Back to Journals » International Journal of Women's Health » Volume 17
Exploring Gut Microbe-Host Genes in Postpartum Depression: Mendelian Randomization and Transcriptomic Analysis
Authors Cong T, Liu J, Yuan L, Chu Z ![]()
Received 18 April 2025
Accepted for publication 23 October 2025
Published 27 November 2025 Volume 2025:17 Pages 4981—4994
DOI https://doi.org/10.2147/IJWH.S535357
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Matteo Frigerio
Ting Cong,1,2,* Jing Liu,3,* Li Yuan,1,2 Zhenchen Chu4
1Department of Anesthesiology, The Affiliated Hospital of Qingdao University, Qingdao, Shandong, 266000, People’s Republic of China; 2Shandong Provincial Key Medical and Health Laboratory of Anesthesia and Brain Function, Qingdao, Shandong, 266000, People’s Republic of China; 3Department of Psychiatry, Dalian Seventh People’s Hospital, Dalian, Liaoning, 116023, People’s Republic of China; 4Department of Orthopedics, Qingdao Municipal Hospital, Qingdao, Shandong, 266011, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Li Yuan, Department of Anesthesiology, The Affiliated Hospital of Qingdao University, 16 Jiangsu Road, Shinan District, Qingdao, Shandong, 266000, People’s Republic of China, Email [email protected] Zhenchen Chu, Department of Orthopedics, Qingdao Municipal Hospital, No. 5 Donghai Middle Road, Shinan District, Qingdao, Shandong, 266011, People’s Republic of China, Email [email protected]
Purpose: To suggest a potential causal role of gut microbiota in Postpartum depression (PPD) and identify susceptible microbiota-host genes.
Patients and Methods: The two-sample Mendelian randomization (MR) study was carried out with the genome-wide association studies (GWAS) data of 196 gut microbial taxa and PPD. Causal relationships were evaluated with inverse variance weighted (IVW), MR‒Egger, weighted mode, and weighted median approaches. Cochran’s Q test, the MR‒Egger intercept test, the MR-PRESSO test, and leave-one-out analysis were performed for assessing pleiotropy and heterogeneity. Additionally, false discovery rate (FDR) correction was completed via q-value method. A microarray dataset was carried out to identify susceptible microbiota-host genes.
Results: IVW suggested that the family Veillonellaceae [odds ratio (OR) = 0.82, 95% confidence interval (CI) = 0.71– 0.94, p = 0.004, q = 0.030] decreased the PPD risk, but the class Alphaproteobacteria (OR = 1.22, 95% CI = 1.01– 1.47, p = 0.041, q = 0.074) and the genus Family XIII AD3011 group (OR = 1.24, 95% CI = 1.04– 1.48, p = 0.019, q = 0.065) increased the incidence of PPD. Additionally, we extracted SNP-related genes from the Family XIII AD3011 group and identified four gut microbe-host genes (AQP9, ALDH1A2, DGUOK, and STAMBP) in combination with the transcriptome dataset GSE45603.
Conclusion: The findings support the genetically predicted causal relationship of gut microbiota with PPD and identify susceptible microbiota-host genes as potential therapeutic targets or diagnostic biomarkers, providing new insights into the prevention and intervention of PPD.
Keywords: postpartum depression, gut microbiota, Mendelian randomization, gut microbe‒host genes, transcriptome dataset
Introduction
Postpartum depression (PPD), a frequently observed complication after delivery, is a characteristic depressive episode that occurs 1 month after childbirth and is generally featured by a labile mood, low concentration, low self-esteem, sleep disorders, and a lack of excitement.1 One meta-analysis involving 27 studies reported that the global morbidity of PPD is 5.0%–26.32%.2 PPD has severe influence on daily activity and life quality of mothers and has negative impacts on newborns.3 Consequently, exploring the neurobiological mechanisms associated with PPD is important for ensuring physical and psychological health of both mothers and their children. Recently, more and more studies suggest that host gut microbial composition changes are closely related to PPD.4
Gut microbiota within human body accounts for dynamic and complex ecosystems with spatial heterogeneity and is the habitat of diverse microorganisms, which interact with each other and their host.5 Dysbiosis of gut microbiota can regulate the gastrointestinal (GI) immune response and affect central nervous system immunity, therefore, the microbiota–gut–brain (MGB) axis concept is formed6. The MGB axis is characterized as the bidirectional communication network comprising gut microbiota, its associated metabolites, immune system, neuroendocrine system, as well as enteric nervous system.4 A relation of maternal gut microbial alterations with depression during pregnancy or after childbirth has been observed in several studies. Zhou et al assessed the relationship between gut microbiota and PPD. According to their results, the abundances of several gut microbiota constituents of PPD cases, like Faecalibacterium, Phascolarctobacterium, and Lachnospiraceae, showed close relations to depressive symptom severity and sex hormone levels.7 The genus Lactobacillus had elevated abundance in both the high-fat diet (HFD)-induced PPD mouse model and the gestational diabetes mellitus (GDM)-mediated PPD mouse model.8,9 Supplementing probiotics with L. rhamnosus HN001 during pregnancy and after childbirth led to a reduction in postnatal depression and anxiety.10 These scientific findings support that gut microbial changes are strongly related to the occurrence of PPD. This association is likely mediated by mechanisms such as sex hormone regulation, neurotransmitter synthesis, short-chain fatty acid (SCFA) contents, immune responses, and hypothalamic–pituitary–adrenal (HPA) axis.11 However, the current research is associated with drawbacks, including inadequate statistical robustness due to the small sample size, disparities between human and animal studies, and inherent limitations of observational studies. The actual causal relationship of gut microbiota with PPD remains unknown and should be further explored.
In Mendelian randomization (MR) analysis, genetic variants tightly related to exposure are used as instrumental variables (IVs) for inferring the causality of risk factors with health outcomes. In comparison with observational epidemiology, MR can provide stronger evidence for causal inference due to its capability to mitigate the influence of potential confounders, reverse causation, and various additional sources of bias on the inferred relationships.12 Two-sample MR analysis is widely employed for exploring the relationship of gut microbiota with psychiatric disorders.13 Nonetheless, studies concerning the causal relationship of gut microbiota with PPD are currently lacking. Gene expression profiling is emerging as a powerful approach for the in-depth exploration of diagnostic biomarkers and pathogenic mechanisms associated with PPD.14 Nevertheless, related studies are mostly conducted to identify differentially expressed genes (DEGs) and not to analyze their associations with gut microbes. Therefore, it is necessary to clarify relationships of host genes with the gut microbiota among PPD patients.
Consequently, this present work focused on investigating the potential causality of gut microbiota with PPD using MR study, and identifying gut microbe-host genes and their potential molecular mechanisms through the use of data from the PPD transcriptome dataset.
Materials and Methods
Study Design
Figure 1 presents our study flowchart. In order to obtain unbiased estimates, IVs should meet three assumptions: (1) IVs are tightly related to exposure factors but have no direct connection to the outcome; (2) IVs do not demonstrate any association with confounders affecting the exposure-outcome relation; and (3) IVs influence the outcome via exposure factor pathway.
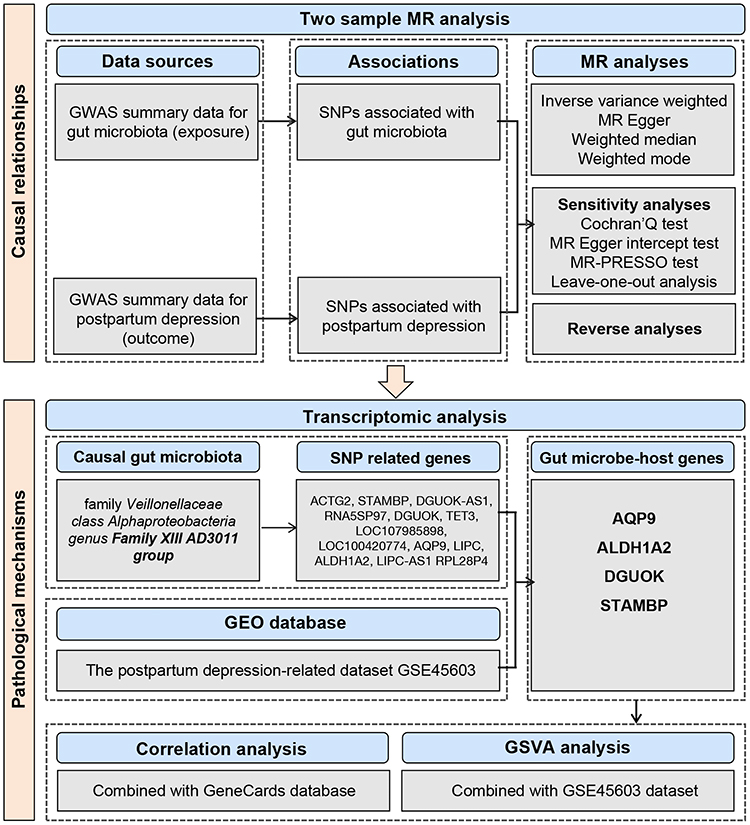 |
Figure 1 The study flowchart. |
Data Sources
We acquired latest GWAS data on PPD based on FinnGen database (https://gwas.mrcieu.ac.uk/datasets/finn-b-O15_POSTPART_DEPR/). The definition of PPD was made according to delivery status and the International Classification of Diseases, Tenth Revision (ICD-10).15 Age, sex, genetic relevance, genotyping batch, and the top 10 principal components were adjusted; thereafter, MR analysis was conducted on 7604 PPD patients and 59601 controls.
We acquired GWAS summary data on the gut microbiota in MiBioGen consortium (https://mibiogen.gcc.rug.nl/), the greatest, multiethnic, genome-wide meta-analysis of the gut microbiota thus far containing 18,340 subjects of 24 cohorts. Genetic variants located on the sex chromosomes were excluded, and sex was adjusted for in the analysis. Gut microbial composition was examined in three different regions within 16S rRNA gene (V4, V3-V4, and V1-V2).16 In addition, we performed microbiota quantitative trait locus (mbQTL) mapping for detecting genetic variants affecting relative abundances of microbial taxa. In our study, we used an average abundance >1% as the standard, and 196 taxa were included (9 phylum, 16 classes, 20 orders, 32 families, and 119 genus) for subsequent MR study.
We acquired the PPD-related microarray dataset GSE45603 based on Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE45603), including 210 blood samples. Finally, 21 of these samples, including 16 PPD and 5 healthy samples, were chosen; samples collected before conception, during the three trimesters of pregnancy and after delivery were excluded from the analysis.
We obtained a total of 152 PPD-related human genes based on GeneCards database (v5.10, https://www.genecards.org) with the keyword “postpartum depression” and a relevance score ≥ 7, as described previously.17
Inclusion and Exclusion of IVs
We chose IVs by criteria below: (1) SNPs related to gut microbial taxa with genome-wide significance (P < 5 × 10–8). Due to insufficient eligible IVs (P < 5 × 10–8), we chose an integrative threshold (P < 1 × 10–5) for obtaining the integrative outcomes. (2) We adopted data from 1,000 Genomes Project European samples to be a reference panel for calculating linkage disequilibrium (LD) among SNPs. For SNPs satisfying R2 <0.001 (clumping window size = 10,000 kb), just those having smallest p values were selected. (3) Following harmonization, F statistics of every SNP were determined, with the SNPs satisfying F statistics <10 being identified as weak instruments and eliminated. (4) SNPs showing minor allele frequency (MAF) ≤0.01 were discarded; and (5) when there were palindromic SNPs, we utilized allele frequency data for inferring forward stranded alleles, which avoided allele coding or strand orientation distortion.
MR Analysis
MR analysis was conducted by employing four distinct methods, namely, inverse variance weighted (IVW), MR‒Egger, weighted mode, and weighted median. IVW represents an essential meta-analysis approach, which assumes that instruments impact outcomes only via the interested exposure rather than via other pathways. When there is no heterogeneity or horizontal pleiotropy, IVWs make credible estimates.18 MR‒Egger can reliably estimate causality even in the presence of pleiotropic effects for every genetic variant, if the relation between the genetic variant and exposure is not dependent on pleiotropic effects. With respect to the weighted median, ≥50% of the weight utilized for analysis was considered to meet validity criteria. If the greatest instrument subset for identifying identical causality is valid, weighted mode still comes to similar results, although most of the other instruments are invalid. A causal relationship was assumed for P < 0.05 according to the four approaches used for MR analysis. For different findings obtained from those four MR analysis approaches, IVW results were used.
There were various sensitivity analyses conducted for analyzing significance of estimates for identifying possible heterogeneity and pleiotropy. Additionally, we utilized Cochran’s Q test to determine heterogeneity. Directional horizontal pleiotropic effects were tested by using MR–Egger intercepts. We identified potential horizontal pleiotropy and outliers through MR-Pleiotropy Residual Sum and Outlier (MR-PRESSO) test, after which outlier SNPs were removed to obtain correct outcomes. Lack of heterogeneity or pleiotropy was defined as p > 0.05. This study also performed leave-one-out analysis for examining the possible biased or driven causal estimates resulting from one individual SNP, where one instrumental SNP was removed each time for repeated IVW analysis.
To examine whether PPD was causally related to key gut microbial taxa obtained from forward MR study, we conducted reverse MR study by an identical method used for forward MR analysis, in which PPD served as the exposure agent, whereas key gut microbial taxa were the outcomes.
False discovery rate (FDR) was corrected using q-value method. The results were deemed statistically significant when the FDR q < 0.1 but suggestive of an association when q ≥ 0.1 and p < 0.05.19
R version 4.3 was adopted in statistical analysis. MR study was completed with two-sample MR (version 0.5.6) and MR-PRESSO R package. p < 0.05 (two-tailed) stood for a significant difference.
SNP-Related Gene Extraction
The retrieval function get_variants (), a component of the R package gwasrapidd, was employed to extract all genes associated with SNP in accordance with prior methods.20
GSVA Analysis
GSVA is an unsupervised and non-parametric technique that evaluates the enrichment of gene sets in the transcriptome.21 To analyze the possible changes in biological functions between diverse samples, gene sets were obtained based on Molecular Signatures Database (v7.4) (http://www.gseamsigdb.org/gsea/msigdb/). The GSVA algorithm was subsequently applied for comprehensive scoring of these gene sets.
Results
Causality of Gut Microbiota with PPD
Following the IVs selection criteria, 2037 SNPs were chosen (103 SNPs for phylum, 179 for class, 217 for order, 339 for family, and 1199 for genus) upon p < 1 × 10–5. Three bacterial genera, namely, the family Veillonellaceae, class Alphaproteobacteria, and genus Family XIII AD3011 group, were related to PPD according to one or more MR approaches. The family Veillonellaceae (OR = 0.82, 95% CI = 0.71–0.94, p = 0.004, q = 0.030) was related to a reduced incidence of PPD, according to IVW analysis. The results were verified via weighted median analysis (OR = 0.79, 95% CI = 0.65–0.97, p = 0.024) and weighted mode (OR = 0.76, 95% CI = 0.60–0.97, p = 0.044) analysis (Table 1, Figure 2A). However, the class Alphaproteobacteria (OR = 1.22, 95% CI = 1.01–1.47, p = 0.041, q = 0.074) (Table 1, Figure 2B) and the genus Family XIII AD3011 group (OR = 1.24, 95% CI = 1.04–1.48, p = 0.019, q = 0.065) were significantly related to elevated PPD incidence upon IVW analysis (Table 1, Figure 2C).
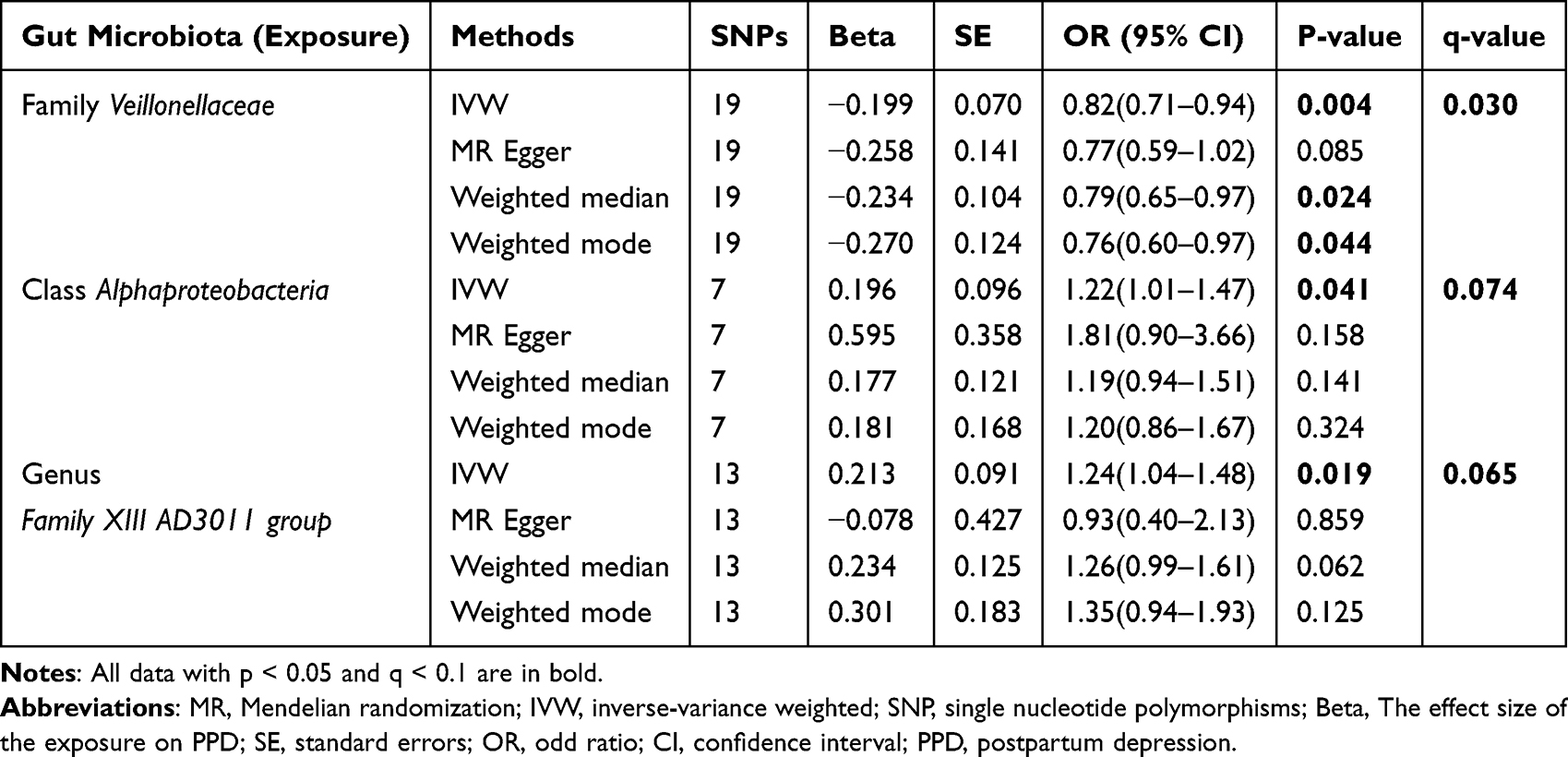 |
Table 1 MR Estimates for the Association Between the Gut Microbiota and PPD Incidence |
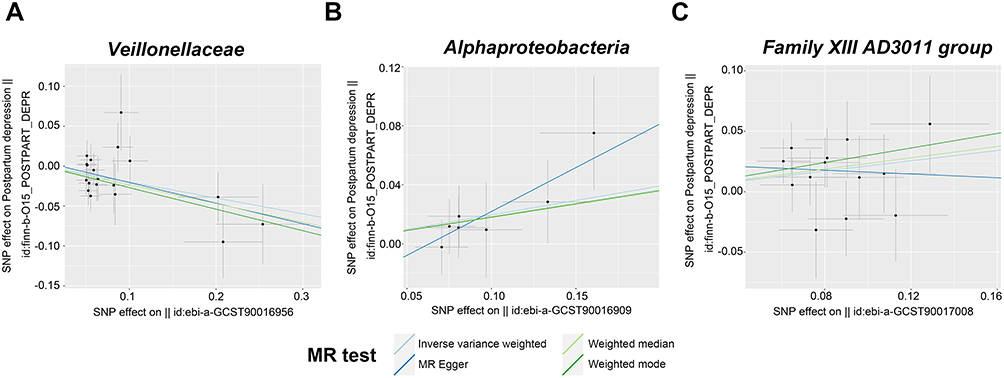 |
Figure 2 Scatter plots showing causal association of the gut microbiota with PPD. (A) Veillonellaceae. (B) Alphaproteobacteria. (C) Family XIII AD3011 group. |
According to Table 2, we did not detect any horizontal pleiotropy from MR-PRESSO global test or MR‒Egger intercept test (p > 0.05). Moreover, heterogeneity was not detected by Cochran Q test (p > 0.05). Besides, leave-one-out analysis suggested no significantly different causal estimates of family Veillonellaceae, class Alphaproteobacteria, and genus Family XIII AD3011 group on PPD (Figure 3A–C), indicating that no individual IV drove those detected causal relationships.
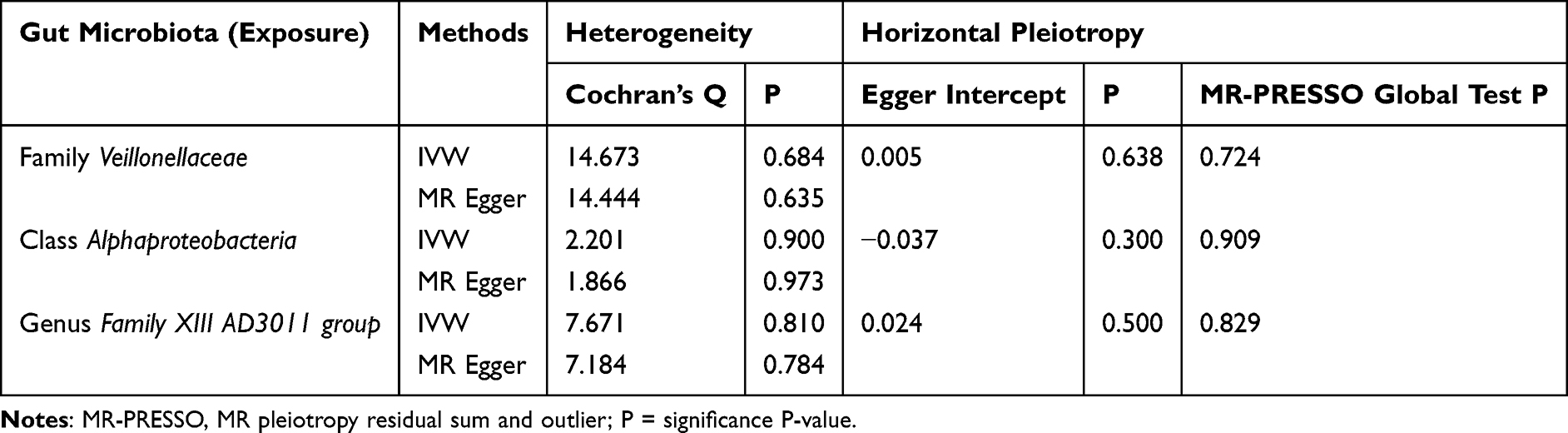 |
Table 2 Heterogeneity and Horizontal Pleiotropy Analyses |
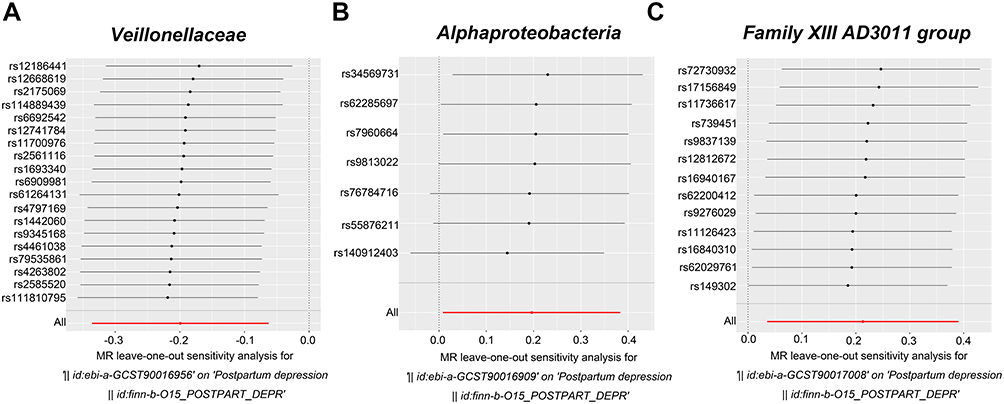 |
Figure 3 Leave one-out plots showing the causality of gut microbiota with PPD. (A) Veillonellaceae. (B) Alphaproteobacteria. (C) Family XIII AD3011 group. |
Reverse MR Study
PPD showed no causal relation to these three bacterial genera according to reverse MR analysis. Supplementary Table 1 displays more details about the IVs adopted for reverse MR analysis.
Gut Microbe-Host Gene Identification
Given the reliability and significance of the Family XIII AD3011 group from MR studies, SNPs were annotated in the Family XIII AD3011 group at a locus-wide significance level, as a result, 13 SNP-related genes were extracted from the functional get_variants. Among them, rs11126423 was associated with 8 genes, and rs16940167 was associated with 5 genes; most of these genes were intergenic, followed by upstream. The 13 SNP related genes were as follows: ACTG2, STAMBP, DGUOK-AS1, RNA5SP97, DGUOK, TET3, LOC107985898, LOC100420774, AQP9, LIPC, ALDH1A2, LIPC-AS1, and RPL28P4 (Supplementary Table 2). To further identify relevant genes that impact the onset and progression of PPD, we conducted a cross-reference analysis between these 13 SNP-related genes and the transcriptome dataset GSE45603. Consequently, four gut microbe-host genes were identified: AQP9, ALDH1A2, DGUOK, and STAMBP.
Correlation Analysis of Gut Microbe-Host Genes and PPD Pathogenic Genes
We crossed 152 PPD-related human genes obtained from the GeneCards database with those from the GSE45603 dataset and screened the top 20 genes as PPD pathogenic genes based on the relevance score (Supplementary Table 3). Correlation analysis between 4 gut microbe-host genes and 20 PPD pathogenic genes was further conducted (Figure 4A). AQP9 was strongly positively related to IL-1β (p = 0.00016, cor = 0.81) (Figure 4B). ALDH1A2 exhibited an obvious positive correlation with PER3 (p = 0.000054, cor = 0.84) (Figure 4C). DGUOK demonstrated a markedly positive correlation with TNF-α (p = 0.0051, cor = 0.66) (Figure 4D), and STAMBP showed an obvious negative correlation with HP1BP3 (p = 0.0095, cor =−0.63) (Figure 4E).
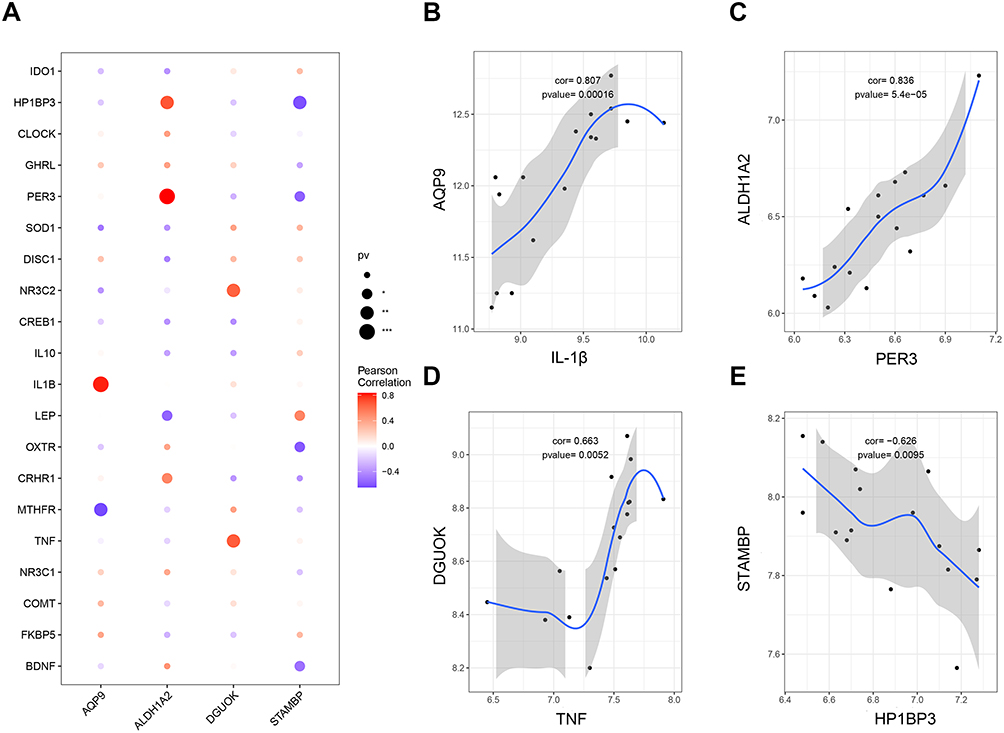 |
Figure 4 Correlation analysis between gut microbe-host genes and PPD pathogenesis genes. (A) Correlations between 4 gut microbe-host genes and 20 PPD pathogenic genes. (B) Relation of AQP9 with IL-1β. (C) Relation of ALDH1A2 with PER3. (D) Correlation between DGUOK and TNF-α. (E) Correlation between STAMBP and HP1BP3. |
GSVA of Gut Microbe-Host Genes and Pathways
Subsequently, possible molecular mechanisms were elucidated, and pathways associated with those four core genes were analyzed through GSVA. As a result, AQP9 was significantly associated with 24 signaling pathways with an absolute value ≥1.0 and a p-value <0.05, including IL6_JAK_STAT3_SIGNALING, INFLAMMATORY_RESPONS, TNFA_SIGNALING_VIA_NFKB, and FATTY_ACID_METABOLISM (Figure 5A). The pathways associated with the enrichment of ALDH1A2 included INFLAMMATORY_RESPONSE, PI3K_AKT_MTOR_SIGNALING, and ESTROGEN_RESPONSE_LATE (Figure 5B). The DGUOK-enriched pathways included OXIDATIVE PHOSPHORYLATION, GLYCOLYSIS, FATTY_ACID_METABOLISM, and IL6_JAK_STAT3_SIGNALING (Figure 5C). The STAMBP-enriched signaling pathways included the COMPLEMENT, KRAS SIGNALING, and PI3K_AKT_MTOR_SIGNALING pathways (Figure 5D).
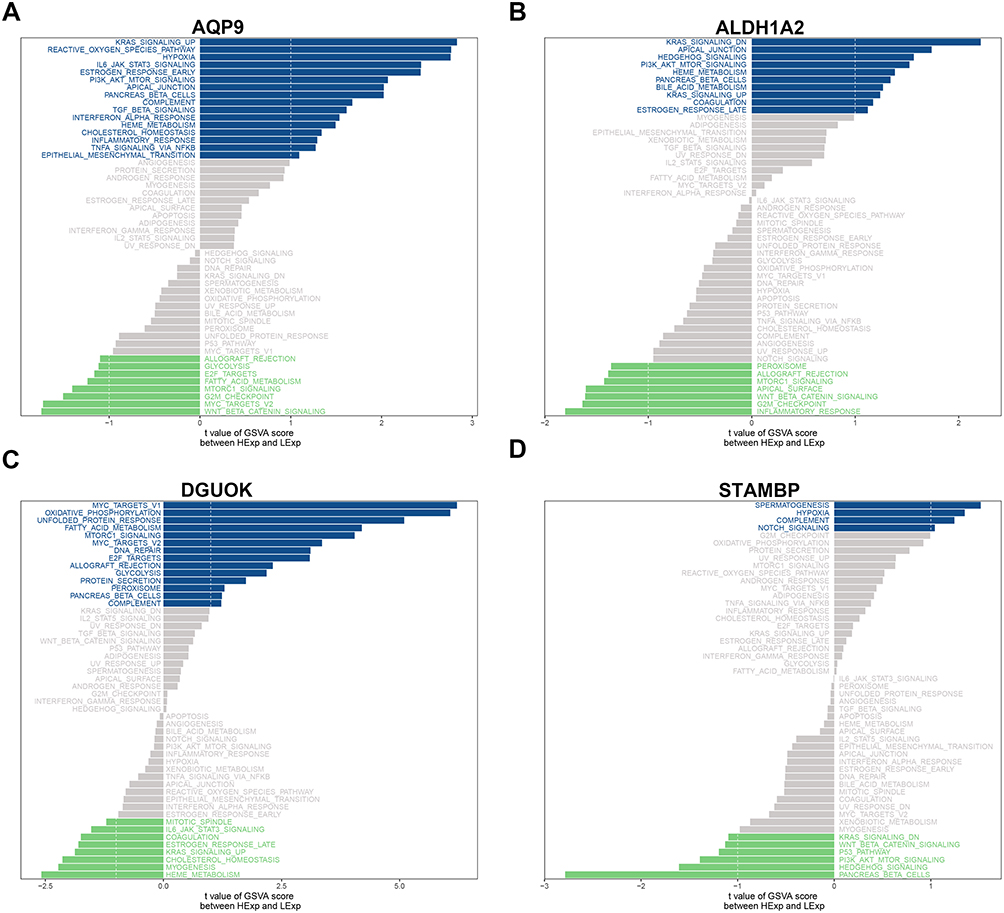 |
Figure 5 GSVA of four gut microbe-host genes. (A) Pathways enriched by AQP9. (B) Pathways enriched by ALDH1A2. (C) Pathways enriched by DGUOK. (D) Pathways enriched by STAMBP. Blue and green bands indicate positive and negative relations, separately. |
Discussion
Our work focuses on assessing the causality of gut microbiota with PPD by two-sample bidirectional MR study and identifying the potential relationships between pathogenic bacteria and host genes for PPD. Our study suggested that the family Veillonellaceae dramatically reduced the incidence of PPD, whereas the class Alphaproteobacteria and genus Family XIII AD3011 group significantly increased the risk of PPD. Furthermore, we identified four microbial (Family XIII AD3011 group) host genes, including AQP9, ALDH1A2, DGUOK, and STAMBP, and investigated their potential molecular mechanisms for preventing PPD.
The family Veillonellaceae is in the class Clostridia of phylum Firmicutes. In previous studies, the relative abundance of Veillonellaceae decreased markedly among depression patients and autistic children.17,22 Additionally, one longitudinal cohort study suggested that maternal psychological distress is significantly related to lower Veillonellaceae abundance in infant fecal specimens aged 20–28 weeks.23 Based on these results, Veillonellaceae may protect from neurodevelopmental impairment or psychological distress. Based on the above findings, this work revealed that a decreased relative abundance of Veillonellaceae was causally related to decreased PPD incidence, suggesting its protective role. However, the mechanism through which Veillonellaceae affects PPD is still unclear.
Many members of the Veillonellaceae have the capability to ferment carbohydrates into different types of SCFAs, including butyrate, propionate, and acetate. They participate in host immunological regulation and inflammatory homeostasis. Additionally, these SCFAs can cross blood-brain barrier, which serves as pivotal mediators in gut-brain axis.24,25 In most cases, the abundance of fecal SCFAs is strongly associated with depressive symptoms of major depressive disorder (MDD) patients.26,27 Furthermore, a high-fiber diet prevents gut microbial disturbance as well as depressive-like behaviors in the HFD-induced PPD mouse model through microbial remodeling and elevating SCFA contents.8 The potential mechanisms through which SCFAs influence emotional states may be attributed to their ability to regulate serotonin metabolism, the HPA axis, and the expression of G-protein-coupled receptors.28–30 Moreover, some reports have proposed that butyrate may exhibit antidepressant-like properties. Animal experiments revealed that sodium butyrate improved depression-like behaviors by increasing 5-HT and BDNF levels, restoring BBB function, while suppressing microglial activation.31–33 In addition, exogenous supplementation with SCFAs was found to ameliorate depression- and anxiety-like behaviors through suppressing conversion of Trp to Kyn in order to rescue 5-HT content decline within the gut and hippocampi.34 Based on the aforementioned evidence, SCFA intake and generation in pregnancy are significant factors for PPD occurrence. Therefore, we speculate that Veillonellaceae may exert a protective effect on PPD by elevating SCFA abundance.
The class Alphaproteobacteria consists of free-living, gram-negative, obligate and symbiotic intracellular bacterial strains, key human, plant and animal pathogens.35 In several studies, Alphaproteobacteria abundance is tightly related to depression severity, but results are variable, with no consistent trends. Paweł et al reported that depression severity was negatively related to Alphaproteobacteria abundance among MDD patients.36 However, two other studies reported the opposite results. The relative abundance of Alphaproteobacteria is positively related to worse depression and anxiety scores and to decrease with antidepressant use.37,38 Similarly, a higher Alphaproteobacteria abundance was causally related to increased PPD incidence in our study, reflecting its harmful effect. However, the mechanism through which Alphaproteobacteria affect PPD is still unclear.
The genus Family XIII AD3011 group is in order Clostridiales of phylum Firmicutes. In a previous study, Family XIII AD3011 group is positively related to a significant risk factor for depression, namely, negative cognitive processing bias (NCPB).39 Consistent with the findings of this study, the Family XIII AD3011 group might have had a higher incidence of PPD. However, the underlying influencing mechanism remains unclear.
One recent study revealed a significant reduction of Family XIII AD3011 group abundance in 2nd and 3rd trimesters during pregnancy; this group is recognized as critical gut microbial genera throughout pregnancy and lactation. Furthermore, the Family XIII AD3011 group exhibited a strong relationship to comprehensive alterations in serum and fecal hormones, particularly decreased progesterone and estradiol levels.40 Fluctuations in reproductive hormones in pregnancy and postpartum are significant factors for PPD development. Progesterone concentrations in pregnant women increase by 20 times relative to normal levels, whereas blood estradiol concentrations can increase by 200–300 times at the end of pregnancy.41 Their blood concentrations apparently decline after placental expulsion, which supports the occurrence of behavioral, affective, and anxiety-like symptoms within 2 weeks after childbirth.42 According to several clinical articles, estrogen and progesterone supplementation decrease depression symptoms after childbirth.43 In contrast, reproductive hormone withdrawal can cause depression-like behaviors in animal models of PPD.44,45 The mechanisms are thought to involve epigenetic changes, HPA axis function, GABAergic signaling, and gut-brain axis.46 Interestingly, certain gut microbes secrete β-glucuronidase, which decomposes estrogen to the bioactive form.47 Gut microbial disruption after childbirth can dramatically reduce estrogen and progesterone levels through influencing relevant enzymes (eg, β-glucuronidase) involved in hormone generation, thereby promoting PPD. These findings provided strong evidence for our study, and we hypothesized that an excessive Family XIII AD3011 group level in the gut might contribute to PPD by reducing estradiol and progesterone levels. However, this assumption warrants further investigation.
Another potential mechanism by which the Family XIII AD3011 group affects PPD is inflammatory response. In recent years, limited articles have shown the correlation between elevated Family XIII AD3011 group levels and proinflammatory responses.48 PPD is related to increased proinflammatory factor levels, which is suggested to be related to depressive symptoms. The IL-6, IL-1β, and TNF-α levels exhibit positive relation to depressive scores among females after childbirth.49 In contrast, T-cell number and function are negatively related to depression symptoms after childbirth.46 It has been proposed that proinflammatory factors induce indoleamine-2,3 dioxygenase (IDO) activation, thus metabolizing tryptophan (a serotonin precursor) to kynurenine. Elevated levels of kynurenine and inflammation-mediated degradation of tryptophan restrict serotonin production, thereby exacerbating depression symptoms during PPD.50 Although the Family XIII AD3011 group likely exerts an important effect on PPD occurrence through activating the pro-inflammatory response, direct empirical evidence is currently lacking, and further research is needed.
Gut microbes may affect gene expression levels through the MGB axis to regulate host physiology or induce psychiatric disorders.51 Therefore, this study also analyzed how specific interactions of pathogenic gut microbes with host genes affected PPD onset. Given the reliability and importance of the Family XIII AD3011 group in MR studies, we identified 4 pivotal host genes (AQP9, ALDH1A2, DGUOK and STAMBP) that may be associated with the Family XIII AD3011 group among the PPD-specific populations, as well as being expressed in the GSE45603 dataset. Aquaporin 9 (AQP9) belongs to the membrane protein family and exerts significant roles in glucose and lipid metabolism, immune responses, and sex hormone regulation.52 Besides, AQP9 is the central gene associated with both Parkinson’s disease (PD) and MDD, suggesting that it may be the therapeutic target of these conditions.53 Recent research has identified a proinflammatory function of AQP9 in the brain, suggesting that it may exacerbate inflammation through metabolic byproducts.54 Additionally, AQP9 is abundantly expressed in the intestine, where probiotics may influence intestinal water metabolism by regulating AQP9 levels.55 Nevertheless, the role of AQP9 within the MGB axis in PPD remains inadequately understood. Our study demonstrates a positive correlation between AQP9 and IL-1β, as well as its association with inflammation-related signaling pathways, including IL6_JAK_STAT3_SIGNALING, INFLAMMATORY_RESPONSE, and TNFA_SIGNALING_VIA_NFKB. We hypothesize that the Family XIII AD3011 group may trigger an inflammatory response by modulating AQP9 expression, thereby contributing to PPD. However, this hypothesis necessitates further validation in future studies due to the current lack of direct evidence.
ALDH1A2, which is capable of encoding an enzyme associated with astrocyte-derived retinoic acid, accounts for an essential neuronal morphogen linked to schizophrenia.56 ALDH1A2 has been identified as a hub gene associated with Cesarean section (CS) birth and affecting adult anxiety and self-harm risk, shedding novel lights in mental disorder occurrence.57 It has been shown that elevated levels of ALDH1A2 expression have been detected in the hippocampus of suicide decedents.58 Additionally, ALDH1A2 is crucial for maintaining intestinal homeostasis. Its expression in intestinal dendritic cells is induced by SCFAs, which modulate intestinal immune responses.59 Recent studies have identified interactions between L. intestinalis and ALDH1A2, leading to the production of retinoic acid, which helps preserve epithelial integrity and mitigate inflammatory responses.60 Our correlation analysis revealed a positive association between ALDH1A2 and PER3, a key gene linked to PPD. The pathways associated with the enrichment of ALDH1A2 included PI3K_AKT_MTOR_SIGNALING, INFLAMMATORY_RESPONSE, and ESTROGEN_RESPONSE_LATE. We hypothesize that the underlying mechanism by which the Family XIII AD3011 group regulates ALDH1A2 expression to participate in the occurrence of PPD is related to inflammatory response and retinoic acid metabolism. However, direct evidence is lacking and this assumption warrants further investigation.
Deoxyguanosine kinase (DGUOK) offers guanosine and adenosine nucleotides to facilitate mitochondrial DNA (mtDNA) replication.61 Mutations in DGUOK are known to cause mtDNA depletion and mitochondrial dysfunction.62 A recent study revealed that gestational stress in female rats leads to reduced mitochondrial respiration in the prefrontal cortex, increased inflammation, and the emergence of postpartum depression-like behaviors, underscoring the role of mitochondrial dysfunction in PPD.63 Our study first revealed that DGUOK was positively associated with NR3C2, a high-risk biomarker related to depression in pregnancy and postpartum.64 Furthermore, gut microbiota dysbiosis may contribute to the development and progression of depression by inducing mitochondrial dysfunction and disrupting energy metabolism.65 GSVA results indicated that DGUOK is enriched in mitochondrial function-related pathways, including OXIDATIVE_PHOSPHORYLATION, GLYCOLYSIS, and FATTY_ACID_METABOLISM. Thus, we propose the hypothesis that the Family XIII AD3011 group may regulate DGUOK expression, leading to mitochondrial dysfunction and thereby contributing to the development of PPD.
STAM-binding protein (STAMBP) is a deubiquitinase that plays a key role in upregulating inflammasome activity.66 A recent study identified STAMBP as an inflammatory biomarker in late pregnancy that may help assess the risk of PPD.67 Additionally, gut microbiota may promote increased secretion of STAMBP, which has been implicated in tumor progression.68 Consistent with previous findings, our correlation analysis revealed a significant negative relationship between STAMBP and HP1BP3, an important epigenetic biomarker associated with PPD. Furthermore, STAMBP was enriched in immune-related signaling pathways, including COMPLEMENT, KRAS_SIGNALING_UP, and PI3K_AKT_MTOR_SIGNALING. Based on these results, we hypothesize that the Family XIII AD3011 group may contribute to the development of PPD by upregulating STAMBP expression. Nevertheless, the precise mechanism of action of these four genes within the MGB axis of PPD remains unclear. Additional large-scale experimental studies are warranted for validation.
To examine whether PPD exerts any causal effect on the three identified bacterial genera, a reverse MR analysis was conducted. The results did not support reverse causality from PPD to the gut microbiota, indicating that the association observed in the forward MR analysis is unlikely to be driven by pre-existing PPD altering the gut microbiota. MR analysis relies on the assumption that IVs influence the outcome only through the exposure. The absence of reverse causality suggests that the IVs used for PPD did not operate via an effect on these specific gut microbiota. Thus, the reverse MR results strengthen our main conclusion that gut microbiota has a causal influence on PPD. However, we acknowledge that during the postpartum period, it cannot be ruled out that PPD may affect the gut microbiota through behavioral, immunological or neuroendocrine pathways. This needs to be confirmed by further studies.
Certain limitations must be noted in the present work. First, European GWAS summary data were mostly utilized; as a result, the conclusions might not be generalizable to other ethnicities. Second, the genus level was the lowest, making it impossible to further explore the causality of gut microbiota with PPD at a species level. Third, GWAS meta-analyses on the gut microbiota were not limited to females, which might induce potential sex bias. However, we eliminated genetic variants in sex chromosomes and adjusted sex for analysis, further minimizing the influence of sex on the results.16 Fourth, SNPs utilized for analysis were not of conventional GWAS significance (p < 5 × 10−8) (Supplementary Table 4). More genetic variants should be included as an IV to analyze the sensitivity and ensure the precise horizontal polymorphism detection. Therefore, a series of multi-faceted sensitivity analyses and the FDR correction were conducted to mitigate potential bias and limit the possible false positives. Fifth, GSE45603 had a small sample size, and validation of independent external cohorts was not performed due to the limited accessibility of clinical data in public databases and ethical constraints. We analyzed the GSE70732 dataset, targeting brain tissue of postpartum mice. The expressions of AQP9 and ALDH1A2 were upregulated in the hypothalamus, while other genes remained unchanged, possibly due to species differences (Supplementary Figure 1). Additional large-scale experimental studies are needed for validation. At last, due to reliance on summary-level GWAS data, the MR analysis lacked stratification by parity, delivery mode, breastfeeding status or other relevant demographic and clinical factors. The potential for interaction between diet, genes or environment, which could impact our results, could not be fully ruled out.
Conclusion
This work provided evidence for genetically predicted causal associations between three bacterial genera, namely, the family Veillonellaceae, class Alphaproteobacteria and genus Family XIII AD3011 group, and PPD using two-sample bidirectional MR method. The Family XIII AD3011 group significantly increases PPD risk. Therefore, we further identified four microbial (Family XIII AD3011 group)-host genes using a transcriptome dataset and investigated their related enrichment pathways. Our study explored the pathogenesis of gut microbiota on PPD and identified candidate biomarkers, providing therapeutic targets for PPD and novel directions for further clinical studies. Nevertheless, it was not possible to completely rule out the presence of confounding factors, including diet, lifestyle choices and antibiotic use, which might weaken causal inference.
Ethics Approval and Consent to Participate
All data used in this study came from publicly available GWAS summary statistics on gut microbiota and postpartum depression and had obtained prior ethical approvals and participant consent. Exemption from institutional ethics review for this study is granted under Article 32 of China’s Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (effective February 18, 2023), specifically with the meeting of the following criteria:
(1) Item 1: Research conducted using public databases that does not involve the identification or processing of personal information.
(2) Item 2: Analysis of previously collected data without additional data acquisition or direct involvement with human subjects. Our institutional review board did not require any additional ethical approval because this work exclusively involved secondary analysis of aggregated genetic summary data, and no access to individual-level information was required.
Acknowledgments
Our thanks should go to all participants and investigators of studies or consortiums included in this work for sharing GWAS data.
Funding
This work was supported by the Qingdao Key Medical and Health Discipline Project.
Disclosure
The authors declare no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Stewart DE, Vigod SN. Postpartum depression: pathophysiology, treatment, and emerging therapeutics. Annu Rev Med. 2019;70:183–196. doi:10.1146/annurev-med-041217-011106
2. Liu X, Wang S, Wang G. Prevalence and risk factors of postpartum depression in women: a systematic review and meta-analysis. J Clin Nurs. 2022;31(19–20):2665–2677. doi:10.1111/jocn.16121
3. Weissman MM. Postpartum depression and its long-term impact on children: many new questions. JAMA Psychiatry. 2018;75(3):227–228. doi:10.1001/jamapsychiatry.2017.4265
4. Simpson CA, Diaz-Arteche C, Eliby D, et al. The gut microbiota in anxiety and depression - A systematic review. Clin Psychol Rev. 2021;83:101943. doi:10.1016/j.cpr.2020.101943
5. Gomaa EZ. Human gut microbiota/microbiome in health and diseases: a review. Antonie van Leeuwenhoek. 2020;113(12):2019–2040. doi:10.1007/s10482-020-01474-7
6. Cryan JF, O’Riordan KJ, Cowan CSM, et al. The microbiota-gut-brain axis. Physiol Rev. 2019;99(4):1877–2013. doi:10.1152/physrev.00018.2018
7. Zhou Y, Chen C, Yu H, et al. Fecal microbiota changes in patients with postpartum depressive disorder. Front Cell Infect Microbiol. 2020;10:567268. doi:10.3389/fcimb.2020.567268
8. Liu Z, Li L, Ma S, et al. High-dietary fiber intake alleviates antenatal obesity-induced postpartum depression: roles of gut microbiota and microbial metabolite short-chain fatty acid involved. J Agric Food Chem. 2020;68(47):13697–13710. doi:10.1021/acs.jafc.0c04290
9. Zhao R, Zhou Y, Shi H, et al. Effect of gestational diabetes on postpartum depression-like behavior in rats and its mechanism. Nutrients. 2022;14(6).
10. Slykerman RF, Hood F, Wickens K, et al. Effect of Lactobacillus rhamnosus HN001 in pregnancy on postpartum symptoms of depression and anxiety: a randomised double-blind placebo-controlled trial. EBioMedicine. 2017;24:159–165. doi:10.1016/j.ebiom.2017.09.013
11. Zhang S, Lu B, Wang G. The role of gut microbiota in the pathogenesis and treatment of postpartum depression. Ann Gen Psychiatry. 2023;22(1):36. doi:10.1186/s12991-023-00469-8
12. Smith GD, Ebrahim S. Mendelian randomization: prospects, potentials, and limitations. Int J Epidemiol. 2004;33(1):30–42. doi:10.1093/ije/dyh132
13. Ni JJ, Xu Q, Yan SS, et al. Gut microbiota and psychiatric disorders: a two-sample Mendelian randomization study. Front Microbiol. 2021;12:737197. doi:10.3389/fmicb.2021.737197
14. Pan D, Xu Y, Zhang L, et al. Gene expression profile in peripheral blood mononuclear cells of postpartum depression patients. Sci Rep. 2018;8(1):10139. doi:10.1038/s41598-018-28509-4
15. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613(7944):508–18.
16. Kurilshikov A, Medina-Gomez C, Bacigalupe R, et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet. 2021;53(2):156–165. doi:10.1038/s41588-020-00763-1
17. Xu XJ, Lang JD, Yang J, et al. Differences of gut microbiota and behavioral symptoms between two subgroups of autistic children based on gammadeltaT cells-derived IFN-gamma Levels: a preliminary study. Front Immunol. 2023;14:1100816. doi:10.3389/fimmu.2023.1100816
18. Bowden J, Del Greco MF, Minelli C, et al. A framework for the investigation of pleiotropy in two-sample summary data Mendelian randomization. Stat Med. 2017;36(11):1783–1802. doi:10.1002/sim.7221
19. Li P, Wang H, Guo L, et al. Association between gut microbiota and preeclampsia-eclampsia: a two-sample Mendelian randomization study. BMC Med. 2022;20(1):443. doi:10.1186/s12916-022-02657-x
20. Magno R, Maia AT. gwasrapidd: an R package to query, download and wrangle GWAS catalog data. Bioinformatics. 2020;36(2):649–650. doi:10.1093/bioinformatics/btz605
21. Hanzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinf. 2013;14:7. doi:10.1186/1471-2105-14-7
22. Kang DW, Park JG, Ilhan ZE, et al. Reduced incidence of Prevotella and other fermenters in intestinal microflora of autistic children. PLoS One. 2013;8(7):e68322. doi:10.1371/journal.pone.0068322
23. Naude PJW, Claassen-Weitz S, Gardner-Lubbe S, et al. Association of maternal prenatal psychological stressors and distress with maternal and early infant faecal bacterial profile. Acta Neuropsychiatr. 2020;32(1):32–42. doi:10.1017/neu.2019.43
24. Dalile B, Van Oudenhove L, Vervliet B, et al. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. 2019;16(8):461–478. doi:10.1038/s41575-019-0157-3
25. Park J, Kim CH. Regulation of common neurological disorders by gut microbial metabolites. Exp Mol Med. 2021;53(12):1821–1833. doi:10.1038/s12276-021-00703-x
26. Liu RT, Rowan-Nash AD, Sheehan AE, et al. Reductions in anti-inflammatory gut bacteria are associated with depression in a sample of young adults. Brain Behav Immun. 2020;88:308–324. doi:10.1016/j.bbi.2020.03.026
27. Skonieczna-Zydecka K, Grochans E, Maciejewska D, et al. Faecal short chain fatty acids profile is changed in Polish depressive women. Nutrients. 2018;10(12):1939. doi:10.3390/nu10121939
28. Dinan TG, Microbes CJF. Immunity, and behavior: psychoneuroimmunology meets the microbiome. Neuropsychopharmacology. 2017;42(1):178–192. doi:10.1038/npp.2016.103
29. Pascale A, Marchesi N, Marelli C, et al. Microbiota and metabolic diseases. Endocrine. 2018;61(3):357–371. doi:10.1007/s12020-018-1605-5
30. Yu S, Wang L, Jing X, et al. Features of gut microbiota and short-chain fatty acids in patients with first-episode depression and their relationship with the clinical symptoms. Front Psychol. 2023;14:1088268. doi:10.3389/fpsyg.2023.1088268
31. Wei Y, Melas PA, Wegener G, et al. Antidepressant-like effect of sodium butyrate is associated with an increase in TET1 and in 5-hydroxymethylation levels in the Bdnf gene. Int J Neuropsychopharmacol. 2014;18(2). doi:10.1093/ijnp/pyu032
32. Sun J, Wang F, Hong G, et al. Antidepressant-like effects of sodium butyrate and its possible mechanisms of action in mice exposed to chronic unpredictable mild stress. Neurosci Lett. 2016;618:159–166. doi:10.1016/j.neulet.2016.03.003
33. Shi H, Ge X, Ma X, et al. A fiber-deprived diet causes cognitive impairment and hippocampal microglia-mediated synaptic loss through the gut microbiota and metabolites. Microbiome. 2021;9(1):223. doi:10.1186/s40168-021-01172-0
34. Han W, Zheng Y, Wang L, et al. Disordered gut microbiota and changes in short-chain fatty acids and inflammatory processes in stress-vulnerable mice. J Neuroimmunol. 2023;383:578172. doi:10.1016/j.jneuroim.2023.578172
35. Hallez R, Delaby M, Sanselicio S, et al. Hit the right spots: cell cycle control by phosphorylated guanosines in alphaproteobacteria. Nat Rev Microbiol. 2017;15(3):137–148. doi:10.1038/nrmicro.2016.183
36. Liskiewicz P, Kaczmarczyk M, Misiak B, et al. Analysis of gut microbiota and intestinal integrity markers of inpatients with major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2021;106:110076. doi:10.1016/j.pnpbp.2020.110076
37. Cirstea MS, Kliger D, MacLellan AD, et al. The oral and fecal microbiota in a Canadian cohort of Alzheimer’s disease. J Alzheimers Dis. 2022;87(1):247–258. doi:10.3233/JAD-215520
38. Lukic I, Getselter D, Ziv O, et al. Antidepressants affect gut microbiota and Ruminococcus flavefaciens is able to abolish their effects on depressive-like behavior. Transl Psychiatry. 2019;9(1):133. doi:10.1038/s41398-019-0466-x
39. Xu HM, Xie SW, Liu TY, et al. Microbiota alteration of Chinese young male adults with high-status negative cognitive processing bias. Front Microbiol. 2023;14:989162. doi:10.3389/fmicb.2023.989162
40. Zhang K, Liu G, Wu Y, et al. Gut microbial succession patterns and metabolic profiling during pregnancy and lactation in a goat model. Microbiol Spectr. 2023;11(1):e0295522. doi:10.1128/spectrum.02955-22
41. Brummelte S, Galea LA. Postpartum depression: etiology, treatment and consequences for maternal care. Horm Behav. 2016;77:153–166. doi:10.1016/j.yhbeh.2015.08.008
42. Amodeo G, Laurenzi PF, Santucci A, et al. Advances in treatment for postpartum major depressive disorder. Expert Opin Pharmacother. 2020;21(14):1685–1698. doi:10.1080/14656566.2020.1779702
43. Moses-Kolko EL, Berga SL, Kalro B, et al. Transdermal estradiol for postpartum depression: a promising treatment option. Clin Obstet Gynecol. 2009;52(3):516–529. doi:10.1097/GRF.0b013e3181b5a395
44. Stoffel EC, Craft RM. Ovarian hormone withdrawal-induced “depression” in female rats. Physiol Behav. 2004;83(3):505–513. doi:10.1016/j.physbeh.2004.08.033
45. Suda S, Segi-Nishida E, newton SS, et al. A postpartum model in rat: behavioral and gene expression changes induced by ovarian steroid deprivation. Biol Psychiatry. 2008;64(4):311–319. doi:10.1016/j.biopsych.2008.03.029
46. Payne JL, Maguire J. Pathophysiological mechanisms implicated in postpartum depression. Front Neuroendocrinol. 2019;52:165–180. doi:10.1016/j.yfrne.2018.12.001
47. Baker JM, Al-Nakkash L, Herbst-Kralovetz MM. Estrogen-gut microbiome axis: physiological and clinical implications. Maturitas. 2017;103:45–53. doi:10.1016/j.maturitas.2017.06.025
48. Zhang L, Jing J, Han L, et al. Characterization of gut microbiota, metabolism and cytokines in benzene-induced hematopoietic damage. Ecotoxicol Environ Saf. 2021;228:112956. doi:10.1016/j.ecoenv.2021.112956
49. Worthen RJ, Beurel E. Inflammatory and neurodegenerative pathophysiology implicated in postpartum depression. Neurobiol Dis. 2022;165:105646. doi:10.1016/j.nbd.2022.105646
50. Levin G, Ein-Dor T. A unified model of the biology of peripartum depression. Transl Psychiatry. 2023;13(1):138. doi:10.1038/s41398-023-02439-w
51. Begum N, Mandhare A, Tryphena KP, et al. Epigenetics in depression and gut-brain axis: a molecular crosstalk. Front Aging Neurosci. 2022;14:1048333. doi:10.3389/fnagi.2022.1048333
52. da Silva IV, Garra S, Calamita G, et al. The multifaceted role of Aquaporin-9 in health and its potential as a clinical biomarker. Biomolecules. 2022;12(7):897. doi:10.3390/biom12070897
53. Wang H, Dou S, Wang C, et al. Identification and experimental validation of Parkinson’s disease with major depressive disorder common genes. Mol Neurobiol. 2023;60(10):6092–6108. doi:10.1007/s12035-023-03451-3
54. Zahl S, Skauli N, Stahl K, et al. Aquaporin-9 in the brain inflammatory response: evidence from mice injected with the parkinsonogenic toxin MPP. Biomolecules. 2023;13(4):588. doi:10.3390/biom13040588
55. Shi Y, Chen F, Wang Z, et al. Effect and mechanism of functional compound fruit drink on gut microbiota in constipation mice. Food Chem. 2023;401:134210. doi:10.1016/j.foodchem.2022.134210
56. Wan C, Shi Y, Zhao X, et al. Positive association between ALDH1A2 and schizophrenia in the Chinese population. Prog Neuropsychopharmacol Biol Psychiatry. 2009;33(8):1491–1495. doi:10.1016/j.pnpbp.2009.08.008
57. Jia Y, Cheng S, Liu L, et al. Association between birth by caesarian section and anxiety, self-harm: a gene-environment interaction study using UK Biobank data. BMC Psychiatry. 2023;23(1):237. doi:10.1186/s12888-023-04720-0
58. Das SC, Schulmann A, Callor WB, et al. Altered transcriptomes, cell type proportions, and dendritic spine morphology in hippocampus of suicide decedents. J Affect Disord. 2024;367:118–128. doi:10.1016/j.jad.2024.08.144
59. Wu W, Sun M, Chen F, et al. Microbiota metabolite short-chain fatty acid acetate promotes intestinal IgA response to microbiota which is mediated by GPR43. Mucosal Immunol. 2017;10(4):946–956. doi:10.1038/mi.2016.114
60. Wang Q-W, D-j-c J, He J-M, et al. Lactobacillus intestinalis primes epithelial cells to suppress colitis-related Th17 response by host-microbe retinoic acid biosynthesis. Adv Sci. 2023;10(36):e2303457. doi:10.1002/advs.202303457
61. Zhou X, Curbo S, Zhao Q, et al. Severe mtDNA depletion and dependency on catabolic lipid metabolism in DGUOK knockout mice. Hum Mol Genet. 2019;28(17):2874–2884. doi:10.1093/hmg/ddz103
62. Zhou X, Mikaeloff F, Curbo S, et al. Coordinated pyruvate kinase activity is crucial for metabolic adaptation and cell survival during mitochondrial dysfunction. Hum Mol Genet. 2021;30(21):2012–2026. doi:10.1093/hmg/ddab168
63. Gorman-Sandler E, Robertson B, Crawford J, et al. Gestational stress decreases postpartum mitochondrial respiration in the prefrontal cortex of female rats. Neurobiol Stress. 2023;26:100563. doi:10.1016/j.ynstr.2023.100563
64. Gutierrez-Zotes A, Diaz-Pena R, Costas J, et al. Interaction between the functional SNP rs2070951 in NR3C2 gene and high levels of plasma corticotropin-releasing hormone associates to postpartum depression. Arch Womens Ment Health. 2020;23(3):413–420. doi:10.1007/s00737-019-00989-x
65. Chen X, Liu Y, Pu J, et al. Proteomics reveals mitochondrial dysfunction and energy metabolism disturbance of intestine in a nonhuman primate model of depression. J Affect Disord. 2023;333:562–570. doi:10.1016/j.jad.2023.04.031
66. Bednash JS, Johns F, Patel N, et al. The deubiquitinase STAMBP modulates cytokine secretion through the NLRP3 inflammasome. Cell Signal. 2021;79:109859. doi:10.1016/j.cellsig.2020.109859
67. Brann E, Papadopoulos F, Fransson E, et al. Inflammatory markers in late pregnancy in association with postpartum depression-A nested case-control study. Psychoneuroendocrinology. 2017;79:146–159. doi:10.1016/j.psyneuen.2017.02.029
68. Li X, Mo Y, Shang S, et al. Gut Escherichia coli promotes lung cancer by increasing circulating STAMBP production. Discov Oncol. 2025;16(1):459. doi:10.1007/s12672-025-02206-x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Revealing the Impact of Gut Microbiota on Acne Through Mendelian Randomization Analysis

Ji X, Wu S, Zhao D, Bai Q, Wang Y, Gong K, Zheng H, Zhu M
Clinical, Cosmetic and Investigational Dermatology 2024, 17:383-393
Published Date: 8 February 2024
Genomics-Microbiome Based Assessment of Bidirectional Causality Between Gut Microbiota and Psoriasis
Gao Q, Liu JH, Ma WY, Cheng ZL, Hao PS, Luo NN
Clinical, Cosmetic and Investigational Dermatology 2024, 17:435-445
Published Date: 13 February 2024
Causal Relationships Between Gut Microbiota, Metabolites, and Diabetic Nephropathy: Insights from a Two-Sample Mendelian Randomization Analysis
Song X, Cui J, Li S, Huang B
International Journal of Nephrology and Renovascular Disease 2024, 17:319-332
Published Date: 10 December 2024
Exploring Causal Relationships Between Gut Microbiota, Inflammatory Cytokines, and Inflammatory Dermatoses: A Mendelian Randomization Study
Huang Z, Lu T, Lin J, Ding Q, Li X, Lin L
Clinical, Cosmetic and Investigational Dermatology 2025, 18:579-592
Published Date: 13 March 2025
The Gut Microbiota and the Nerve-Bone Axis: Insights from a Mendelian Randomization and Mediation Analysis
Qiao X, Lou Y, Chen H, Zhang H, Cao J, Feng X, Guo L, Pan Q
Journal of Multidisciplinary Healthcare 2025, 18:4233-4241
Published Date: 26 July 2025