Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Finerenone Ameliorates High Glucose-Induced Podocytes Epithelial-Mesenchymal Transition Through the Regulation of Krüppel-Like Factor 5 in Diabetic Nephropathy
Authors Shu J ![]() , Chen D, Chen W, Zhang X, Wang S, Chong N, Sun Z, Wang Q, Sun J, Xu Y
, Chen D, Chen W, Zhang X, Wang S, Chong N, Sun Z, Wang Q, Sun J, Xu Y ![]()
Received 26 October 2024
Accepted for publication 30 January 2025
Published 27 February 2025 Volume 2025:18 Pages 637—651
DOI https://doi.org/10.2147/DMSO.S503133
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Jianqiang Shu,1 Dandan Chen,1 Wenzhen Chen,2 Xinyu Zhang,1 Simeng Wang,1 Nannan Chong,3 Zhikang Sun,3 Qinglian Wang,3 Jingshu Sun,4 Ying Xu1
1Department of Nephrology, Shandong Provincial Hospital, Shandong University, Jinan, Shandong, People’s Republic of China; 2Clinic Medical College, The Second Hospital of Shandong University, Jinan, Shandong, People’s Republic of China; 3Department of Nephrology, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, People’s Republic of China; 4Department of Nephrology, Weifang People’s hospital, Weifang, Shandong, People’s Republic of China
Correspondence: Ying Xu, Department of Nephrology, Shandong Provincial Hospital, Shandong University, Jinan, Shandong, People’s Republic of China, Email [email protected]
Introduction: Diabetic nephropathy (DN) could impair the function of the glomerular filtration barrier by damaging the podocytes. Extant research has demonstrated that aldosterone plays a crucial role in this progression. Finerenone is a novel, high-selective mineralocorticoid receptor antagonist that has been demonstrated to be efficacious in renal protection in DN patients, albeit with an unclear underlying mechanism.
Methods: Podocytes were stimulated with RPMI 1640 medium containing different concentrations of glucose and treated with finerenone to evaluate the protective effect of finerenone on podocytes in high glucose environment. Intraperitoneal injection of STZ was used to induce diabetic nephropathy rats and intragastric administration with finerenone or vehicles, and the changes of renal function, renal pathological changes and renal tissue protein expression were assayed.
Results: Cell experiment showed that high glucose could induce epithelial–mesenchymal transition (EMT). After finerenone treatment, we accessed the EMT-related protein and found EMT was reversed. Besides, the cell migration capacity and cytoskeleton were also ameliorated. To further explore the mechanism, we found that finerenone could upregulate the expression of Krüppel-like factor 5 (KLF5) which was downregulated in a high glucose environment. After the silence of KLF5 in the presence of finerenone, the rescue experiment showed the protective function of finerenone is counteracted by KLF5. In animal experiment, after the treatment of finerenone, the level of blood creatinine decreased compared with the DN group while blood urea nitrogen (BUN) and potassium showed no significant difference. The pathological alterations of the treatment group also got ameliorated. Finerenone could normalize the level of EMT-related protein, nephrin, and KLF5 of kidney tissue in DN rats.
Conclusion: Our results suggest that finerenone could alleviate high glucose-induced podocyte EMT via regulating KLF5. Further investigation is warranted to elucidate the precise underlying mechanism.
Keywords: finerenone, DN, EMT, KLF5
Introduction
Nowadays, diabetes become a huge health problem in human society: in 2021, the International Diabetes Federation (IDF) estimated that 537 million people have diabetes worldwide, and by 2045, this number is expected to rise to 783 million.1 Diabetics often suffer a lot of complications, such as hypertension, cerebrovascular disease, cardiovascular disease, eye disease, et al. Besides, these patients’ leading causes of death are the vascular complications of the macrovascular system (cardiovascular disease) and microvascular system (diabetic kidney disease).2,3
Diabetic nephropathy (DN) is able to cause serious kidney damage and is the leading cause of end-stage renal disease nowadays. Diabetic nephropathy is caused by a variety of causes, including genetics, renal hemodynamic abnormalities, metabolic changes, hypertension, and abnormal secretion of vasoactive substances.4,5 It presents clinically with proteinuria, hypertension, and progressive decline in renal function.6 Diabetic nephropathy now become an important health problem that not only causes health damage but also causes quite important social finance issues. Most patients with diabetic nephropathy have glomerulosclerosis, kidney inflammation intensified fibrosis, and many other changes.7 Although there are many treatments for diabetic nephropathy, the kidney damage of patients is still irreversible.8
Renal interstitial fibrosis is a common pathway for most chronic kidney diseases to progress to the end stage. Renal fibrosis leads to a decrease in functional renal parenchyma, resulting in impaired renal function.9 What is more serious is that there is no effective treatment for renal fibrosis. Epithelial to mesenchymal transition (EMT) has been shown to be an important factor in inducing renal fibrosis and causing kidney damage.10–12 Therefore EMT might be a new target in the treatment of Diabetic nephropathy.13
Mineralocorticoid receptors (MR) play an important role in kidney damage, and both the cells of the glomeruli and tubules contain mineralocorticoid receptors.14,15 Finerenone is a novel, high-selective mineralocorticoid receptor antagonist, which was confirmed to have the ability to protect against kidney damage in CKD, especially in diabetic nephropathy.15 However, the specific role of finerenone for kidney protection is unclear. In this article, we proved that finerenone could inhibit EMT by restoring the expression of KLF5, thus achieving the effect of protecting the kidneys of diabetic patients, which might provide a new treatment idea in diabetic nephropathy.
Materials and Methods
Chemicals and Reagents
Finerenone was purchased from Biochem Safebuy Bio-Technology company. A 10 mm stocking solution was dissolved and stored at −80°C. RPMI 1640 medium (Contains Glycine (10mg/L), L-Cystine dihydrochloride (65mg/L), L-Glutamine (300mg/L), L-Isoleucine (50mg/L), L-Leucine (50mg/L), L-Lysine hydrochloride (40mg/L), L-Methionine (15mg/L), L-Phenylalanine (15mg/L), L-Serine (30mg/L), L-Threonine (20mg/L), L-Tryptophan (5mg/L), L-Tyrosine disodium salt dihydrate (29mg/L), L-Valine (20mg/L), L-Arginine (200mg/L), L-Asparagine monohydrate (56.82mg/L), L-Aspartic acid (20mg/L), L-Glutamic Acid (20mg/L), L-Histidine (15mg/L), L-Hydroxyproline (20 mg/L), L-Proline (20mg/L), Magnesium Sulfate (48.84mg/L), Potassium Chloride (400mg/L), Sodium Bicarbonate (2000mg/L), Sodium Chloride (6000mg/L), Calcium Nitrate tetrahydrate (100mg/L), Sodium Phosphate dibasic (800mg/L), Choline chloride (3mg/L), D-Calcium pantothenate (0.25mg/L), Folic Acid (1mg/L), i–Inositol (35mg/L), Niacinamide (1mg/L), Pyridoxine hydrochloride (1mg/L), Riboflavin (0.2mg/L), Thiamine hydrochloride (1mg/L), Biotin (0.2mg/L), Para-Aminobenzoic Acid (1mg/L), Vitamin B12 (0.005mg/L), D-(+)-Glucose (2000mg/L), Phenol Red (5mg/L), Glutathione (1mg/L), no HEPES) and fetal bovine serum (FBS) were purchased from Gibco, Invitrogen (Carlsbad, CA, USA). Trypsin/EDTA was purchased from HyClone (Logan, UT, USA). WST-8 cell proliferation and cytotoxicity assay kit was purchased from Dojindo (Kumamoto, Japan). The phalloidin staining reagent was bought from Sigma-Aldrich company. Nile acid was purchased from Macklin Inc. siRNA specific for KLF-5 was designed and provided by Genomeditech company (Shanghai). Primary antibodies were detailed as below: Anti-E-cadherin (proteintech, China), Anti-N-cadherin (proteintech, China), Anti-Vimentin (proteintech, China), Anti-KLF5 (proteintech, China), Anti-Nephrin (Abcam, Cambridge, MA, UK). All other materials and reagents were endotoxin-free and supplied by central lab of Shandong Provincial Hospital.
Animal Experiments
The rats were studied with permission from animal protection authorities and the Animal Research Center of Shandong Provincial Hospital (ethics approval number: No.2022–142; ethics committee: the Institution Animal Care and Use Committee at the Shandong Provincial Hospital). Healthy male SD rats aged six weeks were purchased from Beijing Vital River Laboratory Animal Technology (China). All rats were accommodated for one week in a quiet room at 22–26°C with a 12-hour light/dark cycle. Streptozotocin was dissolved in 0.1mol/L citric acid buffer, intraperitoneal injection in rats at a dose of 40mg/kg body weight two times at a time interval of 5 days. Control rats received the same amount of citrate buffer. One week after the last injection, caudal vein blood samples were tested for random blood glucose two times. Rats with random blood glucose ≥16.7mmol/L twice were considered diabetes. One rat was excluded because its random blood glucose did not meet the criteria. All diabetes rats were randomly divided into 4 groups: STZ+DMSO group (n = 3), STZ+Finerenone10 (10mg/kg/d) group (n = 3), STZ+Finerenone3 (3mg/kg/d) group (n = 3), and STZ+Valsartan (40mg/kg/d) (n = 3). And chose 3 healthy rats which intraperitoneal injection of 0.1mol/L citric acid buffer as the control group. Finerenone was dissolved in DMSO to be stored and was intragastrical administered at a daily dose of 10mg/kg or 3mg/kg body weight. Valsartan was preserved in DMSO and was intragastrical administered at a daily dose of 40mg/kg based on previous studies. Control group was intragastrical administered DMSO at a same dose. Weight and blood glucose were measured periodically during gavage. After 8 weeks of intragastric administration, all rats were anesthetized intraperitoneally with Sodium pentobarbital (150mL/kg) and sacrificed. Serum was collected, then kidneys were collected for subsequent analysis.
Cell Culture
Human podocytes were obtained from professor Fan Yi (Department of Pharmacology, Shandong University School of Medicine) which have been approved by Ethics Committee of Biomedical Research in Shandong Provincial Hospital. Podocytes were cultured in RPIM 1640 medium with 10% FBS and 10 U/mL interferon γ and placed in a humidified incubator with 5% CO2 at 33°C to induce proliferation. When ready for treatment, cells are transferred to 37°C, 5%CO2, and interferon-γ free incubators to induce differentiation for at least 14 days. In the next experiment, different mediums were designed and cells were incubated with low glucose (LG, 5.5mm glucose), and middle glucose (15mM glucose). High osmotic pressure (Man,30mM mannitol), high glucose (HG,30mM glucose), high glucose + different concentrations of finerenone.
Western Blot
First, total cell proteins were lysed with RIPA lysate supplemented with 1% Cocktail. Proteins were then isolated by using 10%SDS-PAGE. The protein was then transferred to polyvinylidene fluoride (PVDF) membranes, which were sealed with skim milk at room temperature for one hour. Then the membrane was incubated with primary antibody at 4°C overnight. On the second day, membranes were washed with TBST and incubated with HRP-conjugated anti-rabbit/rat IgG secondary antibodies for 1h. After all these steps, the membrane is examined by using the ECL system.
Phalloidin Staining
Podocyte was inoculated on the glass coverslips, after a series of treatments, the coverslips were removed, cleaned with PBS 3 times, fixed with 4% paraformaldehyde for 15 min, then cleaned with PBS again for 3 times, and soaked coverslips with 0.3% Triton. The slides were then stained with Phalloidin for 30 minutes in the dark and photographed.
Transwell Assay
Podocytes were grown in 1640 medium containing 10% FBS for 24 h. The cells were digested with trypsin and the sample was centrifuged (900rpm, 4 min). The supernatant was discarded and the cells were resuspended in serum-free 1640 medium, and the number of inoculated cells was adjusted to 5×10*5/mL.1640 medium containing 10% fetal bovine serum (800ul) was added into the lower chamber of a 24-well plate, different stimuli were added, and the cells were cultured in a sterile cell incubator for 24h. The chambers were removed, washed 3 times with PBS, fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The chamber was natural shade drying and analyzed with a microscope.
Immunofluorescence Assay
Podocyocytes were inoculated in glass coverslips for 24 hours and given different stimuli. After 48h, used PBS washing coverslips for three times, fixed with 4% paraformaldehyde, and incubated with 0.1%TritonX-100 at 37°C for 15 min. The goat serum was blocked for 1h, and the coverslip was incubated in KLF5 Vimentin or E-Cadherin primary antibody (incubated at 4°C overnight). On the second day, after washing with PBS three times, the coverslips were incubated with anti-rabbit IgG or anti-mouse IgG in the dark at 37°C for 1h. Washed with PBS three times, incubated with DAPI for 10 min, and observed under a microscope.
WST-8 Assays
WST-8 proliferation and cytotoxicity assay kit was used to detect the cell viability of different groups. Simply put, the cells were grown in 96-well plates and given different stimuli. After 48h of treatment, the medium was changed to 90μL fresh serum-free medium containing 10μLWST-8 solution. Then, depending on the color of the medium, the dish is placed in an incubator for another 2–4 hours. The absorbance of the medium is then measured by a microplate reader with a wavelength of 450nm. Then calculate cell viability based on absorbance.
Results
High Concentrations of Glucose Could Facilitate the EMT of Human Podocytes
Human podocytes were placed in a humidified incubator with 5% CO2 at 33°C to induce proliferation (Figure S1A–C). To confirm the effect of a high glucose environment on podocytes, we used 15 and 30 mmol/L glucose in a culture medium to stimulate podocytes, to avoid the interference of osmolality in a high glucose environment, we used mannitol, configured as a medium with equal osmolality as control group. In terms of results, the high glucose environment leads to alterations in EMT-associated proteins by Western blot (Figure 1A). Compared with the low glucose group (5.5 mmol/L), in the cells of 30 mmol/L glucose group, the expression of vimentin and N-cadherin are increased and the expression of E-cadherin are decreased which means the podocytes have undergone EMT induced by high glucose. While the expression of EMT-related protein of the mannitol group showed no significant difference with the low glucose group (5.5 mmol/L) proved that those changes have no relatiion with high osmolality (Figure 1B–D). Cell scratching experiments found that the cell migration capacity of the 30mmol/L glucose group was increased significantly (Figure 1E). All of those experimental results could confirm the conclusion that high concentration of glucose can facilitate EMT of human podocytes.
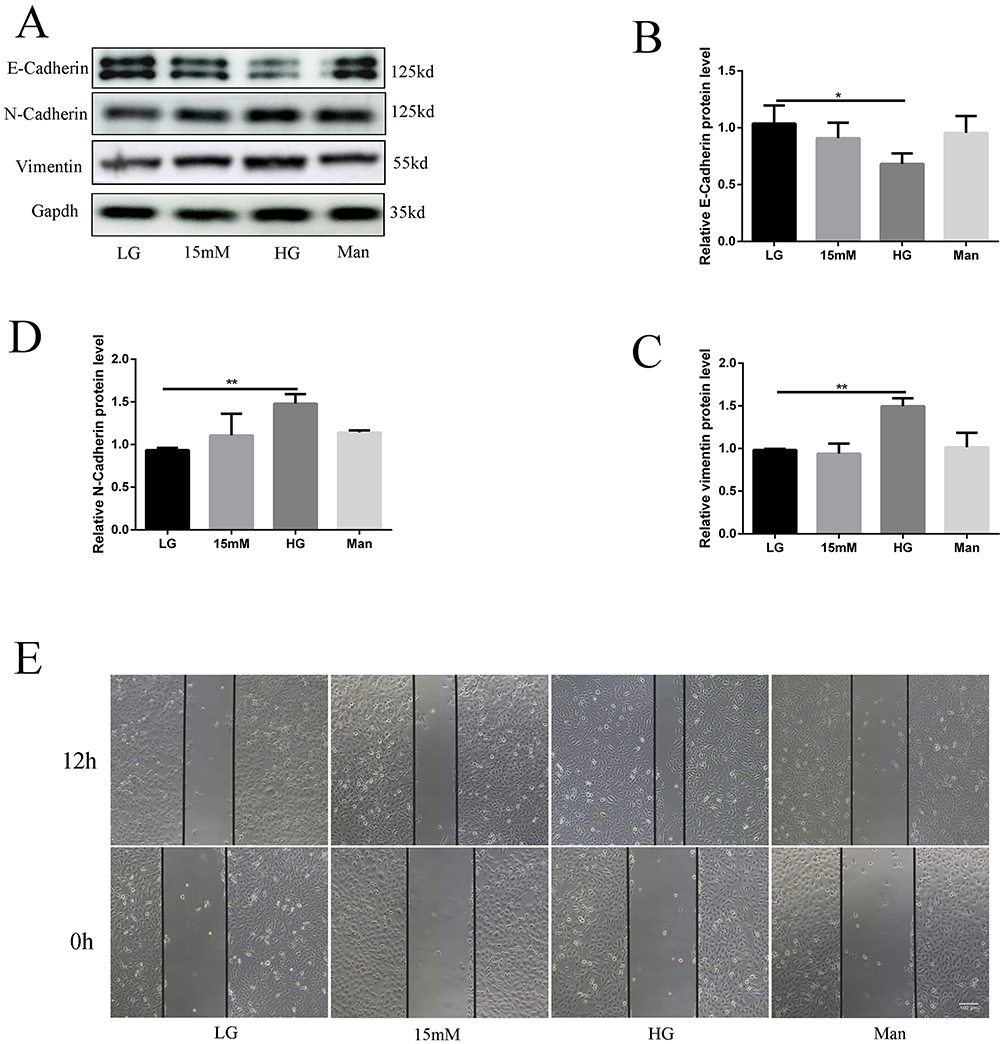 |
Figure 1 High concentrations of glucose could facilitate the EMT of human podocytes. Podocytes were incubated with 11,15,30 mm glucose and 30 mm mannitol for 2 days. The expression of E-cadherin, N-cadherin, and vimentin in podocytes,(A) representative blots. (B–D) representative of the result of densitometric analysis, compared with the expression of EMT-related protein in low glucose (5.5mmol/L) cultured cells, the high glucose group has obvious differences in vimentin, N-cadherin, and E-cadherin.(Data are the mean ± S.D; *P < 0.05 vs LG, **P < 0.01 vs LG, n = 3). The result of cytoskeleton staining(1), cell scratch experiments(1.E). |
Finerenone Could Ameliorate EMT Stimulated by High Glucose in Podocytes
To decide which concentration of finerenone in cell medium is most obvious for its function, we did a cell viability experiment which showed the viability of high glucose (30mmol/L) stimulated cells in the treatment by 2umol/L finerenone was most similar to the viability of cells cultured with low glucose culture (5.5mmol/L) (Figure 2A). Then we used 1 and 2umol/L finerenone in cell medium as treatment to high glucose-stimulated cells, the result of Western blot shows that finerenone could restore the alterations in EMT related proteins stimulated by high glucose (Figure 2B). In 2ummol finerenone treatment group, unlike high glucose group, the expression of vimentin, N-cadherin and E-cadherin all have no significant difference with low glucose group (5.5 mmol/L), which means finerenone could prevent the occurrence of EMT (Figure 2C–E). We also did cell scratch experiments and trans-well experiments which found the migration capacity of the high glucose (30mmol/L) group was improved, and the finerenone alleviated this phenomenon (Figure 3B and C). In cytoskeleton staining, we used phalloidin as dye, the staining results showed that compared with the low glucose group, cytoskeleton arrangement was disordered in the 30mmol/L glucose group while this change could be ameliorated by treatment with 2umol/L finerenone (Figure 3A).
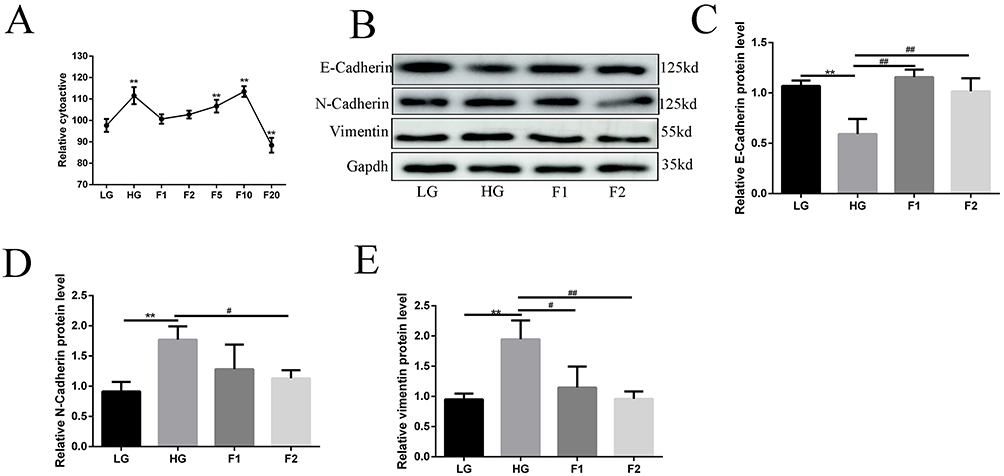 |
Figure 2 Finerenone could ameliorate EMT stimulated by high glucose in podocytes. The result of the cell viability experiment (A), cell viability was measured by CCK-8 assay and expressed as percent cell survival relative to low glucose (5.5mmol/L) controls. (*P < 0.05 vs LG, **P < 0.01 vs LG, n = 3). Podocytes were treated by 1 and 2umol/L finerenone after being stimulated by 30mmol/L glucose. The expression of E-cadherin, N-cadherin, and vimentin in podocytes,(B) representative blots. (C–E) representative of the result of densitometric analysis, compared with the expression of EMT-related protein in high glucose (30mmol/L) cultured cell, 2umol/L finerenone treated group has obvious differences in vimentin, N-cadherin and E-cadherin.(Data are the mean ± S.D; **P < 0.01 vs LG, # P < 0.05 vs HG, ##P < 0.01 vs HG, n = 3). |
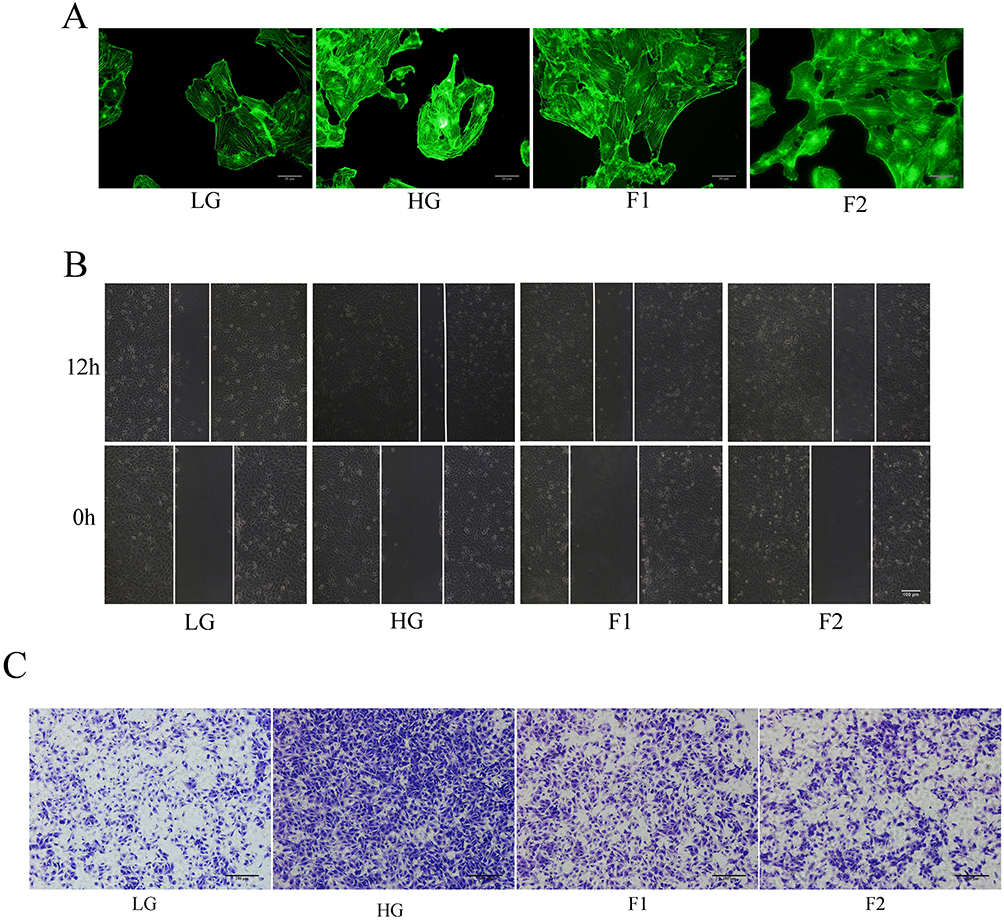 |
Figure 3 Finerenone could ameliorate cytoskeleton and migration capacity of podocytes stimulated by high glucose in podocytes. The result of cytoskeleton staining (A) cell scratch experiments (B) and transwell experiments (C). |
Finerenone Could Upregulate the Expression of KLF5 in High-Glucose Stimulated Cells
In previous studies of our group, KLF5 played an important protective role in diabetic lipotoxicity, so we questioned whether KLF5 could protect podocytes in a high-glucose environment. We did an immunofluorescence experiment and found that finerenone was able to enhance the expression of KLF5 which was downregulated in the high glucose (30mmol/L) group (Figure 4C). The result of the Western blot also proved that finerenone could upregulate the expression of KLF5 in high-glucose stimulated cells (Figure 4A and B).
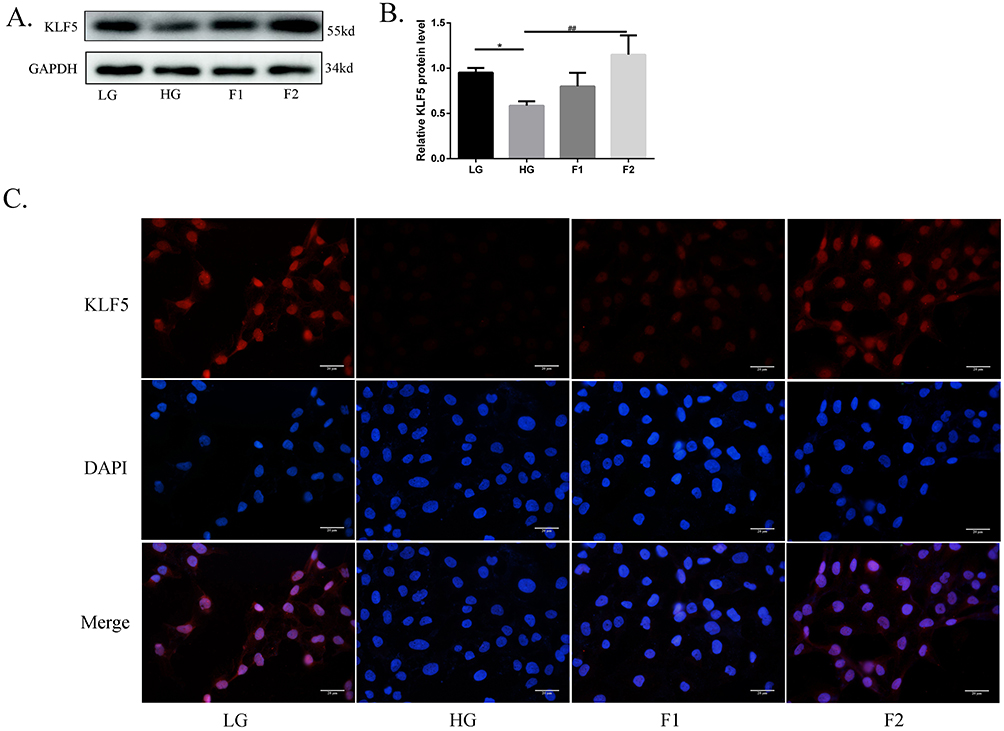 |
Figure 4 Finerenone could upregulate the expression of KLF5 in high-glucose-stimulated cells. Figures 4 and 2 are WB analyses of the same batch of proteins extracted from the same batch of podocytes. The expression of KLF5 in podocytes, (A) representative blots. (B) representative of the result of densitometric analysis, compared with the expression of KLF5 in high glucose (30mmol/L) cultured cells, the 2umol/L finerenone treated group has an obvious difference. (Data are the mean ± S.D; *P < 0.05 vs LG, ##P < 0.01 vs HG, n = 3).The result of the immunofluorescence experiment (C). |
KLF5 Plays a Key Role in the Protective Effect of the Finerenone
To understand whether KLF5 plays an important role in the protection function of finerenone, we decided to silence the expression of KLF5 and then verify the protection function of finerenone. We started to down-regulate the expression of KLF5 by using siRNA. To choose the most effective siRNA, we used three different siRNA to test. After that, we infected the high glucose (30mmol/L) plus 2 umol/L finerenone groups with siRNA and blank vehicle as negative control. By Western blot, we found that NO.1 siRNA has the best-downregulated effect on the expression of KLF5 (Figure 5A and B). Then we stimulated KLF5 silenced podocytes and blank siRNA control podocytes with high glucose, after that we treated them with finerenone (2uM) and negative control. The result of Western blot showed that the protective function of finerenone on EMT was disappear in the KLF5 silenced group (Figure 6A–E), Immunofluorescence experiments which are stained on vimentin and E-cadherin could also prove this conclusion (Figure 7A and B). Cell scratching experiments and trans-well experiments found that finerenone has no ability to restore the migration stimulated by high glucose in KLF5 silenced cells (Figure 6F and G). The cytoskeleton staining results showed that finerenone lost its protective function to cytoskeleton arrangement in a KLF5 silent situation (Figure 6H). These experiments suggested that finerenone probably exerts their protective effect through KLF5.
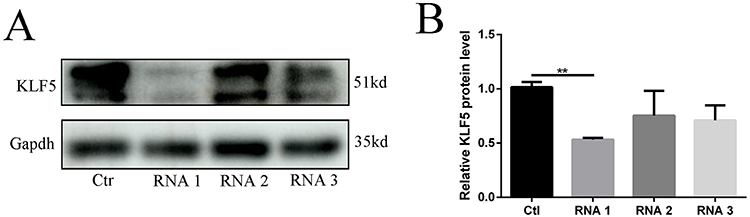 |
Figure 5 Select the most effective KLF5 siRNA Podocytes are transfected by 3 kinds of siRNA and blank vehicle (as a control group). The expression of KLF5 in podocytes,(A) representative blots. (B) representative of the result of densitometric analysis, compared with the expression of KLF5 in the control group, the siRNA1 transfected group has an obvious difference. (Data are the mean ± S.D; **P < 0.01 vs control, n = 3). |
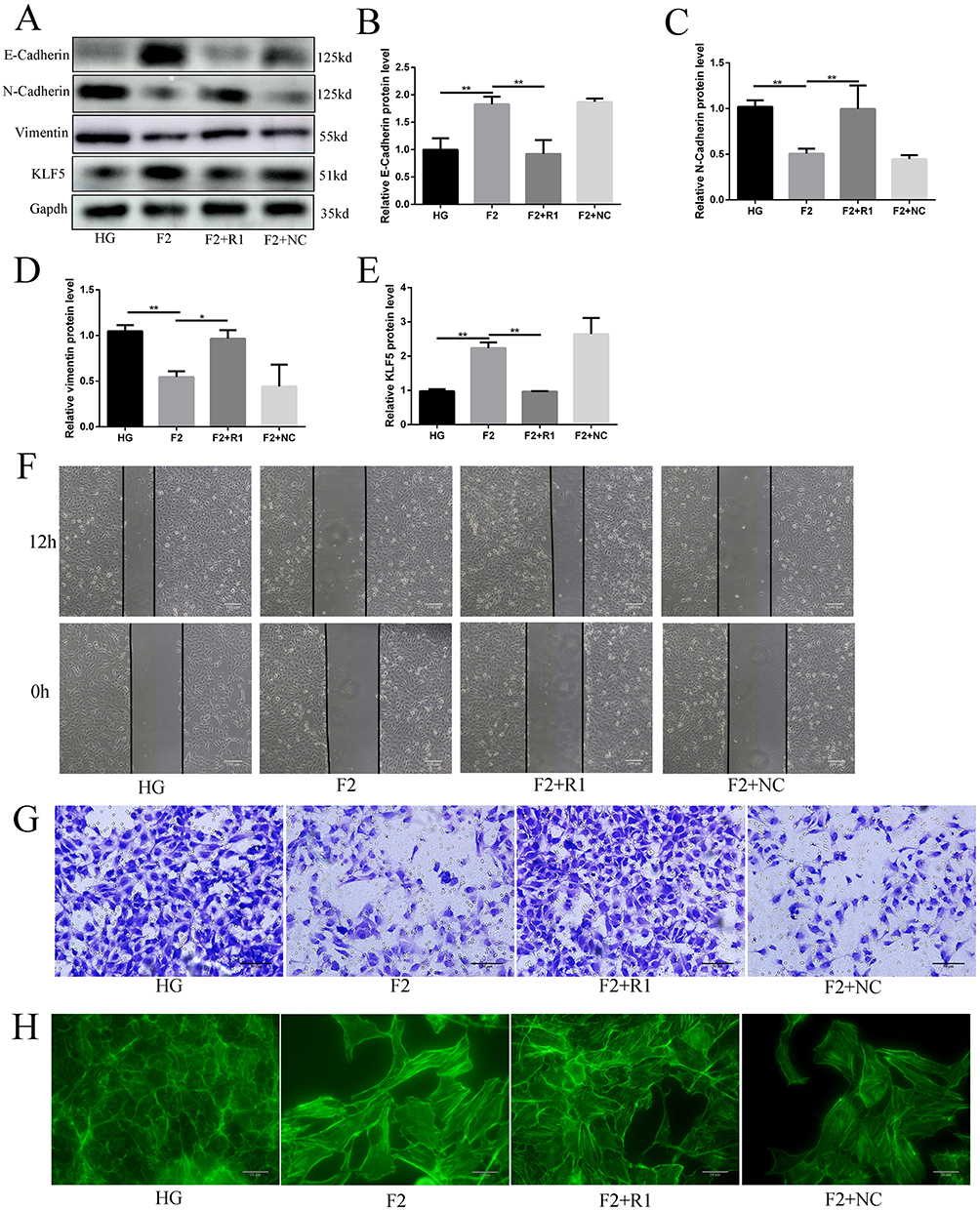 |
Figure 6 KLF5 plays a key role in the protective effect of the finerenone. Podocytes were stimulated with high glucose, and treated with nothing (HG group) or 2umol/L finerenone (F2 group). Podocytes are transfected by siRNA1 (F2+R1 group) and blank vehicle (F2+NC group), then stimulated with high glucose and treated with 2umol/L finerenone. The expression of E-cadherin, N-cadherin, vimentin, and KLF5 in podocytes, (A) representative blots. (B–E) representative of the result of densitometric analysis, compared with the expression of E-cadherin, N-cadherin, vimentin, and KLF5 in the F2 group, HG group, and F2+R1 group have significant differences. (Data are the mean ± S.D; *P < 0.05 vs control, **P < 0.01 vs control, n = 3). The result of cell scratch experiments (F), transwell experiments (G) and cytoskeleton staining (6.H). |
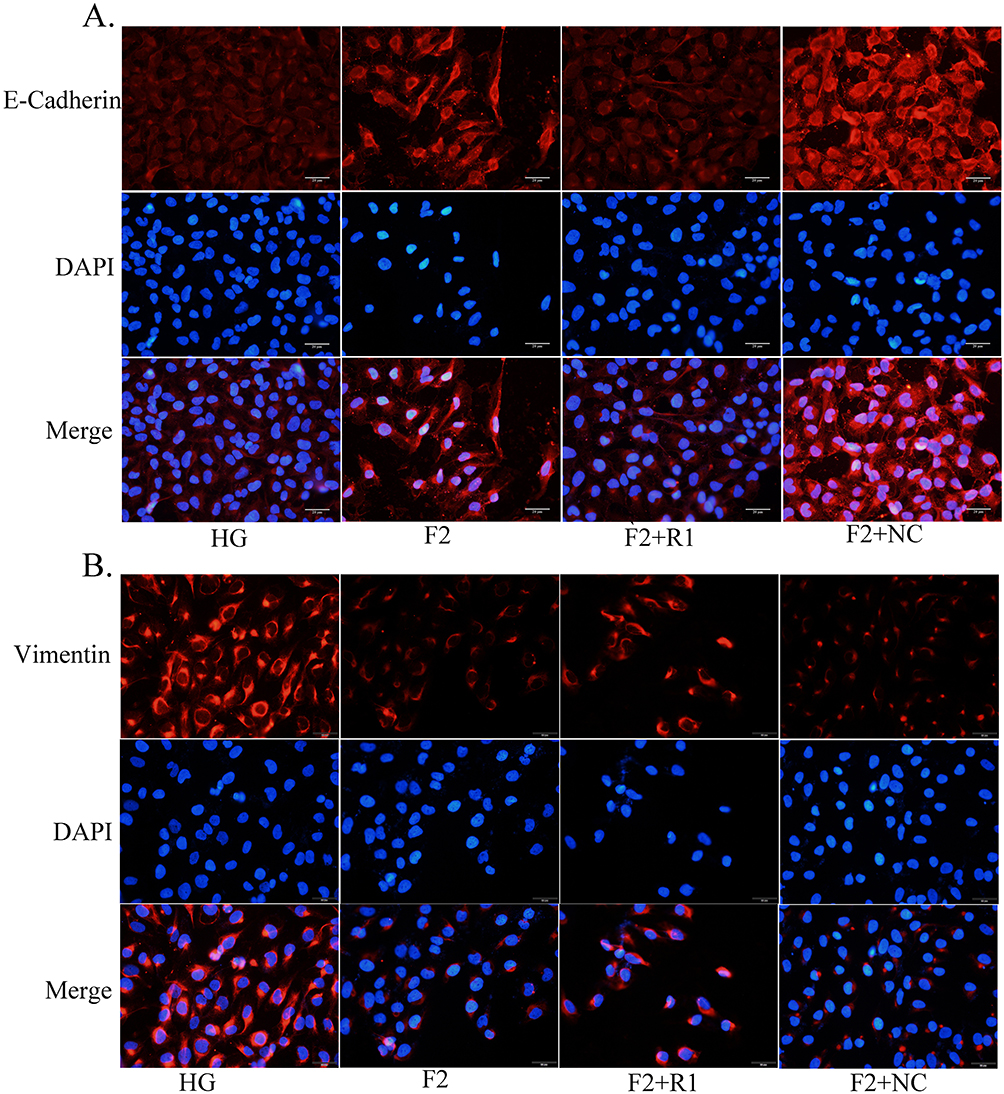 |
Figure 7 KLF5 plays a key role in the protective effect of EMT of the finerenone. The result of the immunofluorescence experiment about E-Cadherin and vimentin (A and B). |
Finerenone Could Ameliorate Kidney Damage in Diabetic Rats
With the progress of gavage, we could find that compared with the control group, all the rats given STZ intraperitoneal injection showed slower weight gain and increased blood glucose (Figure 8A and B). After 8 weeks of gavage, we collected the blood samples of the rats for analysis, and the results showed as follows: Serum creatinine in rats decreased significantly after treatment STZ injection group with finerenone, and both doses of finerenone showed protective effects (Figure 8C). For BUN and potassium, finerenone treatment group has no significant difference with the DN group (Figure 8D and E). In addition, we also found that finerenone had a cholesterol-lowering effect (Figure 8F), which was consistent with our previous findings on the lipid-regulating effect of KLF5.
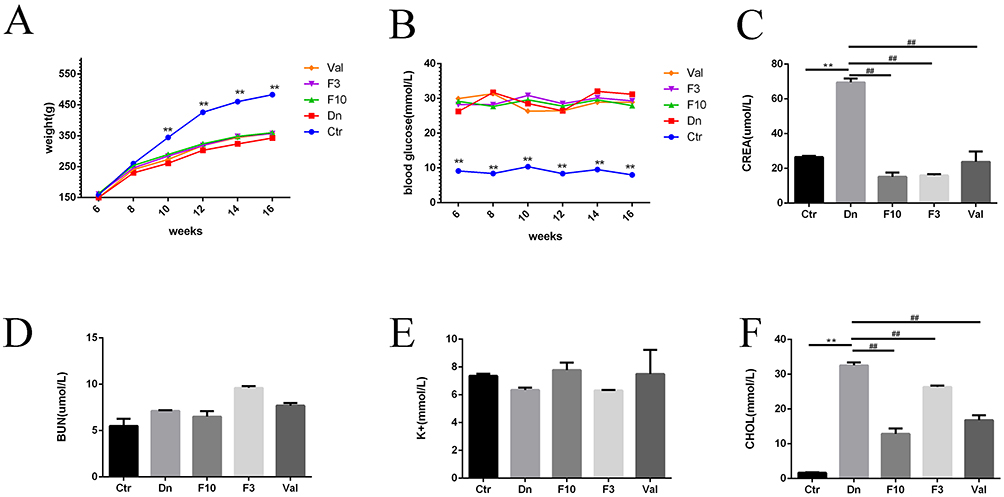 |
Figure 8 Finerenone could ameliorate kidney damage in diabetic rats. The changes in weight and blood glucose of each group of rats (A and B). Analyzing the result of blood samples of the rats: (C–F) representative of the result of serum sample analysis in serum creatinine, BUN, potassium, and cholesterol. (Data are the mean ± S.D; **P < 0.01 vs control group, ##P < 0.01 vs diabetic nephropathy group, n = 3). |
Finerenone Could Ameliorate the Pathological Changes of Glomeruli and Reduce Fibrosis in Diabetic Rats
After PAS, HE, and Masson staining of rat renal tissue sections, it was found that in diabetic rats, glomerulus significantly produced glomerular capillary basement membrane thickening and capillary interstitial (mesangial region) dilatation in response to PAS and HE staining, and these changes were improved by Finerenone treatment. In terms of the degree of fibrosis, Masson staining was significantly deepened in the glomeruli of diabetic rats, suggesting glomerular fibrosis, which was also improved by finerenone (Figure 9F).
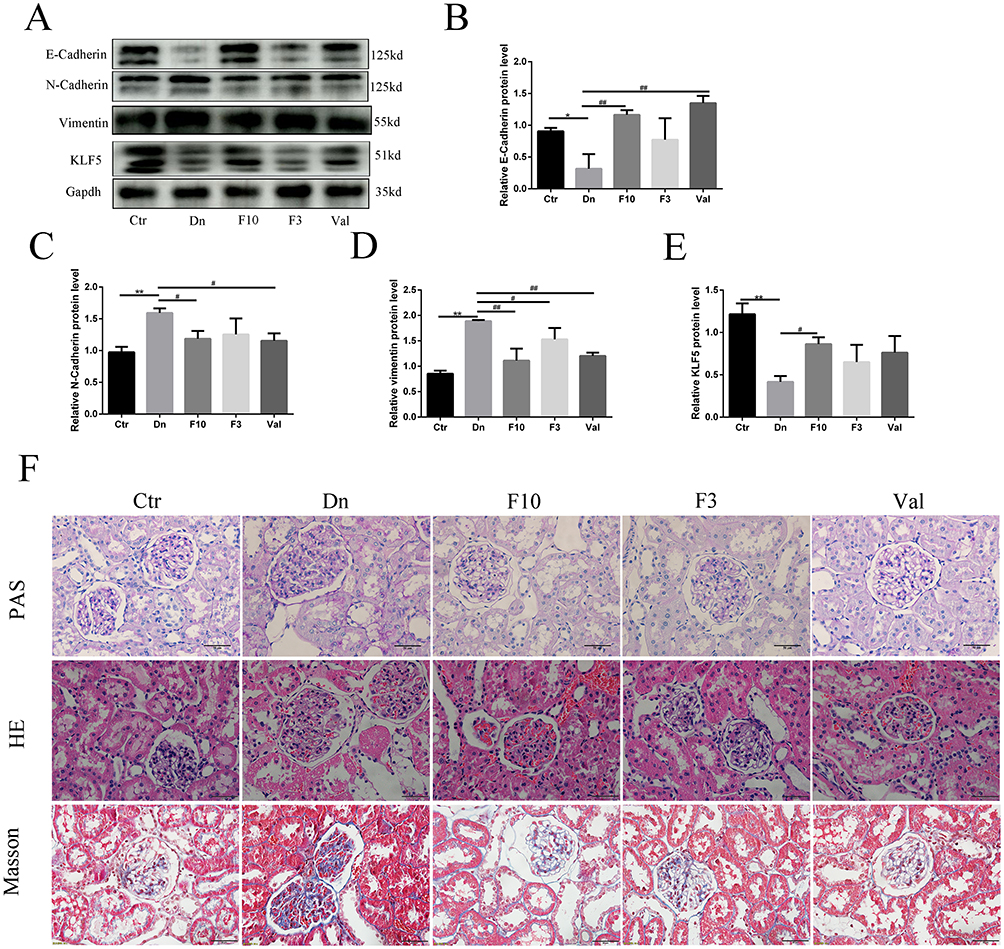 |
Figure 9 Finerenone could ameliorate the pathological changes of glomeruli, reduce fibrosis, ameliorate EMT, and upregulate the expression of KLF5 in diabetic rats. The expression of E-cadherin, N-cadherin, vimentin, and KLF5 in the kidney tissue of rats, (A) representative blots. (B–E) representative the result of densitometric analysis, compared with the expression of E-cadherin, N-cadherin, vimentin, and KLF5 in the control group, diabetic nephropathy group has obvious difference; compared with diabetic nephropathy group, some treated group have a significant difference in above protein. (Data are the mean ± S.D; *P < 0.05 vs control group, **P < 0.01 vs control group,# P < 0.05 vs diabetic nephropathy group, ##P < 0.01 vs diabetic nephropathy group, n = 3). (F) representative PAS, HE, and Masson staining. |
Finerenone Could Ameliorate EMT and Upregulate the Expression of KLF5 in podocytes
Next, we performed a Western blot analysis on the protein extracted from the kidney tissue of the rats: compared with the normal group, EMT-related proteins were found to be changed in the diabetic rats, the expression of E-Cadherin was decreased (Figure 9B) and the expression of N-Cadherin and Vimentin were increased (Figure 9C and D). Same with the cell experiment, the expression of KLF5 was also significantly decreased in the diabetic rats (Figure 9E). These trends of expression of proteins were restored by finerenone (Figure 9A). In finerenone 10mg/kgd group, the expression of these proteins had no statistical difference with the normal group, while in finerenone 3mg/kgd group, the expressions of some EMT-related proteins (N-Cadherin and Vimentin) were improved, and the expression level of KLF5 was improved compared with that of diabetic nephropathy group, however, there was no significant difference with the diabetic nephropathy group. We also found that the expression of vimentin and KLF5 were increasing in glomerular supported by immunofluorescence of kidney tissue sections (Figure 10B and C).
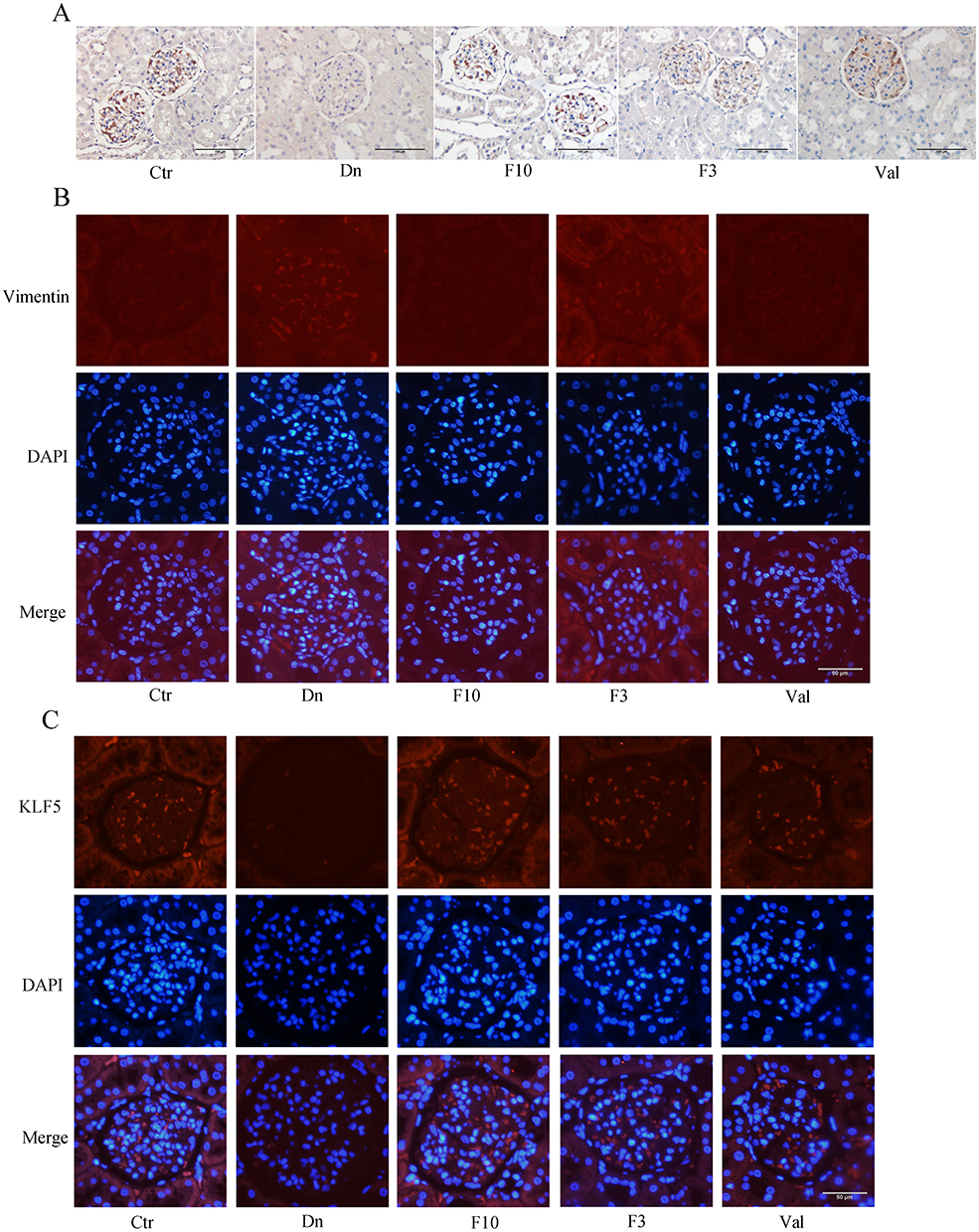 |
Figure 10 Finerenone could protect podocytes in diabetic rats (A) represent the result of an immunohistochemistry experiment for nephrin. Red arrows represent high expression region of KLF5. (B and C) represent the result of the immunofluorescence experiment for vimentin and KLF5. Blue arrows represent high expression region of vimentin, green arrows represent high expression region of KLF5. |
Finerenone Could Protect Podocytes in Diabetic Rats
We selected nephrin as the target protein to conduct immunofluorescence and immunohistochemistry in rat glomerular sections. The results showed that the expression levels of nephrin in the diabetic group were decreased compared with the control group, which indicated the damage of podocin cells in the kidneys of diabetic rats. However, after treatment with finerenone, the expression levels of nephrin were significantly recovered, suggesting the protective effect of finerenone on renal podocytes (Figure 10A).
Discussion
In this study, we found the protective effect of finerenone on diabetic nephropathy and podocytes stimulated by high glucose. The results showed that finerenone alleviated EMT by KLF5, thereby alleviating damage to DN and podocytes. In cell experiment, finerenone inhibited EMT, apoptosis, and cytoskeletal rearrangement by KLF5. In animal experiment, we found that Finerenone reduced EMT, apoptosis, and glomerular damage. Therefore, Finerenone may be an effective drug to treat DN or even reverse kidney damage in DN, and it would provide new ideas for future research.
Finerenone is a novel, high-selective mineralocorticoid receptor antagonist.16 Many articles have already proved that finerenone has a very good effect on kidney disease, especially on kidney disease caused by diabetes. In CKD and type 2 diabetes patients, finerenone treatment could reduce the progress of CKD and the occurrence of cardiovascular events.17 It has been proved that finerenone could improve renal fibrosis in diabetes nephropathy.18 These above articles suggested that finerenone may have a protective effect on patients with diabetes nephropathy.
Diabetes nephropathy (DN) is the main cause of end-stage renal disease worldwide. The main starting mechanism of DN is hyperglycemia-induced vascular dysfunction.19 Its progression is due to different pathological mechanisms, including oxidative stress, inflammatory cell infiltration, inflammation, and fibrosis. In addition, studies have proved that EMT plays a key role in the development of diabetes nephropathy.20
Epithelial-mesenchymal transformation (EMT) is a regulated process in which epithelial cells lose their marker epithelial characteristics and acquire mesenchymal characteristics.21 For podocytes, the expression of nephrin, podocin, E-cadherin, and ZO-1 is down-regulated, the slit diaphragm (SD) will change, and the actin cytoskeleton will rearrange. Diabetes, especially hyperglycemia, has been proven through a variety of molecular mechanisms, such as the TGF- β/Smad classic approach, Wnt /β- Catenin signal pathway, integrin /integrin-linked kinase (ILK) signal pathway, MAPKs signal pathway, Jagged /Match signal pathway, and NF - κB signal pathway to stimulate podocyte EMT.22,23 In our study, high glucose environment will stimulate podocytes to generate EMT, which is one of the reasons for podocyte lesions in diabetes patients. This process could be alleviated by novel, high-selective mineralocorticoid receptor antagonist finerenone.
Abnormal expression of the Krϋppel-like factor (KLF) gene appears in many disease-related pathologies. KLF2 is essential for maintaining endothelial barrier integrity, preventing crevice formation, and preventing glomerular endothelial and podocyte damage. In the AKI environment, KLF4 has a renal protective effect and is a key regulator of proteinuria in mice and humans. Expression of KLF6 in podocytes protects mitochondrial function and prevents podocyte apoptosis, while KLF5 can prevent podocyte apoptosis by blocking the ERK/p38 MAPK pathway.24 Those molecules might be a new target for diabetic nephropathy treatment.
In our previous study, we already proved that the KLF5 could alleviate podocyte injury by regulating cholesterol metabolism. In our current experiment, the cholesterol level of rats treated with finerenone also decreased, and the level of EMT-related protein in the kidney of rats also recovered, which was confirmed in Western blot and immunohistochemistry experiments. However, in the current scientific research field, the effect of KLF5 is not completely determined: KLF5 can regulate apoptosis, proliferation, inflammation, oxidative stress, and fibrosis related to obesity or diabetes-induced nephropathy and other kidney diseases. Different cell types or animal models have different conclusions.25 Baotong Z et al found that KLF5 had an inhibitory effect on EMT.26 Many reports have confirmed that KLF5 has a significant effect on EMT, this may be related to NF-κB,27 HIF-1,28 TGF-β29 and other pathways. A number of studies have reported the interaction between KLF5 and TGF-β in the occurrence of EMT29,30 of tumor. So far, the exact mechanism of KLF5 acting on various kidney diseases is still limited. In our experiment, we proved that KLF5 has a protective effect on the kidney in diabetes: the EMT levels of rats with KLF5 level restored are significantly reduced. In the next step, our research team needs to build more different animal and disease models to further evaluate the role of KLF5 in kidney disease. In this study, we proved for the first time that finerenone exerts its protective effect on DN and high glucose-stimulated podocytes by affecting Podocytes EMT through enhancing the expression of KLF5. However, merely applying rats may not fully simulate patients with diabetic nephropathy; in the future, we consider using large animals (like primate) for further research.
In conclusion, in our current study, we found that finerenone inhibits renal fibrosis and podocyte damage induced by high glucose. Mechanistically, we found that finerenone could restore KLF5 expression and reduce high glucose-induced EMT by KLF5.
Data Sharing Statement
The data generated by the study are available from the corresponding author on reasonable request.
Ethics Statement
The study involving animals met the Animals in Research Reporting In Vivo Experiments (ARRIVE) guidelines and followed the guidelines of the National Institutes of Health guide. All experiments were approved by the hospital ethics committees of Provincial Hospital Affiliated to Shandong First Medical University.
Acknowledgments
The authors wish to thank the staff at the center laboratory of Shandong Provincial Hospital.
Funding
This work was supported by the Natural Science Foundation of Shandong Province of China to Ying Xu (ZR2022MH041), the National Natural Science Foundation of China to Ying Xu (81500553), the National Natural Science Foundation of China to Qinglian Wang (82100791), the Natural Science Foundation of Shandong Province of China to Qinglian Wang (ZR202102181004).
Disclosure
All authors declared that they have no conflicts of interest.
References
1. Guariguata L, Whiting DR, Hambleton I, et al. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabet Res Clin Pract. 2014;103:137–149. doi:10.1016/j.diabres.2013.11.002
2. Cole JB, Florez JC. Genetics of diabetes mellitus and diabetes complications. Nat Rev Nephrol. 2020;16:377–390. doi:10.1038/s41581-020-0278-5
3. Amer Diabet A. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2014;37:S81–S90. doi:10.2337/dc14-S081
4. Umanath K, Lewis JB. Update on Diabetic Nephropathy: core Curriculum 2018. Am J Kidney Dis. 2018;71:884–895. doi:10.1053/j.ajkd.2017.10.026
5. Anders HJ, Huber TB, Isermann B, Schiffer M. CKD in diabetes: diabetic kidney disease versus nondiabetic kidney disease. Nat Rev Nephrol. 2018;14:361–377. doi:10.1038/s41581-018-0001-y
6. Cai RY, Jiang JJ. LncRNA ANRIL silencing alleviates high glucose-induced inflammation, oxidative stress, and apoptosis via upregulation of MME in podocytes. Inflammation. 2020;43:2147–2155. doi:10.1007/s10753-020-01282-1
7. Alicic RZ, Rooney MT, Tuttle KR. Diabetic kidney disease challenges, progress, and possibilities. Clin J Am Soc Nephrol. 2017;12:2032–2045. doi:10.2215/cjn.11491116
8. Bhatia D, Choi ME. Autophagy in Health and Disease. Progress mol Biol Transl Sci. 2020;172:107–133.
9. Humphreys BD. Mechanisms of renal fibrosis. Ann Rev Physiol. 2018;80:309–326. doi:10.1146/annurev-physiol-022516-034227
10. Song AN, Zhang C, Meng XF. Mechanism and application of metformin in kidney diseases: an update. Biomed Pharmacother. 2021;138:12. doi:10.1016/j.biopha.2021.111454
11. Tang GY, Li S, Zhang C, et al. Clinical efficacies, underlying mechanisms and molecular targets of Chinese medicines for diabetic nephropathy treatment and management. Acta Pharm Sin B. 2021;11:2749–2767. doi:10.1016/j.apsb.2020.12.020
12. Lovisa S, LeBleu VS, Tampe B, et al. Epithelial-to-mesenchymal transition induces cell cycle arrest and parenchymal damage in renal fibrosis. Nat Med. 2015;21:998. doi:10.1038/nm.3902
13. Li YJ, Hu Q, Li C, et al. PTEN-induced partial epithelial-mesenchymal transition drives diabetic kidney disease. J Clin Invest. 2019;129:1129–1151. doi:10.1172/jci121987
14. Shibata S, Nagase M, Yoshida S, Kawachi H, Fujita T. Podocyte as the target for aldosterone - roles of oxidative stress and Sgk1. Hypertension. 2007;49:355–364. doi:10.1161/01.HYP.0000255636.11931.a2
15. Rico-Mesa JS, White A, Ahmadian-Tehrani A, Anderson AS. Mineralocorticoid receptor antagonists: a comprehensive review of finerenone. Curr Cardiol Rep. 2020;22:11. doi:10.1007/s11886-020-01399-7
16. Frampton JE. Finerenone: first Approval. Drugs. 2021;81:1787–1794. doi:10.1007/s40265-021-01599-7
17. Bakris GL, Agarwal R, Anker SD, et al. Effect of finerenone on chronic kidney disease outcomes in type 2 diabetes. N Engl J Med. 2020;383:2219–2229. doi:10.1056/NEJMoa2025845
18. Agarwal R, Kolkhof P, Bakris G, et al. Steroidal and non-steroidal mineralocorticoid receptor antagonists in cardiorenal medicine. Eur Heart J. 2021;42:12. doi:10.1093/eurheartj/ehaa736
19. Calle P, Hotter G. Macrophage phenotype and fibrosis in diabetic nephropathy. Int J mol Sci. 2020;21(14):2806. doi:10.3390/ijms21082806
20. Loeffler I, Wolf G. Epithelial-to-Mesenchymal Transition in Diabetic Nephropathy: fact or Fiction? Cells. 2015;4:631–652. doi:10.3390/cells4040631
21. Dai HR, Liu QQ, Liu BL. Research progress on mechanism of podocyte depletion in diabetic nephropathy. J Diabetes Res. 2017;10. doi:10.1155/2017/2615286
22. Ying QD, Wu GZ. Molecular mechanisms involved in podocyte EMT and concomitant diabetic kidney diseases: an update. Ren Fail. 2017;39:474–483. doi:10.1080/0886022x.2017.1313164
23. Lee HS. Mechanisms and consequences of TGF- overexpression by podocytes in progressive podocyte disease. Cell Tissue Res. 2012;347:129–140. doi:10.1007/s00441-011-1169-7
24. Rane MJ, Zhao YG, Cai L. Kr nu ppel-like factors (KLFs) in renal physiology and disease. EBioMedicine. 2019;40:743–750. doi:10.1016/j.ebiom.2019.01.021
25. Li J, Liu L, Zhou W-Q, et al. Roles of Kruppel-like factor 5 in kidney disease. J Cell Mol Med. 2021;25:2342–2355. doi:10.1111/jcmm.16332
26. Zhang BT, Zhang Z, Xia S, et al. KLF5 activates microRNA 200 transcription to maintain epithelial characteristics and prevent induced epithelial-mesenchymal transition in epithelial cells. mol Cell Biol. 2013;33:4919–4935. doi:10.1128/mcb.00787-13
27. Liu FF, Dong L, Yang X, et al. KLF5 silence attenuates proliferation and epithelial-mesenchymal transition induction in Hep-2 cells through NF-κB signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23:3867–3875. doi:10.26355/eurrev_201905_17814
28. Zhou Y, Xu QH, Shang JJ, Lu LH, Chen GY. Crocin inhibits the migration, invasion, and epithelial-mesenchymal transition of gastric cancer cells via miR-320/KLF5/HIF-1α signaling. J Cell Physiol. 2019;234:17876–17885. doi:10.1002/jcp.28418
29. David CJ, Huang Y-H, Chen M, et al. TGF-β tumor suppression through a lethal EMT. Cell. 2016;164:1015–1030. doi:10.1016/j.cell.2016.01.009
30. Zhang BT, Li Y, Wu Q, et al. Acetylation of KLF5 maintains EMT and tumorigenicity to cause chemoresistant bone metastasis in prostate cancer. Nat Commun. 2021;12:21. doi:10.1038/s41467-021-21976-w
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.