")
Back to Journals » Journal of Inflammation Research » Volume 17
Ezrin Contributes to the Damage of Airway Epithelial Barrier Related to Diabetes Mellitus
Authors Yu H , Luo C, Linghu R, Yang J, Wu H
Received 12 November 2023
Accepted for publication 18 April 2024
Published 26 April 2024 Volume 2024:17 Pages 2609—2621
DOI https://doi.org/10.2147/JIR.S449487
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Hongmei Yu,1 Cheng Luo,1 Ru Linghu,2 Juan Yang,3 Haiqiao Wu3
1Department of Geriatrics, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 2Department of Internal Medicine, Hospital of Chongqing University, Chongqing, People’s Republic of China; 3Department of Respiratory Medicine, Chongqing Hospital of Traditional Chinese Medicine, Chongqing, People’s Republic of China
Correspondence: Hongmei Yu, Department of Geriatrics, The First Affiliated Hospital of Chongqing Medical University, No. 1, Youyi Road, Yuzhong District, Chongqing, 400016, People’s Republic of China, Tel +8618996009679, Email [email protected]
Background: Diabetes mellitus predisposes individuals to respiratory infections. The airway epithelial barrier provides defense against inhaled antigens and pathogens. Ezrin, is a component of the membrane-cytoskeleton that maintains the cellular morphology, intercellular adhesion, and barrier function of epithelial cells. This study aimed to explore the role of ezrin in airway epithelial barrier damage and correlate its expression and activation with diabetes mellitus.
Methods: This study was performed in a murine model of diabetes mellitus and with human bronchial epithelial BEAS-2B cells using real-time PCR, Western blotting, immunohistochemical and immunofluorescence staining. Ezrin was knocked down in BEAS-2B cells using siRNA. Ezrin phosphorylation levels were measured to determine activation status. The integrity of the airway epithelial barrier was assessed in vivo by characterizing morphological structure, and in vitro in BEAS-2B cells by measuring tight junction protein expression, transepithelial electrical resistance (TER) and permeability.
Results: We demonstrated that ezrin expression levels were lower in the lung tissue and airway epithelium of diabetic mice than those in control mice. The morphological structure of the airway epithelium was altered in diabetic mice. High glucose levels downregulated the expression and distribution of ezrin and connexin 43, reduced the expression of tight junction proteins, and altered the epithelial barrier characteristics of BEAS-2B cells. Ezrin knockdown had effects similar to those of high glucose levels. Moreover, a specific inhibitor of ezrin Thr567 phosphorylation (NSC305787) inhibited epithelial barrier formation.
Conclusion: These results demonstrate that ezrin expression and activation are associated with airway epithelial damage in diabetes mellitus. These findings provide new insights into the molecular pathogenesis of pulmonary infections in diabetes mellitus and may lead to novel therapeutic interventions for airway epithelial barrier damage.
Keywords: morphological structure, bronchial epithelial cells, high glucose, tight junction, gap junction
Introduction
Diabetes mellitus has become a global health problem due to associated complications over the past several decades.1,2 The International Diabetes Federation estimates that the number of diabetic patients worldwide will increase to 592 million by 2035. Diabetes mellitus predisposes the individual to respiratory infections. Hyperglycemia is an important risk factor for respiratory infections including COVID-193 and is associated with increased morbidity and mortality in patients with inflammatory respiratory disease.4–7 Therefore, understanding the mechanisms underlying respiratory infections caused by hyperglycemia is crucial in patients with diabetes mellitus. The airway epithelial barrier provides resistance to inhaled antigens and pathogens.8 Disruption of epithelial integrity is an important pathological characteristic of chronic airway inflammatory diseases.9 Our previous study reported that hyperglycemia induces dysfunction of the airway epithelial barrier in human airway epithelial cells.10 However, the mechanism by which high glucose concentration induces airway epithelial barrier dysfunction and contributes to respiratory infection is poorly understood.
The epithelial integrity that protects multicellular organisms from the external environment is maintained by intercellular junctional complexes and the orderly arrangement of the cytoskeleton.11,12 The ezrin–radixin–moesin (ERM) family can act as A-kinase anchoring proteins (AKAPs),13–15 building and maintaining the epithelial barrier by connecting transmembrane proteins to the actin cytoskeleton.16 Ezrin (AKAP78), a constituent of microvilli in regions containing densely packed actin filaments, regulates tissue architecture by influencing actin assembly.17 Ezrin has been implicated in microvillus formation, epithelial cell structure, and polarity.18 Moreover, ezrin plays a critical role in tubulogenesis and the migration of kidney epithelial cells during diabetes.19 In addition, ezrin expression is decreased in diabetic kidney glomeruli and controls actin reorganization and glucose uptake via the transporter GLUT1 in cultured podocytes.20
To better understand the pathogenesis of respiratory infections in diabetes mellitus, we hypothesized that alterations in ezrin are associated with breaches in the airway epithelium and that ezrin could be a potential biomarker for patients with respiratory infections and diabetes mellitus.
Materials and Methods
Animals
Briefly, db/db (C57BL/6J-Leprdb/Leprdb) (diabetic group) and age-matched non-diabetic db/+ (C57BL/6J-Leprdb/+) (control group) mice were purchased from the Model Animal Research Center of Nanjing University. All mice were housed in a specific pathogen-free facility with free access to food and water. Body weight and blood glucose levels were monitored weekly using an electronic scale and a glucometer, respectively. Mice were sacrificed at 18 weeks of age and the lungs were recovered. All experimental procedures were performed under the guidelines of the institutional Animal Care and Use Committee at Chongqing Medical University. All animal procedures were also approved by the Research Ethics Committee of Chongqing Medical University.
Lung Tissue Pathology
Right lungs were stored at –80 °C and used for the extraction of RNA and protein, while the left lungs were placed in 10% formalin and used for hematoxylin-eosin (HE) staining, periodic acid-Schiff (PAS) staining, and immunohistochemistry. Paraformaldehyde was carefully rinsed from the left lung with phosphate buffer saline (PBS). Specimens were subjected to gradient dehydration and embedded in paraffin. Wax block embedding, HE and PAS staining were performed using standard methods.
Cell Culture
Immortalized human bronchial epithelial cells, BEAS-2B (ATCC [CRL-9609]), were maintained in bronchial epithelial cell growth medium (BEGM) with SingleQuot kit additives (Product # CC-3170, Lonza, Walkersville, MD, USA). Cells were cultured in serum-free BEGM, as previously described21 and treated with glucose (5 mM normal glucose [NG], ie physiological concentration,22 or 30 mM high glucose [HG]), or an osmotic control (5 mM glucose with 30 mM final concentration mannitol). Cells were treated with 3 μM NSC305787 (specific inhibitor of ezrin Thr567 phosphorylation, dissolved in dimethyl sulfoxide [DMSO]) or vehicle control (DMSO) for 6 h at 37 °C.23 BEAS-2B cells were cultured under normal or high glucose conditions for 72 h.
Immunohistochemistry
Formalin-fixed and paraffin-embedded lung sections were deparaffinized and endogenous peroxidases were inactivated with 3% H2O2. Then, sections were blocked with 3% BSA for 1 h and incubated with anti-ezrin primary antibody (Cell Signaling, 3145) at 4 °C overnight. The next day, the sections were washed thrice with PBS for 5 min each, incubated with secondary antibody for 1 h at room temperature, and developed using a DAB kit (Abcam). For quantitative analysis, the areas of ezrin staining was quantified using ImageJ software. Immunohistochemical staining was semi-quantitatively analyzed using the immunoreactive scoring system. The percentage of positive cells was graded on a scale of 0–4: (0: negative, 1:0–25%, 2:26–50%, 3:51–75%, 4:76–100%). The signal intensity was scored on a scale of 0–3: 0=negative; 1=weak; 2=moderate; and 3=strong. Thus, the final immunoreactive score = (score of staining intensity) × (score of percentage of positive cells).
Small Interfering RNA Preparation and Transfection
One day before transfection, BEAS-2B cells were plated in growth medium without antibiotics so that they were 50–80% confluent at the time of transfection. The cells were transfected with 0.5 μg/mL siRNA duplexes (either scrambled or ezrin) using FuGENE 6 according to the manufacturer’s instructions. The siRNA concentrations were chosen based on dose-response studies (data not shown).
Real-Time Polymerase Chain Reaction Analysis
Total RNA was extracted from BEAS-2B cells or lungs from each experimental group using TRIzol reagent. cDNA was generated using an iScript Complementary DNA Synthesis Kit (Bio-Rad, Hercules, CA, USA). TaqMan qPCR assay reagents for ezrin, ZO-1, and occludin were obtained from Applied Biosystems. To determine relative mRNA expression levels, the comparative cycle of threshold (ΔΔCT) method was performed using the housekeeping gene GAPDH as an internal control.
Western Blotting
Western blotting analysis was performed on samples containing equal amounts of protein (20 µg) using 6% or 10% SDS-PAGE gels. The separated proteins were transferred onto polyvinylidene fluoride (PVDF) membranes. Nonspecific binding sites were blocked with 5% non-fat dry milk in Tris-buffered saline containing 0.1% Tween-20 (TTBS). Membranes were incubated overnight at 4 °C with ezrin (Cell Signaling, 3145), p-ezrin (Santa Cruz Biotechnology, sc-166858), ZO-1 (Cell Signaling, 13663), or occludin (Cell Signaling, 91131) antibodies. After washing with TTBS three times each for 10 min, membranes were incubated with an anti-rabbit secondary antibody conjugated with alkaline phosphatase. Blots were developed using an enhanced chemiluminescence reagent kit (Keygen, Nanjing, China) according to the manufacturer’s instructions.
Immunofluorescence and Confocal Laser Scanning Microscopy
BEAS-2B cells were seeded onto sterile coverslips and plated in 24-well plates. The cells from each group were fixed with 4% paraformaldehyde at 37 °C for 20 min, washed thrice with PBS, and permeabilized with 0.1% Triton X-100 for 15 min. After washing thice with PBS, slides were blocked in 5% BSA for 1 h at room temperature and incubated with anti-ezrin (Cell Signaling, 3145) and anti-Cx43 antibodies (Santa Cruz Biotechnology, sc-271837) overnight at 4 °C. After washing thrice with PBS, the slides were treated with Alexa Fluor 488-labeled goat anti-rabbit IgG and Alexa Fluor 647-labeled goat anti-mouse IgG for 1 h in the dark. The slides were rinsed thrice with PBS, mounted with 50% glycerol, and stored in the dark. Immunofluorescence was examined using a Leica Sp2 confocal microscope.
Transepithelial Electrical Resistance (TER) Measurement
Cells were seeded in the upper chamber of transwell tissue culture plates (12-mm diameter, 0.4-µm pore size, Costar) and allowed to reach confluence. The TER of the cells grown on the filters was measured using an epithelial voltmeter coupled to an A/N converter, monitored using PowerLab software (Chart for Windows, v4.0 AD Instruments) with one acquisition every 0.5 s. The background electrical resistance attributed to the fluid and blank transwell filter was subtracted from the measured TER. The TER measurements were normalized to the area of the monolayer and are expressed in Ω · cm2.
Permeability Assay
Permeability measurements were performed as described previously.24 BEAS-2B cells were seeded at a density of 5×105 cells/cm2 on the apical surface of 12-transwell culture plates, with 0.4 µm polyester filters. Rhodamine B isothiocyanate (RITC)-labeled dextran 70S (Sigma) was added to the cells (50 µL at 100 mM). Over the next 3 h, 50 µL media was sampled from the lower compartments at 30 min intervals and analyzed on a SpectraMax Gemini EM fluorescent plate reader with an excitation wavelength of 530 nm and an emission wavelength of 590 nm. Epithelial permeability was calculated as the percentage of RITC-dextran 70S detected in the bottom chamber over that in the top chamber per unit filter surface area (cm2) per unit time (s).
Statistical Analyses
All statistical analyses were performed using GraphPad Prism 6.0 Software. Values are reported as mean ± SD. Statistical analysis was performed using the Mann–Whitney U-test, one-way analysis of variance, or Student’s t-test. Statistical significance was set at p < 0.05.
Results
Ezrin Expression in Lungs and Airway Epithelium Was Decreased in a Murine Model of Diabetes Mellitus
The lungs of diabetic mice and non-diabetic controls were collected to investigate the effect of diabetes mellitus on ezrin expression. Mice deficient in the leptin receptor (db/db) exhibited significantly higher blood glucose concentrations (Figure 1A) and were heavier (Figure 1B) than mice in the control group. Ezrin mRNA expression in the lungs of the diabetic mice, detected by real-time PCR, was lower than that in the control mice (Figure 1C). Similarly, Western blotting revealed that ezrin protein levels were lower in the diabetic mouse lungs (Figure 1D). Immunohistochemical staining showed decreased ezrin protein in the airway epithelium of diabetic compared with non-diabetic mice (Figure 1E and F).
 |
Figure 1 Ezrin expression is decreased in db/db mouse model of diabetes mellitus. (A) Blood glucose concentration and (B) weight of mice in the control and diabetes groups (n=6 each). Ezrin mRNA and ezrin protein expression in the lungs were detected by (C) real-time PCR and (D) Western blot analysis, respectively (n=6). Ezrin protein expression in airway epithelium of diabetic and control mice was examined by (E and F) immunohistochemical staining (n=6) and Representative images (black arrows indicate ezrin expression in airway epithelium). Staining was analyzed by ImageJ software (scale bar = 50 μm, original magnification, ×200) and results are expressed as fold-change diabetic versus control group. **P<0.01. |
The Morphological Structure of Airway Epithelium Was Altered in the db/db Murine Model of Diabetes Mellitus
To examine the effects of hyperglycemia on the morphological structure of the airway epithelium, we used HE and PAS staining. Hyperglycemia clearly caused thickening of the tracheal wall (Figure 2A). Compared with the control group, goblet cell metaplasia was significantly increased in the airways of the diabetic mice (Figure 2B). These results indicate that hyperglycemia does alter the morphological structure of the airway epithelium.
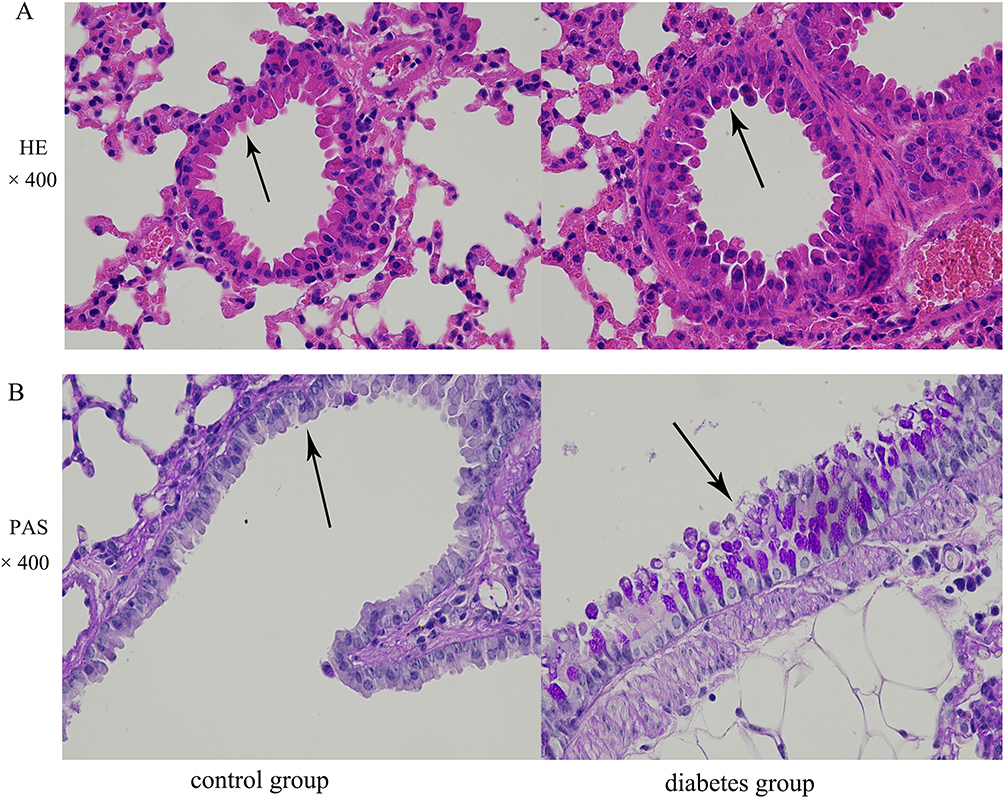 |
Figure 2 The morphological structure of airway epithelium is altered in the db/db murine model of diabetes mellitus. Airway epithelia in lungs of control and diabetic mice were stained with (A) HE (black arrows indicate the tracheal wall, original magnification, ×400) and (B) PAS (black arrows indicate goblet cell metaplasia in the airway epithelium, original magnification, ×400). |
Down-Regulation of Ezrin by High Glucose Concentration Decreased the Expression of Tight Junction Proteins in Human Bronchial Epithelial Cells
To determine the relationship between high glucose and ezrin expression, cells were treated with 30 mM glucose compared with physiological glucose (5 mM). The expression of ezrin mRNA and protein was detected by Western blotting (Figure 3A) and real-time PCR (Figure 3B), respectively. Treatment with high glucose decreased both ezrin mRNA and protein expression in human bronchial epithelial cells. Ezrin siRNA had inhibitory effects on ezrin expression similar to those observed with high glucose.
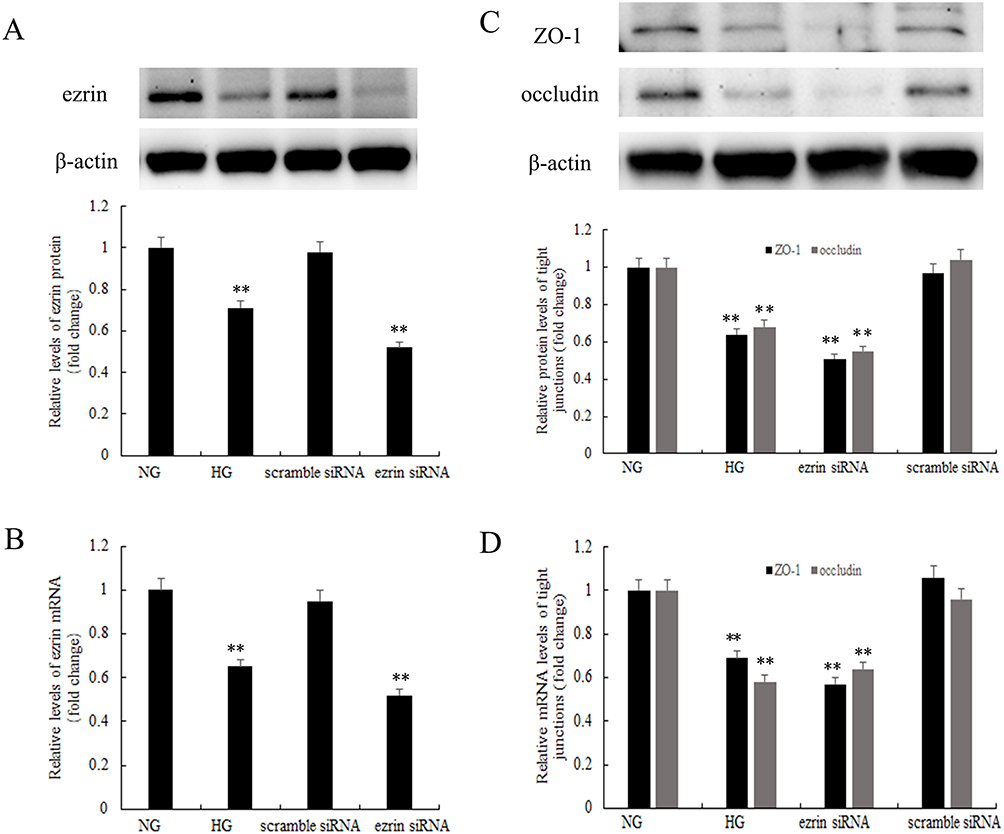 |
Figure 3 Downregulation of ezrin by high glucose levels decreases tight junction proteins in human bronchial epithelial cells. (A and B) Ezrin mRNA and protein expression was examined by Western blotting and real-time PCR, respectively. (C and D) The expression of ZO-1 and occludin was detected by Western blotting and real-time PCR. BEAS-2B cells were pretreated with 100 mM ezrin siRNA or scrambled siRNA and/or stimulated with normal glucose (NG, 5 mM) or high glucose (HG, 30 mM). The HG results were expressed as fold-change over the NG group. **P<0.01. Figure is representative of n=3. |
To investigate whether tight junction proteins were regulated by high glucose concentrations, ZO-1 and occludin protein and mRNA levels were detected by Western blotting (Figure 3C) and real-time PCR (Figure 3D) respectively. High glucose levels led to a significant decrease in the expression of ZO-1 and occludin compared to the normal glucose group. To determine whether ezrin downregulation affects ZO-1 and occludin expression, BEAS-2B cells were transfected with ezrin siRNA or scrambled siRNA. Cells transfected with ezrin siRNA showed a significant decrease in ezrin gene expression and protein production, whereas transfection with scrambled siRNA did not affect ezrin levels. As in the high glucose-treated cells where ezrin was decreased, the expression of ZO-1 and occludin decreased in cells transfected with ezrin siRNA. Transfection with scrambled siRNA had no inhibitory effect on ZO-1 or occludin expression in BEAS-2B cells.
Ezrin Regulated the Distribution of Cx43 Under High Glucose in BEAS-2B Cells
To examine the presence and localization of ezrin and Cx43 in airway epithelial cells, BEAS-2B bronchial epithelial cells were stained for ezrin and Cx43 and immunofluorescence was examined by confocal microscopy (Figure 4). Immunostaining showed that ezrin was abundantly expressed and colocalized with Cx43 in this cell line. A high glucose concentration or ezrin silencing decrease Cx43 expression and colocalization in these bronchial epithelial cells.
 |
Figure 4 Distribution of ezrin and Cx43 under high glucose conditions in BEAS-2B cells. (A–C) The distribution of ezrin and Cx43 in cells was observed by immunofluorescence (original magnification, ×600). BEAS-2B cells were pretreated with 100 mM ezrin siRNA or scrambled siRNA and/or stimulated with normal glucose ([NG], 5 mM) or high glucose (HG, 30 mM). The results are expressed as fold-change versus the NG group. **P<0.01. This figure is representative of n=3. |
A High Glucose Concentration Inhibited Ezrin Phosphorylation in Human Bronchial Epithelial Cells
To explore the effects of high glucose levels on ezrin phosphorylation, a specific inhibitor of ezrin Thr567 phosphorylation (NSC305787) was used. The protein levels of p-ezrin (Figure 5A) and ezrin (Figure 5B) were measured by Western blotting. High glucose significantly decreased levels of p-ezrin and ezrin levels. The amount of p-ezrin was decreased by treatment of the cells with NSC305787, similar to high glucose treatment, indicating that phosphorylation of ezrin at Thr567 was decreased. The overall level of ezrin was not affected by NSC305787 or DMSO.
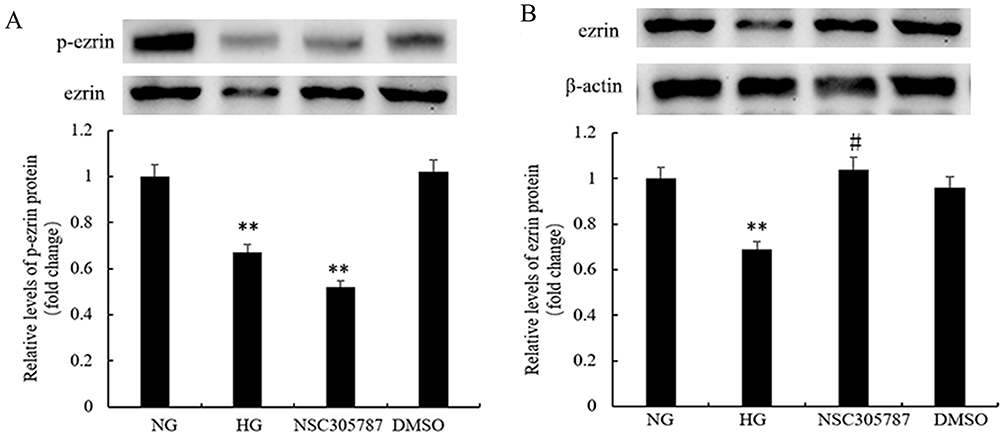 |
Figure 5 High glucose levels inhibit ezrin phosphorylation in human bronchial epithelial cells. The expression levels of p-ezrin and ezrin were detected using (A and B) Western blot analysis. BEAS-2B cells were treated with 3 μM of NSC305787 or vehicle control (DMSO) for 6 h at 37 °C and/or stimulated with normal glucose (NG, 5 mM), or high glucose (HG, 30 mM). The results were expressed as fold-change versus the NG group. **P<0.01, #P>0.05. Figure is representative of n=3. |
Airway Epithelial Transepithelial Electrical Resistance and Permeability Were Modulated by Down-Regulation of Ezrin Induced by High Glucose in Human Bronchial Epithelial Cells
To determine whether high glucose and ezrin downregulation alter cell barrier characteristics, functional integrity, and permeability were evaluated using TER (Figure 6A) and permeability assays (Figure 6B), respectively. BEAS-2B cells grown in high glucose showed decreased cell monolayer TER and increased cell permeability compared to cells grown in physiological concentrations of glucose. Cells transfected with ezrin siRNA showed reduced ezrin expression, significantly decreased cell monolayer TER, and increased cell permeability compared with those transfected with scrambled siRNA. Transfection with scrambled siRNA had no inhibitory effect on cell monolayer TER or permeability of BEAS-2B cells.
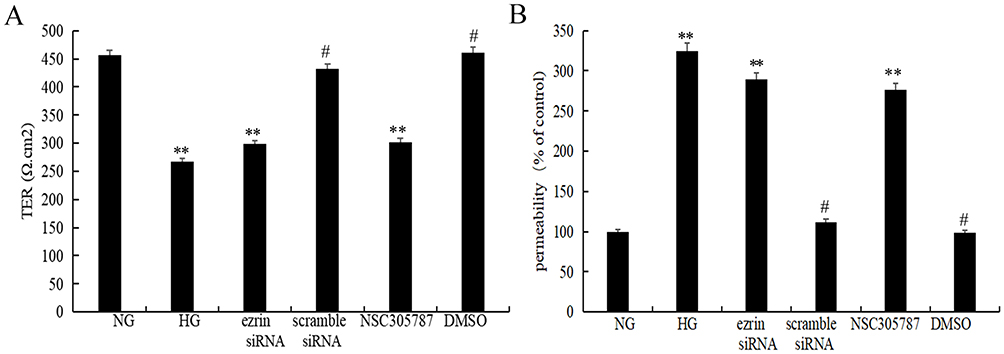 |
Figure 6 TER and permeability are regulated by decreased ezrin levels in human bronchial epithelial cells. Functional integrity and permeability of a BEAS-2B cell layer was evaluated by measuring (A) TER and (B) permeability. BEAS-2B cells were treated with 3 μM NSC305787 or vehicle control (DMSO) for 6 h at 37 °C and/or stimulated with normal glucose (NG, 5 mM), or high glucose (HG, 30 mM). The results were expressed as fold-change versus the NG group (5 mM). **P<0.01, #P<0.01. Figure is representative of n=3. |
Discussion
We demonstrated that ezrin expression was reduced in the lung tissue and airway epithelia of diabetic mice and the morphological structure of the airway epithelium was altered compared to that in control mice. These results indicate that hyperglycemia may be responsible for these changes and that ezrin, a crosslinker protein between membrane proteins and actin filaments, may play a critical role in this process. To explore the role of ezrin in hyperglycemia-induced airway epithelial damage we used human bronchial epithelial BEAS-2B cells. We demonstrated that high glucose downregulated ezrin levels in these cells. High glucose treatment, or knockdown of ezrin using siRNA had similar effects on these cells that could account for epithelial damage: These conditions decreased the expression of tight junction proteins (ZO-1 and occludin) and altered cell barrier characteristics (TER and permeability), suggesting that ezrin plays an important role in hyperglycemia-induced airway epithelial damage. Moreover, we discovered that a specific inhibitor of ezrin Thr567 phosphorylation (NSC305787) showed detrimental effects on cell barrier functions (TER and permeability), similar to those of high glucose concentrations in human bronchial epithelial cells. These results demonstrate that decreased ezrin levels and the inhibition of ezrin phosphorylation due to hyperglycemia may cause airway epithelial damage in diabetes mellitus.
The respiratory epithelium is the major boundary between the environment and the host. Sophisticated barrier functions, sensing, immune regulatory, and antimicrobial mechanisms are involved in maintaining homeostasis and defending the lungs against extraneous substances and pathogens.25 The essential components that contribute to the function of the airway epithelial barrier include mucociliary escalators,26 intercellular tight and adherens junctions that regulate epithelial paracellular permeability,27 and secreted antimicrobial products that kill inhaled pathogens.28 Therefore, impairment of epithelial paracellular permeability leads to barrier dysfunction in the airway epithelium. Diabetes has often been identified as an independent risk factor for respiratory infection. Previous studies have reported that high glucose levels induce endothelial barrier dysfunction in diabetic retinopathy and in brain.29–31 In the present study, we examined airway epithelial paracellular permeability to explore the effects of hyperglycemia on the barrier function of the airway epithelium. Hyperglycemia damages the integrity and permeability of airway epithelium both in vivo and in vitro. These results indicate that hyperglycemia may damage the airway epithelial barrier, leading to epithelial barrier dysfunction.
Ezrin is considered a key regulator of airway cells that modulates membrane-cortex interactions.32 It was initially identified as a crosslinker between the plasma membrane and cortical cytoskeleton.33 Based on its localization and protein-binding activity, ezrin is involved in regulating a variety of cellular processes, including cell polarity,34 epithelial morphology,17 and cell-cell and cell-substrate adhesion,35,36 all of which are essential for maintaining epithelial barrier integrity. Ezrin may participate in bronchial epithelium repair during the early stages of asthma.32 Ezrin knockdown in mice leads to the suppression of ciliary beating activated by procaterol, without changes in ciliary morphology and basal body orientation in the airway epithelium.37 Previous studies reported that ezrin may play a role in the development of renal complications in diabetes.19,20 The present study showed that high glucose concentrations decrease ezrin expression in diabetic mice (in vivo) and human bronchial epithelial cells (in vitro). These results demonstrate that ezrin, a crosslinker of the plasma membrane and cortical cytoskeleton, may participate in hyperglycemia-induced damage to the airway epithelial barrier.
Ezrin acts as a crosslinker between membrane proteins and actin filaments via scaffold proteins.38 The epithelium is a highly regulated and impermeable barrier maintained exclusively by tight junctions.39 Tight junctions are composed of integral membrane proteins such as claudins, occludin, and junctional adhesion molecules, as well as peripheral membrane proteins, including zonula occludens (ZO), scaffold proteins that contain 3 PDZ domains.11 Ezrin acts as a marker of epithelial damage in asthma, and ZO-1 is decreased in the lungs of mice with allergic asthma.40 Human ZO-1 is used clinically to monitor morphological changes and permeability of the bronchial epithelium in respiratory diseases.41 Tight junctions interact with the cytoskeleton via numerous adaptor proteins and form impermeable mechanical barriers. Loosening of tight junctions between airway epithelial cells increases the transepithelial glucose gradient, hyperglycemia increases the glucose concentration of the airway surface liquid, and the airway defense against infection may be dampened, resulting in lung bacterial overgrowth in diabetes mellitus.42 In the present study, high glucose concentrations decreased ZO-1 and occludin expression in human bronchial epithelial cells. Ezrin knockdown had similar effects on the expression of these tight junction proteins, suggesting that ezrin plays an important role in high glucose-induced epithelial injury.
Gap junctions ensure direct cell-to-cell transfer of short peptides, ions, nucleotides, sugars, and second messengers.43 Controlling gap junction coupling is a necessary for cellular function and the response to internal and external stimuli. Cx43 is the major connexin and the most widely distributed connexin in airway epithelial cells44–46 Cx43-mediated gap junction intercellular communication coordinates a signaling network that modulates airway surface liquid volume in human bronchial epithelial cells.45 Regulation of Cx43 expression has been associated with a wide variety of pathological conditions and diseases, including dysfunction of the diabetic retinal vasculature, diabetic nephropathy, and diabetic skin conditions.46,47 Ezrin is the only ezrin-radixin-moesin (ERM) family member involved in the control of gap junction gating. Ezrin regulates gap junction opening and forms a supramolecular complex with Cx43 which, in an epithelial liver cell line, facilitated the regulation of intercellular communication.48,49 Although previous studies have suggested that ezrin and Cx43 play important roles in the process of diabetic dysfunction, little is known about the regulation of Cx43 by ezrin in the airway epithelium under high-glucose conditions, or how perturbed ezrin-Cx43 interactions lead to dysfunction of the airway epithelial barrier which contributes to respiratory infections. In the present study, ezrin was found to be co-distributed with Cx43 in BEAS-2B cells. We demonstrated a central role of ezrin in Cx43 localization under high glucose conditions in human airway epithelial cells.
The effects of ezrin are mediated by interactions between its N- and C-terminal domains that maintain ezrin in a dormant, inactive state: Binding sites for membrane proteins located in the FERM domain and the binding site for actin filaments located in the C-terminal domain are masked in this closed conformation.50,51 Ezrin is activated by phosphorylation, which disrupts the intramolecular interactions between the N- and C-terminal domains and allows the N-terminal domain to interact with membrane receptor complexes and the C-terminal domain to interact with F-actin.16 Thus, the interaction between ezrin and other proteins is facilitated by the phosphorylation of multiple domains by various kinases.52 Phosphorylated ezrin plays a crucial role in the development of diabetic complications,53 regulation of cystic fibrosis transmembrane regulator (CFTR) in airway epithelial cells,54 and maintenance of tight junction organization and barrier function in cystic fibrosis airway epithelial cells.55 In the present study, the culture of airway epithelial cells in high glucose inhibited ezrin phosphorylation and disrupted cell barrier characteristics (TER and permeability). A specific inhibitor of ezrin Thr567 phosphorylation (NSC305787) showed similar inhibitory effects on ezrin phosphorylation and cell barrier characteristics (TER and permeability). Thus, high glucose levels may damage the airway epithelium by decreasing ezrin expression and inhibiting its activation.
The present study had some limitations. First, the signaling mechanisms involved in airway epithelial barrier damage in diabetes mellitus required further exploration. Second, ideal cell culture conditions should include an air-liquid interface, since this type of culture system is more representative of the airway epithelial barrier that exists in vivo. An air-liquid interface will be used in future research on the airway epithelial barrier.
Conclusion
We have shown that high levels of glucose decrease ezrin expression and inhibit ezrin phosphorylation, resulting in changes in tight junction proteins and damage to the airway epithelial barrier in vivo and in vitro. These findings are especially important because hyperglycemia is closely linked to chronic inflammatory airway diseases. Therefore, the regulation of ezrin expression is an ideal goal for reducing pathological effects associated with chronic inflammatory airway diseases related to diabetes mellitus. Our findings expand the current knowledge of the molecular mechanisms underlying the relationship between chronic inflammatory airway disease and diabetes mellitus. This novel recognition of the central role of ezrin regulation is a potential new therapeutic approach for chronic inflammatory airway disease associated with diabetes mellitus.
Acknowledgments
We thank Jing Yu for her assistance with cell culture and Linqiang Ma for his assistance with animal experiments.
Funding
This study was supported by the Scientific and Technological Research Program of the Chongqing Municipal Education Commission (Grant No. KJQN201900428), and Natural Science Foundation of Chongqing (Grant No.CSTB2023NSCQ-MSX0119).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Shaw JE, Sicree RA, Zimmet PZ. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabet Res Clin Pract. 2010;87(1):4–14. doi:10.1016/j.diabres.2009.10.007
2. Yang J, Wu SJ, Dai WT, Li YX, Li YY. The human disease network in terms of dysfunctional regulatory mechanisms. Biol Direct. 2015;10(1):60. doi:10.1186/s13062-015-0088-z
3. Pranata R, Henrina J, Raffaello WM, Lawrensia S, Huang I. Diabetes and COVID-19: the past, the present, and the future. Metabolism. 2021;121:154814. doi:10.1016/j.metabol.2021.154814
4. Baker EH, Janaway CH, Philips BJ, et al. Hyperglycaemia is associated with poor outcomes in patients admitted to hospital with acute exacerbations of chronic obstructive pulmonary disease. Thorax. 2006;61(4):284–289. doi:10.1136/thx.2005.051029
5. Brennan AL, Gyi KM, Wood DM, et al. Airway glucose concentrations and effect on growth of respiratory pathogens in cystic fibrosis. J Cyst Fibros. 2007;6(2):101–109. doi:10.1016/j.jcf.2006.03.009
6. Philips BJ, Redman J, Brennan A, et al. Glucose in bronchial aspirates increases the risk of respiratory MRSA in intubated patients. Thorax. 2005;60(9):761–764. doi:10.1136/thx.2004.035766
7. Tamer A, Karabay O, Ekerbicer H. Staphylococcus aureus nasal carriage and associated factors in type 2 diabetic patients. Jpn J Infect Dis. 2006;59(1):10–14. doi:10.7883/yoken.JJID.2006.10
8. Brune K, Frank J, Schwingshackl A, Finigan J, Sidhaye VK. Pulmonary epithelial barrier function: some new players and mechanisms. Am J Physiol Lung Cell Mol Physiol. 2015;308(8):L731–L745. doi:10.1152/ajplung.00309.2014
9. Winter MC, Shasby SS, Ries DR, Shasby DM. PAR2 activation interrupts E-cadherin adhesion and compromises the airway epithelial barrier: protective effect of beta-agonists. Am J Physiol Lung Cell Mol Physiol. 2006;291(4):L628–L635. doi:10.1152/ajplung.00046.2006
10. Yu H, Yang J, Zhou X, Xiao Q, Lu Y, Xia L. High glucose induces dysfunction of airway epithelial barrier through down-regulation of connexin 43. Exp Cell Res. 2016;342(1):11–19. doi:10.1016/j.yexcr.2016.02.012
11. Schneeberger EE, Lynch RD. The tight junction: a multifunctional complex. Am J Physiol Cell Physiol. 2004;286:C1213–C1228. doi:10.1152/ajpcell.00558.2003
12. Steed E, Balda MS, Matter K. Dynamics and functions of tight junctions. Trends Cell Biol. 2010;20(3):142–149. doi:10.1016/j.tcb.2009.12.002
13. Dransfield DT, Bradford AJ, Smith J, et al. Ezrin is a cyclic AMP-dependent protein kinase anchoring protein. EMBO J. 1997;16(1):35–43. doi:10.1093/emboj/16.1.35
14. Semenova I, Ikeda K, Ivanov P, Rodionov V. The protein kinase A-anchoring protein moesin is bound to pigment granules in melanophores. Traffic. 2009;10(2):153–160. doi:10.1111/j.1600-0854.2008.00852.x
15. Deming PB, Campbell SL, Stone JB, Rivard RL, Mercier AL, Howe AK. Anchoring of protein kinase A by ERM (ezrin-radixin-moesin) proteins is required for proper netrin signaling through DCC (deleted in colorectal cancer). J Biol Chem. 2015;290(9):5783–5796. doi:10.1074/jbc.M114.628644
16. Bretscher A, Reczek D, Berryman M. Ezrin: a protein requiring conformational activation to link microfilaments to the plasma membrane in the assembly of cell surface structures. J Cell Sci. 1997;110(Pt 24):3011–3018. doi:10.1242/jcs.110.24.3011
17. Fehon RG, McClatchey AI, Bretscher A. Organizing the cell cortex: the role of ERM proteins. Nat Rev Mol Cell Biol. 2010;11(4):276–287. doi:10.1038/nrm2866
18. Saotome I, Curto M, McClatchey AI. Ezrin is essential for epithelial organization and villus morphogenesis in the developing intestine. Dev Cell. 2004;6(6):855–864. doi:10.1016/j.devcel.2004.05.007
19. Gallicchio MA, McRobert EA, Tikoo A, Cooper ME, Bach LA. Advanced glycation end products inhibit tubulogenesis and migration of kidney epithelial cells in an ezrin-dependent manner. J Am Soc Nephrol. 2006;17(2):414–421. doi:10.1681/ASN.2005010051
20. Wasik AA, Koskelainen S, Hyvonen ME, et al. Ezrin is down-regulated in diabetic kidney glomeruli and regulates actin reorganization and glucose uptake via GLUT1 in cultured podocytes. Am J Pathol. 2014;184(6):1727–1739. doi:10.1016/j.ajpath.2014.03.002
21. Malm SW, Amouzougan EA, Klimecki WT. Fetal bovine serum induces sustained, but reversible, epithelial-mesenchymal transition in the BEAS-2B cell line. Toxicol In Vitro. 2018;50:383–390. doi:10.1016/j.tiv.2018.04.008
22. Kalsi KK, Baker EH, Fraser O, et al. Glucose homeostasis across human airway epithelial cell monolayers: role of diffusion, transport and metabolism. Pflugers Arch. 2009;457(5):1061–1070. doi:10.1007/s00424-008-0576-4
23. Celik H, Bulut G, Han J, et al. Ezrin inhibition up-regulates stress response gene expression. J Biol Chem. 2016;291(25):13257–13270. doi:10.1074/jbc.M116.718189
24. Vinhas R, Cortes L, Cardoso I, et al. Pollen proteases compromise the airway epithelial barrier through degradation of transmembrane adhesion proteins and lung bioactive peptides. Allergy. 2011;66(8):1088–1098. doi:10.1111/j.1398-9995.2011.02598.x
25. Denney L, Ho L-P. The role of respiratory epithelium in host defence against influenza virus infection. Biomed J. 2018;41(4):218–233. doi:10.1016/j.bj.2018.08.004
26. Knowles MR, Boucher RC. Mucus clearance as a primary innate defense mechanism for mammalian airways. J Clin Invest. 2002;109(5):571–577. doi:10.1172/JCI15217
27. Pohl C, Hermanns MI, Uboldi C, et al. Barrier functions and paracellular integrity in human cell culture models of the proximal respiratory unit. Eur J Pharm Biopharm. 2009;72(2):339–349. doi:10.1016/j.ejpb.2008.07.012
28. Bals R, Hiemstra PS. Innate immunity in the lung: how epithelial cells fight against respiratory pathogens. Eur Respir J. 2004;23(2):327–333. doi:10.1183/09031936.03.00098803
29. Li W, Maloney RE, Aw TY. High glucose, glucose fluctuation and carbonyl stress enhance brain microvascular endothelial barrier dysfunction: implications for diabetic cerebral microvasculature. Redox Biol. 2015;5:80–90. doi:10.1016/j.redox.2015.03.005
30. Chen XL, Zhang XD, Li YY, Chen XM, Tang DR, Ran RJ. Involvement of HMGB1 mediated signalling pathway in diabetic retinopathy: evidence from type 2 diabetic rats and ARPE-19 cells under diabetic condition. Br J Ophthalmol. 2013;97(12):1598–1603. doi:10.1136/bjophthalmol-2013-303736
31. Fernandes R, Girao H, Pereira P. High glucose down-regulates intercellular communication in retinal endothelial cells by enhancing degradation of connexin 43 by a proteasome-dependent mechanism. J Biol Chem. 2004;279(26):27219–27224. doi:10.1074/jbc.M400446200
32. Yin LM, Duan TT, Ulloa L, Yang YQ. Ezrin orchestrates signal transduction in airway cells. Rev Physiol Biochem Pharmacol. 2018;174:1–23. doi:10.1007/112_2017_4
33. Bretscher A. Purification of an 80,000-Dalton protein that is a component of the isolated microvillus cytoskeleton, and its localization in nonmuscle cells. J Cell Biol. 1983;97(2):425–432. doi:10.1083/jcb.97.2.425
34. Bretscher A, Edwards K, Fehon RG. ERM proteins and merlin: integrators at the cell cortex. Nat Rev Mol Cell Biol. 2002;3(8):586–599. doi:10.1038/nrm882
35. Hiscox S, Jiang WG. Ezrin regulates cell-cell and cell-matrix adhesion, a possible role with E-cadherin/beta-catenin. J Cell Sci. 1999;112(Pt 18):3081–3090. doi:10.1242/jcs.112.18.3081
36. Ren L, Hong SH, Cassavaugh J, et al. The actin-cytoskeleton linker protein ezrin is regulated during osteosarcoma metastasis by PKC. Oncogene. 2009;28(6):792–802. doi:10.1038/onc.2008.437
37. Kawaguchi K, Nakayama S, Saito D, et al. Ezrin knockdown reduces procaterol-stimulated ciliary beating without morphological changes in mouse airway cilia. J Cell Sci. 2022;135. doi:10.1242/jcs.259201
38. Kawaguchi K, Asano S. Pathophysiological roles of actin-binding scaffold protein, ezrin. Int J Mol Sci. 2022;23(6):3246. doi:10.3390/ijms23063246
39. Schleimer RP, Kato A, Kern R, Kuperman D, Avila PC. Epithelium: at the interface of innate and adaptive immune responses. J Allergy Clin Immunol. 2007;120(6):1279–1284. doi:10.1016/j.jaci.2007.08.046
40. Jia M, Yan X, Jiang X, et al. Ezrin, a membrane cytoskeleton cross-linker protein, as a marker of epithelial damage in asthma. Am J Respir Crit Care Med. 2019;199(4):496–507. doi:10.1164/rccm.201802-0373OC
41. Evans SM, Blyth DI, Wong T, Sanjar S, West MR. Decreased distribution of lung epithelial junction proteins after intratracheal antigen or lipopolysaccharide challenge: correlation with neutrophil influx and levels of BALF sE-cadherin. Am J Respir Cell Mol Biol. 2002;27(4):446–454. doi:10.1165/rcmb.4776
42. Baker EH, Baines DL. Airway glucose homeostasis: a new target in the prevention and treatment of pulmonary infection. Chest. 2018;153(2):507–514. doi:10.1016/j.chest.2017.05.031
43. Laird DW. The gap junction proteome and its relationship to disease. Trends Cell Biol. 2010;20(2):92–101. doi:10.1016/j.tcb.2009.11.001
44. Losa D, Köhler T, Bellec J, et al. Pseudomonas aeruginosa-induced apoptosis in airway epithelial cells is mediated by gap junctional communication in a JNK-dependent manner. J Immunol. 2014;192(10):4804–4812. doi:10.4049/jimmunol.1301294
45. Scheckenbach KE, Losa D, Dudez T, et al. Prostaglandin E₂regulation of cystic fibrosis transmembrane conductance regulator activity and airway surface liquid volume requires gap junctional communication. Am J Respir Cell Mol Biol. 2011;44(1):74–82. doi:10.1165/rcmb.2009-0361OC
46. Molina SA, Stauffer B, Moriarty HK, Kim AH, McCarty NA, Koval M. Junctional abnormalities in human airway epithelial cells expressing F508del CFTR. Am J Physiol Lung Cell Mol Physiol. 2015;309(5):L475–L487. doi:10.1152/ajplung.00060.2015
47. Wang CM, Lincoln J, Cook JE, Becker DL. Abnormal connexin expression underlies delayed wound healing in diabetic skin. Diabetes. 2007;56(11):2809–2817. doi:10.2337/db07-0613
48. Sawai K, Mukoyama M, Mori K, et al. Redistribution of connexin43 expression in glomerular podocytes predicts poor renal prognosis in patients with type 2 diabetes and overt nephropathy. Nephrol Dial Transplant. 2006;21(9):2472–2477. doi:10.1093/ndt/gfl260
49. Kawaguchi K, Yoshida S, Hatano R, Asano S. Pathophysiological roles of Ezrin/Radixin/Moesin proteins. Biol Pharm Bull. 2017;40(4):381–390. doi:10.1248/bpb.b16-01011
50. Andreoli C, Martin M, Le Borgne R, Reggio H, Mangeat P. Ezrin has properties to self-associate at the plasma membrane. J Cell Sci. 1994;107(Pt9):2509–2521. doi:10.1242/jcs.107.9.2509
51. McRobert EA, Gallicchio M, Jerums G, Cooper ME, Bach LA. The amino-terminal domains of the ezrin, radixin, and moesin (ERM) proteins bind advanced glycation end products, an interaction that may play a role in the development of diabetic complications. J Biol Chem. 2003;278(28):25783–25789. doi:10.1074/jbc.M210433200
52. Lopez JP, Turner JR, Philipson LH. Glucose-induced ERM protein activation and translocation regulates insulin secretion. Am J Physiol Endocrinol Metab. 2010;299(5):E772–E785. doi:10.1152/ajpendo.00199.2010
53. Abbattiscianni AC, Favia M, Mancini MT, et al. Correctors of mutant CFTR enhance subcortical cAMP-PKA signaling through modulating ezrin phosphorylation and cytoskeleton organization. J Cell Sci. 2016;129(6):1128–1140. doi:10.1242/jcs.177907
54. Castellani S, Guerra L, Favia M, Di Gioia S, Casavola V, Conese M. NHERF1 and CFTR restore tight junction organisation and function in cystic fibrosis airway epithelial cells: role of ezrin and the RhoA/ROCK pathway. Lab Invest. 2012;92(11):1527–1540. doi:10.1038/labinvest.2012.123
55. Dukic AR, Haugen LH, Pidoux G, Leithe E, Bakke O, Taskén K. A protein kinase A-ezrin complex regulates connexin 43 gap junction communication in liver epithelial cells. Cell Signal. 2017;32:1–11. doi:10.1016/j.cellsig.2017.01.008
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.