Back to Journals » OncoTargets and Therapy » Volume 12
Fibronectin 1 promotes melanoma proliferation and metastasis by inhibiting apoptosis and regulating EMT
Authors Li B, Shen W, Peng H, Li Y, Chen F, Zheng L, Xu J, Jia L
Received 23 November 2018
Accepted for publication 9 March 2019
Published 1 May 2019 Volume 2019:12 Pages 3207—3221
DOI https://doi.org/10.2147/OTT.S195703
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Gaetano Romano
Bifei Li,1* Weiyu Shen,1* Huayi Peng,2* Yumei Li,1 Fan Chen,1 Liping Zheng,3 Jianhua Xu,2 Lee Jia1,4
1Cancer Metastasis Alert and Prevention Center, College of Chemistry; Fujian Provincial Key Laboratory of Cancer Metastasis Chemoprevention and Chemotherapy, Fuzhou University, Fuzhou, Fujian 350116, People’s Republic of China; 2College of Pharmacy, Fujian Medical University, Fuzhou 350116, People’s Republic of China; 3Pharmacy Department, Fujian Province Children Hospital, Fuzhou, Fujian 350108, People’s Republic of China; 4Institute of Oceanography, Minjiang University, Fuzhou, Fujian 350108, People’s Republic of China
*These authors contributed equally to this work
Background and aims: The complex process of cancer metastasis remains the least understood. Tumor cells alter their protein expression profile to survive from the tumor metastasis. Fibronectin 1 (FN1 gene coding protein) is a member of the glycoprotein family that has been shown to play an important role in cancer metastasis. However, its effects on melanoma metastasis are still unclear.
Methods: We detected the FN1 expression between metastatic cells and primary cells by using Western blot and RT-qPCR assays. And, we analyzed the expressed feature of FN1 in different tissues and examined the clinical relevance of upregulated FN1 in melanoma progression by bioinformatic analysis. Furthermore, we downregulated the expression of FN1 by small interfering RNA technique to reveal the effect of FN1 on melanoma phenotype and expression of related genes. Finally, we used bioinformatics to reveal the possible mechanism of FN1 regulating melanoma progression.
Results: We reported that the expression of FN1 was changed during melanoma metastasis. In this study, we established two metastatic cell lines of melanoma through mouse model, and found that metastatic cells exhibited stronger mesenchyme phenotype and possessed higher FN1 expression level compared to primary cells. Besides, we examined the clinical relevance of upregulated FN1 in tumor progression. Small interfering RNA (siRNA)-mediated downregulation of FN1 suppressed the migration, invasion, adhesion, proliferation capabilities and induced apoptosis of melanoma cells. We detected a diminished EMT-related gene signature including increased expression of E-cadherin and decreased expression of N-cadherin and Vimentin. Downregulation of FN1 also increased Bax/Bcl-2 ratio which might result in apoptosis of melanoma cells. Bioinformatics analysis revealed that FN1 most likely involved in focal adhesion and PI3K-Akt signaling pathway to regulate EMT process and apoptosis.
Conclusions: Taken together, these findings demonstrated a role of FN1 in promoting melanoma metastasis by inhibiting apoptosis and regulating EMT.
Keywords: cancer metastasis, epithelial-mesenchymal transition, survival protein, migration, invasion
Introduction
Recent progress in cancer diagnosis and treatment has contributed to better treatment outcomes and survival rate. However, the complex process of cancer metastasis remains the least understood. Cancer metastasis is still the leading cause of death in cancer patients.1 Melanoma is the most dangerous type of skin cancer.2 There were 3.1 million with active disease which resulted in 59,800 deaths.3 Metastatic melanoma continues to be a challenging disease to treat.4 The 10-year survival rate for patients with metastatic melanoma is less than 10%.5,6 Thus, elucidation of the molecular mechanism is critical to alert and prevent melanoma metastasis.
Only specific tumor phenotypes that result from molecular alterations can penetrate the walls of lymphatic or blood vessels, and therefore are able to circulate in the bloodstream to other tissues in the body.7 Tumor cells must alter expression level of some proteins to survive in blood or lymph and arrive at distant sites.8 Epithelial-mesenchymal transition (EMT), an essential step in tumor progression, was reportedly involved in cancer metastasis. EMT is usually associated with the loss of cell polarity and cell–cell adhesion and acquires migratory and invasive properties and variations of morphological by multiple pathways. Cell surface proteins, E-cadherin (biomarker of epithelial cells) or integrin are replaced by mesenchymal markers (N-cadherin, Vimentin) in EMT process.9–11
Fibronectin 1 (FN1) is a member of the glycoprotein family that is widely expressed by multiple cell types.12 FN1 plays a major role in cell adhesion, growth, migration and differentiation, and it is important for processes such as wound healing and embryonic development.13 Degradation or organization of FN1 expression has been associated with cancer progression,14 such as squamous cell carcinoma,15 nasopharyngeal carcinoma,16 ovarian cancer, renal cancer17 and thyroid cancer.18 Recent studies have shown that increased expression of FN1 in tumor cells is negatively correlated to the prognosis of patients.19 Furthermore, researcher suggested that increased FN1 expression may be associated with lung tumor growth/survival and resistance to therapy.20 Our studies showed that FN1 survived from melanoma metastasis and its expression was upregulated in metastatic tumor cells as compared to primary tumor cells.
Despite the wealth of existing data about the role of FN1 in cancer, its clear picture is yet to be elucidated in melanoma metastasis. To reveal the underlying significance of upregulated FN1 in melanoma metastasis, we examined the clinical relevance of FN1 in tumor progression using public databases of cancer patients and demonstrated that downregulated FN1 expression inhibiting proliferation and metastasis of melanoma cells by inducing apoptosis and suppressing EMT. Meanwhile, we revealed that downregulated FN1 expression significantly decreased the expression of Bcl-2 and increased the expression of Bax. We thus propose that FN1 may function as a metastasis promoter and could be a target for alerting and preventing melanoma metastasis.
Materials and methods
Antibodies and reagents
FN1 was purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Rabbit primary antibodies Bax, Bcl-2, CyclinD1, E-cadherin, N-cadherin and enhanced chemiluminescence (ECL) kit were all obtained from Wanlei Biotechnology. Goat anti-rabbit secondary antibodies conjugated with horseradish peroxidase (HRP) and β-actin were purchased from DingguoChangsheng Biotechnology (Beijing, China). Fetal bovine serum (FBS), RPMI medium and Trizol reagent were obtained from Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Propidium iodide (PI), dimethyl sulfoxide (DMSO), Triton X-100, RNAse, 4% paraformaldehyde and a bicinchoninic acid (BCA) protein assay kit were purchased from Sigma-Aldrich (Merck Kgaa, Darmstadt, Germany). Trypsin-EDTA (0.25%), penicillin/s-treptomycin (100×), tetramethylethylenediamine (TEMED), ammonium persulfate (AP), trihydroxymethyl aminomethane (Tris), glycine, Tween-20, sodium dodecyl sulfonate (SDS), apoptosis assay kit, RIPA lysis buffer and bovine serum albumin (BSA) were obtained from Genview Scientific, Inc. (El Monte, FL, USA). A PrimeScript® RT reagent kit and SYBR®Premix Ex TaqTM PCR kit were purchased from Takara Bio, Inc. (Otsu, Japan). Polyvinylidene difluoride (PVDF) membranes, 30% polyacrylamide were purchased from Bio-Rad Laboratories, Inc. (Hercules, CA, USA).
Animals and ethics statement
Female C57BL/6 mouse and female BALB/C nude mouse were purchased from Shanghai SLAC Laboratory. All animal studies were performed in accordance with animal protocol procedures approved by the Institutional Animal Care and Use Committee (IACUC) of Fuzhou University, which are consistent with AAALAS guidelines. All animal experiments were also approved by the IACUC of Fuzhou University and were monitored for abnormal behaviors to minimize animal pain and suffering. Animals were euthanized if excessive deterioration of animal health was noted.
Cell lines and cell culture
The human melanoma cell line A375 and mouse melanoma cell line B16F10 were purchased from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in RPMI (obtained from HyClone) medium containing 10% FBS (obtained from GEMINI) and 1% penicillin/streptomycin. The corresponding pulmonary metastases were obtained by injecting primary melanoma cells into mouse through tail vein. Briefly, 50,000 B16F10 cells and 4,000,000 A375 cells were injected into C57BL/6 mouse and BALB/C nude mouse, respectively. Six C57BL/6 mice and six BALB/C nude mice are injected with parental B16F10 and A375, respectively, and all mice developed the metastatic spread after 50 days. Pulmonary nodules dissected from BALB/C nude mouse and C57BL/6 mouse were named A375M and B16F10M, respectively.
Apoptosis assay and cell cycle analysis
For cell apoptosis assay, B16F10M and A375M cells were plated in 6-well culture dishes and transfected with negative control siRNA or FN1 siRNA for 48 hrs. Cells were washed, trypsinized, harvested and re-suspended in 400 μL binding buffer. Then, 5 μL Annexin V-FITC and 5 μL propidium iodide (PI) were added to the cell suspension. After incubation for 15 mins following the manufacturer’s instructions at room temperature, these cells were performed on BD FACS Aria III flow cytometer. We collected at least 10,000 cells per sample. Data were analyzed by FlowJo software (FLOWJO, Ashland, OR). Cells stained with only Annexin V-FITC or PI were used as compensation controls. For cell cycle assay, B16F10M and A375M cells were plated in 6-well culture dishes and transfected with negative control siRNA or FN1 siRNA for 48 hrs. Then they were collected, washed with PBS and fixed with 75% ethanol at 4°C for 24 hrs. Fixed cells were washed once with PBS (phosphate buffer saline) and incubated for 30 mins with propidium iodide (PI) solution (20 μg/mL) containing RNAase A (0.1 μg/mL) and TritonX-100 (0.1%). Cell cycle was determined by using the FACS Aria III flow cytometer and analyzed by using the Modfit software.
Western blotting analysis
Whole-cell protein lysates were extracted from cells using ice-cold radio immunoprecipitation assay buffer containing 0.01% protease and phosphatase inhibitor and incubated on ice for 30 mins. The protein concentration was determined using the BCA protein assay. The samples were denatured by adding SDS running buffer. The equal amounts of denatured protein samples were separated by 8% or 12% (w/v) sodium dodecyl sulfate (SDS)-polyacrylamide gel, then transferred to polyvinylidene difluoride (PVDF) membranes, followed by blocking with 5% BSA in Tris‑buffered saline with Tween 20 (TBST) and incubating with corresponding primary antibodies overnight. Membranes were then incubated for 2 hrs at room temperature with horseradish peroxidase-linked secondary antibodies, and intensities were measured on ChemiDoc XPS system (Bio-Rad Laboratories, Inc.) using an enhanced chemiluminescence (ECL) kit. The β-actin was used as internal control. The bands were quantified with Image J software.
Quantitative real-time PCR (RT-qPCR) analysis
Total RNA was extracted from melanoma cells using Trizol reagent (Invitrogen) according to the manufacturer’s protocol. The cDNA synthesis was performed with 450 ng RNA per 10 μL by using PrimeScript® RT Reagent Kit (Takara, Japan) according to the manufacturer’s instructions. The real-time PCR was carried out on the CFX96TM Real-Time PCR Detection Systems (Bio-Rad) with cDNA (<100 ng) per 25 μL reaction volume using SYBR® Premix Ex Taq™ PCR Kit (TaKaRa, Japan). PCR primers are listed in Table 1. Each experiment was performed independently at least three times. Relative expression levels were normalized to β-actin and calculated by 2−ΔΔct method.
 | Table 1 Sequences of RT-qPCR primers and siRNAs |
Cell transfection
The B16F10M cells were plated in 6-well dishes and grew to 70–80% confluence, and then siRNA targeting mouse FN1 mRNA and a non-specific scramble siRNA sequence (negative control siRNA) purchased from Sangon Biotech were transiently transfected into B16F10M cells using lipofectamine 3,000 (Invitrogen) according to the manufacturer's instruction. Similarly, siRNA targeting human FN1 mRNA and a non-specific scramble siRNA sequence (negative control siRNA) were transfected into A375M cells. The sequences of RNAi primer are listed in Table 1. The amount of siRNA is 150 pmol per well. Knockdown of FN1 was determined by western blotting and RT-qPCR.
Cell proliferation and colony formation assay
Cell transfection experiment was conducted as described previously. Cell proliferation was measured using MTT assay. Transfected metastatic cells and control cells were plated in five replicates of wells in a 96-well plate. Cells were incubated with 200 μL MTT solution for 4 hrs. Then, the MTT solution was replaced by 100 μL dimethyl sulfoxide (DMSO), and the plate was shaken at 100 rpm for 30 mins. The result was quantitated using a microplate reader (TECAN, M200PRO, Switzerland) at 490 nm. In colony formation assay, the transfected metastatic cells and control cells were plated into 6-well dishes at a density of 1,000 cells per well and cultured for 2 weeks. Colonies were fixed with cold 4% paraformaldehyde for 30 mins and stained with 0.1% crystal violet for 30 mins. Each experiment was repeated in triplicate.
Invasion and migration assay
For the transwell invasion and migration assay, we prepared transwell culture chambers (24-well, 8 μm pore size, Costar, Corning Incorporated, USA) coated with/without 60 μL Matrigel (Corning, TewksBury, MA). After incubation at 37°C for 2 hrs, the transfected or control cells (1×105 cells) in 200 μL RPMI (0.1% BSA)were added into the upper compartment of the chambers, and 800 μL medium containing 20% FBS was used as the attractant and placed in the bottom chambers. After incubation for 48 hrs at 37°C and 5% CO2, non-invaded cells on the upper chamber were scraped off with a cotton swab. The invaded cells on the lower membrane were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet (genview) for 30 mins. The invading cells were counted and photographed (five random fields) by using optical microscope (Zeiss, Germany).
Wound-healing migration assay
Cells were plated into 6-well plates and grew to confluence. The monolayer was wounded using the tip of a sterile 200 μL pipette. Cell debris was removed by washing twice with PBS. These cells were then cultured in 1% serum medium. Photomicrographs were taken at 0 hrs and 24 hrs of the same wounded areas using the inverted microscope (Zeiss, Germany). The percentage (%) change in migration was used to evaluate the cell migration capability.
Adhesion assay
Briefly, we prepared 12-well plates coated with 1 mL 1% gelatin per well and put the plates in cell culture incubator overnight. Then the unsolidified gelatin was removed and each well was blocked with 5% BSA at room temperature for 30 min. Cells stained with rhodamine-123 for 30 min were added into plates. After incubation at 37°C for 30 mins, the non-adhered cells were removed from the plate by washing twice with PBS, and then the adhered cells were counted and photographed (five random fields) under a fluorescence microscope (Zeiss, Germany).
Bioinformatics analysis
The Gene Expression Profiling Interactive Analysis (GEPIA;
Statistical analysis
Data were analyzed by using the unpaired, two-tailed Student’s t-test with GraphPad Prism 5.0 software (GraphPad Software, La Jolla, CA, USA). Values are presented as the mean ± standard deviation. Differences were considered to be statistically significant at p<0.05 and highly significant at p<0.01.
Results
Metastatic melanoma cells exhibited stronger EMT phenotype compared to primary melanoma cells
We gained two metastatic cell lines of melanoma by injecting primary melanoma cells into mice through tail vein. The metastatic cells from C57BL/6 mice and BALB/C nude mice were named B16F10M and A375M, respectively. Furthermore, we compared the EMT-related phenotype between primary cells and metastatic cells through migration, invasion and proliferation assays in vitro. The results showed that the metastatic melanoma cells possessed stronger invasion, migration and proliferation ability compared to primary melanoma cells (Figure 1A–C).
 | Figure 1 Cell migration, invasion and proliferation comparison between primary melanoma cells (B16F10 and A375) and metastatic melanoma cells (B16F10M and A375M). (A) Cell invasion was evaluated by transwell assay. (B) Cell proliferation was validated by colony formation assay. (C) Cell mobility was analyzed by wound-healing assay. Metastatic melanoma cells showed obviously higher invasion, migration and proliferation capabilities compared to primary melanoma cells. Representative pictures and quantification of each assay were shown. Data were presented as mean ± SD. **P<0.01. |
The expression of FN1 was upregulated in metastatic melanoma cells compared to primary melanoma cells and its overexpression was associated with poor prognosis
RT-qPCR and western blot assay showed that the expression of FN1 was upregulated in metastatic melanoma cells both in mRNA and protein level compared to primary melanoma cells (Figure 2A and B). In addition, we detected higher FN1 expression in metastatic sample than in primary sample from TCGA (Figure 2C). Examination of the correlation between FN1 upregulation and clinical pathological characteristic showed that increased FN1 expression was correlated with advanced stage metastases (Figure 2D).
 | Figure 2 FN1 was upregulated in metastatic melanoma cells compared to primary melanoma cells, and increased expression of FN1 was associated with poor prognosis in SKCM patients. (A) The RT-qPCR result showed that the FN1 transcriptional level was increased in metastatic melanoma cells compared to primary melanoma cells. (B) Western blotting showed FN1 protein expression level was higher in metastatic melanoma cells than in primary melanoma cells. (C) FN1 expression in distant metastatic tumors was higher than that in primary tumors. Statistical significance was derived using Wilcoxon Rank test. The raw data were downloaded from TCGA. (D) The relationship between expression of FN1 and clinical characteristics of skin cutaneous melanoma. The result showed that upregulation of FN1 was positively correlated with advanced stage and metastases. Statistical significance was derived using Chi-square test. Data were presented as mean ± SD. *P<0.05; **P<0.01. Abbreviations: FN1, fibronectin 1; TCGA, The Cancer Genome Atlas; SKCM, Skin Cutaneous Melanoma. |
Downregulation of FN1 inhibits proliferation, adhesion, invasion, migration and mobility of metastatic melanoma cells
To reveal the effect of FN1 on metastasis-related phenotypes, B16F10M and A375M cells were transfected with negative control siRNA (NC-siRNA) or FN1-siRNA. RT-qPCR assays showed that FN1-siRNA transfection in B16F10M and A375M cells decreased the mRNA levels of FN1 (Figure 3A). We performed adhesion assay to explore the effect of FN1 on tumor cell adhesion to matrix. As shown in Figure 3B, downregulation of FN1 in B16F10M and A375M cells significantly inhibited the adhesion of rhodamine-labeled cells to gelatin. For transwell invasion assay, downregulation of FN1 significantly decreased the cells penetrating the matrigel (Figure 3C). For migration assay, B16F10M and A375M cells transfected with FN1-siRNA significantly reduced the cell migration capability (Figure 4A). The cell proliferation was assessed using MTT assay and colony formation assay. In FN1-siRNA-treated B16F10M and A375M cells, the cell proliferation capability was significantly decreased (Figure 4B and D). The wound-healing assay showed that reduced FN1 expression inhibited tumor cells mobility (Figure 4C).
 | Figure 3 Downregulation of FN1 in metastatic melanoma cells significantly inhibited cell adhesion and invasion. (A) The RT-qPCR assay was used to validate the interference of FN1 in metastatic melanoma cells. (B) The cells' adhesive ability was analyzed by adhesion assay. (C) Cell invasion was measured by transwell invasion assay. FN1-decreased cells showed obviously lower penetration and adhesion ability compared to NC group. Representative pictures of these data were shown. Data are presented as mean ± SD. **P<0.01. Abbreviations: FN1, fibronectin 1; NC, negative control. |
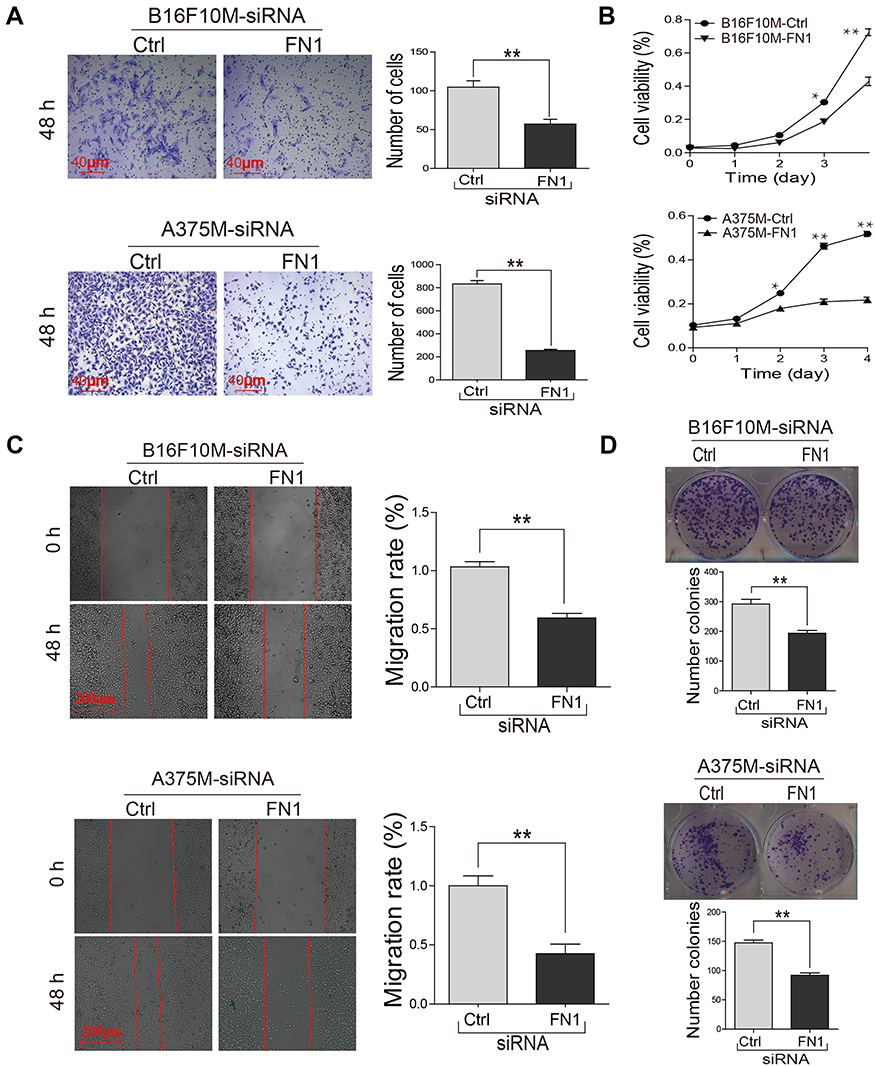 | Figure 4 Knockdown of FN1 in metastatic melanoma cells significantly inhibited cell proliferation, migration and viability. (A) The cell migration was measured by transwell assay. (B) The growth curves were generated based on MTT assay. (C) The cell mobility was performed by wound-healing assay. (D) The cell proliferation was measured by clone formation assay. Representative pictures of the data were shown. The results showed that knockdown of FN1 in metastatic cells significantly inhibited cell proliferation, migration and viability. Data were presented as mean ± SD. *P<0.05; **P<0.01. Abbreviations: FN1, fibronectin 1. |
Downregulation of FN1 blocks the cell cycle progress cycle and induces apoptosis of metastatic melanoma cells
In order to investigate the role of FN1 in cell cycle, B16F10M and A375M cells were transfected with FN1-siRNA or NC-siRNA. Then, the cell cycle of these cells was measured by using the FACS AriaIII flow cytometer. Results indicated that the cell number of G0/G1 phase notably increased and cell number of G2/M phase decreased in FN1-decreased cells (Figure 5A).
 | Figure 5 Downregulation of FN1 blocks the cell cycle progress and induces apoptosis of metastatic melanoma cells. (A) Downregulation of FN1 decreased cell number of G2/M phase in metastatic melanoma cells. (B) Downregulation of FN1 triggered cell apoptosis in metastatic melanoma cells. Representative pictures were shown. Data were presented as mean ± SD. Abbreviations: FN1, fibronectin 1. |
Cell apoptosis was determined by using the FACS AriaIII flow cytometer. As shown in Figure 5B, downregulation of FN1 in B16F10M and A375M cells significantly increased the cell apoptosis compared to NC-siRNA treated cells. Moreover, we found downregulation of FN1 significantly decreased the expression of Bcl-2 and increased the expression of Bax (Figure 6A and B).
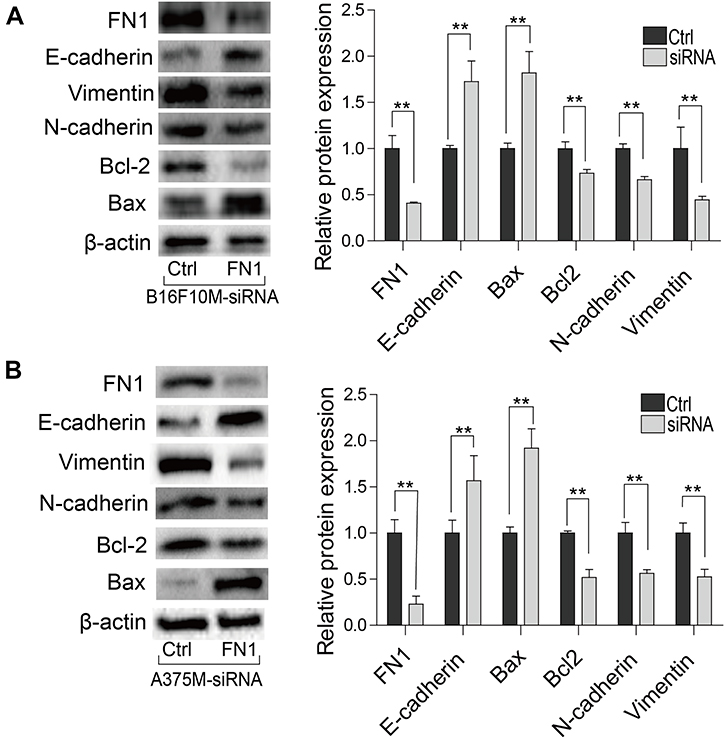 | Figure 6 Effect of FN1 on protein expression in metastatic melanoma cells. (A) Mouse metastatic melanoma cells and (B) human metastatic melanoma cells were transfected with NC-siRNA or FN1-siRNA, and the expression of FN1, BCL2, E-cadherin, N-cadherin and Bax were measured by western blot. The results showed that downregulation of FN1 increased the expression of E-cadherin and Bax, and decreased the expression of N-cadherin, Vimentin and Bcl2. Data were presented as mean ± SD. **P<0.01. Abbreviations: FN1, fibronectin 1; NC, negative control. |
Downregulation of FN1 suppresses the expression of EMT markers in metastatic melanoma cells
To further reveal the possible mechanism of FN1 regulating melanoma cells metastasis, firstly, we studied the role of FN1 in EMT. Western blot showed that FN1-siRNA effectively reduced the FN1 protein levels in metastatic cells. Besides, we found that the expression of E-cadherin increased while N-cadherin and Vimentin decreased in FN1 knockdown cells (Figure 6A and B). In consistent with previous studies, FN1 knockdown reduced the cell migration, invasion and adhesion capabilities dramatically. Furthermore, the protein–protein interaction (PPI) network of proteins that closely correlated with FN1 was established using STRING (Figure 7A). A positive correlation among FN1, ITGB1, ITGA5, TGFB1, HGF, ITGA4 and ITGB3 was uncovered (Figure 7B). In addition, the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of this protein in the PPI network showed that FN1 was tightly associated with focal adhesion, PI3K-AKT signaling pathway and ECM-receptor interaction (Figure 7C). These data suggest that FN1 regulates EMT via interacting with integrin and downstream signaling pathways. Further research is needed to confirm this.
 | Figure 7 The possible signaling pathways by which FN1 regulated cancer metastasis. (A) The PPI network of proteins that were tightly associated with FN1. Search Tool for the Retrieval of Interacting Genes (STRING) was used to make the PPI network. (B) The correlation among FN1, ITGB1, ITGA5, TGFB1, HGF, ITGA4 and ITGB3 was analyzed by GEPIA, and the positive correlations were shown. (C) KEGG pathway analysis of proteins in PPI network showed that FN1 was closely associated with tumor progression including focal adhesion, PI3K-AKT and ECM-receptor signaling pathway. Abbreviations: FN1, fibronectin 1; PPI, protein–protein interaction; FDR, False Discovery Rate; KEGG, Kyoto Encyclopedia of Genes and Genomes. |
Discussion
Metastasis is the major cause of mortality in cancer patients. Although there are many treatment strategies for primary cancer, there is a lack of effective warning and prevention methods for tumor metastasis. Thus, identifying specific markers associated with tumor metastasis may be useful for alertness and prevention of cancer metastasis. It has been reported that cancer cells must make molecular alterations to get specific phenotypes for completing metastasis. In this study, we focused on the changes in expression of FN1 that survived from melanoma metastasis and tried to explain the effects of changed FN1 expression on melanoma metastasis. We established two metastatic cell lines of melanoma through mouse model (Figure S2) and confirmed that FN1 was upregulated in metastatic melanoma cells compared to primary cells in vitro both in protein and mRNA level. Overexpression of FN1 was also reported in hepatocellular carcinoma, renal carcinoma, gastrointestinal carcinoma and head/neck cancer.23,24 Clinical study had revealed that upregulated FN1 was significantly associated with poor prognosis in renal cancer and led to lymphangiogenesis and lymph node metastasis in oral squamous cell carcinoma.25,26 Similarly, we demonstrated increased FN1 expression was evidently associated with metastases and advanced stage in melanoma. In consistence with those known features of FN1, the data from GEPIA confirmed that FN1 expression in tumorous tissues was higher than that in normal non-tumorous tissues (Figure S1A) and the data from the Cancer Cell Line Encyclopedia showed that FN1 was highly expressed in melanoma compared with other cell types (Figure S1B). In addition, we demonstrated that FN1 expression in metastatic tumor was higher than that in primary tumor by using raw sequence data and clinical information from TCGA database. It has been reported that FN1 is an important protein associated with cell migration, apoptosis, invasion and metastasis. The expression of FN1 is tightly related to the development of various malignant tumors. All these data suggest that FN1 upregulation might play an important role in melanoma metastasis.
EMT is a process by which epithelial cells lose their cell polarity and cell–cell adhesion to become mesenchymal-like cells which acquire migratory and invasive properties.27 EMT is important for the cancer metastasis. In our study, we found that metastatic melanoma cells exhibited stronger EMT phenotype with increased capabilities of migration, invasion and proliferation compared to primary cells. This is in consistence with the notion that metastatic tumor cells survived from metastasis possess stronger metastasis-related abilities. To further reveal the role of FN1 in EMT and cell apoptosis, we disturbed the expression of FN1 in metastatic cells. Similar to previous studies,28,29 our results showed that downregulation of FN1 suppressed cell proliferation, migration, invasion and induced cell apoptosis. Moreover, we found that downregulation of FN1 led to an increased expression of E-cadherin and a decreased expression of N-cadherin and Vimentin. Notably, FN1 knockdown in metastatic tumor cells suppressed the EMT process. These data also implied that down-regulation of FN1 induced apoptosis by increasing Bax/Bcl-2 ratio. Thus, we revealed that FN1 upregulation in melanoma might promote melanoma proliferation and metastasis by inhibiting apoptosis and regulating EMT.
Several signaling pathways including PI3K-AKT, TGF-β, EGF, FGF, FAK and Wnt/β-catenin have been shown to regulate EMT.30–32 In this study, we collected some proteins tightly associated with FN1 and made a protein–protein interaction network through STRING. Then, a positive correlation among FN1, ITGB1, ITGA5, TGFB1, HGF, ITGA4 and ITGB3 was uncovered. The KEGG pathway analysis of these proteins in the PPI network showed that FN1 was tightly associated with focal adhesion, PI3K-AKT and ECM-receptor signaling pathway. These data suggest that EMT regulated by FN1 via interacting with integrin and downstream signaling pathways in melanoma. Extensive studies are needed for the precise mechanism.
Conclusion
Our study found that FN1 survived from melanoma metastasis and that FN1 expression level was increased in metastatic tumor cells. The suppression of the FN1 resulted in impaired cell proliferation, increased apoptosis and reduced migration and invasion through regulating EMT process. All these data suggested that FN1 might play a pivotal role in melanoma metastasis, and could be a therapeutic target to alert and prevent melanoma metastasis. However, we have only predicted some signaling pathways regulating EMT process, and further studies are needed to validate this prediction in melanoma.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (NSFC) (U1505225, 81773063, 81273548, 81571802); the Natural Science Foundation of Fujian Province (2016J06020).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Cohen JD, Li L, Wang Y, et al. Detection and localization of surgically resectable cancers with a multi-analyte blood test. Science. 2018;359(6378):eaar3247. doi:10.1126/science.aar3247
2. Kibbi AG, Mihm MC
3. Mortality G;
4. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2010;60(5):277–300. doi:10.3322/caac.20073
5. Bhatia S, Tykodi SS, Thompson JA. Treatment of metastatic melanoma: an overview. Oncology. 2009;23(6):488.
6. Azoury SC, Lange JR. Epidemiology, risk factors, prevention, and early detection of Melanoma. Surg Clin North Am. 2014;94(5):945–962. doi:10.1016/j.suc.2014.07.013
7. Maheswaran S, Haber DA. Circulating tumor cells: a window into cancer biology and metastasis. Curr Opin Genet Dev. 2010;20(1):96–99. doi:10.1016/j.gde.2009.12.002
8. Quail D, Joyce J. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19(11):1423–1437. doi:10.1038/nm.3394
9. Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol. 2006;7(2):131–142. doi:10.1038/nrm1835
10. Luo WR, Chen XY, Li SY, Wu AB, Yao KT. Neoplastic spindle cells in nasopharyngeal carcinoma show features of epithelial-mesenchymal transition. Histopathology. 2012;61(1):113–122. doi:10.1111/j.1365-2559.2012.04205.x
11. Wasil LR, Shair KHY. Epstein–Barr virus LMP1 induces focal adhesions and epithelial cell migration through effects on integrin-α5 and N-cadherin. Oncogenesis. 2015;4(10):e171. doi:10.1038/oncsis.2015.31
12. Gao W, Liu Y, Qin R, Liu D, Feng Q. Silence of fibronectin 1 increases cisplatin sensitivity of non-small cell lung cancer cell line. Biochem Biophy Res Commun. 2016;476(1):35–41. doi:10.1016/j.bbrc.2016.05.081
13. Pankov R, Yamada KM. Fibronectin at glance. J Cell Sci.2002;115(Pt 20):3861–3863.
14. Williams CM, Engler AJ, Slone RD, Galante LL, Schwarzbauer JE. Fibronectin expression modulates mammary epithelial cell proliferation during acinar differentiation. Cancer Res. 2008;68(9):3185. doi:10.1158/0008-5472.CAN-07-2673
15. Yen CY, Huang CY, Hou MF, et al. Evaluating the performance of fibronectin 1 (FN1), integrin α4β1 (ITGA4), syndecan-2 (SDC2), and glycoprotein CD44 as the potential biomarkers of oral squamous cell carcinoma (OSCC). Biomarkers. 2013;18(1):63–72. doi:10.3109/1354750X.2012.737025
16. Ma LJ, Lee SW, Lin LC, et al. Fibronectin overexpression is associated with latent membrane protein 1 expression and has independent prognostic value for nasopharyngeal carcinoma. Tumour Biol. 2013;35(2):1703–1712. doi:10.1007/s13277-013-1235-8
17. Lou X, Han X, Jin C, et al. SOX2 targets Fibronectin 1 to promote cell migration and invasion in Ovarian cancer: new molecular leads for therapeutic intervention. Omics. 2013;17(10):510–518. doi:10.1089/omi.2013.0058
18. Waalkes S, Atschekzei F, Kramer MW, et al. Fibronectin 1 mRNA expression correlates with advanced disease in renal cancer. BMC Cancer. 2010;10(1):1–6. doi:10.1186/1471-2407-10-1
19. Cai X, Liu C, Zhang TN, Zhu YW, Dong X, Xue P. Down-regulation of FN1 inhibits colorectal carcinogenesis by suppressing proliferation, migration and invasion. J Cell Biochem. 2017 ;119(6):4717-4728. doi: 10.1002/jcb.26651.
20. Han S, Khuri FR, Roman J. Fibronectin stimulates non-small cell lung carcinoma cell growth through activation of Akt/mammalian target of rapamycin/S6 kinase and inactivation of LKB1/AMP-activated protein kinase signal pathways. Cancer Res. 2006;66(1):315–323. doi:10.1158/0008-5472.CAN-05-2367
21. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(WebServer issue). doi:10.1093/nar/gkx247
22. Katarzyna T, Patrycja C, Maciej W. The Cancer Genome Atlas (TCGA): an immeasurable source of knowledge. Contemp Oncol. 2015;19(1A):68–77.
23. Torbenson M, Wang J, Choti M, et al. Hepatocellular carcinomas show abnormal expression of fibronectin protein. Mod Pathol. 2002;15(8):826–830. doi:10.1097/01.MP.0000024257.83046.7C
24. Warawdekar UM, Zingde SM, Iyer KSN, Jagannath P, Mehta AR, Mehta NG. Elevated levels and fragmented nature of cellular fibronectin in the plasma of gastrointestinal and head and neck cancer patients. Clinica Chimica Acta. 2006;372(1):83–93. doi:10.1016/j.cca.2006.03.023
25. Boguslawska J, Kedzierska H, Poplawski P, Rybicka B, Tanski Z, Piekielko-Witkowska A. Expression of genes involved in cellular adhesion and extracellular matrix remodeling correlates with poor survival of patients with renal cancer. J Urol. 2016;195(6):1892–1902. doi:10.1016/j.juro.2015.11.050
26. Morita Y, Hata K, Nakanishi M, et al. Cellular fibronectin 1 promotes VEGF-C expression, lymphangiogenesis and lymph node metastasis associated with human oral squamous cell carcinoma. Clin Exp Metastasis. 2015;32(7):739–753. doi:10.1007/s10585-015-9741-2
27. Meng J, Zhang XT, Liu XL, et al. WSTF promotes proliferation and invasion of lung cancer cells by inducing EMT via PI3K/Akt and IL-6/STAT3 signaling pathways. Cell Signal. 2016;28(11):1673–1682. doi:10.1016/j.cellsig.2016.07.008
28. Jia D, Yan M, Wang X, et al. Development of a highly metastatic model that reveals a crucial role of fibronectin in lung cancer cell migration and invasion. BMC Cancer. 2010;10(1):364. doi:10.1186/1471-2407-10-364
29. Wang F, Song G, Liu M, Li X, Tang H. miRNA-1 targets fibronectin1 and suppresses the migration and invasion of the HEp2 laryngeal squamous carcinoma cell line. FEBS Lett. 2011;585(20):3263–3269. doi:10.1016/j.febslet.2011.08.052
30. Wang Y, Shi J, Chai K, Ying X, Zhou BP. The role of snail in EMT and tumorigenesis. Curr Cancer Drug Targets. 2013;13(9):963–972.
31. Micalizzi DS, Farabaugh SM, Ford HL. Epithelial-mesenchymal transition in cancer: parallels between normal development and tumor progression. J Mammary Gland Biol Neoplasia. 2010;15(2):117–134. doi:10.1007/s10911-010-9178-9
32. Pal A, Gascard P, Ravani S, Barcelloshoff M. Inhibition of transforming growth factor beta1 (TGFβ1) signaling increases radiosensitivity in breast cancer cell lines. Cancer Res. 2008;68:423.
Supplementary Materials
 | Figure S1 Clinical evidences for the overexpression of FN1 in melanoma. (A) The data from GEPIA confirmed that FN1 expression in tumorous tissues was higher than in normal non-tumorous tissues. (B) The data from the Cancer Cell Line Encyclopedia showed that FN1 was highly expressed in melanoma compared with other cell types. |
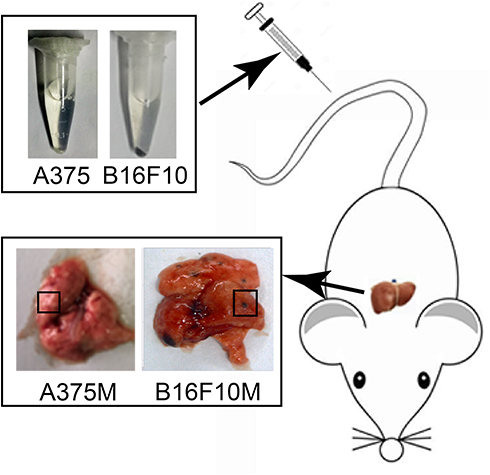 | Figure S2 Schematic of the experiment metastatic model used in the study. B16F10 cells and A375 cells were intravenously injected into mice through the tail vein, and the pulmonary metastases were stripped for the later experiment. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.