benzoate oil, once every 2 days, 3 times in total, to ensure the mice were in a sham state. Then, 20 μL of bacterial suspension (1.5×108 CFU/mL) was inoculated into the vagina of each mouse with a micropipette, and each mouse was placed upside down for 5 mins after inoculation; then, the vaginal orifice of the mouse was gently pinched with tweezers for 30 s to prevent leakage of the bacterial fluid. In the control group, 20 μL of aseptic PBS was injected into the vagina.
In Vivo Drug Treatment
Rats and mice were fed with drugs at a concentration of 10 mg/kg according to their body weight. Animals were held upside down, injected each group of drug solutions from their vaginas and kept still for 30 s. Tests were carried on with the same method once a day for 8 continuous days. In the end, 100 μL of PBS was used to drive into the vagina and then was aspirated, the bacterial count was measured by spread plate method, and the external vaginal surface appearance and allergic reactions were observed. Surgery was carried out to remove the vagina and check the vaginal edema symptom. The percentage of inhibition of bacteria in the results of each group was counted by the following formula: %Bacteriostatic = 100 – CFUd/CFUc × 100, CFUd is the number of bacterial colonies in the drug treatment group, CFUc is the number of bacterial colonies in the control group.31
Statistical Analysis
Statistical analysis was carried out using one-way analysis of variance (ANOVA) in Prism software 5.0 (GraphPad Prism, San Diego, CA, USA). Error bars showed standard deviations of three independent experiments. A p-value < 0.05 was considered to indicate a significant difference, a p-value < 0.01 was considered to indicate a highly significant difference and a p-value of < 0.001 indicated an extremely significant difference.
Ethics Statement
All care and handling of animals were performed in accordance with the guidelines of the animal ethics committee of Jilin University (Approval No. JLUSWLL003, Jilin, China).
Results
Antimicrobial Activity Of The Peptides And CHA
In this study, the sequence of the enantiomers HPRP-A1 and HPRP-A2 is shown in Table 1. HPRP-A1 is composed of all L-amino acids and HPRP-A2 is composed of all D-amino acids, high-performance liquid phase and mass spectrometry analysis and verification (Figures S1A and B).
The antimicrobial activities of HPRP-A1, HPRP-A2 and CHA single against various Gram-negative and Gram-positive bacteria and a fungus were determined as the MICs and the minimum bactericidal concentrations (MBCs), as shown in Table 2. The results of MIC/MBC values of HPRP-A1, HPRP-A2 and CHA showed good effects against E. coli, K. pneumonia and B. subtilis, respectively. HPRP-A1, HPRP-A2 and CHA showed modest antibacterial activity against P. aeruginosa, S. aureus and C. albicans than other strains. The MBC values against each bacterial strain were 2–4-fold greater than those of the MIC values, respectively.
The combined application of two AMPs HPRP-A1 and HPRP-A2 with CHA against Gram-negative and Gram-positive bacteria and a fungus was examined and the results are shown in Table 2. Most of the FIC indices were less than or equal to 0.5, confirming that HPRP-A1, HPRP-A2 and CHA have a synergistic activity to bacteria and fungus. Based on these results, it is clear to see that HPRP-A1 with CHA and HPRP-A2 with CHA showed synergistic effects on three Gram-negative bacteria, two Gram-positive bacteria and one fungus, but had no detectable synergistic effect on S. aureus (Table 2).
Effects Of HPRP-A1, HPRP-A2 And/or CHA On Bacterial And Fungal Viability
The effects of HPRP-A1, HPRP-A2 and/or CHA on the activity and membrane integrity of selected Gram-negative and Gram-positive bacteria and fungus were examined by fluorescence microscopy (FM) following propidium iodide (PI) staining. E. coli and B. subtilis were selected to present the Gram-negative and Gram-positive bacteria, respectively, due to the MIC values in drug combinations. The results obtained with intact E. coli are presented in Figure 1A and showed that more than 99% of the bacterial cells were alive. However, there are 41% and 74% of the bacterial cells (in red color) following treatment with HPRP-A1 and HPRP-A2, respectively, in contrast, following treatment with CHA, indicating the ineffective disruption of membrane with CHA. Interestingly, in the combination groups of HPRP-A1/HPRP-A2 with CHA, the inhibition rate of bacteria was significantly higher than that in the single-drug treatment groups of AMPs and CHA, which reaches more than 99% (Figures 1A, S2A and B).
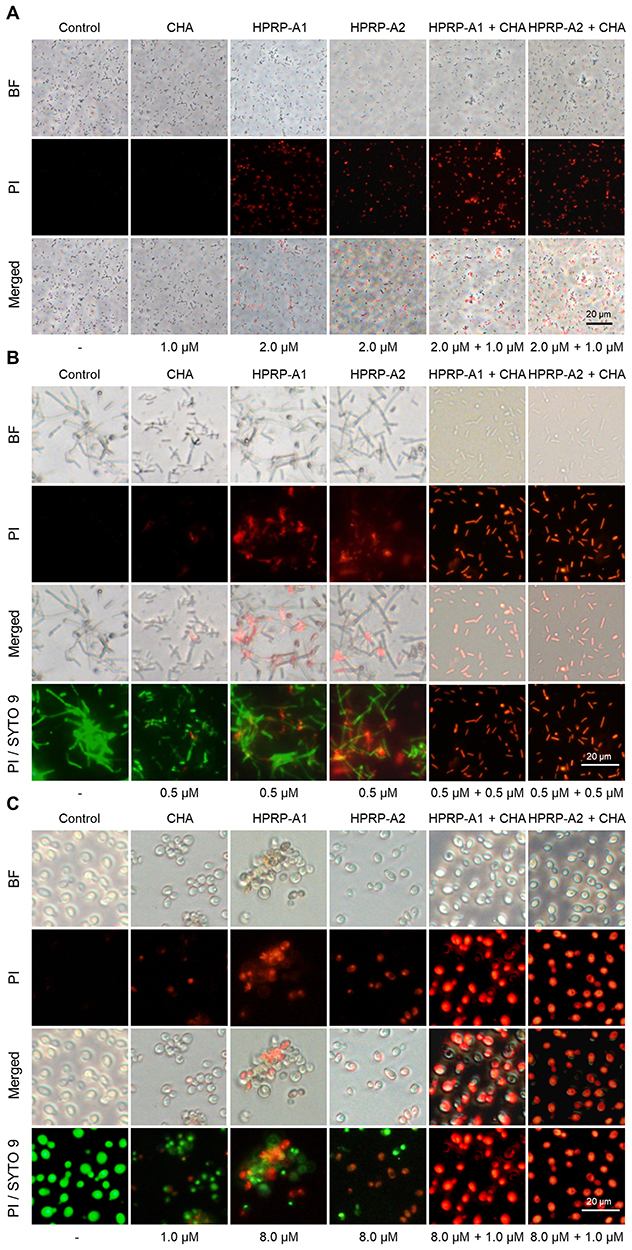 |
Figure 1 Fluorescence microscopy analysis. (A) Lysogeny broth (LB) medium (control), HPRP-A1 (2.0 μM), HPRP-A2 (2.0 μM) and/or CHA (1.0 μM) against E. coli; (B) LB medium (control), HPRP-A1 (0.5 μM), HPRP-A2 (0.5 μM) and/or CHA (0.5 μM) against B. subtilis; (C) Sabouraud dextrose (SD) broth medium (Control), HPRP-A1 (8.0 μM), HPRP-A2 (8.0 μM) and/or CHA (1.0 μM) against C. albicans. Bacterial or fungal cells were treated for 1 hr at 37°C and then stained for 15 mins with PI and/or SYTO 9. E. coli was only stained with PI, which stains dead cells red, B. subtilis and C. albicans were stained with PI and SYTO 9, which stain dead cells red and live cells green, respectively. Cells were visualized in bright field (BF) by fluorescence microscopy. |
The results for B. subtilis and C. albicans are presented in Figure 1B, Figure S2C and Figure 1C, Figure S2D, which show that about 99% of cells are alive (in green color), confirming that they were viable, and their membranes were intact. In contrast, about 22%, 31% and 1% of B. subtilis and about 25%, 53% and 12% of C. albicans cells were dead (in red color), indicating corresponding membrane disruption, respectively. In addition, more than 85% and 99% of bactericidal and fungal membrane damage were observed after combined treatments with HPRP-A1, HPRP-A2 with CHA in B. subtilis and C. albicans, respectively, which was verified by the turbidity method (Figure S3A–3C).
Fluorescence-activated cell sorting (FACS) was also used to detect the integrity of bacterial and fungal membranes following treatment with HPRP-A1, HPRP-A2 and/or CHA by monitoring the uptake of PI. The results showed that HPRP-A1, HPRP-A2 and CHA can induce PI to enter bacterial and fungal cells, resulting in fluorescence intensity increases. The combined application groups showed stronger fluorescence intensity than the single treatment groups shown as follows: E. coli (Figure 2A), B. subtilis (Figure 2B) and C. albicans (Figure 2C). These results were consistent with the results of the FM assay indicating that the combined application groups of HPRP-A1 with CHA, and HPRP-A2 with CHA, exhibited greater membrane degradation than the single application groups.
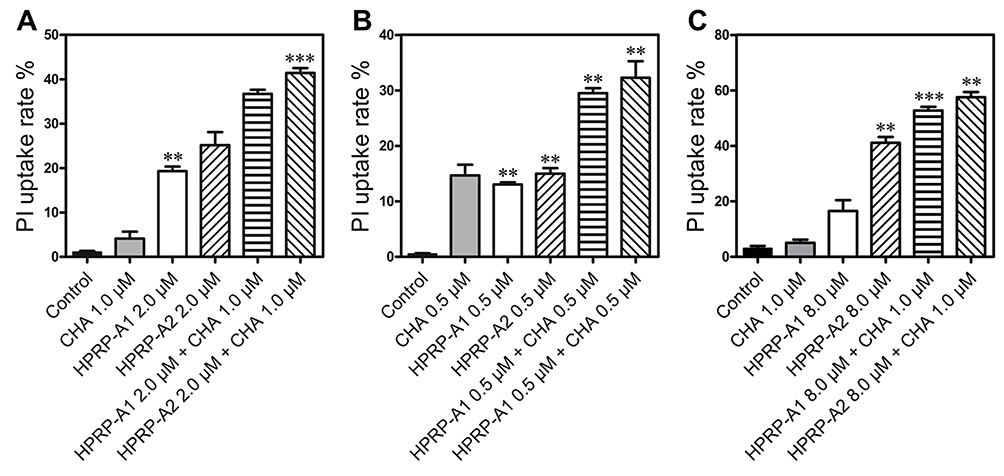 |
Figure 2 FACS analysis of membrane permeabilization. (A) HPRP-A1 (2.0 μM), HPRP-A2 (2.0 μM) and/or CHA (1.0 μM) against E. coli; (B) HPRP-A1 (0.5 μM), HPRP-A2 (0.5 μM) and/or CHA (0.5 μM) against B. subtilis; (C) HPRP-A1 (8.0 μM), HPRP-A2 (8.0 μM) and/or CHA (1.0 μM) against C. albicans. Bacterial or fungal cells were incubated for 1 hrs at 37 °C, followed by 15-min incubation with PI in a dark environment and analyzed by FACS. LB medium was used as control in (A and B), and SD broth medium was used as control in (C). **P < 0.01; ***P < 0.001. |
The Binding Of Peptides And/Or CHA To Components Of Bacterial Cells
Sytox green dye can bind to the bacterial nucleic acid, and it can penetrate a damaged plasma membrane, but not through a living cell membrane.32 Based on the mechanism of action of peptides and agents that target the cell membrane, the binding ability of HPRP-A1, HPRP-A2 and/or CHA to the LPS of Gram-negative bacterial cell wall was determined and the results are shown in Figure 3. Co-incubation of HPRP-A1, HPRP-A2 (2 μM) and/or CHA (1 μM) with LPS (0.25 μg/mL) of Gram-negative bacteria in exogenous E. coli showed significant affinity. On the contrary, the binding ability of CHA and LPS is relatively weak; in addition, no obvious improvement in the LPS binding ability was shown for the combined applications of drugs.
 |
Figure 3 The binding activities of HPRP-A1, HPRP-A2 and/or CHA to bacterial LPS. Fluorescence analysis of HPRP-A1, HPRP-A2 (2 μM) and/or CHA (1 μM) with the LPS (0.25 μg/mL) of exogenous E. coli and then act on E. coli cells. CHA, HPRP-A1, HPRP-A2, HPRP-A1+CHA, HPRP-A2+CHA direct acting bacteria, CHA+LPS, HPRP-A1+LPS, HPRP-A2+LPS, HPRP-A1+CHA+LPS, HPRP-A2+CHA+LPS first combined with LPS of exogenous E. coli for 1 hr and then act on E.coli cells, using E. coli as negative control. HPRP-A1, HPRP-A2 and/or CHA were analyzed by measuring the increase in Sytox green fluorescence in E. coli, and the fluorescence was measured using a fluorospectrophotometer (excitation λ, 510 nm; emission λ, 535 nm). *P < 0.05; **P < 0.01. |
Peptides And/Or CHA Could Induce The Production Of Reactive Oxygen Species From Fungal Cells
Excessive levels of erythropoietin can lead to cell death, and increased levels of reactive oxygen species (ROS) in cells can damage proteins, lipids and DNA.33 Thus, the effect of HPRP-A1, HPRP-A2 and/or CHA on cellular ROS production in fungus was determined by a DCFH-DA assay. DCFH-DA is an intracellular dye that can be oxidized by ROS to form the fluorescent molecule 2,7-dichlorofluorescein with a maximum excitation wavelength of 485 nm and a maximum emission wavelength of 535 nm.29 As shown in Figure 4, HPRP-A1, HPRP-A2 and CHA alone did not produce ROS. In the combination groups, oxygen consumption by the fungus was not increased. The synergistic concentrations of HPRP-A1 with CHA, and HPRP-A2 with CHA, were not significantly different from the individual treatment groups of HPRP-A1, HPRP-A2 and the control, respectively. Significant differences were observed between the HPRP-A1 and CHA group or the HPRP-A2 and CHA group and the CHA alone group, respectively.
 |
Figure 4 Effect of HPRP-A1, HPRP-A2 and/or CHA on cellular ROS formation by C. albicans cells. C. albicans, C. albicans+DCFH-DA, CHA (1 μM)+DCFH-DA, HPRP-A1 (8 μM)+DCFH-DA, HPRP-A2 (8 μM)+DCFH-DA, HPRP-A1 (8 μM)+CHA (1 μM)+DCFH-DA, HPRP-A2 (8 μM)+CHA (1 μM)+DCFH-DA. Each group was compared with the SD broth medium (Control). HPRP-A1/A2 (8 μM) and/or CHA (1 μM) were added and incubated for 1 hr at 35 °C. Then, 10 μM DCFH-DA was added for 30 mins in a dark environment. The fluorescence was measured using a fluorospectrophotometer (excitation λ, 485 nm; emission λ, 535 nm). **P < 0.01. |
The Interaction Between Peptides And/Or CHA And Bacterial Genomic DNA
To confirm the interaction between HPRP-A1, HPRP-A2, CHA and genomic DNA in bacteria, a DNA-binding assay and a nucleic acid electrophoresis were performed; the results are shown in Figure 5A–C. There is an obvious concentration-dependent trend of peptide binding to bacterial genomic DNA bands, suggesting that the genomic DNA of E. coli was tightly bound to extremely high doses of HPRP-A1 (64 μM) and HPRP-A2 (32 μM), but not to CHA (>64 μM). However, the bindings of genomic DNA of E. coli at the synergistic concentrations were not found for both HPRP-A1 (2 μM) with CHA (1 μM), and HPRP-A2 (2 μM) with CHA (1 μM), respectively. Thus, it is obvious to see that the antimicrobial mechanism of low concentrations of the drug combinations is not due to the disruption of bacterial DNA.
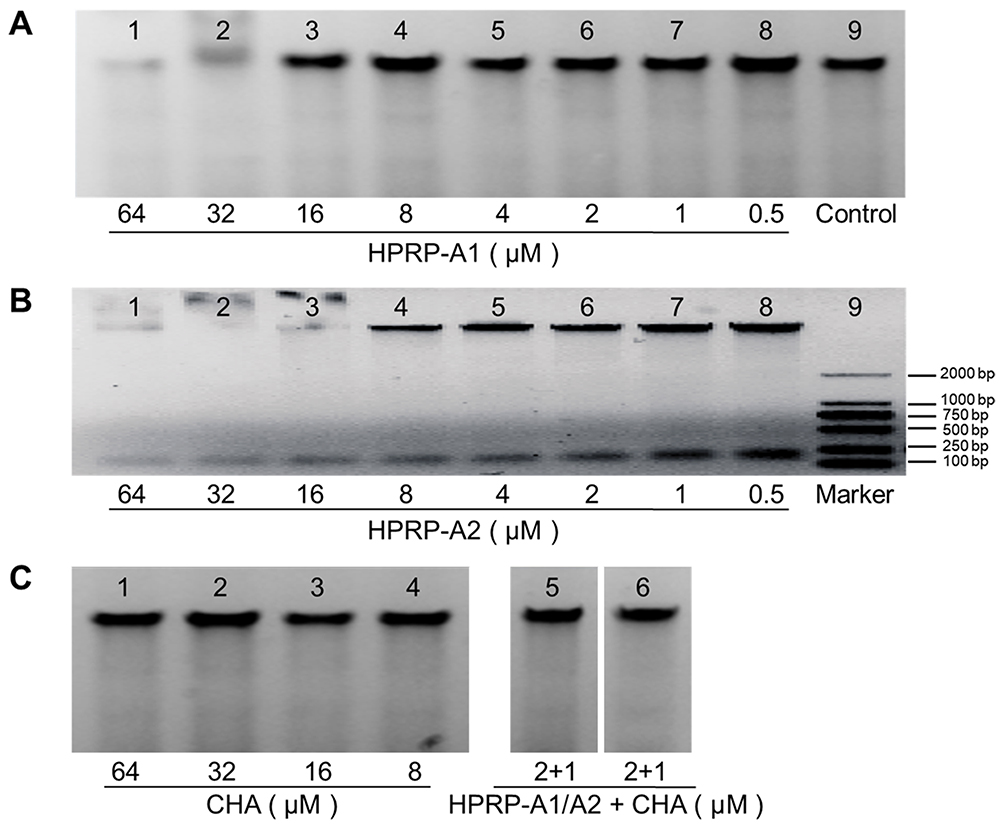 |
Figure 5 The binding activities of HPRP-A1, HPRP-A2 and/or CHA to bacterial genomic DNA. (A) Interaction of HPRP-A1 (64 to 0.5 μM) with E. coli genomic DNA. (B) Interaction of HPRP-A2 (64 to 0.5 μM) with E. coli genomic DNA. (C) Interaction of CHA (64 to 8 μM), HPRP-A1/2 (2 μM) and CHA (1 μM) with E. coli genomic DNA. The marker (DL 2000 DNA marker) is a measure of the molecular weight of. E. coli genomic DNA and LB medium as a negative control, E. coli genomic DNA and peptides and/or CHA at a ratio of 1:1, with 1.0% agarose gel electrophoresis at 100 V and a constant current. |
Evaluating The Therapeutic Effects Of Drug Combinations In Vivo
Gynecological vaginitis is one of the most commonly gynecological diseases in women in the world, most of which are BV or VVC, usually caused by E. coli, S. aureus or C. albicans, respectively. CHA is the most common drug used to treat vaginitis in women. In this study, synergistic antibacterial and antifungal effects of HPRP-A2 and CHA against Gram-positive and Gram-negative bacteria, and fungus were evaluated against BV in rats and VVC in mice at two different doses. The concentrations of experimental animals were determined based on MIC values and pre-tests.
In the low-dose groups (Table 3), the groups of combination of HPRP-A2 and CHA inhibited most of the bacteria and fungus compared to other treatment groups. Compared with the untreated controls, HPRP-A2 alone inhibited E. coli, S. aureus and C. albicans to about 40%. Single dose of CHA inhibited E. coli, S. aureus and C. albicans to about 30%. Moreover, HPRP-A2 and CHA combination group dramatically inhibited E. coli, S. aureus and C. albicans to 80.3%, 78.4% and 84.6%, respectively.
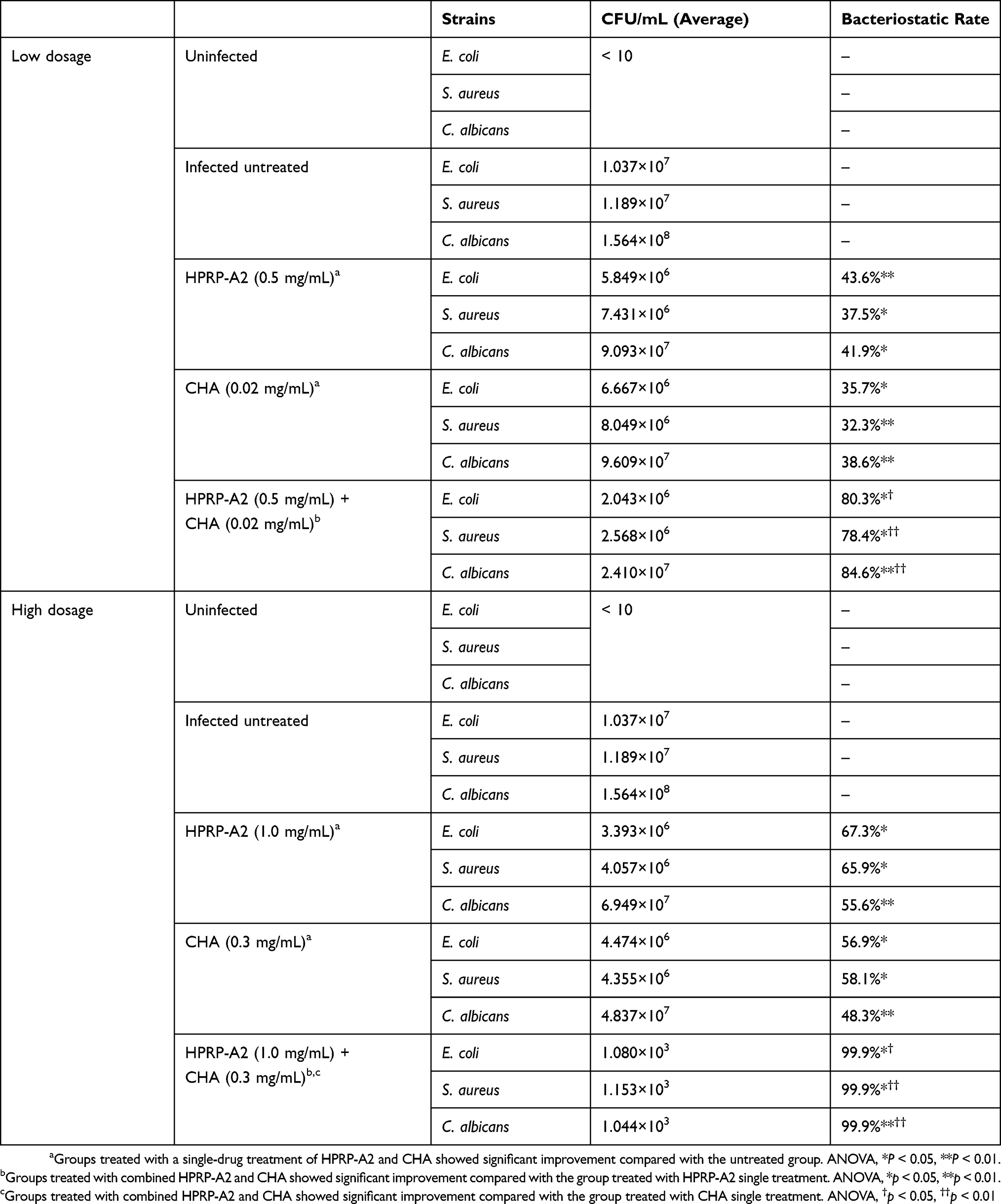 |
Table 3 Quantitative Culture Of Excised Tissues After Drug Administration In E. coli, S. aureus And C. albicans Infected Rats And Mice In The Vaginitis Infection Models |
The high-dose group significantly inhibited all bacteria and fungus compared to the control group. Compared with the control group, the HPRP-A2 alone high-dose group showed inhibition against E. coli, S. aureus and C. albicans to about 50%; similarly, the CHA alone high-dose group showed inhibition against E. coli, S. aureus and C. albicans about 50%. In contrast, HPRP-A2 and CHA combination high-dose group significantly inhibited E. coli, S. aureus and C. albicans to 99.9%.
Thus, whether in low-dose or high-dose treatment groups, the number of vaginal bacteria in mice and rats decreased to different degrees after single or combined treatments of peptide and CHA. Particularly, the high-dose groups inhibited more bacteria and fungus than the low-dose groups; however, the combination groups exhibited excellent bacteriostatic effects with 99.9% inhibition rate compared with the single drug groups. In the mice and rat models virginally infected with E. coli, S. aureus and C. albicans, the combination group of HPRP-A2 and CHA exhibited strong synergistic antimicrobial and antifungal effects both at low and at high doses (Figure S4A and B).
Discussion
In this study, the antibacterial and antifungal activities of antimicrobial peptide HPRP-A1 and HPRP-A2 and antimicrobial agent CHA single or in combination against Gram-positive and Gram-negative bacteria and fungi were investigated in vitro and in vivo. AMPs exhibited a broad spectrum of antimicrobial activities against Gram-positive, Gram-negative bacteria and a fungus.
HPRP-A1 and HPRP-A2 enhance cell membrane permeability through interaction with cell membranes, ultimately leading to cell death. As described in the “membrane discrimination mechanism”, AMPs take a different approach to disrupt the membrane due to the different compositions of phospholipids of eukaryotic or prokaryotic cells.25,34 Their specific role in bacterial sterilization follows the mechanism of the “carpet model”.35 When the peptides interact with the membrane, the ordered phospholipids become unstable. The stability of the local membrane is disrupted by the location of the peptide in phospholipids, and when the peptide concentration reaches a critical point, the membrane would disintegrate due to insufficient energy. In addition, for fungal cells, to follow the “barrel-stave model” mechanism, initially, the peptide may bind to the membrane surface in a monomeric form, and the hydrophobic region of the peptide would vertically insert into the membrane in interaction with the hydrophobic core of the membrane and form channels or pores. At the critical concentration point, these channels or pores cause the disruption of the eukaryotic membrane.36
As the enantiomeric isomer of HPRP-A1 and HPRP-A2 show the almost same activity. The antibacterial and antifungal mechanisms of HPRP-A1 and HPRP-A2 are independent of the chiral characteristics, but depend on the α-helical and amphipathic structure.34 However, HPRP-A2 exhibited slightly better MIC values than HPRP-A1, which could be due to its stability to the degradation of protease.37,38
As a broad-spectrum antibacterial drug, it is generally postulated that the antimicrobial activity of CHA stems from its cationic properties. CHA can interact with the negatively charged cell envelope, resulting in breakage of the outer leaflet. High concentrations of CHA severely compromise the cytoplasmic membrane, leading to cell lysis.39 At lower concentrations near the MIC, CHA distorts the cell walls of Gram-positive and Gram-negative bacteria, leading to morphological changes in the cell surface.13 Uptake of CHA into the cytoplasm causes precipitation of the cytoplasmic components and inhibits ATPase activity.40 However, low concentration of CHA in this study, such as MIC, has a relatively weak effect on membrane breaking of bacteria and fungus.
HPRP-A1 and HPRP-A2 can bind to LPS, the major outer membrane component of Gram-negative bacteria. In bacterial infectious diseases, LPS can cause inflammation, such as sepsis, septicemia and so on.41 LPS is a key stimulating factor for the pathogenesis of Gram-negative bacteria, and not only can it be used as a barrier for bacteria to resist the surroundings, but it also acts as an immune response to invading bacteria, releasing it at cell division or cell death. Hence, LPS can be used as a target for drugs to achieve the treatment of the diseases.42 LPS was delivered into the cytosol and triggered caspase-11-mediated pyroptosis after exposure to antibiotics or antimicrobial peptides, and this is critical for antimicrobial defense and septic shock.43 In this study, HPRP-A1 and HPRP-A2 can strongly bind to LPS of E. coli, which reduces the bactericidal effect on E. coli. On the contrary, the binding ability of CHA and LPS is relatively weak; in addition, there is no obvious improvement in the LPS-binding ability of combined application of drugs. In our previous study,44 HPRP-A1 was shown to kill cancer cells by inducing the decrease in mitochondrial membrane potential and the production of reactive oxygen species in cancer cells. However, as eukaryotic cells, fungus is not associated with reactive oxygen species in mitochondria like cancer cells. This may suggest that the fungal killing ability of HPRP-A1, HPRP-A2 with CHA is not associated with the production of erythropoietin by reactive oxygen species.
In this study, combined with HPRP-A1, HPRP-A2 and CHA, showed excellent synergistic antibacterial and antifungal activity in vitro and in vivo which, we believe, would be due to the different mechanisms of action of AMPs and CHA. Usually, the bactericidal activity of AMPs comes from the disruption of the integrity of membranes, but there are some AMPs interacting with additional intracellular components, such as bacterial DNA. In this study, HPRP-A1 and HPRP-A2 were found to bind to the E. coli DNA of Gram-negative bacteria, suggesting that there may be an intrinsic target of the nuclear material in the mechanism of action. In contrast, CHA (< 64 μM) does not possess the binding ability with bacterial DNA. This suggests that the peptide may kill bacteria by penetrating and disrupting the cell membrane and then targeting the intracellular components of the cell including genomic DNA.
Due to the long-term antibiotic resistance caused by gynecological vaginitis, antibiotics have gradually lost their power to inhibit infections. In addition, many clinically applied CHAs continue to have side effects during the treatment, and the phenomenon of allergies is increasing.17,18 In vivo experiments on mouse and rat vaginitis exhibited that both low and high doses achieved satisfactory therapeutic results after the treatment. The combination application of AMPs and CHA could be a practical approach to fight against antibiotic resistance of gynecological vaginitis in clinical practices.
In conclusion, our results show that the combination of antimicrobial peptide HPRP-A1, HPRP-A2 and CHA has strong synergistic effects on Gram-negative and Gram-positive bacteria and fungus in vitro and in vivo. The antibacterial activity of AMPs relies not only on penetrating the cell membrane rapidly but also on binding with bacterial DNA. Drug combinations achieved 99.9% of inhibition against infections in gynecological vaginitis caused by bacteria and fungus in rat and mouse models. The excellent synergy of AMPs and CHA is due to the different mechanisms of action at different corresponding concentrations. The combination application of AMPs and CHA could be a promising approach to treat gynecological vaginitis in clinical practices.
Acknowledgments
This work was supported by the National Key Science and Technology Projects of China (2018ZX09721001-004-005 to YXC), the Natural Science Foundation of Jilin Province of China (20180101250 JC to YBH) and the “13th Five-Year” Science and Technology Project of Jilin Provincial Department of Education (JJKH20180178 KJ to YBH).
Disclosure
Mingxia Chen is an employee and Yuxin Chen is one of the shareholders of Jiangsu ProteLight Pharmaceutical & Biotechnology Co., Ltd. The authors report no other conflicts of interest in this work.
References
1. Petersen PJ, Labthavikul P, Jones CH, Bradford PA. In vitro antibacterial activities of tigecycline in combination with other antimicrobial agents determined by chequerboard and time-kill kinetic analysis. J Antimicrob Chemother. 2006;57(3):573–576. doi:10.1093/jac/dki477
2. Corsello S, Spinillo A, Osnengo G, et al. An epidemiological survey of vulvovaginal candidiasis in Italy. Eur J Obstet Gynecol Reprod Biol. 2003;110(1):66–72. doi:10.1016/s0301-2115(03)00096-4
3. Foxman B, Muraglia R, Dietz JP, Sobel JD, Wagner J. Prevalence of recurrent vulvovaginal candidiasis in 5 European countries and the United States: results from an internet panel survey. J Low Genit Tract Dis. 2013;17(3):340–345. doi:10.1097/LGT.0b013e318273e8cf
4. Verstraelen H, Verhelst R, Roelens K, Temmerman M. Antiseptics and disinfectants for the treatment of bacterial vaginosis: a systematic review. BMC Infect Dis. 2012;12:148. doi:10.1186/1471-2334-12-166
5. De Bernardis F, Graziani S, Tirelli F, Antonopoulou S. Candida vaginitis: virulence, host response and vaccine prospects. Med Mycol. 2018;56:S26–S31. doi:10.1093/mmy/myx139
6. Mushi MF, Ngeta N, Mirambo MM, Mshana SE. Predictors of esophageal candidiasis among patients attending endoscopy unit in a tertiary hospital, Tanzania: a retrospective cross-sectional study. Afr Health Sci. 2018;18(1):66–71. doi:10.4314/ahs.v18i1.10
7. Jacob L, John M, Kalder M, Kostev K. Prevalence of vulvovaginal candidiasis in gynecological practices in Germany: A retrospective study of 954,186 patients. Curr Med Mycol. 2018;4(1):6–11. doi:10.18502/cmm.4.1.27
8. Workowski KA, Bolan GA, Centers for Disease CPrevention. Sexually transmitted diseases treatment guidelines, 2015. MMWR Recomm Rep. 2015;64:1–137.
9. Gonzalez-Burgos E, Gomez-Serranillos MP. Natural products for vulvovaginal candidiasis treatment: evidence from clinical trials. Curr Top Med Chem. 2018;18:1324–1332. doi:10.2174/1568026618666181002111341
10. Verstraelen H, Verhelst R. Bacterial vaginosis: an update on diagnosis and treatment. Expert Rev Anti Infect Ther. 2009;7(9):1109–1124. doi:10.1586/eri.09.87
11. Wade BE, Zhao J, Ma J, Hart CM, Sutliff RL. Hypoxia-induced alterations in the lung ubiquitin proteasome system during pulmonary hypertension pathogenesis. Pulm Circ. 2018;8(3):1–17. doi:10.1177/2045894018788267
12. Gilbert P, Moore LE. Cationic antiseptics: diversity of action under a common epithet. J Appl Microbiol. 2005;99(4):703–715. doi:10.1111/j.1365-2672.2005.02664.x
13. Cheung HY, Wong MM, Cheung SH, Liang LY, Lam YW, Chiu SK. Differential actions of chlorhexidine on the cell wall of Bacillus subtilis and Escherichia coli. PLoS One. 2012;7(5):e36659. doi:10.1371/journal.pone.0036659
14. Ferris DG, Francis SL, Dickman ED, Miler-Miles K, Waller JL, McClendon N. Variability of vaginal pH determination by patients and clinicians. J Am Board Fam Med. 2006;19(4):368–373.
15. Hsu JF, Chen CL, Lee CC, et al. Characterization of group B Streptococcus colonization in full-term and lete-preterm neonates in Taiwan. Pediatr Neonatol. 2018;18:139.
16. Bernardi A, Teixeira CS. The properties of chlorhexidine and undesired effects of its use in endodontics. Quintessence Int. 2015;46(7):575–582. doi:10.3290/j.qi.a33934
17. Lawrence H. The American College of Obstetricians and Gynecologists supports access to women’s health care. Obstet Gynecol. 2015;125(6):1282–1284. doi:10.1097/AOG.0000000000000867
18. Opstrup MS, Malling HJ, Kroigaard M, et al. Standardized testing with chlorhexidine in perioperative allergy - a large single-centre evaluation. Allergy. 2014;69(10):1390–1396. doi:10.1111/all.12466
19. Hu C, Chen X, Huang Y, Chen Y. Co-administration of iRGD with peptide HPRP-A1 to improve anticancer activity and membrane penetrability. Sci Rep. 2018;8(1):2274. doi:10.1038/s41598-018-20715-4
20. Huang YB, Wang XF, Wang HY, Liu Y, Chen YX. Studies on mechanism of action of anticancer peptides by modulation of hydrophobicity within a defined structural framework. Mol Cancer Ther. 2011;10(3):416–426. doi:10.1158/1535-7163.MCT-10-0811
21. Zhao LJ, Huang YB, Gao S, et al. Comparison on effect of hydrophobicity on the antibacterial and antifungal activities of alpha-helical antimicrobial peptides. Sci China Chem. 2013;56(9):1307–1314. doi:10.1007/s11426-013-4884-y
22. Zhao J, Huang YB, Liu D, Chen YX. Two hits are better than one: synergistic anticancer activity of a-helical peptides and doxorubicin/epirubicin. Oncotarget. 2015;6(3):1769–1778. doi:10.18632/oncotarget.2754
23. Hu C, Huang Y, Chen Y. Targeted modification of the cationic anticancer peptide HPRP-A1 with iRGD to improve specificity, penetration, and tumor-tissue accumulation. Mol Pharm. 2019;16(2):561–572. doi:10.1021/acs.molpharmaceut.8b00854
24. Chen Y, Mant CT, Farmer SW, Hancock RE, Vasil ML, Hodges RS. Rational design of α-helical antimicrobial peptides with enhanced activities and specificity/therapeutic index. J Biol Chem. 2005;280(13):12316–12329. doi:10.1074/jbc.M413406200
25. Chen Y, Guarnieri MT, Vasil AI, Vasil ML, Mant CT, Hodges RS. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob Agents Chemother. 2007;51(4):1398–1406. doi:10.1128/AAC.00925-06
26. Abraham T, Lewis RNAH, Hodges RS, McElhaney RN. Isothermal titration calorimetry studies of the binding of a rationally designed analogue of the antimicrobial peptide gramicidin S to phospholipid bilayer membranes. Biochemistry. 2005;44(6):2103–2112. doi:10.1021/bi048077d
27. Zhao J, Hao X, Liu D, Huang Y, Chen Y. In vitro characterization of the rapid cytotoxicity of anticancer peptide HPRP-A2 through membrane destruction and intracellular mechanism against gastric cancer cell lines. PLoS One. 2015;10(9):e0139578. doi:10.1371/journal.pone.0139578
28. Jin WB, Xu C, Cheng Q, et al. Investigation of synergistic antimicrobial effects of the drug combinations of meropenem and 1,2-benzisoselenazol-3(2H)-one derivatives on carbapenem-resistant Enterobacteriaceae producing NDM-1. Eur J Med Chem. 2018;155:285–302. doi:10.1016/j.ejmech.2018.06.007
29. Wang K, Jia F, Dang W, et al. Antifungal effect and action mechanism of antimicrobial peptide polybia-CP. J Pept Sci. 2016;22(1):28–35. doi:10.1002/psc.2835
30. Wiegand I, Hilpert K, Hancock REW. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. 2008;3(2):163–175. doi:10.1038/nprot.2007.521
31. Huang J, Liu Y, Yang L, Zhou F. Synthesis of sulfonated chitosan and its antibiofilm formation activity against E. coli and S. aureus. Int J Biol Macromol. 2019;129:980–988. doi:10.1016/j.ijbiomac.2019.02.079
32. Zheng Z, Tharmalingam N, Liu Q, et al. Synergistic efficacy of Aedes aegypti antimicrobial peptide cecropin A2 and tetracycline against Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2017;61(7):e00686–e00717. doi:10.1128/AAC.00686-17
33. Zhang M, Harashima N, Moritani T, Huang W, Harada M. The roles of ROS and caspases in TRAIL-induced apoptosis and necroptosis in human pancreatic cancer cells. PLoS One. 2015;10(5):e0127386. doi:10.1371/journal.pone.0127386
34. Chen Y, Vasil AI, Rehaume L, et al. Comparison of biophysical and biologic properties of α-helical enantiomeric antimicrobial peptides. Chem Biol Drug Des. 2006;67(2):162–173. doi:10.1111/j.1747-0285.2006.00349.x
35. Pouny Y, Rapaport D, Mor A, Nicolas P, Shai Y. Interaction of antimicrobial dermaseptin and its fluorescently labeled analogues with phospholipid membranes. Biochemistry. 1992;31(49):12416–12423. doi:10.1021/bi00164a017
36. Ehrenstein G, Lecar H. Electrically gated ionic channels in lipid bilayers. Q Rev Biophys. 1977;10(1):1–34.
37. Wade D, Boman A, Wahlin B, et al. All-D amino acid-containing channel-forming antibiotic peptides. Proc Natl Acad Sci U S A. 1990;87(12):4761–4765. doi:10.1073/pnas.87.12.4761
38. Bland JM, De Lucca AJ, Jacks TJ, Vigo CB, All-D-Cecropin B. synthesis, conformation, lipopolysaccharide binding, and antibacterial activity. Mol Cell Biochem. 2001;218(1–2):105–111. doi:10.1023/a:1007293816634
39. Li FJ, Palmer KL. EfrEF and the transcription regulator ChlR are required for chlorhexidine stress response in Enterococcus faecalis V583. Antimicrob Agents Ch. 2018;62(6):e00267–e00318. doi:10.1128/AAC.00267-18
40. Rölla G, Melsen B. On the mechanism of the plaque inhibition by chlorhexidine. J Dent Res. 1975;54(Spec No B):B57–B62. doi:10.1177/00220345750540022601
41. Chu M, Qian L, Zhu M, Yao J, Xu D, Chen M. Circumferential strain rate to detect lipopolysaccharide-induced cardiac dysfunction: a speckle tracking echocardiography study. Quant Imaging Med Surg. 2019;9(2):151–159. doi:10.21037/qims.2018.11.03
42. Kong W, Kang K, Gao Y, et al. Dexmedetomidine alleviates LPS-induced septic cardiomyopathy via the cholinergic anti-inflammatory pathway in mice. Am J Transl Res. 2017;9(11):5040–5047.
43. Rathinam VAK, Zhao Y, Shao F. Innate immunity to intracellular LPS. Nat Immunol. 2019;20(5):527–533. doi:10.1038/s41590-019-0368-3
44. Hu C, Chen X, Huang Y, Chen Y. Synergistic effect of the pro-apoptosis peptide kla-TAT and the cationic anticancer peptide HPRP-A1. Apoptosis. 2018;23(2):132–142. doi:10.1007/s10495-018-1443-1
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.