")
Back to Journals » International Journal of General Medicine » Volume 16
Expression Profile and Gene Regulation Network of NUSAP1 in Pan Cancers Based on Integrated Bioinformatics Analysis
Authors Zhu X, Wu Y, Liao L, Huang W, Yuan L, Huang J, Zhan Y, Liu L
Received 12 April 2023
Accepted for publication 31 August 2023
Published 18 September 2023 Volume 2023:16 Pages 4235—4248
DOI https://doi.org/10.2147/IJGM.S414270
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Xiaodi Zhu,1,* Yuting Wu,2,* Liwei Liao,1,* Wenqi Huang,1 Lu Yuan,1 Jihong Huang,1 Yongzhong Zhan,1 Laiyu Liu1
1Chronic Airways Diseases Laboratory, Department of Respiratory and Critical Care Medicine, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China; 2Blood Transfusion Department, Ganzhou Hospital-Nanfang Hospital, Southern Medical University, Ganzhou, Jiangxi, 341000 People’s Republic of China
*These authors contributed equally to this work
Correspondence: Laiyu Liu; Yongzhong Zhan, Email [email protected]; [email protected]
Background: Nucleolar and spindle-associated protein 1 (NUSAP1) plays key roles in microtubules and chromosomes in normal cells both structurally and functionally. In malignancies, NUSAP1 is frequently dysregulated and mutated. However, the expression profiles and biological functions of NUSAP1 in tumors remain unclear.
Methods: NUSAP1 expression in BALB/c mice and human normal or tumor tissues was examined using immunohistochemistry. Kaplan–Meier survival analysis was utilized to assess the prognostic significance of NUSAP1 in tumors, and principal component analysis and co-expression analysis were performed to explore the unique roles of NUSAP1. Gene ontology and Kyoto Encyclopedia of Genes and Genomes pathway enrichment analyses were performed with DAVID. The relevance between NUSAP1 and tumor-infiltrating immune cells was investigated using TIMER. A transcriptional regulation network was constructed using data from The Cancer Genome Atlas.
Results: NUSAP1 expression levels in various mice tissues were different. Compared with normal tissues, NUSAP1 was strongly expressed in several human tumor tissues. We believe that NUSAP1 distinctly impacts the prognosis of several cancers and plays various roles in thymoma and testicular germ cell tumors. Further, NUSAP1 expression levels were significantly positively associated with diverse infiltrating levels of immune cells, including B cells, CD4+ and CD8+ T cells, dendritic cells, and macrophages, in thymoma. The expression level of NUSAP1 demonstrated strong relevance with various immune markers in thymoma. Finally, the miR-1236-5p–NUSAP1 and TCF3–NUSAP1 network revealed the tumor-promoting role of NUSAP1 and pertinent underlying mechanisms in human liver hepatocellular carcinoma.
Conclusion: NUSAP1 may be regarded as a therapeutic target or potential prognostic biomarker for various cancer types.
Keywords: nucleolar and spindle-associated protein 1, bioinformatics, the cancer genome atlas, prognostic biomarker, tumor immune microenvironment
Introduction
Cancer poses a fatal threat to human health and life across the world. As per a report published by GLOBOCAN, 18.1 million new cases of cancer were reported in 2018, and 9.6 million patients died due to cancer globally.1 Considering its high incidence and mortality, identifying convenient and effective approaches to improve the diagnosis of cancer and treatment efficacy is pivotal.
Tumor growth and progression is a complex, consecutive process, involving genetic alterations such as oncogene activation, tumor suppressor gene inactivation and mutations, and antiapoptosis.2,3 At present, tumor therapy has shifted from radiotherapy, chemotherapy, and support therapy to biotherapy, which can kill tumor cells without damaging normal cells. Tumor biotherapy, mainly gene therapy, has emerged as a most promising method. Immunotherapeutic strategies are considered promising because immune-related mechanisms play a key role in cancers.4–6 However, with various therapeutics, it is challenging to precisely select most beneficial regimen for different patients. Therefore, there exists an urgent need for understanding as many as oncogenes and identifying new therapeutic biomarkers and targets.
Nucleolar and spindle-associated protein 1 (NUSAP1), a chromatin-binding and microtubule-associated protein, plays an essential role in cytokinesis, spindle assembly, chromosome segregation, microtubule crosslinking, and bundling and attachment to chromosomes.7 NUSAP1 dysregulation has been observed in various tumors, including prostate cancer,8 glioblastoma,9 and colorectal cancer.10 NUSAP1 upregulation has been reported to contribute to the metastasis of cervical cancer by targeting Wnt/β-catenin signaling11 and promote the invasion of astrocytoma by activating the Hedgehog signaling pathway.12 Moreover, NUSAP1 was reported to be an immunogenic antigen in the majority of patients with AML after allogeneic hematopoietic cell transplantation.13 These findings demonstrate that NUSAP1 has diverse important roles in tumor development, however, present studies failed to provide an overview of NUSAP1 among distinct cancer types. Tissue-specific distribution, biological functions, and mechanisms underlying the action of NUSAP1 in tumors remain unclear.
In the current study, we investigated NUSAP1 expression levels in a variety of normal and tumor tissues derived from various organs. Prognostic values of NUSAP1 among pan cancers were discussed. Then, to explain this expression difference, we explored upstream mechanisms regulating NUSAP1 expression. At last, to figure out how NUSAP1 influences prognosis, we analyzed its interacting partners, participating functions and crosstalk with immune microenvironment.
Materials and Methods
Specimen Collection
Our study was authorized by the Review Board of Nanfang Hospital. All paired tumor tissues and adjacent normal tissues were obtained from January 2018 to January 2019 from patients with cancer (8 cases of liver, 7 cases of colon, 8 cases of breast, 6 cases of kidney, 9 cases of stomach, 3 cases of bladder, 11 cases of lung, and 4 cases of esophagus cancer) who underwent a surgical procedure at Nanfang Hospital. None of them received chemotherapy, radiotherapy, or biological immunotherapy or underwent multiple surgeries before. Informed consent was received from every patient. The whole process followed the ethical guidelines of the World Medical Association, Declaration of Helsinki Ethical Principles for Medical Research Involving Human Subjects.
Animals and Tissues
5 weeks BALB/c mice were purchased from The Central Laboratory of Animal Science at Southern Medical University (Guangzhou, China). The animal facility has been accredited by the Institutional Animal Care and Use Committee. The mice were housed in pathogen-free facilities; chow and water were provided ad libitum. The animals were euthanized at 4 weeks of age, and various tissues were subsequently obtained. The process followed the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health.
IHC
Human or mice organs were fixed in 4% paraformaldehyde, dehydrated using gradient alcohol, embedded in paraffin, and cut into 4-µm thick segments. The sections were then incubated at 65°C for 2 h, dewaxed and dehydrated. EDTA buffer was used for antigen recovery at 100°C for 10 min. Subsequently, anti-NUSAP1 monoclonal antibody (Proteintech, 1:100) was added, followed by incubation at 4°C overnight. Next, biotinylated anti-rabbit secondary antibody was added, and the sections were incubated at 37°C for 1 h. Finally, the sections were subjected to color development via diaminobenzidine, counterstained in hematoxylin, mounted with neutral balm, and observed under an optical microscope (Leica, Germany).
Bioinformatics Analysis
The UCSC database (http://genome.ucsc.edu/) was used to analyze the chromosomal localization site of NUSAP1. The gene structure of NUSAP1 was analyzed using the ENSEMBL database (http://asia.ensembl.org/index.html) and was drawn with the Illustrator for Biological Sequences software (IBS, http://ibs.biocuckoo.org/). NUSAP1 protein sequence in mammals was analyzed using GCBI (https://www.gcbi.com.cn). The DNAMAN software (Lynnon BioSoft, USA) was used to compare NUSAP1 protein sequences of human and other mammals. cBioPortal (http://www.cbioportal.org) was used to examine the mutation frequency of NUSAP1 in various human tissues.
TIMER (https://cistrome.shinyapps.io/timer/) was used to assess NUSAP1 expression levels in different tumors and adjacent normal tissues in the TCGA database (https://cancergenome.nih.gov/). The Kaplan–Meier plotter (http://www.kmplot.com/) was employed to predict the overall survival (OS) of patients based on NUSAP1 expression levels in cancers. To elucidate the potential mechanisms underlying the function of NUSAP1 in various tumors, we performed gene coexpression analyses. PCA was performed using ClustVis (http://biit.cs.ut.ee/clustvis/). GeneMANIA, a plugin of Cytoscape v3.7.0 (http://www.cytoscape.org/), was utilized to analyze the network of differentially coexpressed genes, and the MCODE plugin of Cytoscape was used to select the most relevant modules. GO and KEGG pathway enrichment analyses were performed using DAVID (https://david.ncifcrf.gov/summary.jsp). P < 0.05 indicated statistical significance. The correlation between NUSAP1 expression levels and diverse immune infiltrating cells, such as macrophages, dendritic cells, neutrophils, B cells, and CD4+ and CD8+ T cells, was analyzed with TIMER. TIMER was also used to assess the association between NUSAP1 expr ession levels and various gene markers of B cells and CD4+ and CD8+ T cells. GCBI was used to predict TFs of NUSAP1. miRNAs were predicted using the miRWalk (http://zmf.umm.uniheidelberg.de/), TargetScan (http://www.targetscan.org/), miRDB (http://mirdb.org/), and mirDIP (http://ophid.utoronto.ca/mirDIP/index.jsp) databases.
Statistical Analysis
Kaplan–Meier plots were used to construct OS curves, and results are displayed with P values and HR from a Log rank test. Student’s t-test was applied to compare two distinct groups and Pearson correlation method was utilized for correlation analysis of NUSAP1. All our data were analyzed on software GraphPad Prism (version 6.02, San Diego, California, USA).
Results
Structure, Conservation, and Alteration Frequency of NUSAP1 Gene
The gene structure of NUSAP1 showed a 5′-UTR exon, 3′-UTR exon, 10 CDS exons, and 11 introns. It was located at 15q15.1 (Figure 1A). We then analyzed the alteration frequency of NUSAP1 in different human tumors. Uterine cancer showed the highest mutation rate, followed by bladder cancer and cervical squamous cell carcinoma. Adrenocortical carcinoma had the highest fusion rate, while mesothelioma showed the highest deep deletion, suggesting that the alterations of NUSAP1 varied with different cancer types (Figure 1B).
 |
Figure 1 Gene structure, alteration frequency and conservative analysis of NUSAP1. (A) Chromosome location of NUSAP1 (Highlighted in red line) was shown in first panel. Gene structure of NUSAP1 was shown in second panel. Black box represents exons and grey box represents introns. (B) The bar plot presented the alteration frequency of NUSAP1 in different human tumors. Green, blue, purple, red and grey color represent the rate of mutation, involving deep deletion, fusion, amplification and multiple alterations, respectively. (C) Alignments analysis of NUSAP1 protein sequence among human, cattle, chimpanzee, dog, mouse and pig. The degree of conservation of each amino acid residue among these sequences was marked in blue (100% conserved) green (75% conserved) or khaki (50% conserved). |
Further, we compared the protein sequence of NUSAP1 between humans and other mammals to analyze the conservation of NUSAP1 (Figure 1C). It presented the homology between NUSAP1 in humans and in cattle, chimpanzee, dog, mouse, and pig. As evident, NUSAP1 was found to be highly conserved among these mammals.
Biological Expression Levels and Distribution of NUSAP1 in BALB/c Mice Tissues
We first explored biological expression levels of NUSAP1 in 12 diverse types of normal tissue samples obtained from BALB/c mice using IHC. NUSAP1 was positively expressed in the nucleus of cells from the colon, liver, esophagus, thymus, and stomach (Figure 2A). No obvious staining of NUSAP1 was observed in the cytoplasm of all tissues. Further, NUSAP1 expression was strongly positive in the nucleus of cells from the stomach and intestine, followed by those from the thymus and esophagus. Cells from the liver showed weak expression in their nucleus. No expression of NUSAP1 was found in the nucleus of cells from the thyroid, brain, lung, spleen, heart, kidney, and pancreas. These findings indicated that NUSAP1 expression levels in various mice tissues were different.
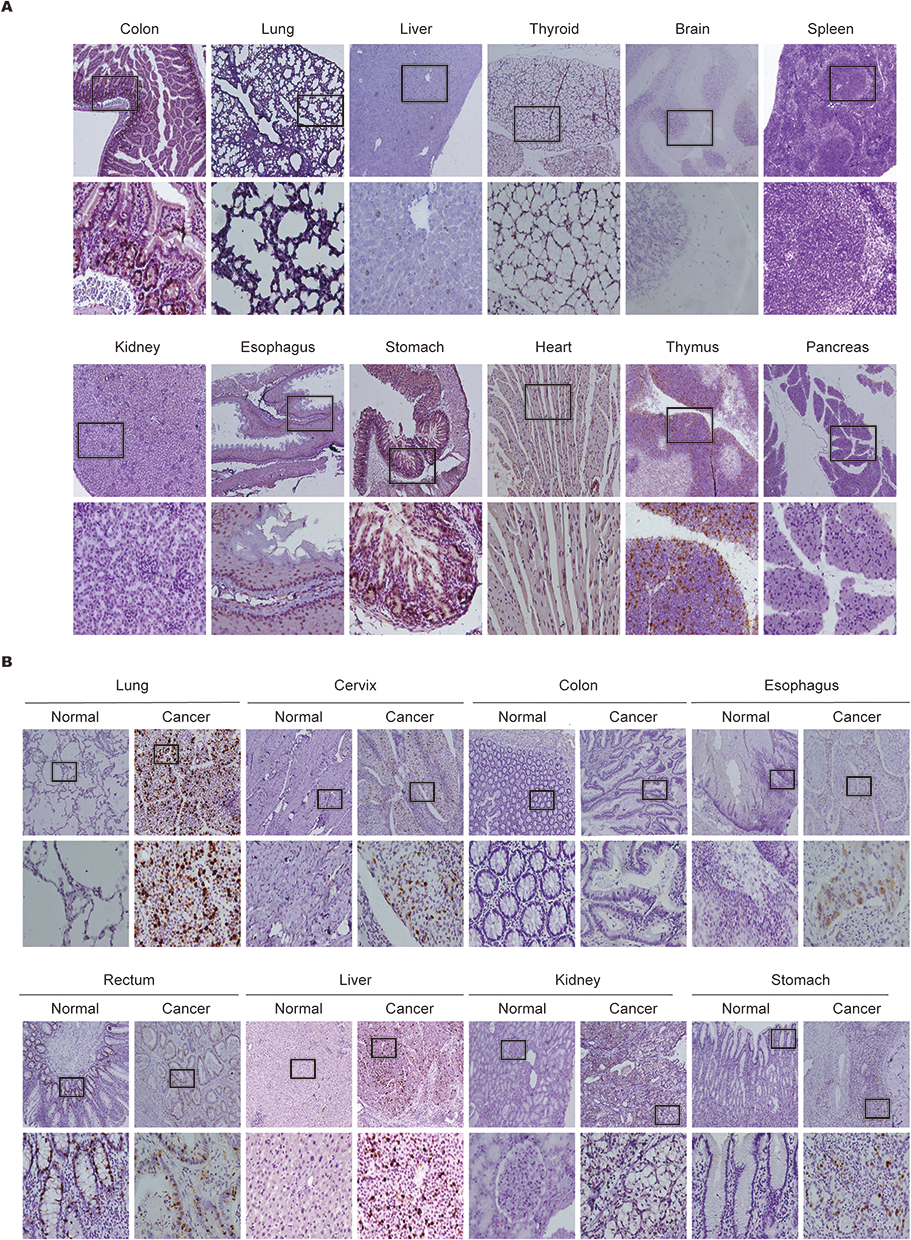 |
Figure 2 Biological distribution and expression of NUSAP1 in tissues of BALB/c mice and human. (A) Expression and distribution of NUSAP1 proteins in BALB/c mice normal issues detected by IHC (100×; 400×; five samples in each group). (B) Expression and distribution of NUSAP1 proteins in paired human tumors and normal tissues examined by IHC assay (100×; 400×; over three samples in each group). |
Expression Levels of NUSAP1 in Human Tumor and Paired Normal Tissues
We assessed the expression level of NUSAP1 in 8 types of human tumor and paired normal tissues via IHC. NUSAP1 was significantly highly expressed in the nucleus of cells from the stomach, liver, esophagus, lung, cervix uteri, and kidney, but negatively expressed in paired normal tissues. In contrast, the protein expression of NUSAP1 was absent in tumor as well as paired normal colon tissues (Figure 2B).
To further examine NUSAP1 expression levels in tumor and normal tissues, we used the RNA-seq data of a variety of tumors in TCGA. Differential expression of NUSAP1 between tumor and normal tissues was shown (sFigure 1). In comparison with corresponding normal tissues, NUSAP1 expression levels were higher in breast invasive carcinoma, colon adenocarcinoma, cholangiocarcinoma, bladder urothelial carcinoma, esophageal cancer, head and neck cancer, lung squamous carcinoma, kidney renal papillary cell carcinoma, liver hepatocellular carcinoma (LIHC), kidney renal clear cell carcinoma, prostate adenocarcinoma, lung adenocarcinoma (LUAD), stomach adenocarcinoma, thyroid carcinoma, rectum adenocarcinoma, uterine corpus endometrial carcinoma. The distribution and expression mRNA levels of NUSAP1 resembled NUSAP1 protein levels in our IHC results.
Impact of NUSAP1 Expression on Prognoses of Different Types of Human Cancers
To further explore the prognostic value of NUSAP1, we used the Kaplan–Meier plotter website to explore the relationship between NUSAP1 expression levels and the OS of patients with cancers. NUSAP1 expression levels were positively correlated with the OS of those with cervical squamous cell carcinoma (Figure 3A), esophageal squamous cell carcinoma (Figure 3C), lung squamous carcinoma (Figure 3E), stomach adenocarcinoma (Figure 3B) and thymoma (Figure 3D). However, in most cases, NUSAP1 was negatively associated with the OS of patients, involving in esophageal adenocarcinoma (Figure 3K), kidney renal papillary cell carcinoma (Figure 3L), kidney renal clear cell carcinoma (Figure 3H), LIHC (Figure 3F), LUAD (Figure 3G), pancreatic ductal adenocarcinoma (Figure 3I) and sarcoma (Figure 3J). Thus, NUSAP1 seems to be closely related to the prognoses of patients with cancer.
 |
Figure 3 Prognostic value of NUSAP1 in the overall survival of Human Cancers. (A–L) Kaplan-Meier curves of overall survival (OS) according to NUSAP1 expression in various tumors, log rank P<0.05 was considered statistically significant. |
Co-Expression Analysis to Explore the Unique Role of NUSAP1 in Solid Cancers
To investigate the biological behavior of NUSAP1 in different cancer types, we performed gene coexpression analyses, which included 29 types of human tumors from TCGA. The correlation coefficient (r value) between NUSAP1 and coexpressed genes was acquired from cBioPortal. To visualize global similarities or differences in NUSAP1 coexpression in cancers, we defined 0.3 ≤ | r | < 0.5 as low correlation, 0.5 ≤ | r | < 0.8 as medium correlation, and | r | ≥ 0.8 as high correlation in order to form a matrix for PCA (Figure 4A). We found that all tumors were distributed in a single cluster, except thymoma (THYM) and testicular germ cell tumor (TGCT). THYM, TGCT, and other tumors were planar in three “clusters”, suggesting that NUSAP1 performs different functions in these “clusters”.
 |
Figure 4 NUSAP1 co-expression patterns in THYM and TGCT are distinct from other tumors. (A) Principal component analysis (PCA) of NUSAP1 co-expression genes in 29 tumor types from TCGA datasets. (B) Principal component analysis (PCA) of NUSAP1 co expression genes in 27 tumor types (THYM and TGCT are excepted) from TCGA datasets. (C) Venn map of the co-expression genes in THYM, LIHC and TGCT. (D–G) Top genes in the most significant module analyzed by MCODE plugin in THYM (D), LIHC (E), TGCT (F) and intersection (G). |
PCA was also utilized to select one representative tumor that still clustered with most other tumors after the analysis to represent “databases that cluster” for further analyses (Figure 4B). LIHC showed the presence of more moderate and highly correlated coexpressed genes (sFigure 2); thus, LIHC was selected as the representative of the cluster for further analyses. Figure 4C shows the number of coexpressed genes in THYM, TGCT, and LIHC. Next, we selected the top 10% genes and all genes in the intersection of the Venn diagram to assess related gene networks via GeneMANIA. Finally, using MCODE, we could identify the top genes that constituted the most significant module (Figure 4D–G). GO and KEGG pathway enrichment analyses were employed to elucidate the biological functions and effects of selected genes, revealing significant enrichment in all types of functional characteristics. In THYM, NUSAP1 was positively associated with immune response, T cell differentiation, and B and T cell receptor signaling pathways (Figure 5A). In TGCT, NUSAP1 was positively correlated with the regulation of transcription, DNA-templated, nucleic acid binding, and metal ion binding (Figure 5B). Whereas in LIHC, higher expression of NUSAP1 was positively correlated with complement activation, oxidation-reduction process, and oxidoreductase activity (Figure 5C). Generally, the top genes selected from the intersection of the Venn diagram were distinctly enriched in pathways correlated with cell division, cell cycle, and DNA replication, representing the main function of NUSAP1 in all selected tumors (Figure 5D). Thus, the biological function of NUSAP1 in some tumors was unique.
 |
Figure 5 Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of co-expressing genes of NUSAP1, consisting with tumor immune infiltration analyses of NUSAP1. (A–D) GO and KEGG pathway enrichment analysis of co-expressing genes of NUSAP1 in THYM (A), TGCT (B), LIHC (C) and in all selected tumors (D). (E) Scatterplots of correlations between NUSAP1 expression and gene markers of B cells, CD8+T cells and CD4+T cells in THYM. (F) NUSAP1 expression has no significant correlation with tumor purity and has significant positive correlations with infiltrating levels of B cell, CD8+T cells, CD4+T cells, macrophages and DC in THYM but negatively correlated with infiltrating levels of neutrophils. (G) NUSAP1 expression is weakly positive related to tumor purity and infiltrating levels of B cell DB+T cells, CD4+T cells, macrophages, neutrophils and DCs in LIHC. (H) NUSAP1 expression has no significant correlations with tumor purity and infiltrating levels of neutrophils and DCs in TGCT. |
Association Between NUSAP1 Expression and Immune Infiltration Levels in Cancers
Enrichment analyses indicated that NUSAP1 was positively associated with T and B cell receptor signaling pathways. Accordingly, we investigated the relationship between NUSAP1 and B cells and CD4+ and CD8+ T cells. Using TIMER, we investigated the correlation between NUSAP1 expression levels and several immune markers of three different immune cells of THYM, TGCT, and LIHC. We found that the NUSAP1 expression levels in THYM were positively associated with most immune markers of the three immune cells (Figure 5E). However, no such association was detected in case of TGCT or LIHC (sFigure 3A and B). These findings were thus consistent with the results of our functional enrichment analyses.
Functional enrichment analyses indicated that NUSAP1 was associated with immune response in THYM. Therefore, we used TIMER to determine whether there existed an association between NUSAP1 expression and immune infiltration levels in different tumors. NUSAP1 expression level was negatively associated with infiltrating levels of neutrophils (r = −0.297, P = 1.34e−3) but positively associated with those of B cells (r = 0.792, P = 9.08e−26), CD4+ T cells (r = 0.747, P = 4.77e−21), CD8+ T cells (r = 0.657, P = 2.14e−15), macrophages (r = 0.547, P = 3.12e−10) and dendritic cells (r = 0.841, P = 1.18e−31) in THYM (Figure 5F). In contrast, no or weaker association was observed between NUSAP1 expression level and infiltrating levels of macrophages, dendritic cells, neutrophils, B cells, CD4+ and CD8+ T cells in LIHC (Figure 5G) as well as TGCT (Figure 5H). In addition, no or weaker association of NUSAP1 expression level with immune infiltration levels was found in other tumors (sfigure 3C–K). These results suggested that NUSAP1 was most likely associated with immune infiltration in THYM.
Construction of Upstream Transcriptional Network Regulating NUSAP1 Expression and Verification in LIHC
On constructing a transcriptional regulatory network of NUSAP1 in LIHC, 117 TFs were found to be involved in the regulation of NUSAP1 expression (Figure 6A). As LIHC represented most tumors according to our gene coexpression analyses, we verified the regulatory network in LIHC, 8 TFs were highly expressed in tumor tissues as compared with paired normal tissues (Figure 6B, P < 0.01). NUSAP1 expression in tumor tissues was higher than that in paired normal tissues (Figure 6C). In comparison with paired normal tissues, TCF3 was significantly highly expressed in tumor tissues (Figure 6D). In addition, Pearson analysis revealed that the correlation between the TF T-cell factor 3 (TCF3) and NUSAP1 expression levels was the most significant in LIHC (Figure 6E, P = 4.30e−16, r = 0.41). Next, using the TargetScan, miRDB, miRWalk and mirDIP databases, we predicted the potential upstream miRNAs targeting NUSAP1. We downloaded 45, 2237, 185, and 263 potential miRNAs targeting NUSAP1 from the miRDB, mirDIP, miRWalk, and TargetScan databases, respectively. Ultimately, hsa-miR-1236-5p was identified to be the only overlapping miRNA in the four databases (Figure 6F), suggesting that hsa-miR-1236-5p is the key regulator in all miRNAs. Based on these results, TCF3 seems to promote and hsa-miR-1236-5p may function either as oncogenes or tumor suppressors by inhibiting NUSAP1 expression in LIHC (Figure 6G), which requires further research for verification.
 |
Figure 6 Construction of transcriptional network of NUSAP1 in LIHC. (A) Prediction of transcription factors (TFs) in NUSAP1 promoter from GCBA online database. (B) Differently expressed TFs between normal and tumour tissues in LIHC derived from TCGA database. (C) Expression of NUSAP1 in LIHC derived from TCGA database. (D)Expression of CTCF in LIHC derived from TCGA database. (E) Pearson analysis of the correlation between the expression of NUSAP1 and TCF3 in LIHC derived from TCGA database. (F) Venn map of the predicted overlapping miRNAs involved in the regulation of NUSAP1. (G) A transcriptional network formed by miR-1236-5P, TCF3 and NUSAP1 in LIHC derived from TCGA database. (***p < 0.001). |
Discussion
NUSAP1 is a chromatin-binding and microtubule-associated protein whose primary physiological function is to stabilize microtubules, maintain spindle integrity, and format spindle networks.7 An increasing number of studies have demonstrated that NUSAP1 dysregulation is correlated with clinical prognosis and tumor progression. In gastric cancer, Ge et al reported that NUSAP1 downregulation suppressed cancer cell proliferation, migration, and invasion by inhibiting the mTORC1 signaling pathway.14 Further, Gordon et al reported that the high expression of NUSAP1 was associated with worse prognosis in prostate cancer.8 In Ovarian cancer cells, Zhang et al found that NUSAP1 gene silencing induces cell apoptosis and inhibited the cell cycle.15 Based on the physiological function of NUSAP1, we believe that NUSAP1 participates in the initiation and development of cancer via cell cycle regulation. Using IHC and bioinformatics, we studied the biological function of NUSAP1, its expression level in tissues, and pertinent underlying mechanisms, which revealed its functions and prognostic importance in different human tumors.
In the present study, we found that NUSAP1 showed a high level of structural conservation in various mammals, which suggests its potential involvement in the development of species. Considering that BALB/c mice are widely used for in vivo studies, we assessed biological expression levels and histological distribution of NUSAP1 in different mice tissues. IHC indicated that NUSAP1 expression levels were upregulated in the colon, liver, esophagus, thymus, and stomach, but no expression was detected in the lung, thyroid, spleen, brain, heart, kidney, and pancreas. Interestingly, we observed that NUSAP1 expression was mainly distributed in the basal cells of tissues. These results suggest the pivotal role of NUSAP1 in spindle microtubule organization and cell proliferation.
Recently, several researches have demonstrated that NUSAP1 could facilitated tumorigenesis and development, such as breast cancer,16 gastric cancer14 and non-small-cell lung cancer.17 Subsequently, using TCGA, we determined the expression level of NUSAP1 in human tumor tissues and adjacent normal tissues. NUSAP1 expression levels were significantly upregulated in 16 tumors, implying that NUSAP1 is closely associated with tumor development. Subsequently, to verify these results, we measured NUSAP1 expression levels in 10 different tumors using IHC; paired normal tissues were used for comparison. Data pertaining to the expression level of NUSAP1 between our study and TCGA showed certain discrepancies, which could be attributed to the fact that data in TCGA reflects global ethnic characteristics. Different biological characteristics may be due to pathological subtypes and ethnic differences, resulting in tumor heterogeneity. Moreover, NUSAP1 expression was found to be associated with the clinical prognosis of patients with different tumors. High expression levels of NUSAP1 resulted in significantly shorter OS in case of patients with kidney renal papillary cell carcinoma, esophageal adenocarcinoma, pancreatic ductal adenocarcinoma, LIHC, kidney renal clear cell carcinoma, LUAD, and sarcoma. On the other hand, patients with lung squamous carcinoma, esophageal squamous cell carcinoma, cervical squamous cell carcinoma, and stomach adenocarcinoma and with high expression levels of NUSAP1 showed longer OS, suggesting that NUSAP1 plays different roles in different tumors and can thus be used as a therapeutic target or prognostic biomarker.
Previous studies have shown that down-regulation of NUSAP1 repressed the expression of cell cycle-regulation related genes in G2/M phase, inhibited the growth of cancer cells.18 Mechanically, NUSAP1 contributes to metastasis of cancer cells by promoting epithelial-mesenchymal transition (EMT) via TGF-β and Wnt/β-catenin signaling.11,16,19 Besides, there was another study showed that lactate upregulated NUSAP1 expression by inhibiting NUSAP1 protein degradation through lysine acetylated (Kla) modification in pancreatic ductal adenocarcinoma (PDAC), the acetyltransferase P300 was identified as a potential histone Kla “Writer” enzyme, the p300 inhibitor which named C646 could significantly reduce the protein expression of NUSAP1.20 Coexpression analysis is one of the many approaches to explore gene function. Tumor-associated genes and gene modules in various cancers can be investigated using differential coexpression networks.21 Such approaches have been used to determine tumor extrinsic and intrinsic biological processes correlated with a particular gene in a specific cancer type.22–25 In this study, we first explored discrepant coexpression networks of NUSAP1 expression levels in several tumors. We identified a coexpression pattern of NUSAP1 in THYM and TGCT that was distinct from that in other tumors. PCA26 revealed global similarities or differences in NUSAP1 coexpression pattern in cancers. GO and KEGG pathway enrichment analyses using DAVID27 helped us elucidate the biological pathways correlated with diverse biomolecular networks. In our study, NUSAP1 was found to be associated with G1/S and G2/m transition of mitotic cell cycle (Figure 5D). Therefore, we hypothesized that NUSAP1 may participate in tumor progression by promoting EMT via transition of mitotic cell cycle. However, it had unique functions in different tumor types.
Immune mechanisms are known to play a vital role in cancer.28–32 Developing robust immunotherapeutic strategies for the treatment of cancers is pivotal. Thymus is a lymphatic organ that is responsible for many immunological functions. THYM is closely related to autoimmune diseases. Herein we observed that NUSAP1 expression level was associated with different immune infiltration levels in tumors, particularly in THYM. Specifically, NUSAP1 expression level was significantly positively correlated with the infiltration levels of B cells, CD4+ and CD8+ T cells, dendritic cells, and macrophages in THYM. Moreover, the relationship between various marker genes of different immune cells and NUSAP1 expression levels indicated the crucial role of NUSAP1 in modulating tumor immune response in THYM. Gene markers of B cells such as CD79A showed moderate correlations with NUSAP1 expression, while CD4+ T cell markers CD4 and CD8+ T cell markers CD8a/CD8b showed significant correlations. Considering these findings and the results of our functional enrichment analyses, we believe that NUSAP1 plays a key role in the regulation and recruitment of tumor immune infiltrating cells in THYM. However, further studies are warranted to elucidate underlying mechanisms and to assess why NUSAP1 expression is associated with immune infiltration and prognosis.
TFs control chromatin and transcription by recognizing specific DNA sequences, guiding expression of the genome. Mutations in TFs may increase or decrease the risk of tumorigenesis.33 miRNAs, a category of small non-coding RNA, are extensively expressed in various human tumors. They can behave either as tumor suppressor genes or oncogenes by regulating the expression level of their target genes.34–36 Herein we first constructed a miRNA–NUSAP1 and TF–NUSAP1 network to elucidate the upstream modulation mechanism underlying NUSAP1 regulation in LIHC. miR-1236-3p inhibits ovarian carcinoma cell invasion and migration by targeting ZEB1;37 moreover, it suppresses the development of tumor cells by targeting MTA2 in gastric cancer38 and plays a tumor suppressive role in LUAD by targeting KLF8.39 To date, the miRNA targeting NUSAP1 in LIHC remains unknown. Our results confirmed the uniqueness of miR-1236-5p but Further exploration of the mechanisms is needed. TCF3 regulates transcription and leads to epigenetic modifications in tumors. In stage II and III colorectal cancer, the upregulation of TCF3 caused by promoter hypomethylation has been correlated with cancer progression and development.40 In human nasopharyngeal carcinoma, TCF3 accelerates tumor cell growth and metastasis.41 In cervical cancer, TCF3 regulates tumor progression via DDN-AS1-miR-15a/16-TCF3.42 In this study, we analyzed LIHC data acquired from TCGA and found that the expression level of TCF3 was stronger in tumor tissues as compared with that in paired normal tissues. Recently, there was a study showed that overexpression of TCF3 drove hepatocarcinoma development by enhancing cell proliferation via activating the Wnt signaling pathway, PKF118-310, which is a kind of inhibitor of TCF/β-catenin, maybe is a potential negative regulator of NUSAP1.43 Thus, we suggest that miR-1236-5p–NUSAP1 and TCF3–NUSAP1 interactions participate in the regulation of LIHC. Nevertheless, further studies are warranted to validate these findings.
To conclude, our findings demonstrate that NUSAP1 was up-regulated in most of cancer tissues and was involved in the development and progression of various cancer types by promoting EMT via transition of mitotic cell cycle and regulating various immune-related pathways. We believe that our results contribute clarify the different functions and potential mechanisms of NUSAP1 in diverse cancer types. Targeting this gene for therapy can be approached from various directions, providing new avenues for our next phase of in-depth research. Overall, NUSAP1 appears to be a potential prognostic biomarker and therapeutic target for pan-cancer.
Ethical Approval and Consent to Participate
This study was ratified by the Ethics Committee of Nanfang Hospital of Southern Medical University. All participants offered written informed consent before surgery. The 4-week-old male BALB/c mice were purchased from the Experimental Animal Center of Southern Medical University. The study conforms to the provisions of the Declaration of Helsinki.
Acknowledgments
Xiaodi Zhu, Yuting Wu, and Liwei Liao are co-first authors for this study. We thank State Key Laboratory of Oncology in Southern Medical University for providing experimental platform.
Author Contributions
All authors read and approved the final manuscript. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported by Natural Science Foundation of Guangdong Province (2023A1515010285); Clinical Research project of Nanfang Hospital, Southern Medical University (2022CR011、2022CR013); President Foundation of Nanfang Hospital, and Southern Medical University (2020C044).
Disclosure
The authors declare no competing interests in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi:10.3322/caac.21492
2. Myant KB, Cammareri P, Hodder MC, et al. HUWE 1 is a critical colonic tumour suppressor gene that prevents MYC signalling, DNA damage accumulation and tumour initiation. Embo Mol Med. 2017;9(9):181–197. doi:10.15252/emmm.201606684
3. Colecchia D, Rossi M, Sasdelli F, Sanzone S, Strambi A, Chiariello M. MAPK15 mediates BCR-ABL1-induced autophagy and regulates oncogene-dependent cell proliferation and tumor formation. Autophagy. 2015;11:1790–1802. doi:10.1080/15548627.2015.1084454
4. Mullard A. New checkpoint inhibitors ride the immunotherapy tsunami. Nat Rev Drug Discov. 2013;12:489–492. doi:10.1038/nrd4066
5. Li Q, Yuan D, Ma C, et al. A new hope: the immunotherapy in small cell lung cancer. Neoplasma. 2016;63:342–350. doi:10.4149/302_151001N511
6. Han Y, Wu Y, Yang C, et al. Dynamic and specific immune responses against multiple tumor antigens were elicited in patients with hepatocellular carcinoma after cell-based immunotherapy. J Transl Med. 2017;15:15. doi:10.1186/s12967-016-1116-1
7. Raemaekers T, Ribbeck K, Beaudouin J, et al. NuSAP, a novel microtubule-associated protein involved in mitotic spindle organization. J Cell Biol. 2003;162:1017–1029. doi:10.1083/jcb.200302129
8. Gordon CA, Gong X, Ganesh D, Brooks JD. NUSAP1 promotes invasion and metastasis of prostate cancer. Oncotarget. 2017;8:29935. doi:10.18632/oncotarget.15604
9. Qian Z, Li Y, Ma J, et al. Prognostic value of NUSAP1 in progression and expansion of glioblastoma multiforme. J Neuro Oncol. 2018;140:199–208. doi:10.1007/s11060-018-2942-1
10. Liu Z, Guan C, Lu C, et al. High NUSAP1 expression predicts poor prognosis in colon cancer. Pathology Res Pract. 2018;214:968–973. doi:10.1016/j.prp.2018.05.017
11. Li H, Zhang W, Yan M, et al. Nucleolar and spindle associated protein 1 promotes metastasis of cervical carcinoma cells by activating Wnt/β-catenin signaling. J Exp Clin Canc Res. 2019;38:38. doi:10.1186/s13046-019-1046-x
12. Wu X, Xu B, Yang C, et al. Nucleolar and spindle associated protein 1 promotes the aggressiveness of astrocytoma by activating the Hedgehog signaling pathway. J Exp Clin Canc Res. 2017;36(36). doi:10.1186/s13046-017-0597-y
13. Persis P, Wadia MC, Armstrong RJ, Mindrinos M, Butte AJ, Miklos DB. Antibodies specifically targetAMLantigen NuSAP1 after allogeneic bone marrow transplantation. Blood. 2010;115:2077–2087. doi:10.1182/blood-2009-03-211375
14. Ge Y, Li Q, Lin L, et al. Downregulation of NUSAP1 suppresses cell proliferation, migration, and invasion via inhibiting mTORC1 signalling pathway in gastric cancer. Cell Biochem Funct. 2019;38:28–37. doi:10.1002/cbf.3444
15. Zhang Y, Huang K, Cai H, Chen S, Sun D, Jiang P. The role of nucleolar spindle‐associated protein 1 in human ovarian cancer. J Cell Biochem. 2020;121(11):4397–4405.
16. Sun L, Shi C, Liu S, et al. Overexpression of NuSAP1 is predictive of an unfavourable prognosis and promotes proliferation and invasion of triple-negative breast cancer cells via the Wnt/β-catenin/EMT signalling axis. Gene. 2020;747:144657. doi:10.1016/j.gene.2020.144657
17. Xu Z, Wang Y, Xiong J, Cui F, Wang L, Peng H. NUSAP1 knockdown inhibits cell growth and metastasis of non‐small‐cell lung cancer through regulating BTG2/PI3K/Akt signaling. J Cell Physiol. 2019;235:3886–3893. doi:10.1002/jcp.29282
18. Zhang X, Pan Y, Fu H, Zhang J. Nucleolar and spindle associated protein 1 (NUSAP1) inhibits cell proliferation and enhances susceptibility to epirubicin in invasive breast cancer cells by regulating cyclin D kinase (CDK1) and DLGAP5 expression. Med Sci Monitor. 2018;24(24):8553–8564. doi:10.12659/MSM.910364
19. Gao S, Yin H, Tong H, et al. Nucleolar and spindle associated protein 1 (NUSAP1) promotes bladder cancer progression through the TGF-β signaling pathway. Oncotargets Ther. 2020;13:813–825. doi:10.2147/OTT.S237127
20. Chen M, Cen K, Song Y, et al. NUSAP1-LDHA-Glycolysis-Lactate feedforward loop promotes Warburg effect and metastasis in pancreatic ductal adenocarcinoma. Cancer Lett. 2023;567:216285. doi:10.1016/j.canlet.2023.216285
21. Serin EAR, Nijveen H, Hilhorst HWM, Ligterink W. Learning from co-expression networks: possibilities and challenges. Front Plant Sci. 2016;7. doi:10.3389/fpls.2016.00444
22. Wang L, Li Y, Chen G. Network-based co-expression analysis for exploring the potential diagnostic biomarkers of metastatic melanoma. PLoS One. 2018;13:e190447.
23. Gov E, Arga KY. Differential co-expression analysis reveals a novel prognostic gene module in ovarian cancer. Sci Rep. 2017;2017:7.
24. Wang Y, Chen L, Ju L, et al. Novel biomarkers associated with progression and prognosis of bladder cancer identified by co-expression analysis. Front Oncol. 2019;2019:9.
25. Zhao Q, Song W, He DY, Li Y. Identification of key gene modules and pathways of human breast cancer by co-expression analysis. Breast Cancer Tokyo. 2018;25:213–223. doi:10.1007/s12282-017-0817-5
26. Wadia PP, Coram M, Armstrong RJ. ClustVis: a web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015;43(43):W566–70. doi:10.1093/nar/gkv468
27. Jiao X, Sherman BT, Huang DW, et al. DAVID-WS: a stateful web service to facilitate gene/protein list analysis. Bioinformatics. 2012;28(28):1805–1806. doi:10.1093/bioinformatics/bts251
28. Kirkwood JM, Butterfield LH, Tarhini AA, Zarour H, Kalinski P, Ferrone S. Immunotherapy of cancer in 2012. CA Cancer J Clin. 2012;62:309–335. doi:10.3322/caac.20132
29. Mahoney KM, Rennert PD, Freeman GJ. Combination cancer immunotherapy and new immunomodulatory targets. Nat Rev Drug Discov. 2015;14:561–584. doi:10.1038/nrd4591
30. Dong Z, Zhong W, Zhang X, et al.Potential Predictive Value of TP53 and KRAS mutation status for response to PD-1 blockade immunotherapy in lung adenocarcinoma. Clin Cancer Res. 2017;(23):3012–3024. doi:10.1158/1078-0432.CCR-16-2554
31. Xu F, Yin C, Wang C, et al. Immunophenotypes and immune markers associated with acute promyelocytic leukemia prognosis. Dis Markers. 2014;2014:1–6.
32. Pan H, Wang X, Huang W, et al. Interferon-induced protein 44 correlated with immune infiltration serves as a potential prognostic indicator in head and neck squamous cell carcinoma. Front Oncol. 2020;10:10. doi:10.3389/fonc.2020.00010
33. Lambert SA, Jolma A, Campitelli LF, et al. The human transcription factors. Cell. 2018;172:650–665. doi:10.1016/j.cell.2018.01.029
34. Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435:834–838. doi:10.1038/nature03702
35. Fan Y, Zhang L, Sun Y, et al. Expression profile and bioinformatics analysis of COMMD10 in BALB/C mice and human. Cancer Gene Ther. 2020;27:216–225. doi:10.1038/s41417-019-0087-9
36. Yang M, Huang W, Sun Y, et al. Prognosis and modulation mechanisms of COMMD6 in human tumours based on expression profiling and comprehensive bioinformatics analysis. Brit J Cancer. 2019;121(121):699–709. doi:10.1038/s41416-019-0571-x
37. Wang YU, Yan S, Liu X, et al. miR-1236-3p represses the cell migration and invasion abilities by targeting ZEB1 in high-grade serous ovarian carcinoma. Oncol Rep. 2014;31:1905–1910. doi:10.3892/or.2014.3046
38. An J, Ma M, Zhang C, Shao S, Zhou N, Dai D. miR-1236-3p inhibits invasion and metastasis in gastric cancer by targeting MTA2. Cancer Cell Int. 2018;18:18. doi:10.1186/s12935-018-0516-0
39. Bian T, Jiang D, Liu J, et al. miR-1236-3p suppresses the migration and invasion by targeting KLF8 in lung adenocarcinoma A549 cells. Biochem Bioph Res Co. 2017;492:461–467. doi:10.1016/j.bbrc.2017.08.074
40. Li C, Cai S, Wang X, Jiang Z. Hypomethylation-associated up-regulation of TCF3 expression and recurrence in stage II and III colorectal cancer. PLoS One. 2014;9:e112005. doi:10.1371/journal.pone.0112005
41. Shen X, Yuan J, Zhang M, et al. The increased expression of TCF3 is correlated with poor prognosis in Chinese patients with nasopharyngeal carcinoma. Clin Otolaryngol. 2017;42:824–830. doi:10.1111/coa.12834
42. Liu Z, Wu M, Shi H, Huang C, Luo S, Song X. DDN‐AS1‐miR‐15a/16‐TCF3 feedback loop regulates tumor progression in cervical cancer. J Cell Biochem. 2018;120:10228–10238. doi:10.1002/jcb.28307
43. Pu X, Zheng D, Lv T, Zhou Y, Yang J, Jiang L. Overexpression of transcription factor 3 drives hepatocarcinoma development by enhancing cell proliferation via activating Wnt signaling pathway. Hepatob Pancreat Dis. 2022;21:378–386. doi:10.1016/j.hbpd.2022.01.003
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.