Back to Journals » Journal of Inflammation Research » Volume 15
Expression of Human Interleukin 8 in Mice Alters Their Natural Behaviors
Authors Tian Z, Shofer FS, Sandroni AZ, Zhao L, Scanzello CR, Zhang Y ![]()
Received 25 December 2021
Accepted for publication 4 April 2022
Published 13 April 2022 Volume 2022:15 Pages 2413—2424
DOI https://doi.org/10.2147/JIR.S355669
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Zuozhen Tian,1 Frances S Shofer,1,2 Alec Z Sandroni,1 Lan Zhao,3 Carla R Scanzello,4,5 Yejia Zhang1,6
1Department of Physical Medicine & Rehabilitation, Hospital of the University of Pennsylvania, Philadelphia, PA, USA; 2Department of Emergency Medicine, University of Pennsylvania, Philadelphia, PA, USA; 3Department of Orthopedic Surgery, Rush University Medical Center, Chicago, IL, USA; 4Division of Rheumatology, Department of Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA; 5Section of Rheumatology, Corporal Michael J. Crescenz Veterans Affairs Medical Center, Philadelphia, PA, USA; 6Section of Rehabilitation Medicine, Corporal Michael J. Crescenz Veterans Affairs Medical Center, Philadelphia, PA, USA
Correspondence: Yejia Zhang, Department of Physical Medicine & Rehabilitation, Hospital of the University of Pennsylvania, Philadelphia, PA, USA, Email [email protected]; [email protected]
Objective: To examine the effects of human interleukin (IL) 8 expression on mouse behavior.
Methods: A mouse line expressing human IL8 in the intervertebral discs (IVD) and cartilaginous tissues (hIL8+) was generated. Mouse spontaneous behaviors, including locomotion, climbing, rearing, grooming, eating, drinking, and immobility were recorded with a fully automatic, non-invasive platform.
Results: Distance traveled by the hIL8+ mice declined with age compared with control littermates, and male hIL8+ mice traveled a shorter distance than male controls and females of either genotype (p < 0.05). The hIL8+ mice also spent less time in locomotion than control mice (p < 0.01), and male hIL8+ mice spent the least amount of time and had lowest count in locomotion compared with the other 3 groups at 12 weeks of age or greater (p < 0.05). The hIL8+ mice spent less time climbing than controls, and male mice spent less time climbing than female mice of the same genotype (p < 0.01). The hIL8+ mice spent more time eating and less time drinking than controls, and all mice spent less time eating and more time drinking with increasing age. Finally, hIL8+ mice spent more time immobile than controls, and male hIL8+ mice spent more time immobile than any other group (p < 0.05).
Conclusion: The hIL8+ mice, especially hIL8+ males, showed reduced ambulation and climbing. Mice showed age-related decrease in eating and increase in drinking and grooming time that was also influenced by expression of hIL8. These changes in natural behaviors in control mice are consistent with functional decline with age. Effects of hIL8 superimposed on the natural aging process could involve systemic (e.g., on the brain) and local (e.g., in the spine and joint tissues) mechanisms. Future exploration of these mechanisms might be productive.
Keywords: interleukin (IL) 8, mouse, behavior, age, decline
Introduction
Patients with chronic pain often withdraw from activities, use pain medication, and display pain-related body postures and facial expressions. These behaviors are collectively described as “pain behavior”, which can be disruptive to work and social functions. Cytokines/chemokines in both neural and non-neural elements of the nervous system play crucial roles in the pathophysiology of chronic pain.1 Therefore, examining pain behaviors in the context of disc, joint and brain inflammation with elevated circulating chemokines is clinically relevant.
Chronic lower back and joint pain related to intervertebral disc (IVD) and articular cartilage degeneration are major socioeconomic problems, and they often coexist.2 Proinflammatory mediators including interleukin (IL) 8 were found elevated in IVD from patients with back pain,3,4 suggesting that these factors may play a role in the genesis of such pain. Patients with chronic pain are often depressed and demotivated.5 Decreased motivation has been elicited in mouse models of chronic pain, via galanin receptor 1-triggered depression of excitatory synaptic transmission in an indirect pathway involving nucleus accumbens medium spiny neurons.6 These findings suggest that the impacts of spinal and joint pain extend far beyond local inflammation.
Human IL8 is barely detectable in healthy tissues. But it is rapidly induced by IL1a in cultured human IVD cells,3 prompting further study of this chemokine. The murine homolog of IL8, CXCL1, is induced by injury in the mouse IVD,7,8 further confirming its significance in the injury-inflammation-repair responses.
IL8, also known as CXCL8, has been implicated in neutrophil and T-cell migration.9 In humans, IL8 binds to IL8 receptors (IL8RA and B).10 Although rodents lack IL8 receptors, human IL8 can bind to rodent receptors for similar chemokines (CXCR1/2).11,12 For example, hIL8 induces murine corneal ulcer formation.13 To examine the systemic effects of elevated hIL8 in the mouse, a transgenic line with the hIL8 gene inserted into its genome using the Cre-Lox system has been created.14 The hIL8 transgenic mouse line was bred with a mouse line expressing Cre recombinase under the control of the promoter for growth differentiation factor (GDF)5, which is active in both the IVD and articular joint tissues,15–18 the two tissues in which we were interested since they are related to back pain and osteoarthritis. The GDF5Cre drives the expression of hIL8 in both IVD and articular cartilage (hIL8/GDF5Cre mice, abbreviated here as hIL8+). The hIL8 protein has been detected in the serum of hIL8+ mice.14 Given that patients with discogenic pain usually have osteoarthritis, this system is relevant to the human disease.
Previously, as a preliminary effort to investigate the relationship between chemokines and mouse nest-making and other natural behaviors, these were measured at 28 weeks of age.14 At this age, both male and female hIL8+ mice showed reduced nesting scores, spent more time immobile and eating and traveled a shorter distance compared with their hIL8− littermates (controls).14 In the current study, we further examined the natural behaviors of hIL8+ mice from 8 weeks to 28 weeks of age, and observed their functional maturity and decline over time.
Materials and Methods
Mice
A total of 24 mice, mixed male and female, were used in this study. Mice were housed (maximum, 5 per cage) under specific pathogen-free (SPF) conditions and are tested for pathogens quarterly. Mice were fed PicoLab diet 5053 (LabDiet, Fort Worth, TX) without restriction and provided with acidified bottled water and environmental enrichment (Nestlets, Ancare, Bellmore, NY; and Mouse Igloo Rodent Enrichment Device, Fisher Scientific, Hampton, NH). The mice were maintained on a 12:12-h light: dark cycle as described by Brent et al.14
Generation of Transgenic Mice
To construct the expression cassette, hIL8 cDNA, synthesized from cultured human annulus fibrosus (AF) cells stimulated with IL1a,3 was inserted into the pCALL2 plasmid (Miami Mice Research, USA). The human AF cells were isolated from deceased human cadaveric donors through Gift of Hope Organ and Tissue Donor Network (Itasca, IL). Use of tissues from deceased donors is exempt from Institutional Review Board approval, as confirmed by Rush University Medical Center (Chicago, IL). Genotyping was conducted using a nested primer pair (forward, 5ʹTGG AAA GGT TTG GAG TAT GTC TTT A3’; reverse, 5ʹCAG CCT TCC TGA TTT CTG CA3’; sequence based on Ensembl),19 with an annealing temperature of 56 °C. One founder pCALL2-hIL8 transgenic mouse expressing hIL8 at the highest level was bred with a C57BL/6J mouse (Jackson Laboratories, Bar Harbor, ME), followed by sibling-to-sibling inbreeding for at least 7 generations before use in experiments. The Cre-inducible pCALL2-hIL8 transgenic mouse line is available from the Jackson Laboratory as JAX no. 035378.14 A mouse line expressing Cre-recombinase under the control of the GDF5 promoter was generously provided by Dr. David Kingsley at Stanford University.15,20 The hIL8 transgenic mice were bred with GDF5Cre mice to conditionally express the transgene in GDF5-expressing tissues. Human IL8 gene expression in various tissues was verified through real-time PCR analysis. For brevity, hIL8+;GDF5Cre mice are shown as hIL8+, and hIL8−;GDF5Cre mice are shown as controls. All adult mice resulting from the breeding were used in the study of natural behaviors.
Confirmation of hIL8 Gene Expression in Various Tissues with Quantitative Real-Time PCR
Nucleus pulposus (NP), AF, joint articular cartilage (AC), meniscus, Achilles tendon and brain tissues from 5-week-old mice were isolated under a dissecting microscope (VistaVision, VWR International, Radnor, PA). Specifically, NP and AF of 4 coccygeal IVDs (C4/5-C7/8) were pooled; cerebral cortex, articular cartilage from the femoral aspects of one knee joint of hindlimb, and the midportion of one hindlimb Achilles tendon were isolated. The isolated tissues were soaked in RNALater (Ambion, Foster City, CA) overnight, and stored at −80°C until extraction. RNA was extracted and further purified using a RNeasy Micro Kit (Qiagen) as described by Tian et al.7 RNA concentration was determined and cDNA was synthesized using a SuperScript VILO cDNA synthesis kit (Life Technologies, Carlsbad, CA) containing random hexamers and added polyDT primers (Invitrogen, Carlsbad, CA). Primers for real-time PCR were identical to the primer pair for genotyping specified above. For each PCR reaction, cDNA, SYBR Select master mix (Life Technologies, Carlsbad, CA), and primers (working concentration 0.5 μM) were mixed, and run in a ViiA7 real-time PCR system (Applied Biosystems, Foster City, CA) using the following program: (1) 50°C for 2 min, (2) 95°C for 2 min, (3) 95°C for 15 seconds, (4) 58°C for 1 min, (5) repeat steps 3 and 4 a total of 40 times. Single products were confirmed by determining melting curves at the conclusion of the reaction as described by Zhang et al.21
Assessment of Spontaneous Behavior
Each mouse was examined once at each time point (8, 12, 16, 20, 24, and 28 weeks of age). Natural behaviors of hIL8+ mice and their control littermates were recorded overnight (for 16 hours) with the LABORAS (laboratory animal behavior observation, registration and analysis system) platform (Metris, Best, the Netherlands),22 a fully automatic and non-invasive system for recording spontaneous behaviors, including climbing, grooming, eating, drinking, rearing, and resting. Climbing (hanging from a wire cage) and rearing (standing on hind legs) were assessed by a validated algorithm (software provided by Metris). Specifically, all 24 mice completed the study, with behaviors at all 6 time points recorded longitudinally, at 8, 12, 16, 20, 24, and 28 weeks of age. At the time of recording, each mouse occupied a single cage, and as many as 6 animals were examined at each time point. Specifically, each mouse was tested for 16 hours, between 1700 and 0900. Time spent in each activity and the distance traveled during the 16-hour observation period were summed, and speeds of ambulation were averaged. The natural behavior at 28 weeks of age was analyzed further by using MATLAB software (MathWorks, Natick, MA).
Statistics
The large dataset on natural behaviors was collected using the LABORAS platform. Time spent in each activity and distance traveled during the 16-hour observation period were summed, and speed of ambulation was averaged, and further organized with MATLAB software (MathWorks, Natick, Massachusetts). The 95% confidence intervals (CIs) (upper–lower limit) for all the outcome variables are shown as error bars in the line graphs (Figures 1–6). To compare the differences between hIL8+ and control mice in each activity (e.g., distance traveled overnight, average speed), three-factor ANOVA in repeated measures was used, where type of mouse and sex were grouping factors and time was the repeated measure. To adjust for multiple comparisons, post-hoc Tukey-Kramer tests were performed to examine sex- or mouse type-associated differences within each time period. A p value of less than 0.05 was considered statistically significant. All analyses were performed using SAS statistical software (version 9.4, SAS Institute, Cary, NC). The graphs were generated using GraphPad Prism version 9.12 for Windows (GraphPad Software, La Jolla, CA).
 |
Figure 1 Distance traveled and maximum speed during locomotion. (A and B) Comparison of distance traveled between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of maximum speed between the 2 mouse types and mice of different sexes. Values: mean ± 95% confidence intervals; ##: p <0.01 when comparing values within the same cohort over age; **: p <0.01 when comparing values between different cohorts at a single age. |
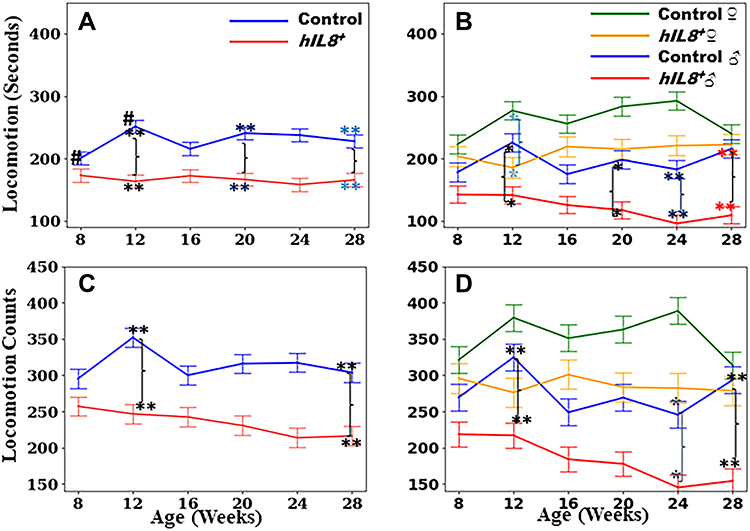 |
Figure 2 Time spent on locomotion and counts in locomotion. (A and B) Comparison of time spent in locomotion between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of locomotion counts between the 2 mouse types, and mice of different sexes. Values: mean ± 95% confidence intervals; #: p <0.05 when comparing values within the same cohort over age; **or *: p <0.01 or p <0.05, respectively, when comparing values between different cohorts at a single age. |
 |
Figure 3 Time spent on climbing and climbing counts. (A and B) Comparison of time spent on climbing between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of climbing counts between the 2 mouse types, and mice of different sexes. Values: mean ± 95% confidence intervals; ## p <0.01 when comparing values within the same cohort over age; **or *: p <0.01 or p <0.05, respectively, when comparing values between different cohorts at a single age. |
 |
Figure 4 Time spent in rearing and rearing counts. (A and B) Comparison of time spent on rearing between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of rearing counts between the 2 mouse types, and mice of different sexes. Values: mean ± 95% confidence intervals; **: p <0.01 when comparing values between different cohorts at a single age. |
 |
Figure 5 Time spent in eating and drinking. (A and B) Comparison of time spent on eating between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of time spent on drinking between the 2 mouse types and mice of different sexes. Values: mean ± 95% confidence intervals; ## or #: p <0.01 or p <0.05, respectively, when comparing values within the same cohort over age; **or *: p <0.01 or p <0.05, respectively, when comparing values between different cohorts at a single age. |
 |
Figure 6 Time spent grooming and immobile. (A and B) Comparison of time spent on grooming between hIL8+ and control mice, and hIL8+ and control mice of different sexes; (C and D) comparison of time spent immobile between the 2 mouse types, and mice of different sexes. Values: mean ± 95% confidence intervals; ##: p <0.01 when comparing values within the same cohort over age; **or *: p <0.01 or p <0.05, respectively, when comparing values between different cohorts at a single age. |
Results
All mice concluded the studies without any dropouts, because the natural behavior study was non-invasive and therefore well tolerated.
Distance Traveled and Maximal Speed
Overall, hIL8+ mice traveled a shorter distance than their control littermates (mean ± 95% CI = 10.7±1.1 m/h vs 12.5±1.1 m/h; n = 12 mice/group; p = 0.03). The hIL8+ mice showed an age-related decline in distance traveled (n = 12, p = 0.02; Figure 1A).
Mice were further divided by sex: hIL8+ males (n = 7), hIL8+ females (n = 5), control males (n = 6), and control females (n = 6). Among these, hIL8+ male mice show the most evident age-related decline in distance traveled, from 11.9±1.4 m/h to 5.6±1.4 m/h between 8 weeks and 28 weeks of age (p <0.01). Consistent with a report by Brent et al.,14 at 28 weeks of age, hIL8+ males traveled a shorter distance than control males, hIL8+ females, or control females (5.6±1.4, 10.1±1.5, 12.9±1.7, and 12.6±1.5 m/h, respectively); hIL8+ males also showed a significant decline with age (p <0.01). Interestingly, control females traveled further than control males at 16–20 weeks of age (p<0.05; Figure 1B).
The hIL8+ mice traveled at a slower maximal speed at age 28 weeks compared with age 12 weeks (mean ± 95% CI = 160.3±13.1 mm/s and 187.4±13.1 mm/s, n = 12, p <0.05). In comparison, maximal speed did not decline significantly in WT controls (n = 12, p >0.05; Figure 1C). The age-related declines in maximum speed among the four subgroups were not statistically significant at any time point (p >0.05; Figure 1D).
Locomotion
The hIL8+ mice spent significantly less time in locomotion than their control littermates (mean ± 95% CI = 166.8±23.8 seconds/hour (s/h) and 229.1±23.3 s/h, n = 12 mice/group; p <0.01). Interestingly, control mice spent more time in locomotion at 12 weeks of age than the same cohort at age 8 weeks (251.4±30.7 s/h and 200.7±20.8 s/h, respectively; p = 0.04), which may represent growth and maturity. However, this peak in locomotion at 12 weeks of age was not observed in the hIL8+ mice (p >0.05; Figure 2A). Male hIL8+ mice spent less time in locomotion than control males at 12 and 28 weeks of age (142.6±27.1 vs 178.3±29.3 s/h, and 109.5±27.2 vs 216.0±29.3 s/h; p <0.05 or 0.01) and hIL8+ females (n = 7 and 5 mice/group, p <0.01). The differences between hIL8+ and control females are significant at the 12-week time point only (141.7±27.1 vs 277.0±29.3 s/h, p = 0.01; Figure 2B).
The pattern of counts in locomotion bears striking similarities to that of time spent in locomotion. Overall, hIL8+ mice walked/ran less frequently than their control littermates (234.7±31.4 vs 314.1±30.9 counts, p<0.01). The hIL8+ moved less frequently than controls at 12 and 20–28 weeks of age (n = 12 mice/group, p <0.01; Figure 2C). Male hIL8+ mice had the lowest counts in locomotion compared with any other group at 12, 24, and 28 weeks of age (n = 5–7 mice/group; p <0.05; Figure 2D).
Climbing
Time spent climbing peaked at 12 weeks of age for control mice, more than the same cohort of mice at 8 or 28 weeks of age (295.9±35.6 vs 196.1±35.5 or 145.2±35.5 s/h; n = 12, p <0.01). The hIL8+ mice did not show this peak activity as 12-week young adults, in contrast to their control littermates (n = 12 mice/group; p >0.05). Accordingly, hIL8+ mice spent less time climbing than control littermates at 12 weeks of age (206.8±36.0 and 295.9±35.6 s/h; p = 0.03; Figure 3A). Control females spent more time climbing than hIL8+ females or males of either genotype at 12 weeks of age (n = 5–7 mice/group; p <0.01). Control females also spent more time climbing than control males at 8, 16, 24, and 28 weeks of age (n = 6 mice/group; p <0.01). Interestingly, control males and hIL8+ males spent a similar amount of time climbing (n = 6 and 7 mice/group, p >0.05; Figure 3B).
Climbing counts in control mice peaked at 12 weeks of age, higher than for the same cohort at 8 or 28 weeks of age (12.5±1.6 vs 7.8±1.6 or 7.3±1.6 counts; n = 12, p <0.01). However, this peak is visibly absent in the hIL8+ group (n = 12, p >0.05). Accordingly, hIL8+ mice climbed significantly less frequently than their control littermates at 12 weeks of age (6.6±1.6 counts and 12.5±1.6 counts, respectively; p <0.01; Figure 3C). At 12 weeks of age, control females spent more time climbing than any other group (hIL8+ females, control males, or hIL8+ males) (n = 5–7 mice/group, p <0.01). Interestingly, control females climbed more frequently than control males at ages 12 weeks and older (n = 6 mice/group; p <0.01), although climbing frequency in male controls and male hIL8+ mice was similar (n = 6 and 7 mice/group; p >0.05; Figure 3D).
Rearing
There were no statistically significant differences in time spent rearing between mice of the two genotypes or sexes (p >0.05, Figure 4A and B). The hIL8+ mice reared less frequently (counts) than controls at 12 weeks of age (45.2±5.3 vs 55.5±5.3; n = 12/group, p <0.01; Figure 4C). Female hIL8+ mice reared less frequently than female controls at 12 weeks of age (38.4±9.5 vs 70.98.6 counts; n = 5 and 6, respectively; p <0.01). There were no statistically significant differences between hIL8+ and control males (n = 7 and 6 mice/group, respectively; p >0.05, Figure 4D).
Eating and Drinking
Time spent eating decreased progressively with age for both the hIL8+ and control mice (hIL8+ at 8 and 28 weeks of age: 284.8±31.1 and 171.4± 31.1 s; controls at 8 and 28 weeks of age: 238.1±30.7 and 146.1±30.6 s; n = 12 mice/group; p <0.01; Figure 5A). The hIL8+ mice spent more time eating than their littermate controls at 16 and 24 weeks of age (n = 12 mice/group; p <0.05). Male hIL8+ mice spent more time eating than controls at age 16 weeks (n = 7 and 6, respectively; p = 0.036; Figure 5B).
Control mice spent increased time drinking with age (8 and 28 weeks of age: 4.9±4.4 and 25.2±4.4 s/h; n = 12; p <0.01), and a similar trend existed for hIL8+ mice, although not statistically significant (n = 12 mice; p >0.05). The hIL8+ mice spent less time drinking than their littermate controls at 28 weeks of age (12.0±4.5 vs 25.2±4.4 s/h, p <0.01; Figure 5C). Male hIL8+ mice spent less time drinking than male littermate controls at 28 weeks of age (n = 7 and 6 mice/group, respectively; p <0.01); this is consistent with findings by Brent et al.14 Both male and female controls spent more time drinking at 28 weeks than at 8 weeks of age (n = 6/group, p <0.01 and p <0.05, respectively; Figure 5D).
Grooming
Control mice spent more time grooming at 28 weeks of age than at 8 weeks of age (343.8±39.7 and 214.5±39.8 s/h, respectively; n = 12, p <0.01). The hIL8+ mice showed a similar trend, but the difference was not statistically significant (n = 12, p = 0.29; Figure 6A). When separating the mice by sex, no statistically significant differences in grooming time were found (n = 5–7 mice/group, p >0.05; Figure 6B). However, no excessive hair loss was noticed with increasing age.
Immobility
The hIL8+ mice spent more time immobile than controls (1452.9±98.6 vs 1226.6±97.2 s/h; n = 12 mice/group; p <0.01; Figure 6C). At 28 weeks of age, hIL8+ males spent the most time in immobility of any group, i.e., more than control males, and hIL8+ and control females (n = 5–7 mice/group; p <0.05; Figure 6D).
Discussion
The hIL8+ mice were generated to express human IL8 in the IVD and articular cartilage. Expression of the hIL8 gene has been confirmed in the intervertebral disc (IVD), joint, and brain tissues in hIL8+ mice, and is absent in the control littermates (n = 3 hIL8+ and control littermate pairs; Figure S1). The hIL8 protein has been detected in the serum of these mice, potentially affecting the brain as well as spinal and joint tissues.14 Reduced activity and nest-making impairment have been shown in both male and female hIL8+ mice compared with littermate controls at 28 weeks of age.14 Findings presented here are a natural extension from that previous study, describing the effects of hIL8 on mouse activity from young adulthood (8–12 weeks of age) to 28 weeks. Multiple cytokines/chemokines have been found in IVD tissues3,23,24 and serum25,26 of patients with back pain.4 Among the cytokines/chemokines found in the painful human IVD tissues, the IL8 gene expression and protein production showed highest magnitude of elevation in response to inflammation.3 Therefore, the hIL8-expressing mouse line was generated to observe any consequent behaviors. Intriguingly, these mice exhibit some of the behavioral characteristics observed in human patients (e.g., reduced voluntary activities). The impacts on behavior by other cytokines/chemokines found in patients with back pain and arthritis such as IL6 are worth investigating in the future.
With increasing age, hIL8+ mice traveled shorter distances at slower maximal speed. The hIL8+ mice spent less time in locomotion than controls, and most of the differences could be accounted for by less travel frequency. This phenomenon was more pronounced in the male than in female hIL8+ mice. Male hIL8+ mice also spent the longest time immobile compared with any other group (female hIL8+, male and female controls). Human IL8 is expressed in hIL8+ mice, which binds to the mouse receptors CXCR1/2.11 Human IL8 is found in serum of hIL8+ mice,14 in addition to expression in disc and cartilaginous tissues. Therefore, systemic effects of this chemokine are likely. One limitation is that we cannot attribute the change in behavior specifically to joint/back pain with the existing data. The GDF5-Cre drives the hIL8 gene expression in IVD, joint, and brain tissues. The hIL8 in any of these tissues could contribute to the behavior changes observed in mice. In addition, the human IL8 protein has been secreted into the blood, as shown in the serum by ELISA.14 Since cytokines/chemokines are found in human patients with back and joint pain, the fact that hIL8+ mouse IVD, joint, and brain expressed the hIL8 gene may be relevant to at least a subset of patients. Time spent rearing did not differ significantly between the hIL8+ and littermate control mice, but the hIL8+ mice made fewer attempts to rear (rearing counts) than littermate control mice. The mechanism of reduced rearing count in the IL8+ mice could be back/joint pain, since these mice expressed human IL8 in the spine and knee joints, which may cause pain. Alternatively, loss of motivation may contribute to the reduced rearing counts. Importantly, peripheral inflammation has been shown to affect the central nervous system.27 The Quan group has shown that Interleukin (IL) 1 receptor 3 mediates IL1-induced potassium current increase through fast activation of Akt kinase,28 which may help in explaining “sickness behavior”.29 The behavioral changes in the hIL8+ mice presented here could be mediated by a combination of peripheral (spine and joint inflammation) and central nervous system mediated mechanisms. Future experiments expressing hIL8 specifically in the IVD or joint tissues, and measurements of local mechanical sensitivities with the Von Frey filament may help to tease out the contributions of peripheral tissues and the central nervous system to pain behaviors, and will be explored in future studies.
Likewise, climbing activity followed similar trends to that seen in travel and rearing in strain, age, and sex-related differences. Laboratory mice in standard laboratory cages spontaneously display cage-grid climbing as one of the major components of spontaneous home-cage behavior.30 Climbing time and counts peaked at 12 weeks of age for control mice, but this peak was not evident in the hIL8+ mice. Both the hIL8+ and control female mice spent more time climbing than males of the same genotypes. Sex-related differences in activities observed are consistent with a recent publication that described female C57BL mice traveling significantly longer distances (by 30%), and showing greater grid-climbing activity (by 50%) than males.31 Estrous cycle is known to affect behaviors of female mice.32 The estrous cycle was not recorded in the current study, and might be an important direction for future research.
Both IL8+ and littermate control mice spent less time eating as their ages increased from 8 weeks to 28 weeks. This may reflect, in part, lower energy needed in the aging mice. Time spent eating at 16 and 24 weeks of age was greater in the hIL8+ mice than in littermate controls, but the mouse weight was not significantly different. Mice spent more time drinking with increased age, and IL8+ mice spent less time drinking than littermate controls at the 28-week time point. Eating and drinking are reported to correlate with cytokine/chemokine levels in the intestinal tract in the mouse colitis model.33 Both activities required that the mice stand up in order to reach the food and water, and may therefore be influenced by pain in the spine/joints. As reported previously, male mice were heavier than females for mice of both genotypes, but there was no significant difference in the weights between hIL8+ and control mice at 28 weeks of age.14 The body weight reflects food intake and energy expenditure. We did not record food and water consumption, which would be important measures to document in the future.
Over-grooming (excessive hair loss) in either hIL8+ or control mice has not been observed, although grooming activity increased in the control mice with increasing age, with a similar trend in the hIL8+ mice. Both hormonal status and age could influence rodent anxiety-like behavior including over-grooming.34–36 Although the hIL8+ and control mice spent a similar amount of time grooming, the hIL8+ mice built worse nests than control mice,14 suggesting that these behaviors reflect related yet separate functions.
It is well known that disc degeneration is poorly correlated with inflammation/pain in human patients.37,38 As mechanisms leading to tissue degeneration and inflammation/pain may be distinct, it is not surprising that no significant differences in histological features of IVD and joint tissues were detected between the hIL8+ mice and their control littermates at 28 weeks of age.14 Similar to our findings, transgenic mice expressing human TNFα did not show overt IVD degeneration, although they experienced early onset spontaneous disc herniation.39 Furthermore, in a mouse line with ADAM8 (ADAM Metallopeptidase Domain 8) inactivated, there is elevated inflammatory gene expression along with better preserved disc structure.21 Behavioral changes in these mutant mice may be informative.
In a hIL8 conditional expression mouse line, expression of hIL8 after birth in the intestinal tract increased mobilization of immature myeloid cells to local sites and accelerated carcinogenesis.40 This hIL8 expression model was generated using a bacterial artificial chromosome encompassing the entire IL8 gene with regulatory elements, thus allowing induction of hIL8 via a local pathophysiological stimulus such as azoxymethane in the gastrointestinal tract. This model is advantageous since temporal control of IL8 gene expression can be achieved, while the hIL8+ mouse used in the present study expressed the gene at inception. In yet another study using human IL8 expression in the liver, IL8 substantially limited hepatic apoptosis, reduced liver hemorrhage and lengthened survival in mice treated with galactosamine and endotoxin.41 These findings suggest that hIL8 expression locally and systemically results in a variety of effects, in addition to influencing leukocyte trafficking. A comparative study of behaviors of mouse lines with conditional expression of IL8 during different developmental stages and in response to injuries would be valuable.
In summary, mice expressing the hIL8 gene show changes in natural behaviors compared with littermate controls. These changes are especially evident in male mice with increasing age. One limitation of this study is that the hIL8 gene is constitutively expressed in the spine, joint, and brain tissues. Future experiments with more specific tissue expression at different stages of life would provide detailed information on the mechanisms of behavioral changes. Another future direction is specific pain-related behavior tests (e.g., Von Frey test) to differentiate central nervous system mediated behavior changes from spine/joint pain.
Conclusion
Both hIL8+ mice and their control littermates traveled shorter distances, and traveled less frequently at 28 weeks of age than when they were younger (8–12 weeks). The decline in activities is most evident in the male hIL8+ mice. Mice also spent less time eating, and more time drinking, grooming and immobile with increased age. These behavior changes in the hIL8+ mice are reminiscent of human patients with chronic back/joint pain, whose local and systemic inflammation often coincides with behavioral manifestations. Further studies to differentiate central or peripheral mechanisms of these changes may shed light on mechanisms of behavioral changes.
Ethics Approval
All animal use in this study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the Corporal Michael J. Crescenz Veterans Affairs Medical Center, Philadelphia, PA. All methods were performed in accordance with the following policies and guidelines for care and use of animals: the United States Humane Care and Use of Laboratory Animals Policy by the Office of Laboratory Animal Welfare (OLAW), the American Association for Accreditation of Laboratory Animal Care (AAALAC) Guide for the Care and Use of Laboratory Animals, and Department of Veterans Affairs policy on studies involving animals. This study is compliant with the Animal Research Reporting of In Vivo Experiments (ARRIVE) guidelines.
Acknowledgments
We gratefully thank Dr. Motomi Enomoto-Iwamoto, DDS, PhD and Randel Swanson II, DO, PhD for valuable advice, and Martin F. Heyworth, MD for critically editing the manuscript. The authors thank Yan Xiu, MD, PhD and Flavia Vitale, PhD, for technical support and organizing data, and Debra Pawlowski and Pierre Conti, VMD for care of the animals. The results were partially presented as a poster presentation at the Orthopaedic Research Society, Philadelphia Spine Research Society (ORS PSRS) 5th International Spine Research Symposium, November 3–7, 2019.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. All authors have read and approved the final submitted manuscript.
Funding
This work was supported, in part, by funds provided by the Department of Physical Medicine and Rehabilitation to Zhang. Funding was also received from the Department of Veterans Affairs Healthcare Network-VISN 4, and the National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIH/NIAMS, R21 AR071623 to Zhang). All authors approved the final version of the manuscript submitted. None of the authors have any professional or financial affiliations that may be perceived to have biased the presentation.
Disclosure
The authors have no financial or non-financial conflicts of interest to disclose in this work.
References
1. White FA, Wilson NM. Chemokines as pain mediators and modulators. Curr Opin Anaesthesiol. 2008;21(5):580–585.
2. Weinstein S, Yelin E, Ahn J, et al. United States bone and joint initiative: the burden of musculoskeletal diseases in the United States (BMUS). 2014.
3. Zhang Y, Chee A, Shi P, et al. Intervertebral disc cells produce interleukins found in patients with back pain. Am J Phys Med Rehabil. 2016;95(6):407–415.
4. Zhang Y, Yao L, Robinson KM, Dillingham TR. Biomarkers in the degenerative human intervertebral disc tissue and blood. Am J Phys Med Rehabil. 2021;2:646.
5. Bletzer J, Gantz S, Voigt T, Neubauer E, Schiltenwolf M. Chronic low back pain and psychological comorbidity: a review. Schmerz. 2017;31(2):93–101.
6. Schwartz N, Temkin P, Jurado S, et al. Chronic pain. decreased motivation during chronic pain requires long-term depression in the nucleus accumbens. Science. 2014;345(6196):535–542.
7. Tian Z, Ma X, Yasen M, et al. Intervertebral disc degeneration in a percutaneous mouse tail injury model. Am J Phys Med Rehabil. 2018;97(3):170–177.
8. Brent JM, Tian Z, Shofer FS, et al. Influence of genetic background and sex on gene expression in the mouse (mus musculus) tail in a model of intervertebral disc injury. Comp Med. 2020;70(2):131–139.
9. Gerszten RE, Garcia-Zepeda EA, Lim YC, et al. MCP-1 and IL-8 trigger firm adhesion of monocytes to vascular endothelium under flow conditions. Nature. 1999;398(6729):718–723.
10. Stillie R, Farooq SM, Gordon JR, Stadnyk AW. The functional significance behind expressing two IL-8 receptor types on PMN. J Leukoc Biol. 2009;86(3):529–543.
11. Fan X, Patera AC, Pong-Kennedy A, et al. Murine CXCR1 is a functional receptor for GCP-2/CXCL6 and interleukin-8/CXCL8. J Biol Chem. 2007;282(16):11658–11666.
12. Lee J, Cacalano G, Camerato T, Toy K, Moore MW, Wood WI. Chemokine binding and activities mediated by the mouse IL-8 receptor. J Immunol. 1995;155(4):2158–2164.
13. Oka M, Norose K, Matsushima K, Nishigori C, Herlyn M. Overexpression of IL-8 in the cornea induces ulcer formation in the SCID mouse. Br J Ophthalmol. 2006;90(5):612–615.
14. Brent JM, Tian Z, Yao L, et al. Functional deficits in mice expressing human interleukin 8. Comp Med. 2020;70(3):205–215.
15. Capellini TD, Chen H, Cao J, et al. Ancient selection for derived alleles at a GDF5 enhancer influencing human growth and osteoarthritis risk. Nat Genet. 2017;49(8):1202–1210.
16. Chen H, Capellini TD, Schoor M, Mortlock DP, Reddi AH, Kingsley DM. Heads, shoulders, elbows, knees, and toes: modular Gdf5 enhancers control different joints in the vertebrate skeleton. PLoS Genet. 2016;12(11):e1006454.
17. Koyama E, Shibukawa Y, Nagayama M, et al. A distinct cohort of progenitor cells participates in synovial joint and articular cartilage formation during mouse limb skeletogenesis. Dev Biol. 2008;316(1):62–73.
18. Rountree RB, Schoor M, Chen H, et al. BMP receptor signaling is required for postnatal maintenance of articular cartilage. PLoS Biol. 2004;2(11):e355.
19. Howe KL, Achuthan P, Allen J, et al. Ensembl 2021. Nucleic Acids Res. 2021;49(D1):D884–D891.
20. Storm EE, Kingsley DM. GDF5 coordinates bone and joint formation during digit development. Dev Biol. 1999;209(1):11–27.
21. Zhang Y, Tian Z, Gerard D, et al. Elevated inflammatory gene expression in intervertebral disc tissues in mice with ADAM8 inactivated. Sci Rep. 2021;11(1):180.
22. Van de Weerd HA, Bulthuis RJ, Bergman AF, et al. Validation of a new system for the automatic registration of behaviour in mice and rats. Behav Processes. 2001;53(1–2):11–20.
23. Kepler CK, Markova DZ, Dibra F, et al. Expression and relationship of proinflammatory chemokine RANTES/CCL5 and cytokine IL-1beta in painful human intervertebral discs. Spine. 2013;38(11):873–880.
24. Shamji MF, Setton LA, Jarvis W, et al. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010;62(7):1974–1982.
25. Sowa GA, Perera S, Bechara B, et al. Associations between serum biomarkers and pain and pain-related function in older adults with low back pain: a pilot study. J Am Geriatr Soc. 2014;62(11):2047–2055.
26. Weber KT, Satoh S, Alipui DO, et al. Exploratory study for identifying systemic biomarkers that correlate with pain response in patients with intervertebral disc disorders. Immunol Res. 2015;63(1–3):170–180.
27. Quan N, Whiteside M, Kim L, Herkenham M. Induction of inhibitory factor kappaBalpha mRNA in the central nervous system after peripheral lipopolysaccharide administration: an in situ hybridization histochemistry study in the rat. Proc Natl Acad Sci U S A. 1997;94(20):10985–10990.
28. Qian J, Zhu L, Li Q, et al. Interleukin-1R3 mediates interleukin-1-induced potassium current increase through fast activation of akt kinase. Proc Natl Acad Sci U S A. 2012;109(30):12189–12194.
29. Quan N, Banks WA. Brain-immune communication pathways. Brain Behav Immun. 2007;21(6):727–735.
30. Buttner D. Climbing on the cage lid, a regular component of locomotor activity in the mouse. J Exp Anim Sci. 1991;34(5–6):165–169.
31. Borbelyova V, Janisova K, Myslivecek J, Riljak V. Sex-related differences in locomotion and climbing of C57Bl/6NTac mice in a novel environment. Physiol Res. 2019;68(Suppl 3):S353–S359.
32. Datta S, Samanta D, Tiwary B, Chaudhuri AG, Chakrabarti N. Sex and estrous cycle dependent changes in locomotor activity, anxiety and memory performance in aged mice after exposure of light at night. Behav Brain Res. 2019;365:198–209.
33. Anderson SJ, Lockhart JS, Estaki M, et al. Effects of azithromycin on behavior, pathologic signs, and changes in cytokines, chemokines, and neutrophil migration in C57BL/6 mice exposed to dextran sulfate sodium. Comp Med. 2019;69(1):4–15.
34. Domonkos E, Borbelyova V, Csongova M, et al. Sex differences and sex hormones in anxiety-like behavior of aging rats. Horm Behav. 2017;93:159–165.
35. Domonkos E, Hodosy J, Ostatnikova D, Celec P. On the role of testosterone in anxiety-like behavior across life in experimental rodents. Front Endocrinol (Lausanne). 2018;9:441.
36. Trankner D, Boulet A, Peden E, Focht R, Van Deren D, Capecchi M. A microglia sublineage protects from sex-linked anxiety symptoms and obsessive compulsion. Cell Rep. 2019;29(4):791–799.e3.
37. Boden SD, McCowin PR, Davis DO, Dina TS, Mark AS, Wiesel S. Abnormal magnetic-resonance scans of the cervical spine in asymptomatic subjects. A prospective investigation. J Bone Joint Surg Am. 1990;72(8):1178–1184.
38. Boden SD, Davis DO, Dina TS, Patronas NJ, Wiesel SW. Abnormal magnetic-resonance scans of the lumbar spine in asymptomatic subjects. A prospective investigation. J Bone Joint Surg Am. 1990;72(3):403–408.
39. Gorth DJ, Shapiro IM, Risbud MV. Transgenic mice overexpressing human TNF-alpha experience early onset spontaneous intervertebral disc herniation in the absence of overt degeneration. Cell Death Dis. 2018;10(1):
40. Asfaha S, Dubeykovskiy AN, Tomita H, et al. Mice that express human interleukin-8 have increased mobilization of immature myeloid cells, which exacerbates inflammation and accelerates colon carcinogenesis. Gastroenterology. 2013;144(1):155–166.
41. Hanson JC, Bostick MK, Campe CB, et al. Transgenic overexpression of interleukin-8 in mouse liver protects against galactosamine/endotoxin toxicity. J Hepatol. 2006;44(2):359–367.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.