Back to Journals » Cancer Management and Research » Volume 15
Fibroblast Growth Factor 11 (FGF11) Promotes Progression and Cisplatin Resistance Through the HIF-1α/FGF11 Signaling Axis in Ovarian Clear Cell Carcinoma
Authors Yin M ![]() , Lu C
, Lu C ![]() , Zhou H, Liu Q, Yang J
, Zhou H, Liu Q, Yang J
Received 18 April 2023
Accepted for publication 19 July 2023
Published 26 July 2023 Volume 2023:15 Pages 753—763
DOI https://doi.org/10.2147/CMAR.S414703
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lu-Zhe Sun
Min Yin,1,* Chunli Lu,2,* Huimei Zhou,1 Qian Liu,1 Jiaxin Yang1
1National Clinical Research Center for Obstetric and Gynecologic Diseases, Department of Obstetrics and Gynecology, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, People’s Republic of China; 2Neurospine Center, China International Neuroscience Institute (CHINA-INI), Xuanwu Hospital, Capital Medical University, National Center for Neurological Disorders, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Huimei Zhou; Jiaxin Yang, Email [email protected]; [email protected]
Background: A poor prognosis is often associated with ovarian clear cell carcinoma (OCCC) due to its relative resistance to platinum-based chemotherapy. Although several studies have been launched to explore the pathogenesis of OCCC, the mechanism of chemoresistance has yet to be uncovered.
Methods: Nanostring nCounter PanCancer Pathways Panel was performed to explore the expression profiles of OCCC tissues from patients showing different platinum sensitivity. Bioinformatic analysis was performed to select genes associated with chemoresistance and cell function assays, including colony formation, wound healing, transwell and flow cytometric analysis, were used to explore the role of the target gene in the progression of OCCC and resistance to cisplatin (DDP).
Results: Gene expression profiles and bioinformatic analysis verified that the expression of fibroblast growth factor 11 (FGF11) was significantly increased in platinum-resistant OCCC tissues and increased FGF11 expression was related to poorer survival. Downregulation of FGF11 inhibited the proliferation, migration, and invasion, reversing the DDP resistance of OCCC cells. Mechanically, FGF11 was regulated by hypoxia-inducible factor-1α (HIF-1α) to modulate the DDP sensitivity.
Conclusion: FGF11 was highly expressed in platinum-resistant OCCC tissues, promoting progression and resistance to DDP through the HIF-1α/FGF11 signaling axis.
Keywords: ovarian clear cell carcinoma, cisplatin chemoresistance, FGF11, HIF-1α
Introduction
Ovarian cancer is the primary cause of mortality among gynecological cancers.1 Epithelial ovarian cancer (EOC) can be divided into two types. Type I EOCs are generally believed to be tumors that develop slowly and maintain genetic stability. These tumors often emerge from identifiable precursor conditions such as endometriosis or tumors with low malignant potential. Conversely, type II EOCs are thought to be highly aggressive tumors from their inception and have a higher likelihood of metastasizing from smaller primary tumors.2 In addition, EOC is classified into five subtypes based on histological criteria: high-grade serous, low-grade serous, mucinous cancer, endometrioid cancer, and clear cell carcinoma (OCCC).
OCCC is a distinct histological entity of EOC with different clinicopathologic characteristics including molecular features and chemosensitivity as well as patient outcomes.3 OCCC is less common, and accounts for only approximately 10–25% of patients with EOC. OCCC exhibits a distinct genetic profile from high-grade serous ovarian cancer. It has a lower frequency of p53 mutations (25%) and BRCA1/2 mutations (6.3%), but has higher mutations rates in ARID1A, PIK3CA, and PTEN. Due to the role of inflammatory and epigenetic mechanisms in the development of OCCC, therapies targeting the PI3K pathway, immune checkpoint inhibitors, and epigenetic treatments could be integral to its treatment.4,5
Although a platinum-based regimen is typically preferred as first-line chemotherapy, OCCC is relatively resistant to chemotherapy, leading to a poor prognosis.6 Response rates of high-grade serous ovarian cancer to conventional platinum-based chemotherapy reach 80%, but the response rate of OCCC ranges from only 11% to 56% in the first-line chemotherapy.7 In OCCC, numerous platinum resistance mechanisms have been identified such as reduced drug accumulation, elevated drug detoxification, and augmented DNA repair activity.6,8,9 Additionally, resistance to chemotherapy has been linked with decreased proliferation activity.10 Despite these findings, the specific causes of resistance to platinum-based chemotherapy have yet to be fully comprehended. Gene expression continues to be an important aspect as cancer therapy strategies broaden to personalized and targeted treatment. Technologies, such as proteomics, have advanced the underlying molecular signaling events and the characteristics of ovarian cancer. New therapeutic options can be discovered through these studies, drug resistance may decrease, and patient outcomes could potentially improve.11
In this study, we explored the molecular alterations among platinum-resistant and platinum-sensitive OCCC patients. We found that the expression of fibroblast growth factor 11 (FGF11) was significantly up-regulated in platinum-resistant OCCC tissues and higher expression of FGF11 was associated with poorer survival. Further functional studies showed that downregulation of FGF11 inhibited the proliferation, migration, and invasion, reversing the cisplatin (DDP) resistance of OCCC cells. Mechanically, FGF11 was regulated by hypoxia-inducible factor-1α (HIF-1α) to modulate the DDP sensitivity.
Materials and Methods
Patients
This study was conducted on patients who had surgery and were diagnosed with OCCC at stages II–IV according to the International Federation of Gynecology and Obstetrics (FIGO) between January 2019 and December 2021. The exclusion criteria consisted of being diagnosed with mixed subtypes via histopathology, not receiving standard platinum-based adjuvant chemotherapy following surgery, and suffering from chronic systemic ailments or other malignant tumors. According to their response to first-line platinum-based chemotherapy, patients were classified into two groups: the platinum-resistant (PR) group and the platinum-sensitive (PS) group. Patients were considered platinum-resistant or platinum-sensitive if they had relapsed within 6 months or 6 to 12 months after receiving platinum-based treatment, respectively. After a selection process, a total of 24 patients were chosen, evenly divided into two groups of 12 patients each.
RNA Extraction and Nanostring nCounter Analysis
Total RNA was extracted from formalin-fixed paraffin-embedded (FFPE) tissues and quality check were performed as we previously described.12 NanoString nCounter is a multiplex nucleic acid hybridization technology that allows accurate and reproducible measurement of gene expression.13 As previously described, we conducted gene expression profiling using the PanCancer Pathways Panel and analyzed the data using nSolver 4.0 software.12
Bioinformatic Analysis
The FGF11 expression data in ovarian cancer and normal tissues were obtained from The Cancer Genome Atlas (TCGA) and the Genotype-Tissue Expression (GTEx) databases. It was analyzed and visually displayed in boxplot format in R using ggplot. The Kaplan-Meier plotter (http://kmplot.com/analysis/) is an online tool to assess the correlation between gene expression and survival in tumors, including ovarian cancer. By utilizing the Kaplan-Meier survival curve, the examination of overall survival (OS) and progression-free survival (PFS) among various subgroups of FGF11 expression levels was conducted.
Cell Culture
ES-2 and TOV-21G cell lines of human ovarian clear cell carcinoma (OCCC) were acquired from the American Type Culture Collection (ATCC, USA). The cells were cultured with DMEM (Gibco, USA) supplemented with 10% fetal bovine serum (Gibco, USA), penicillin (100 U/mL), and streptomycin (100 ng/mL) (Gibco, USA) at 37°C, in a humidified incubator containing 5% CO2.
Plasmid Construction and Viral Transfection
GeneChem (Shanghai, China) constructed lentiviral plasmids that contained negative control shRNA (NC), as well as specific shRNAs targeting FGF11 (shFGF11) and HIF-1α (shHIF-1α). In order to transfect the cells, Lipofectamine 2000 (Invitrogen, USA) was used following the manufacturer’s instructions. After seeding OCCC cells in a 24-well plate and allowing them to reach 50–60% confluence, recombinant lentivirus was used for cell infection. Stably transduced cells were screened by puromycin for one week.
Quantitative Real‑time Polymerase Chain Reaction (qRT‑PCR)
The cultured cells were subjected to RNA extraction using Trizol (Invitrogen, USA). The extracted RNA was then reverse transcribed utilizing a PrimeScript RT Reagent kit (TaKaRa, Japan). The expression was examined on a 7500 Fast Real-Time PCR (Applied Biosystems, USA) by following the SYBR-Green PCR Master Mix Kit (Takara, Japan) instructions. The 2−ΔΔCt method was used to calculate the expression levels with β-actin serving as an internal control. All RT reactions were carried out in triplicates. The primer sequences were as follows: FGF11 forward, 5’-GGCATCGTCACCAAACTGTT-3’ and reverse, 5’-GCAGTCCCTCAGCATTCATG-3’; HIF-1α forward, 5’-GACAGCCTCACCAAACAGAG-3’ and reverse, 5’-GTAGCTGCATGATCGTCTGG-3’; β-actin forward, 5’- CGTGCGTGACATCAAAGAGAAG-3’ and reverse, 5’-CCAAGAAGGAAGGCTGGAAAA-3’.
Western Blot Analysis
Total protein was extracted from cells using RIPA buffer (Solarbio, China) mixed with 1% protease inhibitor PMSF (Solarbio, China). Once quantified by the BCA method, protein samples underwent SDS-PAGE separation and were transferred onto PVDF membranes (Millipore, USA). Following blocking with 5% non-fat milk (Solarbio, China), the membranes were left to incubate at 4 °C overnight with primary antibodies against FGF11 (1:1000; Abcam, ab89713, UK), HIF-1α (1:1000; Abcam, ab179483, UK), and β-tubulin (1:5000; Abcam, ab6046, UK). After washing with PBST, the membrane was incubated with an HRP-linked secondary antibody. An enhanced chemiluminescence kit was used to visualize the protein bands (Sangon, China). Image J (V2.0) was used to quantify the bands. Three independent experiments were performed for final analyses.
Colony Formation Assay
Cells were seeded at a density of 500 cells per well in six-well plates and cultured for two weeks. Every three days, the medium was replaced. The colonies were enumerated under a microscope after being fixed with 4% paraformaldehyde for 15 minutes and stained with 0.1% crystal violet (Solarbio, China). Three replicates per sample were measured.
Cell Migration and Invasion Assay
The cells were seeded on a six-well plate and a scratch was made on the cell monolayer using a sterile 100μL tip when the confluency reached 70%-80%. The cells were then cultured for another 24 hours without serum. Migration images were acquired at 0 h and 24h after scraping.
Invasive abilities were evaluated using 24-well transwell chamber inserts. Matrigel (BD, USA) was applied onto the top surface of the chamber, and the upper chamber was seeded with starved cells in a serum-free medium, while the bottom chamber was filled with a medium containing 10% serum. Twenty-four hours later, using a cotton swab, the cells in the upper chamber were taken out, and the invasive cells were fixed and stained with 0.1% crystal violet. By examining five random fields at a magnification of 100, the number of invasive cells was counted. Each sample was performed in triplicate.
Apoptosis Assay by Flow Cytometry
Cells were plated in six-well culture plates for overnight incubation. The half-maximal drug inhibitory concentration (IC50) of DDP in ES-2 cells was measured using the cell counting kit-8 as we previously described, and the IC50 was 52 µM.14 In this study, the experimental and negative control (NC) groups ES-2 cells were treated with 50 µM DDP (Sigma-Aldrich; Millipore) for 48h. Then cells were harvested by trypsinization and washed twice in ice-cold PBS. Apoptosis was assessed via flow cytometry after cells were stained with Annexin V-FITC detection kit (eBioscience, USA). Collected data were analyzed by Flow Jo software (Version 13.0, BD). Each sample was performed in triplicate.
Statistical Analysis
The mean ± standard deviation was used to present the data obtained from the in vitro experiments. Student’s t-test was used to assess the variations between the two groups, and the results were analyzed using GraphPad Prism 9.0 software (GraphPad Software, USA). If P value was less than 0.05, it was deemed to be a significant statistical difference.
Results
FGF11 Was Up-Regulated in Platinum-Resistant OCCC Tissues and Associated with Poor Prognosis
A total of 32 differentially expressed genes (DEG) were identified between the PR and PS groups (P < 0.05 and |log2FC| >1) (Table 1). Among them, 17 genes were up-regulated and 15 genes were down-regulated (Figure 1A). Based on the gene set defined by the PanCancer Pathways Panel, most DEGs were involved in the MAPK pathway, PI3K pathway, and Cell Cycle-Apoptosis pathway (Figure 1B). Some genes were involved in two or all these three pathways, and FGF11 was the most significant up-regulated (Figure 1C).
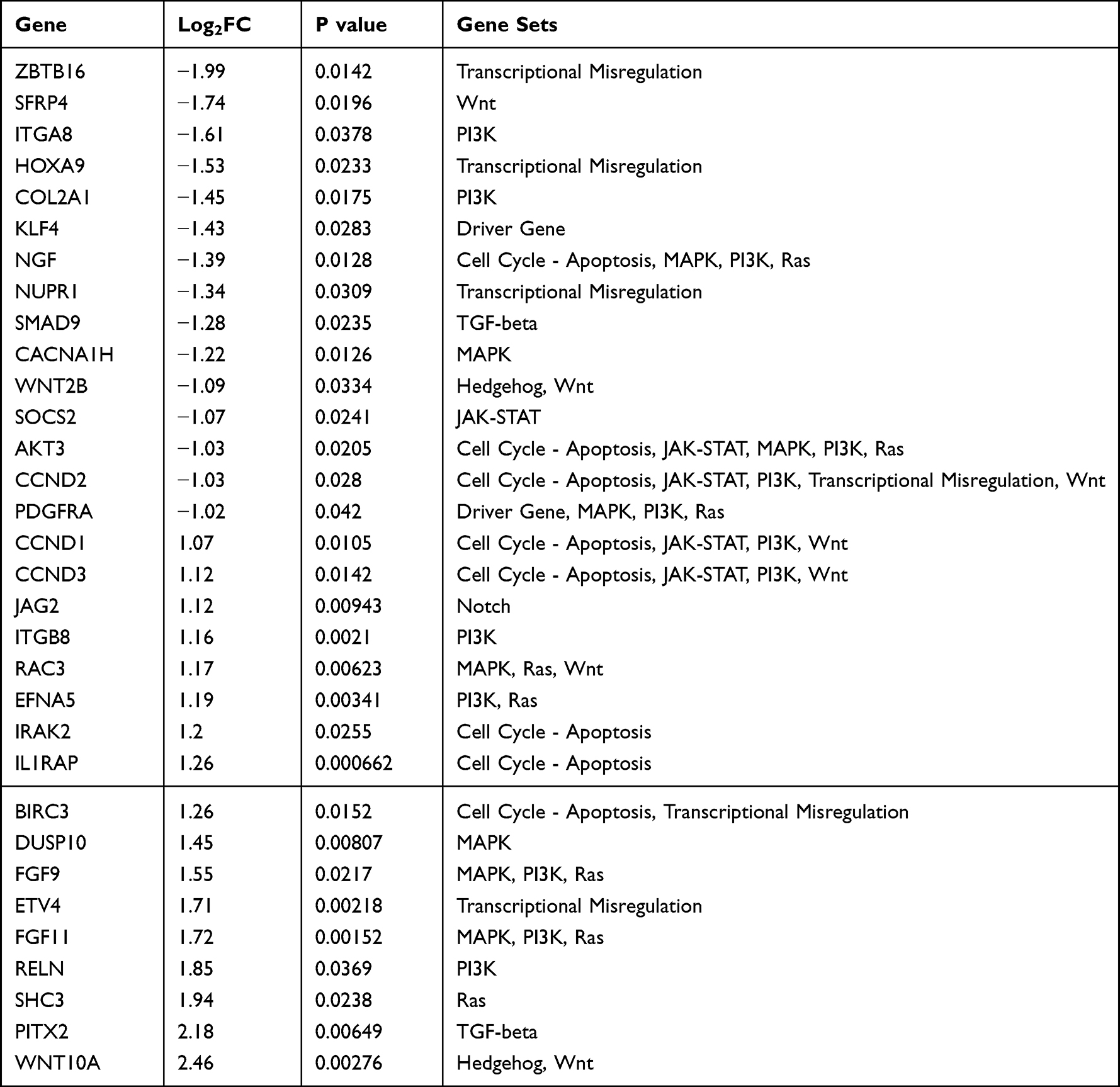 |
Table 1 List of 32 Differentially Expressed Genes Between PR and PS Groups |
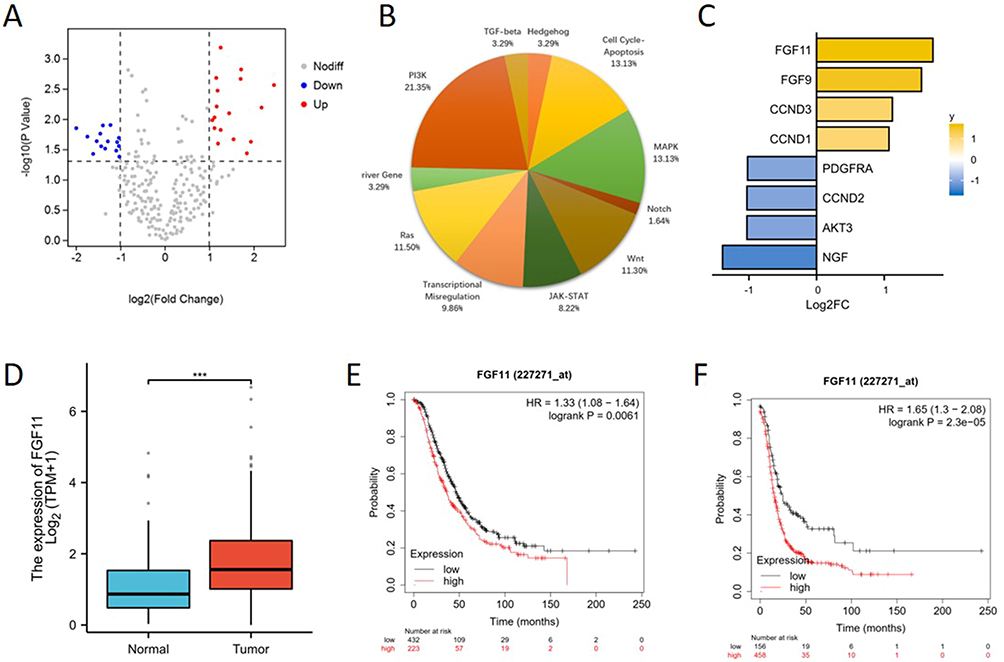 |
Figure 1 FGF11 is up-regulated in platinum-resistant OCCC tissues and associated with poor prognosis. (A) The differentially expressed genes (DEGs) in the platinum-resistant group compared to the platinum-sensitive group. (B) The distribution of pathways of DEGs based on the gene set defined by the PanCancer Pathways Panel. (C) Out of the PI3K pathway, the MAPK pathway, and the Cell Cycle-Apoptosis pathway, eight genes have been involved in two or all of them. (D) The mRNA expression level of FGF11 in ovarian cancer and normal ovarian tissues from the TCGA and GTEx databases. (E) Kaplan-Meier survival curve for overall survival (HR=1.33, 95% CI 1.08–1.64, P=0.0061). (F) Kaplan-Meier survival curve for progression-free survival (HR=1.65, 95% CI 1.3–2.08, P=2.3e-05). ***, P < 0.001. |
Then we analyzed the expression level of FGF11 in ovarian cancer and normal tissues from the TCGA and GTEx databases. The expression level of FGF11 was shown to be significantly higher in ovarian cancer than in normal tissues (Figure 1D). To further evaluate the clinical significance of FGF11 on prognosis, the Kaplan-Meier plotter showed that a high level of FGF11 was associated with poor OS (HR=1.33, 95% CI 1.08–1.64, P=0.0061) and PFS (HR=1.65, 95% CI 1.3–2.08, P=2.3e-05) (Figure 1E and F).
Downregulation of FGF11 Inhibited OCCC Cell Proliferation, Migration and Invasion
After observing a significant increase in FGF11 expression in platinum-resistant OCCC tissues as previously mentioned, we proceeded to investigate the impact of FGF11 on OCCC cells. Through lentiviral transduction, we successfully created shFGF11 ES-2 and TOV-21G cell lines with stable FGF11 knockdown (Figure 2A and B). The results of colony formation experiments demonstrated a decrease in cell proliferation ability in the shFGF11 groups when compared to the NC groups, indicating that the knockdown of FGF11 can reduce the proliferation ability of OCCC cells (Figure 2C and D). Our findings from wound healing experiments showed a significant decrease in migration capacity in both cell lines following FGF11 knockdown (Figure 2E and F). Additionally, the results of cell invasion assays indicated that FGF11 knockdown also decreased cell invasion in both cell lines when compared to the NC groups (Figure 2G). Thus, our study suggests that FGF11 knockdown can weaken the proliferation, migration and invasion ability of OCCC cells in vitro.
 |
Figure 2 FGF11 knockdown inhibited OCCC cell proliferation, migration and invasion. (A and B) FGF11-knockdown cell lines were established with ES-2 and TOV-21G cells, and the transfection efficiency was confirmed by qRT-PCR and WB assays. (C and D) Colony formation results in ES-2 and TOV-21G cells. (E and F) Wound healing results in ES-2 and TOV-21G cells. (G) Transwell results in ES-2 and TOV-21G cells. **, P <0.01. |
Downregulation of FGF11 Reversed DDP Resistance in OCCC
To explore whether FGF11 could influence chemoresistance in OCCC, we conducted apoptosis assays in ES-2 cell line. As shown in Figure 3, shFGF11 group had a noteworthy increase in the number of apoptotic cells compared to the NC group. Treatment with DDP at a concentration of 50 µM for 48 hours resulted in a significant increase in the number of apoptotic cells in the DDP group compared to the NC group. Furthermore, in comparison to the NC group, the shFGF11+DDP group showed a further increase in the number of apoptotic cells. These findings suggested that FGF11 down-regulation could reverse DDP resistance and work together with DDP by inducing cell apoptosis.
 |
Figure 3 Downregulation of FGF11 could reverse DDP resistance via inducing cell apoptosis. (A and B) Cell apoptosis results in the NC, shFGF11, NC+DDP and shFGF11+DDP groups in ES-2 cells. **, P < 0.01, ***, P < 0.001. |
FGF11 Was Regulated by HIF-1α to Participate in DDP Resistance
In solid tumors, hypoxia of the tumor microenvironment is a common phenomenon. A hypoxic environment promotes the production of hypoxia-inducible factors and activates hypoxia-dependent signaling pathways.15 Hypoxia-inducible factor-1 (HIF-1) is made up of an alpha and a beta subunit (HIF-1α and HIF-β) and is a member of the basic helix-loop-helix family of transcription factors.16 HIF-1α is a key regulatory factor for the cell hypoxia response, involved in tumor invasion, metastasis, angiogenesis, and drug resistance.17 Studies showed that the expression level of HIF-1α in OCCC was significantly higher than that in other subtypes of ovarian cancer, which may be related to OCCC drug resistance.18 Furthermore, FGF11 has been reported to be induced in hypoxic cancer cells.19
To investigate if FGF11 is regulated via the hypoxia signaling pathway in OCCC, we knocked down HIF-1α in ES-2 cell lines. At the same time, the protein level of FGF11 was significantly reduced after the knockdown of HIF-1α (Figure 4A). In order to investigate the potential role of HIF-1α in the regulation of FGF11 and its promotion of chemoresistance to DDP, we performed apoptosis assays in ES-2 cell lines. Our findings showed a significant increase in the number of apoptotic cells in the shHIF-1α group compared to the NC group. Moreover, upon treatment with DDP, the shHIF-1α+DDP group exhibited a significantly greater number of apoptotic cells compared to the NC group (Figure 4B). These results suggest that the knockdown of HIF-1α may have a synergistic effect in vitro with DDP, and that FGF11 plays a role in the modulation of DDP resistance through HIF-1α.
 |
Figure 4 FGF11 was regulated by HIF-1α to participate in DDP resistance. (A) Knockdown of HIF-1α inhibited the expression of FGF11. (B) Cell apoptosis results in the NC, shHIF-1α, and shHIF-1α+DDP groups in ES-2 cell lines. *, P < 0.05; **, P < 0.01. |
Discussion
Chemoresistance is a major cause of a poor prognosis in advanced OCCC, but the mechanisms of resistance in OCCC are still unclear.20 In this study, a total of 32 DEGs were screened by expression profiling of 12 platinum-sensitive and 12 platinum-resistant OCCC samples. We found that FGF11 was overexpressed in platinum-resistant patients and was related to unfavorable survival. In vitro experiments revealed that overexpressed FGF11 promoted cell proliferation, migration, invasion, and resistance to DDP, implying that FGF11 has an oncogenic function. Furthermore, FGF11 was modulated by HIF-1α to participate in DDP resistance.
The tumor microenvironment (TME) plays an important role in malignant biological characteristics, including ovarian cancer.15 Hypoxia is a noticeable trait of TME, which is positively associated with tumor development, angiogenesis, chemotherapy resistance, and tumor spreading.21,22 Numerous studies have found that HIF-1α promotes the progression of ovarian cancer through many biological processes, such as inhibiting the function of p53, promoting IL-6 expression, stimulating the AKT/mTOR pathway, and promoting the glycolysis pathway in ovarian cancer.17 HIF-1α activates hypoxia-dependent signaling pathways and affects the reactions to both chemotherapy and immunotherapy in ovarian cancer.17,23,24 In OCCC, HIF-1α expression levels were significantly higher than in other histological types.18 Moreover, HIF-1α has been detected in putative precursor lesions of OCCC, indicating its involvement in the progression of OCCC.25
Fibroblast growth factors (FGFs) are a kind of polypeptide growth factor that exists widely in various tissues of the body.26 In mammals, there are 22 members of the FGFs family, which can be classified into endocrine, paracrine, typical autocrine, and intracellular FGFs.27 Canonical FGFs include paracrine and autocrine subfamilies, which can bind and trigger the activation of fibroblast growth factor receptors (FGFRs). FGF/FGFR signaling network influences oncogenesis through a variety of mechanisms.28 Many human malignancies, including nasopharyngeal,29 prostate,30 and lung cancers,31 have been shown to have elevated levels of different FGFs, suggesting FGFs/FGFR as therapeutic targets for the treatment of cancer.32
FGF11 is a member of the intracellular FGFs subfamily (FGF11/12/13/14) whose action is not related to FGFRs due to a lack of the N-terminal secretion signal. It is reported that the level of FGF11 expression was greater in cancerous tissue compared to the noncancerous tissue in lung adenocarcinoma. Additionally, patients with high FGF11 expression had a less favorable prognosis than those with low FGF11 expression. The correlation between FGF11 and immune cells suggests that it may serve as a likely biomarker and drug target of lung adenocarcinoma.33 It has also been found that patients with human papillomavirus (HPV)-positive oropharyngeal squamous cell carcinoma (OPSCC) who have high levels of FGF11 are more likely to have a poorer survival rate. Therefore, FGF11 could potentially be used as a prognostic biomarker.34
Recently, some studies showed that FGF11 could be induced by hypoxia and promoted cancer growth and metastasis.35,36 In non-small cell lung cancer (NSCLC), the oncogenic role of FGF11 depends on the up-regulation of HIF-1α.31 Chen et al created a hypoxia model for thyroid cancer cells and observed that FGF11 inhibited the degradation of HIF-1α, thereby increasing its stability. This led to the formation of a positive feedback loop between FGF11 and HIF-1α that facilitated the growth and metastasis of thyroid cancer in hypoxic environments.19 Similarly, our results showed that FGF11 was regulated by HIF-1α and overexpressed FGF11 promoted cell proliferation, migration, and invasion of OCCC cells in vitro. Furthermore, apoptosis assays further demonstrated that downregulation of FGF11 could induce cell apoptosis therefore reversing DDP resistance in OCCC (Figure 5).
 |
Figure 5 The function of the HIF-1α/FGF11 signaling axis in the regulation of proliferation, migration, and DDP resistance in OCCC cells. |
Conclusions
In our study, we indicated that FGF11 is up-regulated in platinum-resistant OCCC tissues, and patients with overexpressed FGF11 had a poor prognosis. FGF11 could enhance cell proliferation, migration, invasion, and DDP resistance in OCCC. Targeting the HIF-1α/FGF11 signaling axis may improve the effectiveness of DDP in OCCC, and more research is warranted.
Ethics Approval
The study was ethically approved by the institutional review board of Peking Union Medical College Hospital (I-22PG84).
Acknowledgments
We thank all the participants for their contribution and participation.
Funding
This study was financially supported by National Key Research and Development Program of China (2022YFC2704202).
Disclosure
All authors declare no conflict of interest.
References
1. Iida Y, Okamoto A, Hollis RL, et al. Clear cell carcinoma of the ovary: a clinical and molecular perspective. Int J Gynecol Cancer. 2021;31(4):605–616. doi:10.1136/ijgc-2020-001656
2. Pavlidis N, Rassy E, Vermorken JB, et al. The outcome of patients with serous papillary peritoneal cancer, fallopian tube cancer, and epithelial ovarian cancer by treatment eras: 27 years data from the SEER registry. Cancer Epidemiol. 2021;75(102045):102045. doi:10.1016/j.canep.2021.102045
3. Ogasawara A, Sato S, Hasegawa K. Current and future strategies for treatment of ovarian clear cell carcinoma. J Obstet Gynaecol Res. 2020;46(9):1678–1689. doi:10.1111/jog.14350
4. Boussios S, Rassy E, Moschetta M, et al. BRCA mutations in ovarian and prostate cancer: bench to bedside. Cancers. 2022;14(16):3888. doi:10.3390/cancers14163888
5. Revythis A, Limbu A, Mikropoulos C, et al. Recent insights into PARP and immuno-checkpoint inhibitors in epithelial ovarian cancer. Int J Environ Res Public Health. 2022;19(14):8577. doi:10.3390/ijerph19148577
6. Matsuzaki S, Yoshino K, Ueda Y, et al. Potential targets for ovarian clear cell carcinoma: a review of updates and future perspectives. Cancer Cell Int. 2015;15(117). doi:10.1186/s12935-015-0267-0
7. Buechel M, Herzog TJ, Westin SN, et al. Treatment of patients with recurrent epithelial ovarian cancer for whom platinum is still an option. Ann Oncol. 2019;30(5):721–732. doi:10.1093/annonc/mdz104
8. Kim A, Serada S, Enomoto T, et al. Targeting annexin A4 to counteract chemoresistance in clear cell carcinoma of the ovary. Expert Opin Ther Targets. 2010;14(9):963–971. doi:10.1517/14728222.2010.511180
9. Kitamura S, Yamaguchi K, Murakami R, et al. PDK2 leads to cisplatin resistance through suppression of mitochondrial function in ovarian clear cell carcinoma. Cancer Sci. 2021;112(11):4627–4640. doi:10.1111/cas.15125
10. Itamochi H, Kigawa J, Sugiyama T, et al. Low proliferation activity may be associated with chemoresistance in clear cell carcinoma of the ovary. Obstet Gynecol. 2002;100(2):281–287. doi:10.1016/s0029-7844(02)02040-9
11. Ghose A, Gullapalli N, Chohan N, et al. Applications of proteomics in ovarian cancer: dawn of a new era. Proteomes. 2022;10(2):16. doi:10.3390/proteomes10020016
12. Yin M, Lu C, Zhou H, et al. Differential molecular pathway expression according to chemotherapeutic response in ovarian clear cell carcinoma. BMC Womens Health. 2023;23(1):298. doi:10.1186/s12905-023-02420-1
13. Zheng M, Piao XM, Byun YJ, et al. Study on the use of Nanostring nCounter to analyze RNA extracted from formalin-fixed-paraffin-embedded and fresh frozen bladder cancer tissues. Cancer Genet. 2022;2022:268–269.
14. Lyu C, Zhang Y, Zhou X, et al. ARID1A gene silencing reduces the sensitivity of ovarian clear cell carcinoma to cisplatin. Exp Ther Med. 2016;12(6):4067–4071. doi:10.3892/etm.2016.3863
15. Devlin J, Miller R, Laforets F, et al. The tumor microenvironment of clear-cell ovarian canCer. Cancer Immunol Res. 2022;10(11):1326–1339. doi:10.1158/2326-6066.CIR-22-0407
16. Ioannidou E, Moschetta M, Shah S, et al. Angiogenesis and anti-angiogenic treatment in prostate cancer: mechanisms of action and molecular targets. Int J Mol Sci. 2021;22(18):9926. doi:10.3390/ijms22189926
17. Wang X, Du W, Xu M, et al. HIF-1alpha is a rational target for future ovarian cancer therapies. Front Oncol. 2021;11(785111). doi:10.3389/fonc.2021.785111
18. Lee S, Garner I, Welch R, et al. Over-expression of hypoxia-inducible factor 1 alpha in ovarian clear cell carcinoma. Gynecol Oncol. 2007;106(2):311–317. doi:10.1016/j.ygyno.2007.03.041
19. Chen B, Feng M, Yao Z, et al. Hypoxia promotes thyroid cancer progression through HIF1alpha/FGF11 feedback loop. Exp Cell Res. 2022;416(1):113159. doi:10.1016/j.yexcr.2022.113159
20. Stewart J, Cunningham N, Banerjee S. New therapies for clear cell ovarian carcinoma. Int J Gynecol Cancer. 2023;33(3):385–393. doi:10.1136/ijgc-2022-003704
21. Khalaf K, Hana D, Chou T, et al. Aspects of the tumor microenvironment involved in immune resistance and drug resistance. Front Immunol. 2021;12(656364). doi:10.3389/fimmu.2021.656364
22. Jing X, Yang F, Shao C, et al. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol Cancer. 2019;18(1):157. doi:10.1186/s12943-019-1089-9
23. Jin Y, Wang H, Liang X, et al. Pathological and prognostic significance of hypoxia-inducible factor 1alpha expression in epithelial ovarian cancer: a meta-analysis. Tumour Biol. 2014;35(8):8149–8159. doi:10.1007/s13277-014-2059-x
24. Klemba A, Bodnar L, Was H, et al. Hypoxia-mediated decrease of ovarian cancer cells reaction to treatment: significance for chemo- and immunotherapies. Int J Mol Sci. 2020;21(24):9492. doi:10.3390/ijms21249492
25. Kato M, Yamamoto S, Takano M, et al. Aberrant expression of the mammalian target of rapamycin, hypoxia-inducible factor-1alpha, and glucose transporter 1 in the development of ovarian clear-cell adenocarcinoma. Int J Gynecol Pathol. 2012;31(3):254–263. doi:10.1097/PGP.0b013e318237d66c
26. Hui Q, Jin Z, Li X, et al. FGF family: from drug development to clinical application. Int J Mol Sci. 2018;19(7):1875. doi:10.3390/ijms19071875
27. Mahapatra S, Jonniya A, Koirala S, et al. The FGF/FGFR signalling mediated anti-cancer drug resistance and therapeutic intervention. J Biomol Struct Dyn. 2023;2023:1–25.
28. Ghedini C, Ronca R, Presta M, et al. Future applications of FGF/FGFR inhibitors in cancer. Expert Rev Anticancer Ther. 2018;18(9):861–872. doi:10.1080/14737140.2018.1491795
29. Ye B, Zhang H, Cai T, et al. Exosomal miR-24-3p impedes T-cell function by targeting FGF11 and serves as a potential prognostic biomarker for nasopharyngeal carcinoma. J Pathol. 2016;240(3):329–340. doi:10.1002/path.4781
30. Hu S, Li L, Yeh S, et al. Infiltrating T cells promote prostate cancer metastasis via modulation of FGF11-->miRNA-541-->androgen receptor (AR)-->MMP9 signaling. Mol Oncol. 2015;9(1):44–57. doi:10.1016/j.molonc.2014.07.013
31. Wu X, Li M, Li Y, et al. Fibroblast growth factor 11 (FGF11) promotes non-small cell lung cancer (NSCLC) progression by regulating hypoxia signaling pathway. J Transl Med. 2021;19(1):353. doi:10.1186/s12967-021-03018-7
32. Weaver A, Bossaer JB. Fibroblast growth factor receptor (FGFR) inhibitors: a review of a novel therapeutic class. J Oncol Pharm Pract. 2021;27(3):702–710. doi:10.1177/1078155220983425
33. Zhai B, Wu J, Li T. Fibroblast growth factor 11 enables tumor cell immune escape by promoting T cell exhaustion and predicts poor prognosis in patients with lung adenocarcinoma. J Oncol. 2023;2023(9303632):1–13. doi:10.1155/2023/9303632
34. Flon H, Haeggblom L, Holzhauser S, et al. High levels of FGF11 correlate with poor survival in patients with Human Papillomavirus (HPV)-positive oropharyngeal squamous cell carcinoma. Cancers. 2023;15(7):1954. doi:10.3390/cancers15071954
35. Yang J, Kim WJ, Jun HO, et al. Hypoxia-induced fibroblast growth factor 11 stimulates capillary-like endothelial tube formation. Oncol Rep. 2015;34(5):2745–2751. doi:10.3892/or.2015.4223
36. Lee W, Yim S, Shin J, et al. FGF11 induced by hypoxia interacts with HIF-1alpha and enhances its stability. FEBS Lett. 2017;591(2):348–357. doi:10.1002/1873-3468.12547
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.