Back to Journals » The Application of Clinical Genetics » Volume 17
Functional Analysis of BRCA1 3’UTR Variants Predisposing to Breast Cancer
Authors Sierra-Díaz DC, Cabrera R ![]() , Gonzalez-Vasquez LA, Angulo-Aguado M, Llinás-Caballero K
, Gonzalez-Vasquez LA, Angulo-Aguado M, Llinás-Caballero K ![]() , Fonseca-Mendoza DJ
, Fonseca-Mendoza DJ ![]() , Contreras-Bravo NC, Restrepo CM, Ortega-Recalde O, Morel A
, Contreras-Bravo NC, Restrepo CM, Ortega-Recalde O, Morel A ![]()
Received 2 November 2023
Accepted for publication 29 February 2024
Published 23 May 2024 Volume 2024:17 Pages 57—62
DOI https://doi.org/10.2147/TACG.S444546
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Martin Maurer
Diana Carolina Sierra-Díaz,1 Rodrigo Cabrera,1,2 Laura Alejandra Gonzalez-Vasquez,1 Mariana Angulo-Aguado,1,3 Kevin Llinás-Caballero,1,4 Dora Janeth Fonseca-Mendoza,1 Nora Constanza Contreras-Bravo,1 Carlos Martin Restrepo,1 Oscar Ortega-Recalde,1,5 Adrien Morel1
1Center for Research in Genetics and Genomics (CIGGUR), Institute of Translational Medicine (IMT), School of Medicine and Health Sciences, Universidad Del Rosario, Bogotá, Colombia; 2Laboratorio de Biología Molecular y Pruebas Diagnósticas de Alta Complejidad, Fundación Cardioinfantil-Instituto de Cardiología, Bogotá, Colombia; 3Growth Factors, Nutrients and Cancer Group, Molecular Oncology Programme, Centro Nacional Investigaciones Oncológicas (CNIO), Madrid, Spain; 4Institute for Immunological Research, University of Cartagena, Cartagena, Colombia; 5Departamento de Morfología, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá, D.C, Colombia
Correspondence: Adrien Morel, School of Medicine and Health Sciences, Center for Research in Genetics and Genomics (CIGGUR), Institute of Translational Medicine (IMT), Universidad Del Rosario, Cra 24# 63c - 69, Bogotá, 111221, Colombia, Tel +57-6012970200, Email [email protected]
Purpose: Breast Cancer (BC) is the main female cancer diagnosed worldwide, and it has been described that few genes, such as BRCA1, have a high penetrance for this type of cancer. In this manuscript, we were interested in evaluating the effect of 3’UTR variants on BRCA1 expression.
Patients and Methods: To accomplish this objective, Whole Exome Sequencing (WES) data of 400 patients with unselected BC was used to filter variants located in the region of interest of BRCA1 gene, finding two of them (c.*36C>G and c.*369_373del). miRGate and miRanda in silico tools were used to predict microRNA (miRNA) interaction.
Results: The two variants (c.*36C>G, c.*369_373del) were predicted to affect miRNA interaction. After cloning of BRCA1 3’UTR into pMIR-Report vector, the construct was transfected into two BC cell lines (MDA-MB-231 and MCF-7), and the variant c.*36C>G evidenced overexpression of reporter gene luciferase, showing that the transcript was not being degraded by the miRNA in MDA-MB-231 cells.
Conclusion: The variant seems to protect against Triple Negative BC probably due to the expression level of miRNA in this particular cell line (MDA-MB-231). This is consistent with the clinical history of the patients who harbor BC Hormone Receptors positive (HR+).
Keywords: miRNA, 3’UTR variant, breast cancer, BRCA1
Introduction
Breast cancer (BC) is the first cancer in the world in terms of incidence and mortality for women. It has been estimated that 2,261,419 were newly diagnosed and 684,996 died of BC in 20201 and is predicted an increase of approximately 10% of both rates for 2025 (www.gco.iarc.fr). The majority of BC is sporadic (93–95%) and approximately 5–7% of BC is hereditary and associated with family history.2
BC is a disease with genetic and environmental risk factors.3 The main environmental factors are age, alcohol consumption, obesity, and smoking among others.4–7 In the last three decades, 12 genes have been associated with the risk of developing BC, ATM, BRCA1, BRCA2, BRIP1, CDH1, CHEK2, PALB2, PTEN, RAD51C, RAD51D, STK11 and TP53. Some of them are high penetrance genes such as BRCA1 and BRCA2, and others are moderate penetrance genes, for example CHEK2 and BRIP1.8
The two major genes associated with BC are BRCA1 and BRCA2 discovered in 1990 and 1994, respectively. These two genes are located on chromosomes 17 and 13 and are involved in the maintenance of genome stability, especially in the double-strand DNA break repair through homologous recombination. These genes show an autosomal dominant inheritance pattern.9 In fact, the cumulative risk of BC increases with aging among people having BRCA1 and BRCA2 pathogenic variants, reaching a maximum of 72% and 69% at the age of 80 years, respectively.10 BRCA1/2 genes account for the cause of 25% of hereditary BC cases and in the general population, the prevalence of pathogenic variants in BRCA1/2 genes has been estimated as 1 in 300 and 1 in 800, respectively.11 In the Ashkenazi Jewish ethnic group, the prevalence is higher than the general population, due to endogamic behavior, and estimated as 1 in 40.12 It has been demonstrated that BRCA1 pathogenic variants are more frequent in Triple Negative BC (TNBC) compared to BRCA2 gene which are more prevalent in less aggressive BC subtypes.13 Nowadays, it has been described around 4900 pathogenic variants in BRCA1/2 genes (BRCA Exchange https://brcaexchange.org/) and the majority of them are located in coding regions. Few pathogenic variants are reported in non-coding regions including splicing regions (70 pathogenic variants) or UTR regions (0 pathogenic variant).
Non-coding regions have paramount importance in gene expression regulation. In particular, 3’UTR interacts with several types of miRNAs, which are a type of non-coding RNAs (~22 nucleotides) that interact with 3’UTR and can inhibit the translation or/and promote mRNA degradation. Briefly, the miRNAs are transcribed by RNA pol II or III in pri-miRNA, then they are processed into short 70-nt stem-loop RNA, pre-miRNAs by the Microprocessor complex (Drosha enzyme – DiGeorge syndrome critical region 8 gene (DGCR8)). These RNAs are then exported by exportin-5 from nucleus to cytoplasm and processed into double-strand miRNAs by the RNAse II enzyme, Dicer. The duplex is loaded onto Argonaute protein (AGO) and RISC complex (RNA inducing silencing complex). The strand less thermodynamically stable is removed and the complex RISC-miRNA is able to recognize 3’UTR of target mRNA. The seed region, a sequence spanning from position 2 to 8 at the 5’ end of the miRNA, recognizes the miRNA response element (MRE) in the target mRNA.14,15 A variant in the 3’UTR or in the miRNA can inhibit the mRNA-miRNA interaction or create a new interaction with a different miRNA that has been previously described in breast and ovarian cancer.16–19
In this study, we performed the functional analysis of germline variants located on 3’UTR of BRCA1 gene. This study is the first analysis of these type of variants described in Colombian patients diagnosed with unselected BC.
Materials and Methods
Patients
Four hundred unselected BC, regarding cancer family history, female patients were aged over 18 years old, with a diagnosis of invasive BC confirmed histopathologically, without personal and family history of germline BC risk pathogenic variants. These women were attended to Fundación CardioInfantil, Hospital Universitario de Mederi, Bogota, Colombia and different cancer centers located throughout Colombia (Bucaramanga, Cali, Medellin, Manizales, Pereira).
This study was performed in compliance with the Helsinki Declaration and was approved by both, Fundación Cardioinfantil–Instituto de Cardiología and Universidad del Rosario Ethics Committee (approval numbers: 402,018 7-11-2018, DVO005 1805-CV1469 3-12-2021, Pfizer: WI241988 – Investigator initiate research, independent review board: 28–08-2018, GF1147 2018). All patients approved the informed consent and were part of a previously described study https://doi.org/10.1158/1538-7445.SABCS21-P3-07-05. The study was financially supported by the grant “Capital Semilla” of the University of Rosario, IV-FCS036.
Variants Analysis
Germline variants located on 3’UTR of BRCA1 gene (~500pb) were filtered by population frequencies, taking into consideration a Minor Allele Frequency (MAF) ≤1% exomes in the gnomAD database (https://gnomad.broadinstitute.org v2.1.1). Levels of expression of miRNAs in breast and/or BC were used to use in silico tools miRGate (http://mirgate.bioinfo.cnio.es) and to filter germline variants (supplementary Tables 1–3). miRanda package v1.9 on Linux (https://cbio.mskcc.org/miRNA2003/miranda.html) was used to predict the miRNA-3’UTR interaction.
None of the women, harboring the c.*36C>G variant, have pathogenic/likely pathogenic germline variants in the ORF of 10 genes with known penetrance for BC (ATM, BARD1, BRCA1, BRCA2, CHEK2, CDH1, PALB2, TP53, RAD51C, and RAD51D).
pMIR-Report Construction
The 3’UTR of BRCA1 was amplified from patient DNA using GoTaq Master Mix (Promega, cat#M7122) with the following primers (huBRCA1_3UTR_For:CCCACTAGTCTGCAGCCAGCCACAGGTAC, huBRCA1_3UTR_Rev: CCCAAGCTTCGATCCCAAGCACTCTCCTTC), PCR products were cloned into pCR4-TOPO vector (Invitrogen, cat# 45–0030), and then cloned into the pMIR-ReportTM Luciferase vector (Invitrogen, cat#AM5795) through enzyme digestion (SpeI, HindIII, New England Biolabs, cat# R0133S and # R0104S).
Cell Culture
MDA-MB-231 and MCF-7 cells (ATCC, cat#HTB-26, cat#HTB-22) were maintained in culture with DMEM-F12 (10% FBS, 1%P/S) at 37°C and 5% of CO2.
Luciferase Assay
Cells were transfected through Fugene 6 (Promega, cat#E2692) with 1μg of vector and co-transfected with 30ng of Renilla reporter vector and incubated for 48h. Cells were lysed with Passive Buffer of Dual-Glo luciferase assay (cat#E2980) among the manufacturer’s recommendations. Briefly, 20µL of cell extracts were used to the read of the luminescence by the GloMax system (Promega). Luciferase was read at 570nm through LARII reagent and Renilla at 480nm through STOP&Glo reagent. Three experiments were realized, and RLUs were normalized by experiment and compared.
GEO Database
The GSE146477 reports an array of miRNA expression and the miR-99a expression was compared in MDA-MB-231 and MCF-7 cells.
Statistical Tests
The replicates were compared by Mann–Whitney test using Prism 10 (https://www.graphpad.com/features).
Results
On the 400 women diagnosed with BC, 2 germline variants were observed on 3’UTR of BRCA1 gene with a MAF ≤1%. The variants c.*36C>G and c.*369_373del were tested on miRGate and were located on two miRNA interaction sites, miR-99a-3p and miR-26a-2-3p, respectively (supplementary Tables 1 and 2). The first variant was observed in three patients and the second in only one patient, all in heterozygote state. The c.*36C>G variant is located on the complementary interaction site of miR-99a-3p (Figure 1A) and the c.*369_373del variant is located on the miR-26a-2-3p interaction seed sequence (Figure 1B). These variants were then tested on two cell models of breast cancer, MDA-MB-231 (TNBC cells) and MCF7 cells (human breast cancer cell line with estrogen, progesterone and glucocorticoid receptors) (Figure 1C and D, respectively). One variant, which has not been detected by WES, c.*421G>T was found and included in the experiments. Higher (Relative Luciferase Units) RLU were observed for the c.*36C>G variant compared to the WT BRCA1 3’UTR in the MDA-MB-231 cells (p-value 0.0286) (Figure 1C) but not in the MCF-7 cell line (Figure 1D). The other variants had no difference in the RLU compared to WT. The expression level of the miR-99a-3p was verified and compared between both BC cell lines from the GEO database (GSE146477) and the expression level of miR99a-3p is higher in MDA-MB-231 in comparison with MCF-7 (supplementary Figure 1).
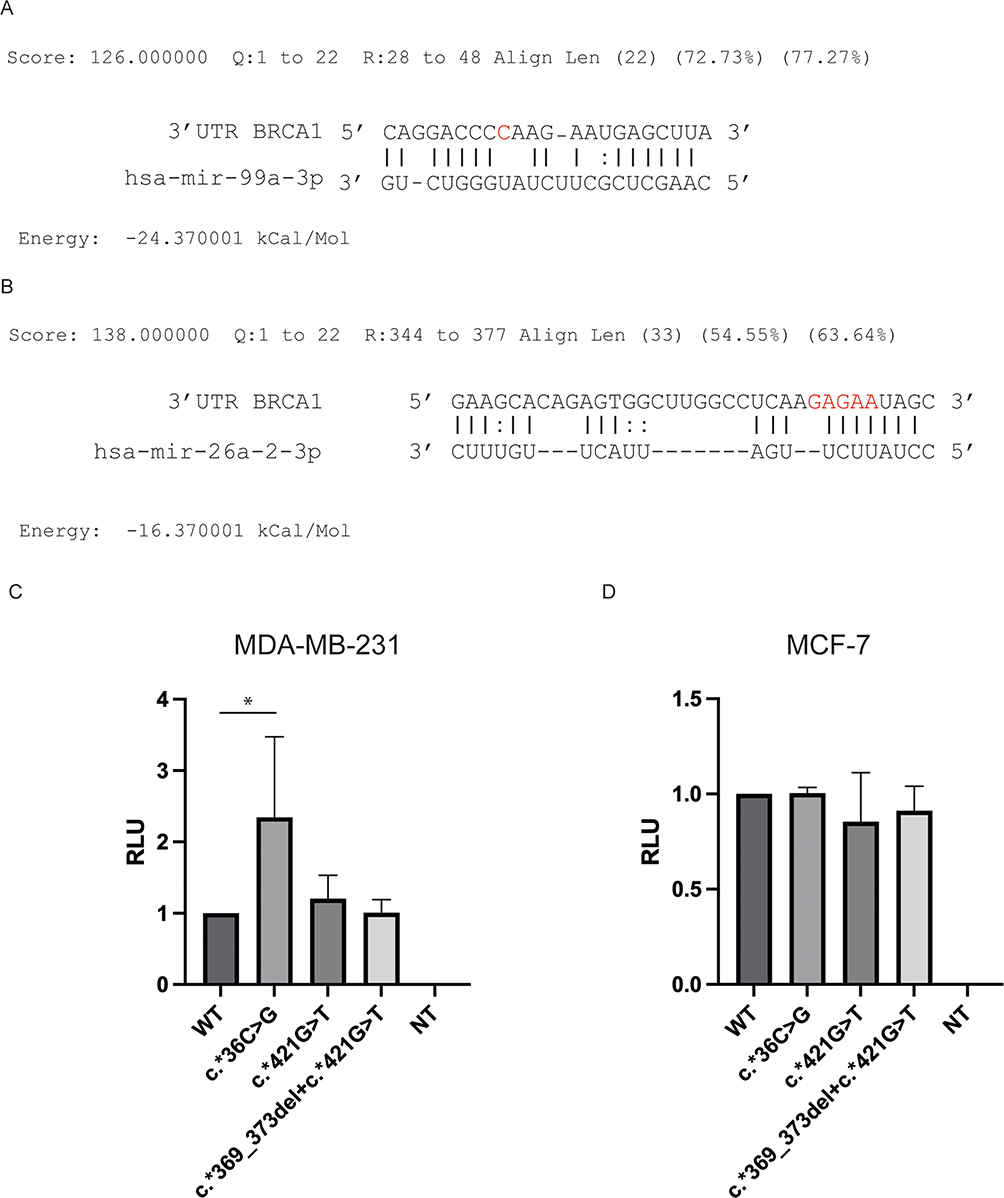 |
Figure 1 Involvement of variants on 3’UTR on BRCA1 expression. (A) Interaction of miR-99a-3p with BRCA1 3’UTR in presence of c.*36C>G variant (red) (B) Interaction of miR-26a-2-3p with BRCA1 3’UTR in presence of c.*369_373del variant (red) (C and D) Luciferase assay representing the transfection of pMIR-Report with the different variants in MDA-MB-231 and MCF-7 cell lines, respectively. Statistical significance *p-value<0.05. |
Discussion
BC is the cancer most frequently diagnosed in women and causes 15% of cancer-related deaths in the same population. BC is a multifactorial disease, with a high percentage of unexplained heritability. It has been well described that approximately 10% of BC are hereditary and only the 5.6% of all BC cases are caused by germline pathogenic variants on 12 BC predisposition genes.8,20 At present, it is important to explore new mechanisms capable of explaining the cancer development and the heritability of this disease.
miRNAs are small non-coding RNA with an important role in the regulation of gene expression. It has been described that these miRNAs can be involved in different molecular processes in BC, such as apoptosis, epithelial-mesenchymal transition, and angiogenesis, among others.21–23 The miRNAs can exert their function via their union on the 3’UTR of the gene. It has been proposed that some miRNAs can be biomarkers of breast, prostate and ovarian cancer.24,25 It has been described that some miRNAs have an oncogenic and tumor suppressive function.26,27 The dysregulation of miRNAs expression could explain in part the dysregulation of some genes involved in BC.
A few years ago, different studies have shown the impact of 3’UTR variants on the expression and susceptibility to cancer development.18,28,29 In this study, we were interested in exploring the impact of 3’UTR variants on the gene expression of BRCA1. In fact, BRCA1, as well as BRCA2, have an important role in the biology of the cell, especially in tumor microenvironment in BC. The function of BRCA1/2 consists essentially in the maintenance of genome stability through participation in DNA repair processes including homologous recombination.30 Pathogenic variants on this gene have a penetrance of approximately 69% up to the age of 80 years and seem to exhibit an autosomal dominant inheritance.10 On the BRCA Exchange database (brcaexchange.org) there are reported more than 2000 pathogenic variants located on coding regions and splicing sites of BRCA1/2 genes. All 3’UTR variants are probably considered non-pathogenic, according to the current ACMG/AMP guidelines, but these variants could be considered risk factors as described by Zhang and collaborators.18 In this study, we observed that the c.*36C>G variant evidenced an overexpression of the reporter gene luciferase in MDA-MB-231 cells, a TNBC cell line. This finding is consistent with the clinicopathological characteristics of the BC in the three patients harboring this variant, who present a positive hormone receptor (HR+) subtype (supplementary Table 4). This variant seems to protect against the development of TNBC, which is a subtype of BC with a worse prognosis compared with HR+ BC. Expression of the miR-99a was validated in the two cell lines, by the GEO database (GSE146477), and we observed that the miR-99a-3p is overexpressed in MDA-MB-231 cell line in comparison with MCF-7 cell line (1 vs 0.73 fold-change). The difference in miRNA expression could explain the lack of RLU difference between the variants and WT in MCF-7 cells.
It has been demonstrated that miR-99a correlates negatively with inflammation in adipose tissue in humans. It has been well described that obesity/overweight is a pro-inflammatory status, and a BC risk factor. The mechanism, involved in obesity and hormonal changes is related to adipokines, such as leptin, adiponectin, and resistin among others. The hormonal changes in BC patients with obesity/overweight also have oxidative stress in the tumoral microenvironment regulated by the adipokines, supporting carcinogenesis in breast tissue.31–34 The MCF-7 cell model could explain this situation since it expresses HR and a lower level of miR-99a. 2/4 of the patients harboring the 3’UTR variant (c.*36C>G) have a BMI >25 kg/m2 (supplementary Table 4) this finding could reinforce the hypothesis of obesity/overweight role in breast carcinogenesis.
In summary, the c.*36C>G variant does not allow the degradation of BRCA1, as a protector effect in TNBC model, and not in the HR+ BC model, probably due to differential miRNA expression levels.
We suggest that the scientific community should analyze the impact of 3’UTR variants on gene expression, given that in a model of multifactorial disease, each variants have a little additive effect that, when added, can permit the development of the phenotype. Furthermore, 3’UTR variants allow gene expression dysregulation in different diseases with a Mendelian inheritance pattern.35,36 Evolution of technologies with bioinformatics algorithm like miRGate or miRanda could help to expand the knowledge in genetics and allow us to propose new molecular mechanisms involved in different diseases including BC.
Data Sharing Statement
Data supporting this study are available under request.
Funding
This study was financed by the grant “Capital Semilla” of the Universidad del Rosario, IV-FCS036.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Melchor L, Benítez J. The complex genetic landscape of familial breast cancer. Hum Genet. 2013;132(8):845–863. doi:10.1007/s00439-013-1299-y
3. Rudolph A, Chang-Claude J, Schmidt MK. Gene–environment interaction and risk of breast cancer. Br J Cancer. 2016;114(2):125–133. doi:10.1038/bjc.2015.439
4. Benz CC. Impact of aging on the biology of breast cancer. Crit Rev Oncol Hematol. 2008;66(1):65–74. doi:10.1016/j.critrevonc.2007.09.001
5. Catsburg C, Miller AB, Rohan TE. Active cigarette smoking and risk of breast cancer. Int J Cancer. 2015;136(9):2204–2209. doi:10.1002/ijc.29266
6. García-Estévez L, Cortés J, Pérez S, et al. Obesity and breast cancer: a paradoxical and controversial relationship influenced by menopausal status. Front Oncol. 2021:11. doi:10.3389/fonc.2021.705911
7. McDonald JA, Goyal A, Terry MB. Alcohol intake and breast cancer risk: weighing the overall evidence. Curr Breast Cancer Rep. 2013;5(3):208–221. doi:10.1007/s12609-013-0114-z
8. Yoshimura A, Imoto I, Iwata H. Functions of breast cancer predisposition genes: implications for clinical management. Int J Mol Sci. 2022;23(13):7481. doi:10.3390/ijms23137481
9. Varol U. BRCA genes: BRCA 1 and BRCA 2. J BUON. 2018;23(4):862–866.
10. Kuchenbaecker KB, Hopper JL, Barnes DR, et al. Risks of breast, ovarian, and contralateral breast cancer for BRCA1 and BRCA2 mutation carriers. JAMA. 2017;317(23):2402–2416. doi:10.1001/jama.2017.7112
11. Kast K, Rhiem K, Wappenschmidt B, et al. Prevalence of BRCA1/2 germline mutations in 21 401 families with breast and ovarian cancer. J Med Genet. 2016;53(7):465–471. doi:10.1136/jmedgenet-2015-103672
12. Daly MB, Pal T, Berry MP, et al. Genetic/familial high-risk assessment: breast, ovarian, and pancreatic, version 2.2021, NCCN clinical practice guidelines in oncology. J Natl Compr Canc Netw. 2021;19(1):77–102. doi:10.6004/jnccn.2021.0001
13. Chen H, Wu J, Zhang Z, et al. Association between BRCA status and triple-negative breast cancer: a meta-analysis. Front Pharmacol. 2018;9:909. doi:10.3389/fphar.2018.00909
14. Chatterjee S, Pal JK. Role of 5′- and 3′-untranslated regions of mRNAs in human diseases. Biol Cell. 2009;101(5):251–262. doi:10.1042/BC20080104
15. Shang R, Lee S, Senavirathne G, Lai EC. microRNAs in action: biogenesis, function and regulation. Nat Rev Genet. 2023;1–18. doi:10.1038/s41576-023-00611-y
16. Garcia AI, Buisson M, Damiola F, et al. Mutation screening of MIR146A/B and BRCA1/2 3′-UTRs in the GENESIS study. Eur J Hum Genet. 2016;24(9):1324–1329. doi:10.1038/ejhg.2015.284
17. Mir R, Javid J, Al Balawi IA, et al. A germline mutation in the BRCA1 3’UTR variant predicts susceptibility to breast cancer in a Saudi Arabian population. Asian Pac J Cancer Prev. 2018;19(3):859–866. doi:10.22034/APJCP.2018.19.3.859
18. Zhang L, Liu Y, Song F, et al. Functional SNP in the microRNA-367 binding site in the 3′UTR of the calcium channel ryanodine receptor gene 3 (RYR3) affects breast cancer risk and calcification. Proc Natl Acad Sci U S A. 2011;108(33):13653–13658. doi:10.1073/pnas.1103360108
19. Santana Dos Santos E, Lallemand F, Burke L, et al. Non-coding variants in BRCA1 and BRCA2 genes: potential impact on breast and ovarian cancer predisposition. Cancers. 2018;10(11):453. doi:10.3390/cancers10110453
20. Harbeck N, Penault-Llorca F, Cortes J, et al. Breast cancer. Nat Rev Dis Primers. 2019;5(1):66. doi:10.1038/s41572-019-0111-2
21. Ghafouri-Fard S, Khanbabapour Sasi A, Abak A, et al. Contribution of miRNAs in the pathogenesis of breast cancer. Front Oncol. 2021:11. doi:10.3389/fonc.2021.768949
22. Hussen BM, Abdullah ST, Rasul MF, et al. MicroRNAs: important players in breast cancer angiogenesis and therapeutic targets. Front Mol Biosci. 2021;8:764025. doi:10.3389/fmolb.2021.764025
23. Tucci P. The role of microRNAs in cancer: functions, biomarkers and therapeutics. Cancers. 2022;14(4):872. doi:10.3390/cancers14040872
24. Galvão-Lima LJ, Morais AHF, Valentim RAM, Barreto EJS. miRNAs as biomarkers for early cancer detection and their application in the development of new diagnostic tools. Biomed Eng Online. 2021;20:21.
25. Zou R. Development and validation of a circulating microRNA panel for the early detection of breast cancer. Br J Cancer. 2022;126(3):472–481. doi:10.1038/s41416-021-01593-6
26. Kent OA, Mendell JT. A small piece in the cancer puzzle: microRNAs as tumor suppressors and oncogenes. Oncogene. 2006;25(46):6188–6196. doi:10.1038/sj.onc.1209913
27. Loh H-Y. The regulatory role of MicroRNAs in breast cancer. Int J Mol Sci. 2019;20(19):4940. doi:10.3390/ijms20194940
28. Skeeles LE, Fleming JL, Mahler KL, Toland AE. The impact of 3’UTR variants on differential expression of candidate cancer susceptibility genes. PLoS One. 2013;8(3):e58609. doi:10.1371/journal.pone.0058609
29. Zhang N, Huang D, Jiang G, et al. Genome-Wide 3’-UTR single nucleotide polymorphism association study identifies significant prostate cancer risk-associated functional loci at 8p21.2 in Chinese population. Adv Sci. 2022;9(23):2201420. doi:10.1002/advs.202201420
30. Miklikova S, Trnkova L, Plava J, et al. The role of BRCA1/2-mutated tumor microenvironment in breast cancer. Cancers. 2021;13(3):575. doi:10.3390/cancers13030575
31. Bhardwaj P. Obesity promotes breast epithelium DNA damage in women carrying a germline mutation in BRCA1 or BRCA2. Sci Transl Med. 2023;15(684):eade1857. doi:10.1126/scitranslmed.ade1857
32. Verras G-I, Tchabashvili L, Chlorogiannis -D-D, Mulita F, Argentou M-I. Updated clinical evidence on the role of adipokines and breast cancer: a review. Cancers. 2023;15(5):1572. doi:10.3390/cancers15051572
33. Hanusek K, Karczmarski J, Litwiniuk A, et al. Obesity as a risk factor for breast cancer—the role of miRNA. Int J Mol Sci. 2022;23(24):15683. doi:10.3390/ijms232415683
34. Jaiswal A, Reddy SS, Maurya M, Maurya P, Barthwal MK. MicroRNA-99a mimics inhibit M1 macrophage phenotype and adipose tissue inflammation by targeting TNFα. Cell Mol Immunol. 2019;16(5):495–507. doi:10.1038/s41423-018-0038-7
35. Simon D, Laloo B, Barillot M, et al. A mutation in the 3’-UTR of the HDAC6 gene abolishing the post-transcriptional regulation mediated by hsa-miR-433 is linked to a new form of dominant X-linked chondrodysplasia. Hum Mol Genet. 2010;19(10):2015–2027. doi:10.1093/hmg/ddq083
36. Wang Y, Du X, Zhou Z, et al. A gain-of-function ACTC1 3’UTR mutation that introduces a miR-139-5p target site may be associated with a dominant familial atrial septal defect. Sci Rep. 2016;6(1):25404. doi:10.1038/srep25404
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Management Strategies of Breast Cancer Patients with BRCA1 and BRCA2 Pathogenic Germline Variants
Edaily S, Abdel-Razeq H
OncoTargets and Therapy 2022, 15:815-826
Published Date: 27 July 2022
Guideline-Based, Multi-Gene Panel Germline Genetic Testing for at-Risk Patients with Breast Cancer
Abdel-Razeq H, Abujamous L, Al-Azzam K, Abu-Fares H, Bani Hani H, Alkyam M, Sharaf B, Elemian S, Tamimi F, Abuhijla F, Edaily S, Salama O, Abdulelah H, Daoud R, Abubaker M, Al-Atary A
Breast Cancer: Targets and Therapy 2023, 15:1-10
Published Date: 13 January 2023
Noncoding RNAs, Vital Players in Breast Cancer Metastasis
Kang Y, Wang X, Liu Y, Cao J, Lin J, Qiao G
Breast Cancer: Targets and Therapy 2025, 17:1463-1492
Published Date: 30 December 2025
Advancing Germline Genetic Testing for Breast Cancer in Resource-Restricted Settings: Evidence, Barriers, and a Practical Roadmap for Implementation
Abdel-Razeq H
Breast Cancer: Targets and Therapy 2026, 18:618067
Published Date: 24 June 2026