Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 21
Exploring the Roles of Wnt Signaling Pathway and Non-Coding RNAs in COPD
Authors Xu Y, Lu P, Duan H, Liu F ![]()
Received 26 March 2026
Accepted for publication 8 July 2026
Published 14 July 2026 Volume 2026:21 612141
DOI https://doi.org/10.2147/COPD.S612141
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Zijing Zhou
Yanling Xu,1 Peng Lu,1 Hanmei Duan,2 Feng Liu1
1Department of Emergency Medicine, Hengyang Central Hospital, Hengyang, Hunan, 421200, People’s Republic of China; 2Department of Endocrinology, Hengyang Central Hospital, Hengyang, Hunan, 421200, People’s Republic of China
Correspondence: Feng Liu, Department of Emergency Medicine, Hengyang Central Hospital, 10 Yancheng Road, Yanfeng, Hengyang, Hunan, 421200, People’s Republic of China, Email [email protected]
Abstract: Chronic obstructive pulmonary disease (COPD) is a chronic respiratory disease with one of the leading causes of global morbidity and mortality. Its core pathological features include irreversible airflow limitation, chronic airway inflammation, and lung tissue remodeling. In recent years, the key roles of the Wnt signaling pathway and non-coding RNAs (ncRNAs) in the pathogenesis of COPD have gained increasing attention. As a pivotal signaling cascade governing cell proliferation, differentiation and tissue repair, the Wnt pathway undergoes aberrant activation or suppression in COPD, linking closely to airway inflammation, alveolar epithelial damage and pulmonary fibrosis. Non-coding RNAs including miRNAs, lncRNAs and circRNAs modulate COPD pathogenesis via targeting core molecules of the Wnt pathway. This review summarizes the basic components and functions of the Wnt pathway and the classification and biological roles of ncRNAs and discusses their interactive mechanisms and therapeutic strategies in COPD. In addition, this review discusses the current controversies and future directions. Studies indicate that the synergistic regulatory network between the Wnt pathway and ncRNAs provides new molecular targets for the early diagnosis, prognosis assessment, and targeted therapy of COPD, highlighting potential avenues for future therapeutic development.
Keywords: COPD, Wnt signaling pathway, ncRNAs
Introduction
Chronic obstructive pulmonary disease (COPD) is a common chronic respiratory disease with its prevalence and mortality rates showing a continuous upward trend.1,2 Recent studies have shown that COPD is the third leading cause of death worldwide, posing a serious threat to human health and representing a major public health issue.3 COPD affects 13.1% of people worldwide and 13.7% of Chinese people over 40 years old.4,5 To date, the etiology of COPD remains unclear. Current studies indicate that the development and progression of COPD result from the interplay of genetic and environmental factors.6 Studies have indicated that genetic predisposition plays a significant role in COPD pathogenesis. For instance, genome-wide association studies (GWAS) have identified multiple genetic loci associated with COPD that may increase an individual’s disease risk by influencing lung function and disease progression.7 Additionally, variations in specific genes, such as SERPINA1, EPHX1, GST, MMP12, TGF-β1, and SERPINE2 have been linked to susceptibility to COPD.6,8,9 Environmental factors also play a critical role in COPD development. While smoking is the primary environmental risk factor, other factors, such as air pollution, occupational exposure, and biomass fuel use are also recognized as significant environmental risk factors.10 These epidemiological characteristics suggest that the prevention and management of COPD requires comprehensive consideration of environmental exposure, genetic susceptibility, and comorbidity management.
The molecular mechanisms underlying COPD involve multiple signaling pathways and molecular regulatory networks. Recent studies have indicated that the Wnt signaling pathway, non-coding RNAs (nc-RNAs), and their interactions may serve as key molecular bridges connecting these risk factors to disease progression.11–13 MicroRNAs (miRNAs) participate in the inflammatory response and airway remodeling in COPD by regulating the expression of target genes. For example, miR-29c and miR-126 are significantly downregulated in patients with COPD and are associated with disease severity.13 Additionally, long non-coding RNAs (lncRNAs), such as MALAT1 and TUG1 are involved in the pathogenesis of COPD.14,15 For example, lncRNA TUG1 is significantly upregulated in the lung tissues of patients with COPD and promotes airway remodeling by regulating the miR-145-5p/DUSP6 axis. Aberrant activation of the canonical Wnt/β-catenin pathway is associated with airway smooth muscle hyperplasia and fibrosis in COPD patients. For instance, β-catenin expression is significantly elevated in airway smooth muscle cells of patients with COPD and is positively correlated with airway wall thickness.16 The non-canonical Wnt pathway, such as the Wnt5a pathway, is also involved in the pathogenesis of COPD. Wnt5a expression is significantly upregulated in the lung tissues of patients with COPD and promotes emphysema formation by inhibiting the classical Wnt pathway.17 These results indicate that abnormal regulation of nc-RNAs and the Wnt pathway is a critical component in the pathological progression of COPD. Although increasing studies have confirmed that the interaction between non-coding RNAs and the Wnt signaling pathway regulates the occurrence and progression of COPD, relevant reviews summarizing their interaction mechanisms are still lacking. Therefore, this review aimed to elucidate the pivotal role of the ncRNA-Wnt signaling axis in COPD, highlighting potential avenues for future therapeutic development (Figure 1).
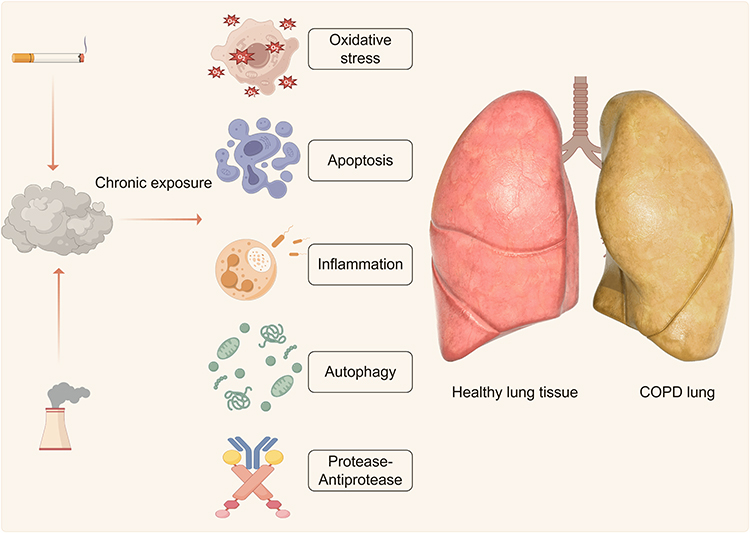 |
Figure 1 The etiology of COPD. Chronic exposure to smoke can induce oxidative stress, apoptosis, inflammation, autophagy, and protease-antiprotease biological process. These contribute to the development of COPD. |
Wnt Signaling Pathway and COPD
Basic Components and Function of the Wnt Signaling Pathway
The Wnt signaling pathway is a highly conserved signaling transduction pathway that plays a central role in embryonic development, tissue homeostasis maintenance, tissue regeneration, and disease pathogenesis.18–20 It is primarily classified into three major branches: the canonical Wnt/β-catenin signaling pathway, the non-canonical planar cell polarity (PCP)/Wnt signaling pathway, and the non-canonical calcium-dependent Wnt signaling pathway.21 Canonical Wnt/β-catenin signaling pathway is dependent onβ-catenin.22 However, the non-canonical Wnt signaling pathway is not dependent on β-catenin activation.23 Activation of the canonical Wnt signaling pathway depends on the binding of Wnt ligands to transmembrane co-receptors composed of the Frizzled (FZD) receptor and low density lipoprotein receptor-related protein 5 or 6 (LRP5/6), which inhibit the activity of the β-catenin degradation complex (composed of adenomatous polyposis coli (APC), glycogen synthase kinase 3 protein (GSK3), AXIN, etc), leading to the accumulation of β-catenin in the cytoplasm and its entry into the nucleus.β-catenin binds to TCF/LEF transcription factors to regulate the expression of target genes such as c-Myc and Cyclin D1.24 For example, inhibition of the canonical Wnt signaling pathway promotes epithelial branching during morphogenesis of submandibular gland branches in embryonic mice.25 The non-canonical Wnt signaling pathway regulates cell polarity, migration, and inflammatory responses by activating kinases such as calcium/calmodulin-dependent protein kinase II (CaMKII), c-Jun N-terminal kinases (JNK), and protein kinase C (PKC).26 For instance, Wnt5a promotes chondrocyte degradation in osteoarthritis through a nonclassical pathway27 (Figure 2).
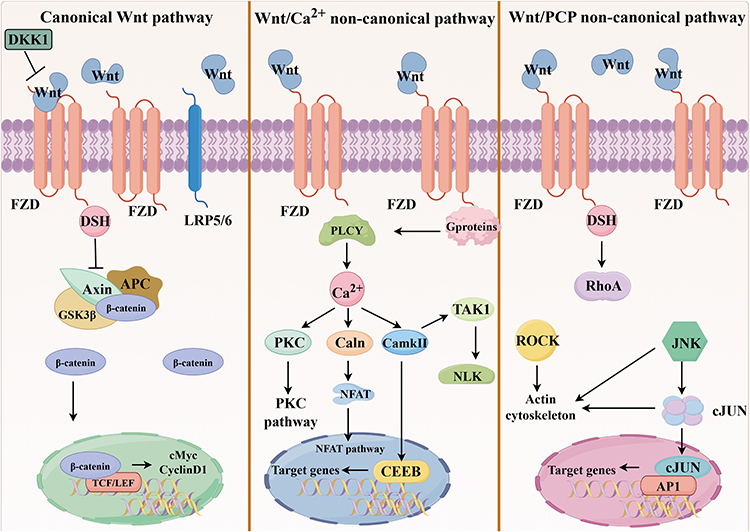 |
Figure 2 The Wnt signaling pathway. It is primarily classified into three major branches: the canonical Wnt/β-catenin signaling pathway, the non-canonical calcium-dependent Wnt signaling pathways, and the non-canonical PCP/Wnt signaling pathway. |
Pathological Role of Wnt Signaling Pathway in COPD
The Wnt signaling pathway plays a complex role in the pathogenesis of COPD, with abnormal activation or inhibition participating in multiple pathophysiological processes. The Wnt signaling pathway regulates the pathogenesis of COPD through multiple mechanisms, such as the inflammatory response, epithelial apoptosis, airway remodeling, and the imbalance of the canonical and non-canonical pathway”.28–30 These multiple mechanisms may work together to regulate COPD pathogenesis.
The Role of Wnt Signaling Pathway in Inflammatory Response of COPD
The Wnt signaling pathway plays a critical role in the inflammatory response in COPD. Abnormal activation or inhibition can influence the progression of COPD by regulating the infiltration of inflammatory cells and release of inflammatory factors.31 Previous studies have confirmed that Wnt4 is upregulated in primary bronchial epithelial cells (PBEC) from patients with COPD and amplifies inflammation triggered by cigarette smoke.31,32 Interestingly, it did not affect the canonical WNT target genes but triggered the activation of the non-canonical signaling molecule p38. In addition, activation of the canonical Wnt/β-catenin pathway can suppress the inflammatory responses in COPD. Studies have shown that Wnt3a can promote the expression of antioxidant enzymes by activating the (nuclear factor erythroid-2 related factor-2) Nrf2 pathway, thereby alleviating oxidative stress-induced inflammation.30 Wnt3a overexpression significantly reduced IL-6 and IL-8 secretion in human bronchial epithelial cells (NHBE) treated with cigarette smoke extract (CSE), whereas Wnt3a knockdown further increased the secretion of these inflammatory factors.
Activation of the non-canonical Wnt signaling pathway promotes inflammatory responses in COPD. Wnt5a, a key ligand of the non-classical Wnt pathway, exhibits significantly elevated expression in the lung tissues of patients with COPD and can promote the expression of inflammatory factors (such as IL-6 and IL-8) by activating the NF-κB pathway. In alveolar macrophages of patients with COPD, Wnt5a treatment increasedIL-6 secretion.33,34 In patients with COPD, the levels of Wnt5a- and Wnt5b-induced IL-6 and CXCL8 secretion in pulmonary fibroblasts were significantly higher than those in healthy controls.33 Additionally, the Wnt signaling pathway can influence the inflammatory response in COPD by regulating the activation and proliferation of T lymphocytes.
Previous studies have shown that secreted frizzled-related protein 2 (sFRP2) inhibits the Wnt pathway by competitively binding to Wnt ligands of the frizzled protein receptor, thereby exacerbating airway inflammation and disrupting Th17/Treg homeostasis.35,36 CSE exposure increases sFRP2 expression, which in turn inhibits Wnt/β-catenin signaling, leading to Th17 cell expansion and Treg cell suppression. This suggests that sFRP2-induced Wnt suppression is a key driver of Th17/Treg imbalance. Additionally, sFRP2 may indirectly affect T cell balance by modulating dendritic cells (DCs) function. DCs are critical antigen-presenting cells that regulate Th17 and Treg differentiation, and their function is impaired in COPD.37 Results found that COPD patients had reduced mature DC (mDC) levels and increased immature DC (imDC) levels, with a positive correlation between imDC frequency and Th17/Treg ratio. Furthermore, sFRP2 has been shown to regulate macrophage polarization.38 These findings indicate that sFRP2 might also modulate the Th17/Treg balance via inhibiting Wnt signaling pathway in COPD.
The Effect of Wnt Signaling Pathway on Apoptosis in COPD
The Wnt signaling pathway plays a critical role in apoptosis in COPD. Abnormal activation or inhibition can influence cell survival and death in the lung tissue of patients with COPD by regulating the expression of apoptosis-related genes. Previous studies have indicated that inhibition of the Wnt signaling pathway alleviates pulmonary fibrosis by promoting lung fibroblast apoptosis.39 In addition, studies have shown that bone marrow mesenchymal stem cell (BMSCs)-derived exosomes can decrease apoptosis of pulmonary microvascular endothelial cells in CSE-induced COPD by targeting Wnt5a.40 In a cigarette smoke-induced COPD mouse model, treatment with the Wnt pathway activator, LiCl, significantly reduced the apoptosis rate of alveolar epithelial cells and improved lung function.30 Similar studies have indicated that activation of the Wnt5a signaling pathway promotes proliferation and inhibits apoptosis of normal lung fibroblasts. This effect is mediated by the non-canonical Wnt signaling pathway.41
Regulatory Mechanism of Wnt Signaling Pathway in Airway Remodeling of COPD
The Wnt signaling pathway plays a pivotal role in airway remodelling in COPD. Its abnormal activation or inhibition can influence the progression of airway remodeling by regulating the proliferation of airway smooth muscle cells, epithelial-mesenchymal transition (EMT), and extracellular matrix (ECM) deposition.28,42–44 Activation of the non-canonical Wnt signaling pathway promotes airway remodeling in COPD. Studies have shown that Wnt5a enhances the proliferation and migration of airway smooth muscle cells by activating the β-catenin pathway.43 In TGF-β1 induced airway smooth muscle cells, overexpression of Wnt5a significantly increased cell proliferation rate and migration capacity, whereas Wnt5a knockdown markedly reduced these parameters. Additionally, the activation of the Wnt pathway can facilitate EMT and contribute to airway fibrosis. In airway epithelial cells of COPD patients, the expression level of Wnt5a was positively correlated with the expression of EMT-related genes.45 Inhibition of the Wnt signaling pathway also inhibits the expression of matrix metalloproteinases (MMPs), degrades the ECM, and leads to airway remodeling damage.46 Research has demonstrated that miR-149-3p modulates the expression of Wnt1, Wnt5a, β-catenin, and RhoA, and alters the expression of alveolar inflammatory factors. These alterations subsequently influence the expression of alveolar inflammatory factors, thereby affecting the pathological progression of COPD.47–49 The canonical Wnt signaling pathway also plays an important role in airway remodellingin COPD. Studies have shown that nicotine exposure induces upregulation of Wnt3a and activation of the canonical Wnt signaling pathway, along with an increase incollagen type I, vimentin, and α-SMA, and downregulation of E-cadherin.50,51
Nc-RNAs and COPD
NC-RNA plays multiple roles in the pathological process of COPD, including airway inflammation, oxidative stress, epithelial injury, and airway remodeling. In this section, we summarize the roles of ncRNAs in COPD.
Classification and Biological Functions of Nc-RNAs
Of the diverse RNA families present within cells, long non-coding RNAs (lncRNAs, >200 nt), microRNAs (miRNAs, 20–24 nt), and circular RNAs (circRNAs, covalently closed loop structures) are categorized as nc-RNAs, a class which exerts significant regulatory functions in both fundamental physiological processes and the development of various pathological conditions in humans.52–55 miRNAs regulate gene expression by binding to the 3’-UTR of their target mRNA, thereby inhibiting translation or promoting degradation.54 LncRNAs exhibit a broad range of functions, including (1) acting as endogenous competing RNAs (ceRNAs) to sponge adsorbed miRNAs and regulate target gene expression; (2) binding to proteins to modulate their localization or activity; and (3) participating in chromatin remodeling.56,57 CircRNAs exhibit high stability and function primarily as ceRNAs.
Pathological Role of Nc-RNAs in COPD
Nc-RNAs play crucial regulatory roles in the pathological processes of COPD, with miRNAs and lncRNAs being the most extensively studied. miRNAs regulate gene expression by targeting the 3’-UTR region of mRNA, participating in inflammatory responses, airway remodeling, and apoptosis associated with COPD. Recent studies have confirmed that miRNAs are critical modulators of macrophage activity, demonstrating their capacity to both drive and protect against inflammatory responses.12 For instance, miR-21 promotes COPD pathogenesis by regulating the SATB1/S100A9/NF-κB signaling axis. Targeting miR-21 suppresses the inflammatory response and improves lung function.58 miRNAs derived from epithelial cell-exosomes, such as miR-125a-5p and miR-221-3p, promote M1 macrophage polarization.59,60 Other pro-inflammatory nc-RNAs like miR-486-5p, miR-27-3p, miR-1260, miR-155, and lncRNA MEG3, also regulate macrophages via different signaling pathways.12,61–63 In contrast, some miRNAs play a protective role in the inflammatory response and emphysema formation. For example, miR-195-5p, miR-146a, miR-513a-5p, miR-150, miR-344b-1-3p, Cox2, and IL7R inhibit COPD development by targeting NF-κB and TLR signaling.64–70 However, another study showed that M2 macrophage polarization might play a role in promoting rather than inhibiting COPD development by regulating mirR-let-7c.71 Interestingly, MIR155HG has the ability to regulate in two directions. Overexpression LncRNA MIR155HG promotes M1 macrophages polarization and pro-inflammatory cytokine release, whereas downregulation LncRNA MIR155HG enhances M2 macrophages development.72,73 nc-RNAs are a large family of transcripts. However, its role in COPD requires further investigation (Figure 3).
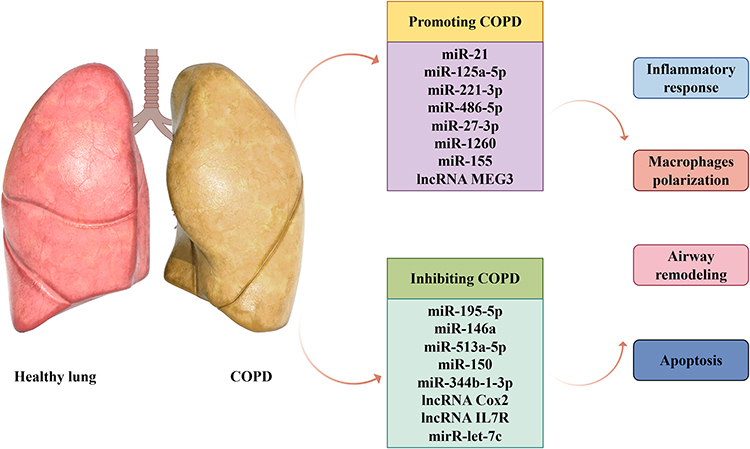 |
Figure 3 The biological function role of nc-RNAs in COPD. Nc-RNAs participated in the inflammatory responses, airway remodeling, and apoptosis associated with COPD, and nc-RNAs were recognized as critical modulators of macrophage activity, demonstrating a capacity for both driving and protecting against inflammatory responses. |
The Interaction Mechanisms Wnt Signaling Pathway and Nc-RNAs in COPD
As reported before, nc-RNAs can promote or inhibit disease development by regulating Wnt signaling pathway.21,74,75 However, the crosstalk between the Wnt signaling pathway and nc-RNAs remains unclear. Recent studies have reported that crosstalk between the Wnt signaling pathway and nc-RNAs plays an important role in COPD. In this section, we summarize the role of ncRNAs intertwined with the Wnt signaling pathway in COPD (Table 1).
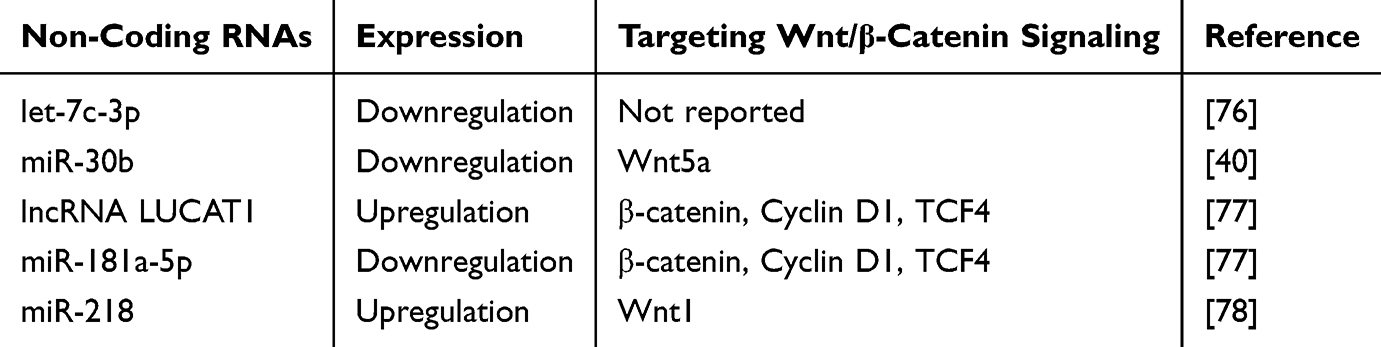 |
Table 1 Crosstalk of Non-Coding RNA and Wnt/β-Catenin Signaling Proteins in COPD |
Interaction between ncRNAs and the Wnt pathway is one of the core mechanisms regulating cell fate and disease progression. lncRNA, as a ceRNA, sponges miRNAs and indirectly regulates the expression of key molecules in the Wnt signaling pathway. The lncRNA GATA3-AS1 is highly expressed in pancreatic cancer, upregulating Tex10 expression by sponging miR-30b-5p and activating the Wnt signaling pathway to promote tumor proliferation.79 miRNAs directly target key components of the Wnt signaling pathway. Furthermore, circRNAs are involved in Wnt signaling pathway regulation via sponge adsorption of miRNAs.80 The interaction between ncRNAs and the Wnt signaling pathway also involves epigenetic regulation.81
It has been shown that nc-RNAs could inhibit COPD development by targeting Wnt signaling pathway. Previous studies have indicated that let-7c-3p plays a protective role in COPD by regulating PPAR/RXR, WNT/β-catenin, and pulmonary fibrosis pathways.76 However, the mechanisms by which let-7c-3p regulates the Wnt/β-catenin signaling pathway have not yet been reported. In patients with CSE-induced COPD, miR-30b was down-regulated.40 Overexpression of miR-30b reduces PMVEC apoptosis by targeting Wnt5a. This indicates that Wn5a plays an important role in COPD development.
Some ncRNAs can promote COPD development by targeting the Wnt signaling pathway. Studies have shown that the lncRNA LUCAT1 is upregulated in patients with COPD. Subsequent mechanistic studies indicated that knockdown of lncRNA LUCAT1 could reverse cell apoptosis by targeting the miR‐181a‐5p/Wnt/β‐catenin axis.77 In a CSE-induced mouse model, miR-130a was upregulated. Inhibition of miR-130a can rescue CSE-induced injury by targeting the Wnt signaling pathway.78
The above findings have confirmed the interaction between Wnt signaling pathway and some nc-RNAs. Advances in bioinformatics also provide new insights into their underlying interaction mechanisms. Resent bioinformatics studies have indicated that some nc-RNAs can regulate the Wnt signaling pathway participating in COPD pathogenesis. For instance, miR-218, miR-133a/b, miR-455-3p, miR-21-3p, miR-23a, miR-25, miR-145, and miR-224 have been reported to be differentially expressed in COPD. Gene set enrichment analysis (GSEA) and/or Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis showed that nc-RNAs might strongly target the Wnt signaling pathway.82–84 However, there is a lack of evidence to clarify their regulatory effect on the Wnt pathway and their specific involvement in COPD development. Further studies are required to confirm these results.
Therapeutic Strategies Targeting Nc-RNAs and Wnt Signaling Pathway in COPD
An increasing number of studies have confirmed that nc-RNAs and Wnt signaling pathways play a vital role in COPD pathogenesis. Therapeutic strategies targeting nc-RNAs and the Wnt signaling pathway may provide new insights into the treatment of COPD. Based on previous studies, a diverse array of pharmacological agents has been developed to modulate Wnt signaling.85–94 These pharmacological agents target different molecules in the Wnt signaling pathway. Some of the agents are listed in Table 2. Although many drugs target the Wnt signaling pathway, their use in COPD has seldom been documented. Previous studies have reported that GSK3 inhibitors can improve CS-induced lung injury and inflammation by activating the Wnt pathway.95–98 It should be noted that activation of the Wnt signaling pathway may lead to the development of tumors or other diseases. Therefore, side effects of pharmacological agents should also be considered, and targeting nc-RNAs may serve as a more promising therapeutic strategy.
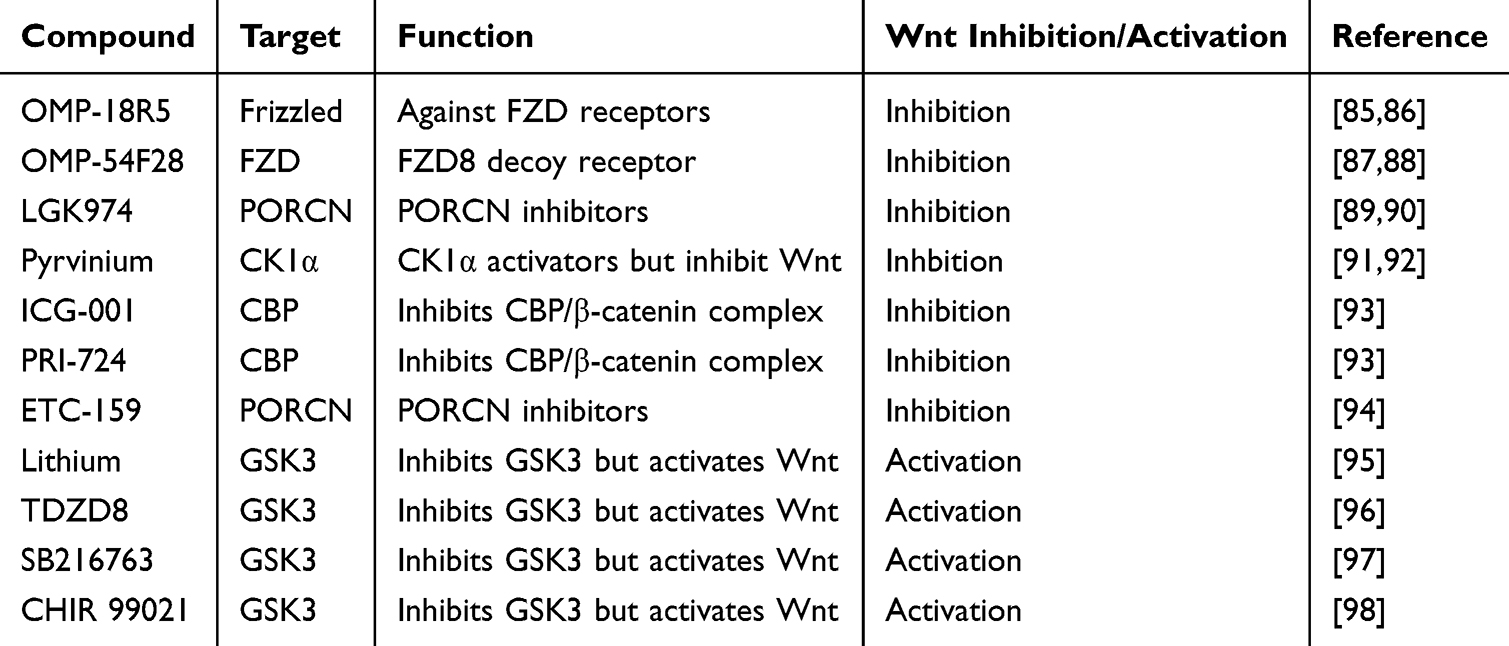 |
Table 2 Pharmacological Agents Target Wnt Signaling Pathway |
In recent years, therapeutic strategies targeting nc-RNAs, particularly miRNAs, have attracted significant attention. High-throughput sequencing technology and bioinformatics analysis have confirmed that an increasing number of miRNAs have been identified in COPD. Targeted miRNA therapy has also demonstrated efficacy. For instance, miR-3202, miR-145-5p, and miR-181c are downregulated in COPD, and upregulation of these miRNAs could improve COPD development via anti-inflammatory and anti-apoptosis.99–101 In addition, miR-206 and miR-27-3p are upregulated in COPD, and downregulation of these miRNAs improves COPD development via anti-inflammatory and anti-apoptotic effects.61,102 However, these therapeutic strategies remain at the stage of animal and cell experiments, and remain a significant challenge for clinical applications.
Challenges and Future Directions of Nc-RNAs and Wnt Signaling Pathway in COPD
Although the roles of the Wnt signaling pathway and nc-RNAs in COPD have been extensively studied, some challenges remain. First, the role of the Wnt signaling pathway in COPD remains to be investigated. Some studies suggest that inhibition of the Wnt signaling pathway is a key mechanism in COPD, whereas other studies indicate that its activation may promote airway remodeling.30–32 These differences may be related to the choice of subjects, severity of COPD, and different detection methods. Second, the mechanism of the Wnt signaling pathway in COPD remains controversial. Some studies have suggested that activation of the Wnt signaling pathway may suppress inflammatory responses and apoptosis in COPD, whereas other studies have demonstrated that activation of this pathway can promote airway remodeling in COPD.30,36,46 The inconsistent research conclusions may be attributed to multiple confounding factors. First, the activation level and duration of Wnt signaling vary significantly across different stages of COPD. Wnt signaling exerts a protective effect in the early compensatory repair stage of lung injury, while persistent overactivation in the advanced stage promotes pathological remodeling of lung tissue. Second, the functional differences of Wnt signaling are cell-type specific. Wnt pathway activation plays distinct roles in alveolar epithelial cells, airway smooth muscle cells and inflammatory immune cells, leading to opposite biological effects. In addition, differences in animal models, intervention methods, and detection indicators in existing studies also contribute to conflicting experimental results. Despite the current progress, there remain considerable knowledge gaps in clarifying the ncRNA-Wnt regulatory network in COPD. The specific upstream and downstream regulatory mechanisms of different ncRNA subtypes targeting the Wnt pathway in COPD progression have not been fully elucidated. Moreover, the dynamic changes and precise threshold of Wnt signaling activation mediated by ncRNAs at different disease stages remain unclear. Further studies are required to resolve existing controversies, clarify the spatiotemporal regulatory pattern of the ncRNA-Wnt axis, and provide more accurate and targeted theoretical basis for the clinical treatment of COPD. The resolution of these points depends on multi-center, large-sample clinical studies and rigorous experimental design.
Emerging technologies, such as artificial intelligence (AI), machine learning, single-cell RNA sequencing, spatial transcriptomics, CRISPR-Cas9 technology, and synthetic biology, have provided new directions for the study of nc-RNAs and Wnt signaling pathways in COPD. AI can be utilized for predicting the regulatory networks of ncRNA and Wnt pathways, machine learning can be employed for screening biomarkers of COPD, single-cell RNA sequencing can reveal changes in the Wnt signaling pathway and nc-RNA expression across different cell types in the lung tissue of COPD patients, spatial transcriptomics can elucidate the spatial distribution of the Wnt signaling pathway and nc-RNAs in lung tissue, CRISPR-Cas9 technology can be utilized for functional validation of ncRNA, and synthetic biology can be utilized in the design of ncRNA-targeted therapies. Additionally, exosomes, as carriers of ncRNAs, deserve attention for their potential applications in COPD treatment.103–107 Additionally, integrated multi-omics analyses, such as a combination of genomics, transcriptomics, proteomics, and metabolomics, can comprehensively elucidate the regulatory networks of ncRNAs and Wnt pathways in COPD. The advancement of these research directions is expected to drive breakthroughs in the study of ncRNAs and Wnt pathways, thereby providing novel strategies for the precise diagnosis and treatment of COPD.
Conclusion
In summary, the Wnt signaling pathway acts as a core regulatory axis participating in the entire pathological process of COPD, including airway inflammation, airway remodeling, and apoptosis. Aberrant activation or inhibition of Wnt signaling exerts dual, stage-dependent biological effects on COPD progression, which explains the inconsistent research conclusions in current studies. As critical regulatory molecules, nc-RNAs including miRNAs, lncRNAs, and circRNAs can precisely modulate the activation status of the Wnt pathway by targeting its key molecules, thereby mediating the occurrence and development of COPD. The interactive regulatory network between nc-RNAs and Wnt signaling provides a novel molecular mechanism for exploring the pathological basis of COPD.
Data Sharing Statement
All data generated or analyzed in this study are included in this published article.
Acknowledgment
We acknowledge the Figdraw (www.figdraw.com) as the figures were drawn by Figdraw.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research received no external funding.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Chen J, Zhang X, Sun G. Identification and validation of biomarkers related to mitophagy in chronic obstructive pulmonary disease. Mol Med Rep. 2025;31(4):93. doi:10.3892/mmr.2025.13458
2. Ding H, Wang H, Wu D, et al. Serum metabolomics analysis of patients with chronic obstructive pulmonary disease and ‘frequent exacerbator’ phenotype. Mol Med Rep. 2024;30(2):137. doi:10.3892/mmr.2024.13261
3. MH Stanikzai, M Nassar, S Sankar, et al. GBD CODC: Global burden of 292 causes of death in 204 countries and territories and 660 subnational locations, 1990-2023: a systematic analysis for the Global Burden of Disease Study 2023. Lancet. 2025;406(10513):1811–12. doi:10.1016/S0140-6736(25)01917-8
4. Blanco I, Diego I, Bueno P, Casas-Maldonado F, Miravitlles M. Geographic distribution of COPD prevalence in the world displayed by geographic information system maps. Europ Resp J. 2019;54(1):1900610. doi:10.1183/13993003.00610-2019
5. Wang C, Xu J, Yang L, et al. Prevalence and risk factors of chronic obstructive pulmonary disease in China (the China Pulmonary Health [CPH] study): a national cross-sectional study. Lancet. 2018;391(10131):1706–1717. doi:10.1016/S0140-6736(18)30841-9
6. Werder RB, Liu T, Abo KM, et al. CRISPR interference interrogation of COPD GWAS genes reveals the functional significance of desmoplakin in iPSC-derived alveolar epithelial cells. Sci Adv. 2022;8(28):eabo6566. doi:10.1126/sciadv.abo6566
7. Zhao X, Qiao D, Yang C, et al. Whole genome sequence analysis of pulmonary function and COPD in 19,996 multi-ethnic participants. Nat Commun. 2020;11(1):5182. doi:10.1038/s41467-020-18334-7
8. Nakamura H. Genetics of COPD. Allergol Int. 2011;60(3):253–258. doi:10.2332/allergolint.11-RAI-0326
9. Silverman EK. Genetics of COPD. Annu Rev Physiol. 2020;82:413–431. doi:10.1146/annurev-physiol-021317-121224
10. Ramdani I, Pescatore KA, Bouazza B. Causes of hospitalization and characteristics of Algerian chronic obstructive pulmonary disease patients in Tizi-Ouzou: a retrospective study. Monaldi Arch Chest Dis. 2020;90(3):10–4081.
11. Li P, Han X, Li J, et al. Aerobic exercise training engages the canonical wnt pathway to improve pulmonary function and inflammation in COPD. BMC Pulm Med. 2024;24(1):236. doi:10.1186/s12890-024-03048-z
12. Yong X, Luo X, Chen X, Yu C. Orchestrating inflammation: non-coding RNAs as master regulators of macrophage function in chronic obstructive pulmonary disease-an update. Front Immunol. 2025;16:1679730. doi:10.3389/fimmu.2025.1679730
13. Kara M, Kirkil G, Kalemci S. Differential expression of MicroRNAs in chronic obstructive pulmonary disease. Adv Clin Exp Med. 2016;25(1):21–26. doi:10.17219/acem/28343
14. Gu W, Yuan Y, Wang L, et al. Long non-coding RNA TUG1 promotes airway remodelling by suppressing the miR-145-5p/DUSP6 axis in cigarette smoke-induced COPD. J Cell Mol Med. 2019;23(11):7200–7209. doi:10.1111/jcmm.14389
15. Liu X, Ma Y, Zong D, Chen Y. LncRNA MALAT1 regulates cigarette smoke induced airway inflammation by modulating miR-30a-5p/JNK signaling pathway. Int Immunopharmacol. 2024;140:112826. doi:10.1016/j.intimp.2024.112826
16. Ge Z, Li B, Zhou X, Yang Y, Zhang J. Basic fibroblast growth factor activates β-catenin/RhoA signaling in pulmonary fibroblasts with chronic obstructive pulmonary disease in rats. Mol Cell Biochem. 2016;423(1–2):165–174. doi:10.1007/s11010-016-2834-7
17. Baarsma HA, Skronska-Wasek W, Mutze K, et al. Noncanonical WNT-5A signaling impairs endogenous lung repair in COPD. J Exp Med. 2017;214(1):143–163. doi:10.1084/jem.20160675
18. Tian Y, Liu Y, Xiao Y, et al. Alternative biological material for tissue engineering of the vagina: porcine-derived acellular vaginal matrix. Tissue Eng Regen Med. 2024;21(2):277–290. doi:10.1007/s13770-023-00604-2
19. Liu J, Xiao Q, Xiao J, et al. Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduction Tar. 2022;7(1):3.
20. Wang H, Zhang L, Hu C, Li H, Jiang M. Wnt signaling and tumors (Review). Mol Clin Oncol. 2024;21(1):45. doi:10.3892/mco.2024.2743
21. Tian Y, Lai T, Li Z, et al. Role of non‑coding RNA intertwined with the Wnt/β‑catenin signaling pathway in endometrial cancer (Review). Mol Med Rep. 2023;28(2):150. doi:10.3892/mmr.2023.13037
22. Xie B, Cao Y, Li W, Chen R, Zhou Q, Xie X. Nuclear β-catenin: molecular regulation, challenges and therapeutic opportunities. Cell Commun Signal. 2025;2025:10–1186.
23. Alam S, Duncan D, Hasan S. Profilin and non-canonical wnt signaling: coordinating cytoskeletal dynamics from development to disease. J Dev Biol. 2025;13(3):31. doi:10.3390/jdb13030031
24. Li P, Ma X, Huang D. Role of the lncRNA/Wnt signaling pathway in digestive system cancer: a literature review. Eur J Med Res. 2024;29(1):447. doi:10.1186/s40001-024-02033-w
25. Gou L, Ren X, Ji P. Canonical Wnt signaling regulates branching morphogenesis of submandibular gland by modulating levels of lama5. Int J Dev Biol. 2021;65(7–8–9):497–504. doi:10.1387/ijdb.200307lg
26. Liang J, Pan Y, Yang J, Zeng D, Li J. WNT signaling in cancer: molecular mechanisms and potential therapies. Mol Biomed. 2025;6(1):83. doi:10.1186/s43556-025-00327-x
27. Feng L, Fan X, Shao L, et al. Wnt5a deficiency in osteocalcin-expressing cells could not alleviate the osteoarthritic phenotype in a mouse model of post-traumatic osteoarthritis. Iran J Basic Med Sci. 2024;27(6):671–677. doi:10.22038/IJBMS.2024.71417.15527
28. Liu M, Huo Y, Cheng Y. Mechanistic regulation of wnt pathway-related progression of chronic obstructive pulmonary disease airway lesions. Int J Chronic Obstr. 2023;18:871–880.
29. Eapen MS, Sohal SS. WNT/β-catenin pathway: a novel therapeutic target for attenuating airway remodelling and EMT in COPD. Ebiomedicine. 2020;62:103095. doi:10.1016/j.ebiom.2020.103095
30. Cui W, Zhang Z, Zhang P, et al. Nrf2 attenuates inflammatory response in COPD/emphysema: crosstalk with Wnt3a/β-catenin and AMPK pathways. J Cell Mol Med. 2018;22(7):3514–3525. doi:10.1111/jcmm.13628
31. Heijink IH, de Bruin HG, van den Berge M, et al. Role of aberrant WNT signalling in the airway epithelial response to cigarette smoke in chronic obstructive pulmonary disease. Thorax. 2013;68(8):709–716. doi:10.1136/thoraxjnl-2012-201667
32. Durham AL, McLaren A, Hayes BP, et al. Regulation of Wnt4 in chronic obstructive pulmonary disease. FASEB J. 2013;27(6):2367–2381. doi:10.1096/fj.12-217083
33. van Dijk EM, Menzen MH, Spanjer AIR, Middag LDC, Brandsma CA, Gosens R. Noncanonical WNT-5B signaling induces inflammatory responses in human lung fibroblasts. Am J Physiol Lung Cell Mol Physiol. 2016;310(11):L1166–L1176. doi:10.1152/ajplung.00226.2015
34. Wu X, van Dijk EM, Ng-Blichfeldt J, et al. Mesenchymal WNT-5A/5B signaling represses lung alveolar epithelial progenitors. Cells Basel. 2019;8(10):1147. doi:10.3390/cells8101147
35. Zhou M, Jiao L, Liu Y. sFRP2 promotes airway inflammation and Th17/Treg imbalance in COPD via Wnt/β-catenin pathway. Resp Physiol Neurob. 2019;270:103282. doi:10.1016/j.resp.2019.103282
36. Ma R, Su H, Jiao K, Liu J. Role of Th17 cells, Treg cells, and Th17/Treg imbalance in immune homeostasis disorders in patients with chronic obstructive pulmonary disease. Immun Inflamm Dis. 2023;11(2):e784. doi:10.1002/iid3.784
37. Zheng X, Zhang L, Chen J, Gu Y, Xu J, Ouyang Y. Dendritic cells and Th17/Treg ratio play critical roles in pathogenic process of chronic obstructive pulmonary disease. Biomed Pharmacothe. 2018;108:1141–1151. doi:10.1016/j.biopha.2018.09.113
38. Xu H, Zhan F, Wang H, Lin J, Niu H. Down-regulation of RTEL1 improves M1/M2 macrophage polarization by promoting SFRP2 in fibroblasts-derived exosomes to alleviate COPD. Cell Biochem Biophys. 2024;82(3):2129–2139. doi:10.1007/s12013-024-01320-x
39. Chen L, Tang R, Ruan J, Zhu X, Yang Y. Up-regulation of THY1 attenuates interstitial pulmonary fibrosis and promotes lung fibroblast apoptosis during acute interstitial pneumonia by blockade of the WNT signaling pathway. Cell Cycle. 2019;18(6–7):670–681. doi:10.1080/15384101.2019.1578144
40. Song Q, Zhou A, Cheng W, et al. Bone marrow mesenchymal stem cells-derived exosomes inhibit apoptosis of pulmonary microvascular endothelial cells in COPD mice through miR-30b/Wnt5a pathway. Int J Nanomed. 2025;20:1191–1211. doi:10.2147/IJN.S487097
41. Vuga LJ, Ben-Yehudah A, Kovkarova-Naumovski E, et al. WNT5A is a regulator of fibroblast proliferation and resistance to apoptosis. Am J Resp Cell Mol. 2009;41(5):583–589. doi:10.1165/rcmb.2008-0201OC
42. Zhou H, Lai Y, Zhu Y, et al. Quercetin improves airway remodeling in COPD rats by suppressing phenotypic switch of ASMCs via inhibiting the Wnt5a/β-catenin pathway. Phytomedicine. 2025;139:156491. doi:10.1016/j.phymed.2025.156491
43. Zou W, Wang X, Sun R, et al. PM2.5 induces airway remodeling in chronic obstructive pulmonary diseases via the Wnt5a/β-Catenin pathway. Int J Chronic Obstr. 2021;16:3285–3295.
44. Su X, Chen J, Lin X, et al. FERMT3 mediates cigarette smoke-induced epithelial-mesenchymal transition through Wnt/β-catenin signaling. Resp Res. 2021;22(1):286. doi:10.1186/s12931-021-01881-y
45. Ng-Blichfeldt J, de Jong T, Kortekaas RK, et al. TGF-β activation impairs fibroblast ability to support adult lung epithelial progenitor cell organoid formation. Am J Physiol Lung Cell Mol Physiol. 2019;317(1):L14–L28. doi:10.1152/ajplung.00400.2018
46. Ge Z, Yang Y, Zhou X, et al. Overexpression of the hyperplasia suppressor gene inactivates airway fibroblasts obtained from a rat model of chronic obstructive pulmonary disease by inhibiting the Wnt signaling pathway. Mol Med Rep. 2019;20(3):2754–2762. doi:10.3892/mmr.2019.10504
47. Naskar D, Maiti G, Chakraborty A, Roy A, Chattopadhyay D, Sen M. Wnt5a-Rac1-NF-κB homeostatic circuitry sustains innate immune functions in macrophages. J Immunol. 2014;192(9):4386–4397. doi:10.4049/jimmunol.1302817
48. Neumann J, Schaale K, Farhat K, et al. Frizzled1 is a marker of inflammatory macrophages, and its ligand Wnt3a is involved in reprogramming Mycobacterium tuberculosis-infected macrophages. FASEB J. 2010;24(11):4599–4612. doi:10.1096/fj.10-160994
49. Zhang X, Wang Y, He X, Sun Z, Shi X. Diagnosis of chronic obstructive pulmonary disease and regulatory mechanism of miR-149-3p on alveolar inflammatory factors and expression of surfactant proteins A (SP-A) and D (SP-D) on lung surface mediated by wnt pathway. Comput Intel Neurosc. 2022;2022:7205016.
50. Krebs M, Sakurai R, Torday JS, Rehan VK. Evidence for in vivo nicotine-induced alveolar interstitial fibroblast-to-myofibroblast transdifferentiation. Exp Lung Res. 2010;36(7):390–398. doi:10.3109/01902141003714023
51. Sakurai R, Cerny LM, Torday JS, Rehan VK. Mechanism for nicotine-induced up-regulation of Wnt signaling in human alveolar interstitial fibroblasts. Exp Lung Res. 2011;37(3):144–154. doi:10.3109/01902148.2010.490288
52. Liao Y, Cao W, Zhang K, et al. Bioinformatic and integrated analysis identifies an lncRNA-miRNA-mRNA interaction mechanism in gastric adenocarcinoma. Genes Genom. 2021;43(6):613–622. doi:10.1007/s13258-021-01086-z
53. Huang Y, Liu G, Ma H, et al. Plasma lncRNA FEZF1-AS1 as a potential biomarker for diagnosis of non-small-cell lung carcinoma. Medicine. 2020;99(26):e21019. doi:10.1097/MD.0000000000021019
54. Nan F, Liu B, Yao C. Discovering the role of microRNAs and exosomal microRNAs in chest and pulmonary diseases: a spotlight on chronic obstructive pulmonary disease. Mol Genet Genom. 2024;299(1):107. doi:10.1007/s00438-024-02199-2
55. Markelov VA, Korytina GF, Aznabaeva YG, et al. Expression of long noncoding RNAs and protein-coding genes involved in oxidative stress and cell senescence in patients with chronic obstructive pulmonary disease. Mol Biol. 2024;58(5):821–839. doi:10.31857/S0026898424050119
56. Lin Q, Zhang C, Weng H, Lin Y, Lin Y, Ruan Z. The utility of long non-coding RNAs in chronic obstructive pulmonary disease: a comprehensive analysis. BMC Pulm Med. 2023;23(1):340. doi:10.1186/s12890-023-02635-w
57. De Smet EG, Mestdagh P, Vandesompele J, Brusselle GG, Bracke KR. Non-coding RNAs in the pathogenesis of COPD. Thorax. 2015;70(8):782–791. doi:10.1136/thoraxjnl-2014-206560
58. Kim RY, Sunkara KP, Bracke KR, et al. A microRNA-21-mediated SATB1/S100A9/NF-κB axis promotes chronic obstructive pulmonary disease pathogenesis. Sci Transl Med. 2021;13(621):eaav7223. doi:10.1126/scitranslmed.aav7223
59. Wang R, Zhu Z, Peng S, et al. Exosome microRNA-125a-5p derived from epithelium promotes M1 macrophage polarization by targeting IL1RN in chronic obstructive pulmonary disease. Int Immunopharmacol. 2024;137:112466. doi:10.1016/j.intimp.2024.112466
60. Jia H, He W, Wu B, et al. Cigarette smoke-induced exosomal miR-221-3p facilitates M1 macrophage polarization via the STAT3 pathway in chronic obstructive pulmonary disease. Aging. 2024;16(17):12379–12391. doi:10.18632/aging.206095
61. Wang D, He S, Liu B, Liu C. MiR-27-3p regulates TLR2/4-dependent mouse alveolar macrophage activation by targetting PPARγ. Clin Sci. 2018;132(9):943–958. doi:10.1042/CS20180083
62. Yang X, Zeng X, Shu J, Bao H, Liu X. MiR-155 enhances phagocytosis of alveolar macrophages through the mTORC2/RhoA pathway. Medicine. 2023;102(35):e34592. doi:10.1097/MD.0000000000034592
63. Zhang J, Xu Z, Kong L, Gao H, Zhang Y, Zheng Y. Wan Y: miRNA-486-5p promotes COPD progression by targeting HAT1 to regulate the TLR4-triggered inflammatory response of alveolar macrophages. Int J Chronic Obstr. 2020;15:2991–3001.
64. Li S, Jiang L, Yang Y, et al. MiR-195-5p inhibits the development of chronic obstructive pulmonary disease via targeting siglec1. Hum Exp Toxicol. 2020;39(10):1333–1344. doi:10.1177/0960327120920923
65. Yoshikawa H, Sato T, Horikoshi K, et al. miR-146a regulates emphysema formation and abnormal inflammation in the lungs of two mouse models. Am J Physiol Lung Cell Mol Physiol. 2024;326(1):L98–L110. doi:10.1152/ajplung.00080.2023
66. Zhou J, Du J, Xu R, Wu X, Zhang G. Reduced miR-513a-5p expression in COPD may regulate airway mucous cell hyperplasia through TFR1-dependent signaling. Kaohsiung J Med Sci. 2024;40(2):139–149. doi:10.1002/kjm2.12777
67. Xu H, Wu Y, Li L, et al. MiR-344b-1-3p targets TLR2 and negatively regulates TLR2 signaling pathway. Int J Chronic Obstr. 2017;12:627–638.
68. Xue H, Li MX. MicroRNA-150 protects against cigarette smoke-induced lung inflammation and airway epithelial cell apoptosis through repressing p53: microRNA-150 in CS-induced lung inflammation. Hum Exp Toxicol. 2018;37(9):920–928. doi:10.1177/0960327117741749
69. Salih MM, Robinson EK, Malekos E, et al. LincRNA-Cox2 regulates smoke-induced inflammation in murine macrophages. Am J Resp Cell Mol. 2023;68(5):511–522. doi:10.1165/rcmb.2022-0413OC
70. Wu S, Feng P, Chuang H, et al. Impaired lnc-IL7R modulatory mechanism of Toll-like receptors is associated with an exacerbator phenotype of chronic obstructive pulmonary disease. FASEB J. 2020;34(10):13317–13332. doi:10.1096/fj.202000632R
71. Liu T, Zhang Z, Shen W, Wu Y, Bian T. MicroRNA Let-7 induces M2 macrophage polarization in COPD emphysema through the IL-6/STAT3 pathway. Int J Chronic Obstr. 2023;18:575–591.
72. Li N, Liu Y, Cai J. LncRNA MIR155HG regulates M1/M2 macrophage polarization in chronic obstructive pulmonary disease. Biomed Pharmacothe. 2019;117:109015. doi:10.1016/j.biopha.2019.109015
73. Song J, Wang Q, Zong L. LncRNA MIR155HG contributes to smoke-related chronic obstructive pulmonary disease by targeting miR-128-5p/BRD4 axis. Biosci Rep. 2020;40(3):BSR20192567. doi:10.1042/BSR20192567
74. Zhang Y, Sun X, Li Z, et al. Interactions between miRNAs and the Wnt/β-catenin signaling pathway in endometriosis. Biomed Pharmacothe. 2024;171:116182. doi:10.1016/j.biopha.2024.116182
75. Zhong Y, He J, Huang C, et al. The NcRNA/Wnt axis in lung cancer: oncogenic mechanisms, remarkable indicators and therapeutic targets. J Transl Med. 2025;23(1):326. doi:10.1186/s12967-025-06326-4
76. Liu Y, Xu G, Li L. LncRNA GATA3‑AS1‑miR‑30b‑5p‑Tex10 axis modulates tumorigenesis in pancreatic cancer. Oncol Rep. 2021;45(5):59. doi:10.3892/or.2021.8010
77. Khalaji A, Nazari Y, Pandeh M, et al. Regulatory roles of circular RNAs in Wnt and other oncogenic signaling pathways in breast cancer progression: a comprehensive review. Eur J Med Res. 2025;30(1):750. doi:10.1186/s40001-025-02967-9
78. Wu Y, Li H, Long Y, et al. Epigenetic Suppression of miR-137 Induces RNF4 Expression, Facilitating Wnt Signaling in Colorectal Cancer. Mol Carcinog. 2025;64(3):475–489. doi:10.1002/mc.23859
79. Hsieh M, Lai P, Chen P, et al. Exosomal MicroRNA let-7 modulates lipid metabolism and inflammation in foamy macrophages of chronic obstructive pulmonary disease. Int J Mol Sci. 2025;26(18):8800. doi:10.3390/ijms26188800
80. Zhao S, Lin C, Yang T, Qian X, Lu J, Cheng J. Expression of long non-coding RNA LUCAT1 in patients with chronic obstructive pulmonary disease and its potential functions in regulating cigarette smoke extract-induced 16HBE cell proliferation and apoptosis. J Clin Lab Anal. 2021;35(7):e23823. doi:10.1002/jcla.23823
81. Wu Y, Guan S, Ge Y, Yang Y, Cao Y, Zhou J. Cigarette smoke promotes chronic obstructive pulmonary disease (COPD) through the miR-130a/Wnt1 axis. Toxicol vitro. 2020;65:104770. doi:10.1016/j.tiv.2020.104770
82. Wang G, Wang R, Strulovici-Barel Y, et al. Persistence of smoking-induced dysregulation of miRNA expression in the small airway epithelium despite smoking cessation. PLoS One. 2015;10(4):e120824.
83. Ong J, Timens W, Rajendran V, et al. Identification of transforming growth factor-beta-regulated microRNAs and the microRNA-targetomes in primary lung fibroblasts. PLoS One. 2017;12(9):e183815. doi:10.1371/journal.pone.0183815
84. Liu X, Qu J, Xue W, et al. Bioinformatics-based identification of potential microRNA biomarkers in frequent and non-frequent exacerbators of COPD. Int J Chronic Obstr. 2018;13:1217–1228.
85. Gurney A, Axelrod F, Bond CJ, et al. Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc Natl Acad Sci U S A. 2012;109(29):11717–11722. doi:10.1073/pnas.1120068109
86. Davis SL, Cardin DB, Shahda S, et al. A Phase 1b dose escalation study of Wnt pathway inhibitor vantictumab in combination with nab-paclitaxel and gemcitabine in patients with previously untreated metastatic pancreatic cancer. Invest New Drug. 2020;38(3):821–830. doi:10.1007/s10637-019-00824-1
87. Jimeno A, Gordon M, Chugh R, et al. A first-in-human Phase I study of the anticancer stem cell agent Ipafricept (OMP-54F28), a decoy receptor for Wnt Ligands, in patients with advanced solid tumors. Clin Cancer Res. 2017;23(24):7490–7497. doi:10.1158/1078-0432.CCR-17-2157
88. Moore KN, Gunderson CC, Sabbatini P, et al. A phase 1b dose escalation study of ipafricept (OMP54F28) in combination with paclitaxel and carboplatin in patients with recurrent platinum-sensitive ovarian cancer. Gynecol Oncol. 2019;154(2):294–301. doi:10.1016/j.ygyno.2019.04.001
89. Rodon J, Argilés G, Connolly RM, et al. Phase 1 study of single-agent WNT974, a first-in-class Porcupine inhibitor, in patients with advanced solid tumours. Brit J Cancer. 2021;125(1):28–37. doi:10.1038/s41416-021-01389-8
90. Tabernero J, Van Cutsem E, Garralda E, et al. A Phase Ib/II study of WNT974 + Encorafenib + cetuximab in patients with BRAF -mutant KRAS wild-type metastatic colorectal cancer. Oncologist. 2023;28(3):230–238.
91. Thorne CA, Hanson AJ, Schneider J, et al. Small-molecule inhibition of Wnt signaling through activation of casein kinase 1α. Nat Chem Biol. 2010;6(11):829–836. doi:10.1038/nchembio.453
92. Li B, Flaveny CA, Giambelli C, et al. Repurposing the FDA-approved pinworm drug pyrvinium as a novel chemotherapeutic agent for intestinal polyposis. PLoS One. 2014;9(7):e101969. doi:10.1371/journal.pone.0101969
93. Kimura K, Ikoma A, Shibakawa M, et al. Safety, tolerability, and preliminary efficacy of the anti-fibrotic small molecule PRI-724, a CBP/β-Catenin inhibitor, in patients with Hepatitis C virus-related cirrhosis: a single-center, open-label, dose escalation Phase 1 trial. Ebiomedicine. 2017;23:79–87. doi:10.1016/j.ebiom.2017.08.016
94. He B, You L, Uematsu K, et al. A monoclonal antibody against Wnt-1 induces apoptosis in human cancer cells. Neoplasia. 2004;6(1):7–14. doi:10.1016/S1476-5586(04)80048-4
95. Hedgepeth CM, Conrad LJ, Zhang J, Huang HC, Lee VM, Klein PS. Activation of the Wnt signaling pathway: a molecular mechanism for lithium action. Dev Biol. 1997;185(1):82–91. doi:10.1006/dbio.1997.8552
96. Eldar-Finkelman H, Martinez A. GSK-3 inhibitors: preclinical and clinical focus on CNS. Front Mol Neurosci. 2011;4:32. doi:10.3389/fnmol.2011.00032
97. Baarsma HA, Meurs H, Halayko AJ, et al. Glycogen synthase kinase-3 regulates cigarette smoke extract- and IL-1β-induced cytokine secretion by airway smooth muscle. Am J Physiol Lung Cell Mol Physiol. 2011;300(6):L910–L919. doi:10.1152/ajplung.00232.2010
98. Uhl FE, Vierkotten S, Wagner DE, et al. Preclinical validation and imaging of Wnt-induced repair in human 3D lung tissue cultures. Europ Resp J. 2015;46(4):1150–1166. doi:10.1183/09031936.00183214
99. Shen W, Liu J, Fan M, et al. MiR-3202 protects smokers from chronic obstructive pulmonary disease through inhibiting FAIM2: an in vivo and in vitro study. Exp Cell Res. 2018;362(2):370–377. doi:10.1016/j.yexcr.2017.11.038
100. Dang X, Yang L, Guo J, Hu H, Li F, Liu Y. Pang Y: miR-145-5p is associated with smoke-related chronic obstructive pulmonary disease via targeting KLF5. Chem Biol Interact. 2019;300:82–90. doi:10.1016/j.cbi.2019.01.011
101. Du Y, Ding Y, Chen X, et al. MicroRNA-181c inhibits cigarette smoke-induced chronic obstructive pulmonary disease by regulating CCN1 expression. Resp Res. 2017;18(1):155. doi:10.1186/s12931-017-0639-1
102. Sun Y, An N, Li J, et al. Zhang X: miRNA-206 regulates human pulmonary microvascular endothelial cell apoptosis via targeting in chronic obstructive pulmonary disease. J Cell Biochem. 2019;120(4):6223–6236. doi:10.1002/jcb.27910
103. Reid LV, Spalluto CM, Watson A, Staples KJ, Wilkinson TMA. The role of extracellular vesicles as a shared disease mechanism contributing to multimorbidity in patients with COPD. Front Immunol. 2021;12:754004. doi:10.3389/fimmu.2021.754004
104. Lv J, Xiong X. Extracellular vesicle microRNA: a promising biomarker and therapeutic target for respiratory diseases. Int J Mol Sci. 2024;25(17):9147. doi:10.3390/ijms25179147
105. Soccio P, Moriondo G, Lacedonia D, et al. EVs-miRNA: the new molecular markers for chronic respiratory diseases. Life. 2022;12(10):1544. doi:10.3390/life12101544
106. Wang H, Zhong Y, Li N, et al. Transcriptomic analysis and validation reveal the pathogenesis and a novel biomarker of acute exacerbation of chronic obstructive pulmonary disease. Resp Res. 2022;23(1):27. doi:10.1186/s12931-022-01950-w
107. Yang H, Wu Y, Wu T, Ji J, Lei S, Xu W. Accuracy of deep learning in diagnosing chronic obstructive pulmonary disease: systematic review and meta-analysis. J Med Internet Res. 2026;28:e83459. doi:10.2196/83459
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Mechanistic Regulation of Wnt Pathway-Related Progression of Chronic Obstructive Pulmonary Disease Airway Lesions
Liu M, Huo Y, Cheng Y
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:871-880
Published Date: 15 May 2023