Back to Journals » Journal of Pain Research » Volume 19
Experimental Gestational Diabetes Mellitus Prolongs Post-Incisional Mechanical Hypersensitivity and Increases Cutaneous Neuro-Immune Markers in Adult Female Offspring
Authors Cantú-Cepeda CA ![]() , Villarreal-Salcido J
, Villarreal-Salcido J ![]() , Elizondo-Martinez CE
, Elizondo-Martinez CE ![]() , Vazquez-Mora JA, Martínez-Martínez A, Acosta-González RI, Jiménez-Andrade JM, Muñoz-Islas E
, Vazquez-Mora JA, Martínez-Martínez A, Acosta-González RI, Jiménez-Andrade JM, Muñoz-Islas E ![]()
Received 3 February 2026
Accepted for publication 7 May 2026
Published 19 May 2026 Volume 2026:19 594897
DOI https://doi.org/10.2147/JPR.S594897
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Carlos A Cantú-Cepeda, Jaira Villarreal-Salcido, Cecilia Esther Elizondo-Martinez, Juan Antonio Vazquez-Mora, Arisai Martínez-Martínez, Rosa Issel Acosta-González, Juan Miguel Jiménez-Andrade, Enriqueta Muñoz-Islas
Laboratory of Pharmacology, Multidisciplinary Academic Unit Reynosa-Aztlan, Autonomous University of Tamaulipas, Reynosa, Tamaulipas, Mexico
Correspondence: Enriqueta Muñoz-Islas, Multidisciplinary Academic Unit Reynosa-Aztlan, Autonomous University of Tamaulipas, Calle 16 y Lago de Chapala S/N Col. Aztlán, Reynosa, Tamaulipas, 88740, México, Tel +525532076849, Email [email protected]
Purpose: Gestational diabetes mellitus (GDM) can lead to health complications in adult offspring. It was recently reported that experimental GDM increases intraepidermal nerve fiber density (INFD) in adult mouse offspring. However, the impact of GDM on acute postsurgical pain and the neuroimmune response to incision has not been examined.
Methods: GDM was induced through intraperitoneal administrations of streptozotocin (STZ) in ICR pregnant mouse dams. At 12 weeks old, all female mouse offspring underwent an incision in the glabrous skin of the right hindpaw. The mechanical sensitivity of both incised and non-incised hindpaws was assessed before and during the 3 weeks following the incision. At 16 weeks old, offspring born from dams with and without STZ-induced GDM were euthanized and processed for immunohistochemistry. The INFD (CGRP+ and PGP9.5+ axons) and the density of CD68+ macrophages around the wounded skin were quantified.
Results: Offspring mice of dams with STZ-induced GDM exhibited decreased body weight, yet their blood glucose levels were comparable to those of offspring born from vehicle-treated dams. These offspring of dams with STZ-induced GDM experienced significantly prolonged incision-induced mechanical hypersensitivity compared to offspring of control dams. At 3 weeks post-incision, offspring of STZ-treated dams had an increased density of macrophages and greater INFD of CGRP+ and PGP9.5+ axons compared to the values of offspring of vehicle-treated dams.
Conclusion: Offspring of dams with STZ-induced GDM exhibited a delayed resolution of post-incisional pain that was accompanied by increased epidermal nerve fiber density and peri-incisional macrophage activation.
Keywords: gestational diabetes mellitus, post-incisional, recovery, pain
Introduction
Gestational diabetes mellitus (GDM), a metabolic disorder characterized by glucose intolerance first diagnosed during pregnancy,1–3 represents a significant concern with short and long-term consequences for both maternal and neonatal health. Its acute complications, such as preeclampsia, fetal macrosomia, and birth injuries, are well-documented.4–6 Beyond these immediate outcomes, emerging evidence indicates that intrauterine exposure to a hyperglycemic environment can induce persistent physiological alterations in the offspring, predisposing them to metabolic and cardiovascular disorders later in life.6–8 This phenomenon, often referred to as fetal metabolic programming,9 highlights the profound role of the intrauterine environment in shaping long-term health trajectories.
Recent studies, including our own,10 have revealed that maternal hyperglycemia disrupts baseline nociceptive processing in adult offspring, and this effect appears to be independent of sex, which is accompanied by increased intraepidermal nerve fiber density (INFD) of calcitonin gene-related peptide (CGRP)- and protein gene product 9.5 (PGP9.5)-positive axons innervating skin.10 These findings suggest that GDM may permanently alter sensory innervation patterns, predisposing offspring to heightened mechanical and thermal sensitivity under baseline conditions.10,11 However, a critical question remains unexplored: Does GDM exacerbate or prolong pain-like responses in the offspring following tissue injury, such as a skin incision surgery?
Post-surgical pain recovery is a complex process influenced by neuroimmune interactions, yet the role of developmental programming in this context is poorly understood.12,13 While prior work has established that GDM offspring exhibit baseline nociceptive abnormalities, no studies have investigated whether these alterations translate into delayed recovery after surgical insult.9–11 This gap is clinically significant, as prolonged post-surgical pain in the offspring may reflect maladaptive neuroplasticity or dysregulated inflammatory mechanisms potentially rooted in gestational hyperglycemia.14,15
To address this, a widely accepted murine model of GDM was employed10,16,17 to evaluate the duration of mechanical hypersensitivity in the offspring following plantar incision,12 along with quantitative changes in a subset of sensory nerve fibers and macrophages at the wound site. The present study uniquely integrates behavioral pain assessments with immunohistochemical analyses to elucidate how GDM may alter post-injury recovery. It was hypothesized that offspring of dams with GDM would exhibit: (1) prolonged post-surgical mechanical hypersensitivity, (2) epidermal hyperinnervation, and (3) enhanced macrophage-driven inflammation compared to offspring of control dams.
Material and Methods
Animals and Experimental GDM Induction
To establish the GDM model, 60 timed-pregnant, nulliparous ICR mouse dams were utilized. These dams were bred with 20 male ICR mice (Bioinvert Laboratories, Mexico), all at an initial age of 8–10 weeks and with a body weight of 20–30 g. All animals were maintained under standard laboratory conditions: 22±2°C, 40–60% humidity, and a 12-hour light/12-hour dark cycle, with ad libitum access to food and water. Procedures involving mice and their care were conducted in conformity with the Mexican Official Norm for Animal Care and Handling (NOM- 062-ZOO-1999)18 and the Guide for the Care and Use of Laboratory Animals.19 The Institutional Research and Ethics Committee of Multidisciplinary Academic Unit Reynosa-Aztlan (Approval No. CEI-UAMRA-2022-0008) approved all experimental procedures. GDM was induced by intraperitoneal administrations of streptozotocin (STZ; Sigma-Aldrich, St. Louis, MO, USA), dissolved in a 0.1 M sodium citrate buffer (pH 4.5), at decreasing doses of 100 mg/kg on 6.5 and 7.5 gestation days and 80 mg/kg on 8.5 gestation day.16,17,20 Control dams received an equivalent volume of the vehicle solution. On gestation day 10.5, a 6-hour fasting period preceded serum glucose level assessment (Accu-Check Guide, Roche S.A. de C.V., Mexico) via a tail-vein blood sample. Dams with diabetes were considered and included in the GDM group if their fasting blood glucose levels were >200 mg/dL.21 Following weaning at 3–4 weeks of age, female offspring of dams with GDM and from control dams were maintained under standard housing conditions with weekly body weight monitoring until week 16. Blood glucose measurements were conducted at 12 and 16 weeks of age using prior standardized sampling protocols (Figure 1A).
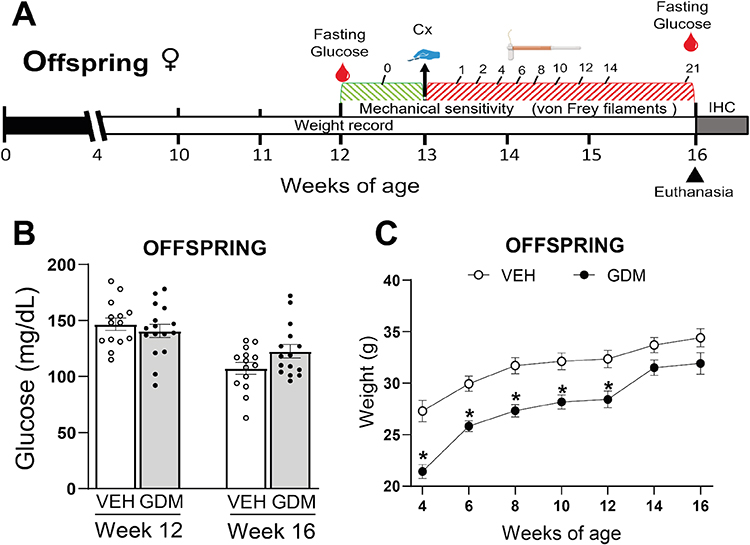 |
Figure 1 Experimental design, blood glucose levels, and body weight of female offspring from control dams or from dams with gestational diabetes mellitus. (A) The experimental timeline for the offspring is illustrated as follows: offspring are weaned at 4 weeks, followed by weekly weight measurements and a fasting glucose measurement at week 12. After baseline mechanical sensitivity recording, a right hindpaw plantar incision was performed at week 13, and the left hindpaw (contralateral) served as its control. Mechanical sensitivity in both hindpaws was evaluated over 21 subsequent days, concluding with a final glucose measurement and tissue collection for IHC. (B) Maternal streptozotocin (STZ) administration did not alter glucose concentrations in offspring at weeks 12 or 16. (C) Offspring of dams with gestational diabetes mellitus (GDM) exhibited significantly lower body weight from weeks 4 to 12. The data are presented as mean ± SEM; *p < 0.05 vs. vehicle (VEH)-treated using two-way RM ANOVA followed by the Tukey post-hoc analysis test. |
Plantar Incision Surgery
At 12 weeks of age, female mouse offspring underwent a unilateral plantar incision to induce acute pain, a procedure based on a previously established method.12 Animals were anesthetized with 2% isoflurane (SofloranVet, Reg. S.A.G.A.R.P.A. Q-7833-222, PiSA Agropecuaria, Mexico City) delivered in medical-grade oxygen (INFRA, Tamaulipas, Mexico) at a flow rate of 500 cc/min via a nose cone (VetFlo®, Kent Scientific Co., Torrington, CT, USA). Temperature was controlled at 37°C using an infrared thermometer gun (CEM DT-811, Kent Scientific Co., Torrington, CT, USA) and a temperature-controlled heating pad (Repti Therm UTH, #RH-4, ZooMed Laboratories Inc., CA, USA). The room temperature for all experiments was 22±2°C.
The right hindpaw’s plantar surface was cleansed with 10% povidone-iodine (Dermo Dine, Re. 0822C87 SSA, Mexico City) and 70% ethanol (JALMEK Científica, No. E5325-17, N.L. Mexico). A standardized plantar incision was performed. Briefly, a 5-mm longitudinal incision was made with a No. 11 blade (HERGOM; Reg. 2038E2014 SSA; Mexico City), traversing the skin, fascia, and muscle layers of the glabrous hindpaw. The incision was initiated approximately 2.5 mm from the heel and continued distally. The plantaris muscle was carefully elevated during the incision. Following the incision, the wound was apposed with two 6–0 nylon mattress sutures (TAGUM; R.S.: DM0100N; Mexico City), and antibiotic ointment (Neomycin, Tranexamic Acid; RECOVERON NC, Reg. 443M94 SSA IV; Armstrong; Mexico City) was applied to prevent infection. To minimize the number of offspring mice used in the present study, the left hindpaw (contralateral) served as a non-incised control.12,13
Evaluation of Hindpaw Mechanical Sensitivity in the Adult Offspring
Hindpaw mechanical sensitivity was evaluated using the up-down method with calibrated von Frey filaments (Stoelting Co., Wood Dale, IL, USA).10,11 Before testing, mice were acclimated for 30 minutes in elevated plastic cages with wire-mesh floors. The von Frey filaments were applied perpendicularly to the plantar surface of both the incised (right) and contralateral (left) hindpaws, with sufficient force to cause slight bending, for approximately 6 seconds. Withdrawal responses, including paw lifting, licking, or flinching, were recorded. Measurements were taken at baseline (pre-surgery at 12 weeks of age) and on postoperative days: 1, 2, 4, 6, 8, 10, 12, 14, and 21. The 50% probability of withdrawal threshold (in grams) for each paw was calculated using the formula described elsewhere.22 All behavioral assessments were conducted by an observer blinded to the animals’ GDM status.
Tissue Harvesting and Immunohistochemistry
At 16 weeks of age (21 days after incisional surgery), offspring were deeply anesthetized via intraperitoneal injection with a mixture of ketamine (KETANIL; Reg. Q-0609-004; Wildlife Pharmaceuticals, Mexico City) and xylazine (PROCIN; Reg. Q-7833-099; PiSA Agropecuaria, Mexico City) at a dose of 100/10 mg/kg, respectively. Adequate anesthesia was confirmed by the absence of withdrawal response to a firm toe pinch according to the Guidelines for the Euthanasia of Animals. Animals were transcardially perfused with 0.9% phosphate-buffered saline (PBS, 0.1 M, pH 7.4, 4°C), followed by 4% paraformaldehyde in PBS (Cat. No. F1635, Sigma-Aldrich, EEUU). Glabrous skin from both the incised and contralateral hindpaws was carefully harvested, post-fixed in 4% paraformaldehyde for 24 hours, and then cryoprotected by immersion in 30% sucrose in PBS for 48–72 hours. Tissue samples were then embedded in optimal cutting temperature (OCT) compound (Tissue-Tek, Sakura Finetek, Torrance, CA, USA) and snap frozen. Serial frozen sections (20 µm thick) were prepared using a cryostat instrument (Leica CM1950; Leica Biosystems, Wetzlar, Germany) and mounted onto Superfrost Plus glass microscope slides (Fisher Scientific, Waltham, MA, USA).
Immunohistochemical staining was performed as follows: sections were initially blocked for 1 hour at room temperature in PBS containing 0.3% Triton X-100 and 3% Normal Donkey Serum (Jackson ImmunoResearch Laboratories, West Grove, PA, USA). This was followed by overnight incubation at 4°C with primary antibodies. The primary antibodies used were: against CGRP (polyclonal rabbit anti-mouse 1:3,000; Sigma Aldrich, Cat# C8198) for primary sensory peptidergic nerve fibers; antibody against PGP9.5 (Protein gene product 1:3,000; Cedarlane: Cat# CL7756AP) as a pan-neuronal marker; and primary antibody against CD68 marker aimed towards macrophages (Macrosialin; monoclonal rat anti-mouse 1:3,000; Bio-Rad; Cat# CA1957). After primary antibody incubation, sections were washed thoroughly in PBS and then incubated for 3 hours at room temperature with the following secondary antibodies (Cy3 monoclonal donkey anti-rabbit 1:600; Jackson ImmunoResearch; Cat# 711–165-152 and Cy2 monoclonal donkey anti-rat 1:300; Jackson ImmunoResearch; Cat# 712–225-150). Following washes, nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI 1:20,000; Invitrogen; Cat# D21490). Slides were then washed, dehydrated through an ascending ethanol series, cleared with xylene, and coverslipped using DPX mounting medium. Images were captured using an epifluorescence microscope (Axio Scope A1, Carl Zeiss, Jenna, Germany) and a confocal laser microscope (LSM 800, Carl Zeiss, Jena, Germany). Confocal Z-stack images were acquired with a 20 µm slice thickness using a 20x objective for detailed analysis.
Quantification of the Density of Nerve Fibers and Macrophages Around the Wounded Skin
Quantification of nerve fiber and macrophage densities was performed by investigators blinded to the experimental conditions. Analyses were performed on within-animal comparison between incised (right hindpaw) and non-incised (left hindpaw) samples obtained from each animal (n = 5 GDM offspring; n = 6 VEH offspring). For nerve fibers, CGRP+ axons were quantified in the epidermis, and PGP9.5+ axons were quantified in both the epidermis and the superficial dermal region (up to 150 µm from the dermo-epidermal junction). A minimum of five distinct sections per mouse were analyzed at 20x magnification. Nerve fibers were manually traced using ImageJ software (National Institutes of Health, Bethesda, MD, USA), and the data were expressed as the mean total nerve fiber length (µm) per 100 µm2 of epidermis.23 For macrophage quantification, CD68-immunoreactive cell density was assessed specifically within the dermal region.24 Three confocal images were acquired per sample using the 20x objective lens. CD68+ profiles containing a DAPI-stained nucleus were meticulously counted at the incision site. The total number of cells was calculated and expressed per 1 mm3 of volume.
Statistical Analysis
Data is presented as mean ± standard error of the mean (SEM). The comparison of glucose measurements from the dams was performed using a Student’s t-test. Offspring body weight and mechanical withdrawal thresholds were analyzed using a two-way repeated-measure ANOVA, followed by Tukey’s post-hoc test when appropriate. Immunohistochemical data (nerve fiber and macrophage densities) were analyzed with a two-way ANOVA, followed by Tukey’s post-hoc test. A statistical significance level of p < 0.05 was accepted for all analyses. Statistical software (GraphPad Prism v8.0; GraphPad Software, San Diego, CA, USA) was used for all computations.
Results
Effect of Experimental GDM on Blood Glucose Levels and Body Weight in Dams and Offspring
Consistent with successful GDM induction, STZ-treated dams exhibited significantly elevated fasting blood glucose levels at gestational day 10.5 compared to vehicle-treated dams (303 ± 84.20 mg/dL vs. 123 ± 8.28 mg/dL; p < 0.05). In contrast, offspring from dams with STZ-induced GDM showed no significant differences in glucose levels at 12 or 16 weeks of age compared to those of offspring from vehicle-treated dams (Figure 1B). Offspring of dams with STZ-induced GDM had significantly reduced body weight compared to offspring from vehicle-treated dams between 4 and 12 weeks old. Although a trend to have lower body weight persisted at 14 and 16 weeks of age, these differences were not statistically significant (Figure 1C).
Experimental GDM Prolongs the Post-Surgery Pain Recovery in Adult Offspring
The assessment of baseline tactile thresholds revealed that offspring from dams with STZ-induced GDM had significantly lower withdrawal thresholds in both hindpaws than offspring from vehicle-treated dams (Figure 2A and C). After the plantar skin incision in the right hindpaw, mechanical sensitivity in offspring of dams with GDM did not return to baseline values until day 21 post-surgery. In contrast, offspring of vehicle-treated dams exhibited values of similar magnitude to baseline records at day 10 post-incision. Offspring of dams with STZ-induced GDM displayed a delayed recovery compared to offspring of control dams, starting from day 4 through 14 (Figure 2B). Additionally, no significant differences were observed in the contralateral paw (without incision) when comparing post-incision values to baseline measurements within each group (Figure 2C and D).
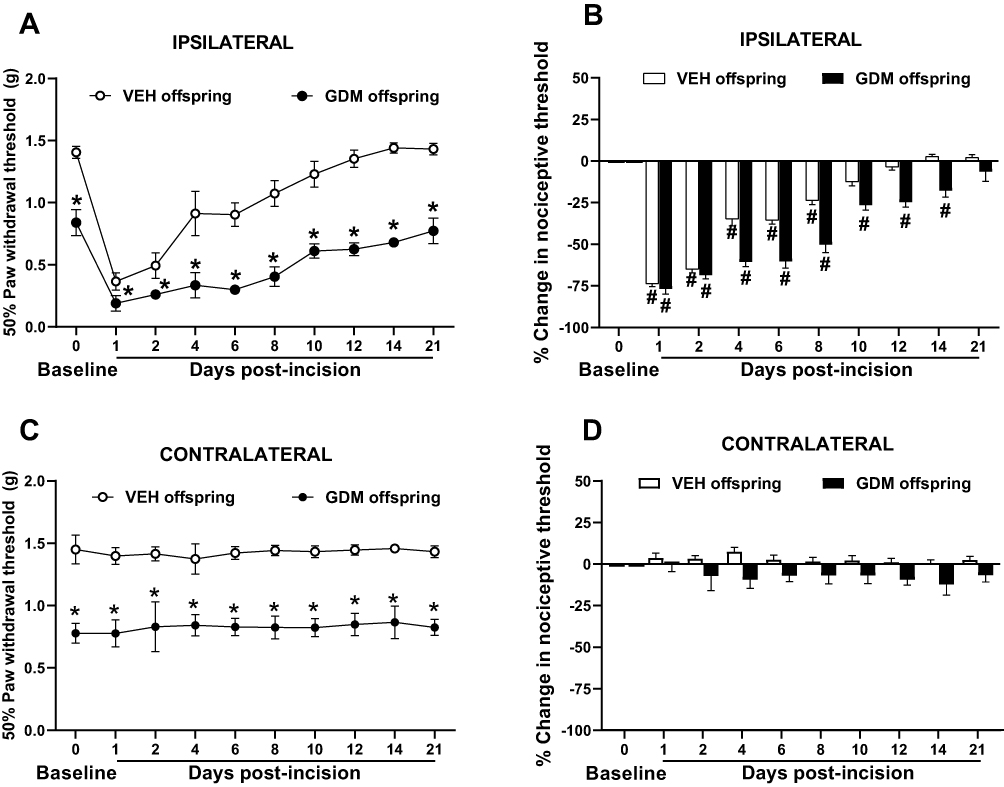 |
Figure 2 Gestational diabetes mellitus significantly delayed the resolution of mechanical sensitivity in adult offspring following skin incision. Repeated administration of streptozotocin (STZ) during gestation significantly reduces the 50% paw withdrawal threshold of the incised hindpaw of adult female offspring compared to values of offspring of vehicle (VEH)-treated dams, indicating greater surgery-induced mechanical hypersensitivity (A–C). Notably, this hypersensitivity persisted until 21 days post-incision in offspring from dams with STZ-induced GDM, whereas it resolved by day 10 in offspring from vehicle-treated dams. Additionally, from postoperative day 4 through 14, offspring of dams with gestational diabetes mellitus (GDM) showed a prolonged recovery period compared with offspring of dams without GDM (B). In contrast, mechanical sensitivity evaluations of the non-incised (contralateral) hindpaw showed no statistically significant changes in either experimental group following the plantar incision compared to their baseline values (C and D). Data is presented as mean ± SEM; *p < 0.05 vs. VEH offspring; #p < 0.05 vs. respective pre-incision values. Two-way RM ANOVA followed by Tukey’s post-hoc test. |
Experimental GDM Increases the Density of Intraepidermal CGRP+ and PGP9.5+ Nerve Fibers Around the Wounded Skin of the Hindpaw from Adult Offspring Following a Plantar Incision
Immunohistochemical analysis and confocal microscopy were employed to assess the density of CGRP, a neuropeptide primarily found in unmyelinated C-fiber and some thinly myelinated A-delta sensory nerve fibers, in the epidermis, as well as PGP9.5, a pan-neuronal marker, in the epidermis and upper dermis of the wounded skin from the offspring. Representative confocal images were taken of CGRP+ axons (Figure 3A–D) and PGP9.5+ axons (Figure 4A–D) in the hindpaw glabrous skin of 16-week-old offspring born from vehicle-treated and STZ-induced GDM dams. Consistent with previous reports,10 the basal densities of CGRP+ and PGP9.5+ were significantly higher in non-surgery offspring of dams with STZ-induced GDM compared to non-surgery offspring from vehicle-treated dams (Figures 3E and 4E). Additionally, in offspring from vehicle-treated dams, the nerve fiber density in the skin at 3 weeks post-incision was similar in magnitude compared to values of the non-incised hindpaw. Interestingly, in the incised hindpaw from offspring of dams with GDM, the density of CGRP+ and PGP9.5+ axons innervating skin at 3 weeks post-incision was significantly higher compared to the non-incised hindpaw from offspring of dams with GDM and compared to the incised hindpaw skin of offspring of vehicle-treated dams (Figures 3E and 4E).
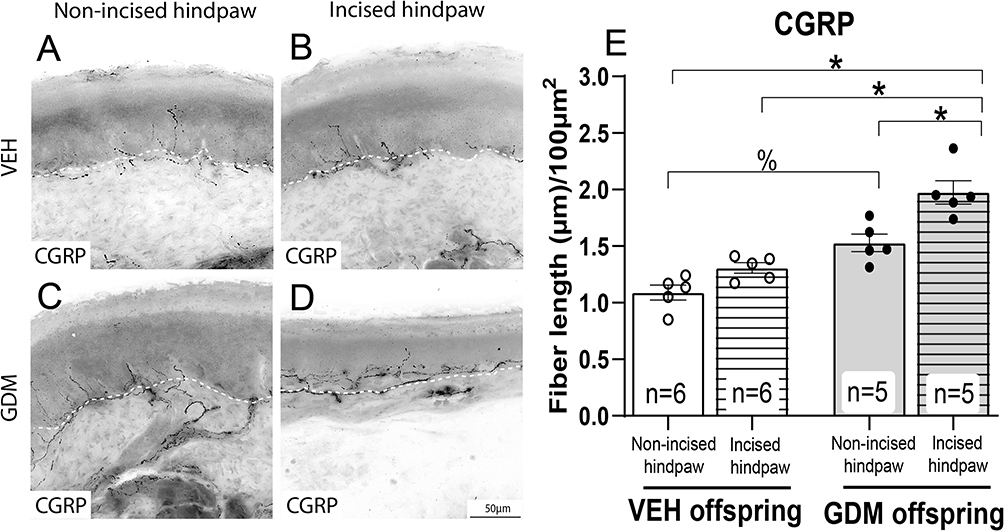 |
Figure 3 Adult offspring of dams with gestational diabetes mellitus have an increased density of intraepidermal CGRP+ nerve fibers at 3 weeks post-plantar incision. Representative confocal images of calcitonin gene-related peptide (CGRP)-positive axons in the glabrous hindpaw skin of 16-week-old female offspring. (A) Non-incised hindpaw of offspring of vehicle (VEH)-treated dams, (B) Incised hindpaw of offspring of VEH-treated dams, (C) Non-incised hindpaw of dams with gestational diabetes mellitus (GDM), (D) Incised hindpaw of offspring with GDM at 21 days post-incision. Scale bar = 50 µm (applies to all panels). (E) Quantitative analysis of CGRP⁺ intraepidermal nerve fiber density expressed as mean total fiber length (µm) per 100 µm2 of epidermis. The data are presented as mean ± SEM. *p < 0.05, %p < 0.05. Two-way ANOVA followed by the Tukey post-hoc test. |
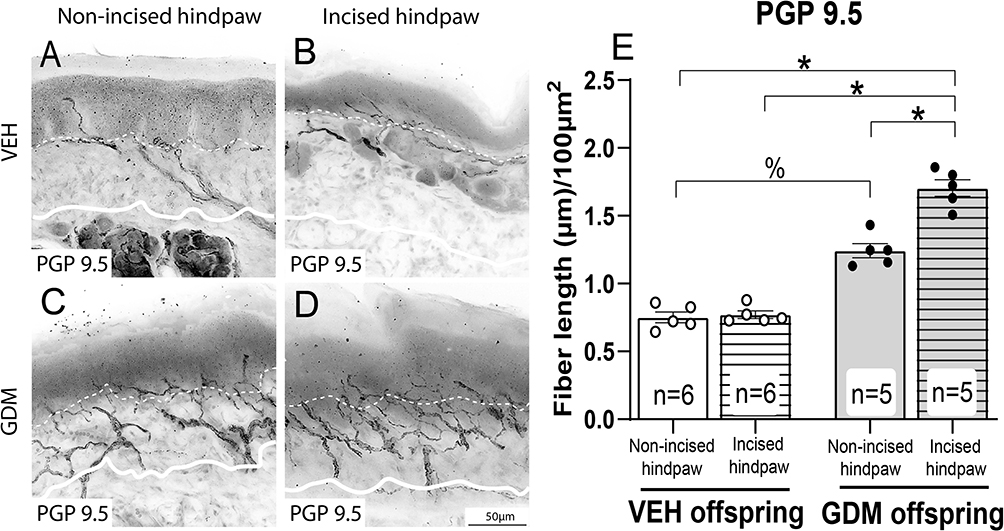 |
Figure 4 Adult offspring of dams with gestational diabetes mellitus display an increased density of PGP9.5+ axons in the intraepidermal and upper dermal regions at 3 weeks post-plantar incision. Representative confocal images of intraepidermal (dotted line) and upper dermal (solid line) protein gene product-9.5 (PGP9.5) in the hindpaw skin of offspring. (A) Non-incised hindpaw of offspring of vehicle (VEH)-treated dams, (B) Incised hindpaw of offspring of VEH-treated dams, (C) Non-incised hindpaw of dams with gestational diabetes mellitus (GDM), (D) Incised hindpaw of offspring with GDM at 21 days post-incision. (E) Quantitative analysis of PGP9.5+ intraepidermal and upper dermal nerve fiber density expressed as mean total fiber length (µm) per 100 µm2. The data are presented as mean ± SEM; *p < 0.05, %p < 0.05. Two-way ANOVA followed by the Tukey post-hoc test. |
Experimental GDM Increases the Density of Macrophages Around the Glabrous Hindpaw Skin of Adult Offspring Following a Plantar Incision
Representative confocal images and quantitative analyses showed no significant changes in the density of macrophages between offspring from vehicle-treated dams and from dams with STZ-induced GDM (Figures 5A, C, E) in the non-incised skin at 3 weeks post-incision. However, the plantar incision in the offspring of dams with STZ-induced GDM, but not in the offspring of dams without GDM, resulted in a significant increase in the macrophage density around the wound site (Figure 5B–E). The offspring of dams with STZ-induced GDM exhibited a significant increase in macrophage density post-incision compared to both their own contralateral hindpaw and the incised hindpaw of offspring from vehicle-treated dams (Figure 5E).
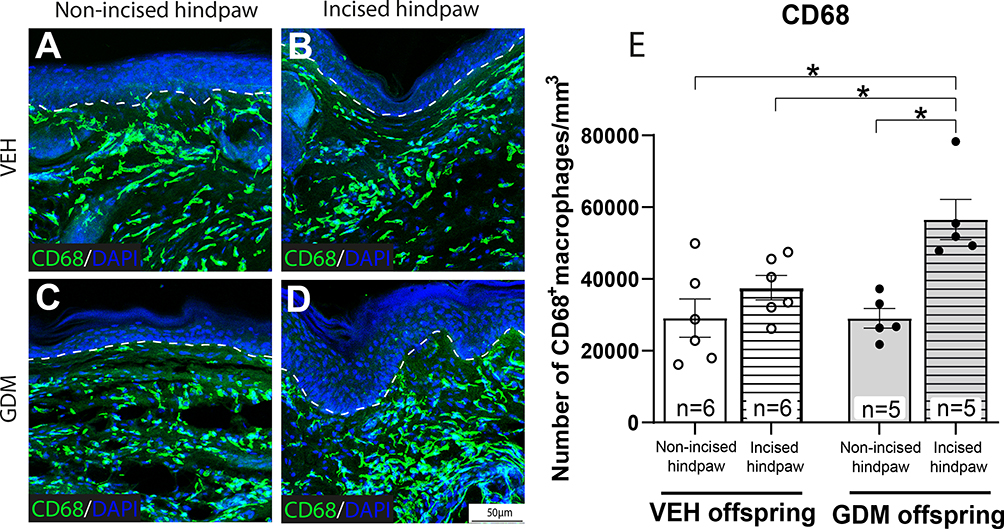 |
Figure 5 Adult offspring of dams with gestational diabetes mellitus exhibited an increased density of macrophages in the intradermal region at 3 weeks post-plantar incision. Representative confocal images of intradermal cells expressing cluster of differentiation 68 (CD68), indicating macrophages, in the hindpaw skin of offspring. (A) Non-incised hindpaw of offspring of vehicle (VEH)-treated dams, (B) Incised hindpaw of offspring of VEH-treated dams, (C) Non-incised hindpaw of dams with gestational diabetes mellitus (GDM), (D) Incised hindpaw of offspring with GDM at 21 days post-incision. (E) Quantitative analysis of CD68+ density expressed as number of macrophages per 1 mm3. The data are presented as mean ± SEM; *p < 0.05. Two-way ANOVA followed by Tukey post-hoc test. |
Discussion
This study reveals that GDM induces lasting alterations in pain processing and wound healing in adult offspring, as evidenced by (1) prolonged post-surgical mechanical hypersensitivity (21 days vs. 10 days in the offspring of control dams), (2) hyperinnervation of CGRP⁺ and PGP9.5⁺ nerve fibers at the incision site, and (3) enhanced CD68⁺ macrophage infiltration post-injury at 3 weeks post-incision. These findings highlight a novel link between developmental metabolic disturbances and maladaptive pain responses, offering critical insights into GDM’s long-term consequences.
Consistent with prior reports,10,21,25 offspring with GDM displayed reduced body weight but normoglycemia in adulthood, suggesting metabolic imprinting independent of glucose dysregulation. This aligns with the concept that fetal programming,9 in which in utero hyperglycemia triggers persistent physiological adaptations. Pancreatic β-cell overstimulation during gestation may impair insulin secretion and anabolism, contributing to offspring growth restriction.25 Crucially, our results indicate that these developmental changes may predispose offspring to delayed pain recovery post-surgery despite healthy metabolic profiles.
To investigate whether maternal GDM prolongs pain-like responses in the offspring following tissue insult, the plantar incision model12 was applied rather than chronic neuropathic (eg., Spared Nerve Injury; SNI)26 or inflammatory (eg., Complete Freund’s Adjuvant; CFA)27 models. Given that offspring with GDM already exhibit baseline mechanical hypersensitivity,10 using severe chronic injury models risks a nociceptive “ceiling effect” that could mask group differences.26,27 The plantar incision model mimics clinical post-operative pain and typically resolves within a week, providing a critical temporal window for assessing pain resolution.13 This approach uniquely enabled us to determine that intrauterine exposure to GDM not only alters baseline nociception but also fundamentally impairs the recovery from acute surgical trauma.
The delayed resolution of incision-induced pain in offspring of dams with STZ-induced GDM was accompanied by an increased density of CGRP⁺ and PGP9.5⁺ nerve fibers in the wounded skin, extending our earlier observations of baseline hyperinnervation.10,11 Similarly, in the early stages of the type 2 diabetes mouse model, mechanical allodynia has been linked to the sprouting of CGRP⁺ sensory fibers.28 This suggests that GDM not only alters baseline nociceptive pathways but may also modify the injury-induced neuroplasticity. Similar hyperinnervation of CGRP⁺ fibers has been reported in areas adjacent to non-healed fractures, a finding linked to chronic pain,29 supporting our hypothesis that aberrant hyperinnervation may underlie prolonged post-incision-induced mechanical hypersensitivity. A possible mechanism driving this nerve fiber sprouting might involve pathways mediated by Nerve Growth Factor (NGF). NGF, secreted by macrophages and keratinocytes,30 promotes axonal growth via TrkA receptors.31 In support of this, rodents with diabetes exhibit elevated NGF and TrkA⁺/CGRP⁺ fiber densities,24 and macrophage-derived NGF32,33 may further amplify post-injury hyperinnervation in offspring of dams with GDM. Future studies should quantify NGF levels in offspring of dams with GDM to validate this hypothesis.
Dermal macrophage density did not differ between non-incised skin from offspring with GDM compared to non-incised skin from offspring of control dams. This result contrasts with our previous observation, in which the exocrine pancreas from offspring with GDM exhibited a greater baseline density of CD68 macrophages than that from offspring of control dams,17 suggesting a GDM-induced tissue-specific response. However, the incised hindpaw of offspring from dams with STZ-induced GDM showed a significantly increased density of CD68⁺ macrophage at 3 weeks post-incision, a response absent in the incised skin of offspring from dams without GDM. This macrophage-driven inflammation likely exacerbates both hyperinnervation and prolonged mechanical hypersensitivity following skin incision. While the mechanisms behind this lasting inflammatory response in the offspring of dams with GDM following a skin incision are unknown, it is proposed that elevated advanced glycation end products (AGEs) and the advanced oxidation protein products (AOPPs) in offspring from dams with GDM34 may perpetuate inflammation via TNF-α/IL-1β.33,35 At the same time, oxidative stress36,37 and hematopoietic epigenetic changes14 could further delay the resolution of inflammation.
Our study has some limitations. Firstly, wound-healing dynamics were not assessed; thus, a delayed closure could confound the assessment of pain persistence. Secondly, the molecular mechanisms behind the hyperinnervation and persistent infiltration of CD68 macrophages at 3 weeks post-injection were not evaluated. Finally, sex-specific effects warrant investigation, as only female offspring were studied.
Conclusion
This study provides preclinical and initial evidence that maternal GDM exposure permanently modifies the offspring’s neuro-immune axis, resulting in a significantly delayed resolution of post-surgical mechanical hypersensitivity. Our findings also show that GDM exposure does not merely shift baseline sensitivity but fundamentally alters pain-like responses to tissue injury. Moreover, a key contribution of this work is the identification of a specific neuro-immune axis, characterized by exacerbated CGRP+ and PGP9.5+ innervation and persistent CD68+ macrophage infiltration in the incised skin at 3 weeks post-incision, as potential drivers of this maladaptive pain recovery. In conclusion, these findings underscore the critical need for clinical monitoring of GDM-exposed individuals in postoperative settings as well as the causes underlying these functional and anatomical findings, to develop mechanism-based therapies to improve pain resolution after surgery.
Abbreviations
GDM, Gestational diabetes mellitus; VEH, Vehicle; INFD, Intraepidermal nerve fiber density; STZ, Streptozotocin; CGRP, Calcitonin gene-related peptide; PGP9.5, Protein gene product 9.5; NGF, Nerve growth factor; TrkA, Tropomyosin receptor kinase A; AGEs, Advanced Glycation End Products; AOPPs, Advanced Oxidation Protein Products; TNF-α, Tumor Necrosis Factor alpha; IL-1β, Interleukin-1 beta.
Acknowledgments
EMI and JMJA designed and coordinated the project; CACC, JVS, CEEM, JAVM, AMM, and RIAG performed the experiments, acquired and analyzed data; EMI, JMJA, CACC, JVS, and CEEM drafted the manuscript; and all authors contributed to interpreting the results, critically reviewing the paper, and approving the final version of the article.
We would like to thank Christopher M. Peters for the English language editing.
Funding
This work was funded by the Ministry of Science, Humanities, Technology, and Innovation of Mexico (SECIHTI), grant number CBF-2025-I-3397.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Magliano DJ, Boyko EJ. scientific committee. IDF Diabetes Atlas. In: IDF Diabetes Atlas
2. Tsakiridis I, Giouleka S, Mamopoulos A, et al. Diagnosis and management of gestational diabetes mellitus: an overview of national and international guidelines. Obstet Gynecol Surv. 2021;76(6):367–11. doi:10.1097/OGX.0000000000000899
3. ElSayed NA, Aleppo G, Aroda VR, et al. 2. Classification and diagnosis of diabetes: standards of care in Diabetes—2023. Diabetes Care. 2023;46(Supplement_1):S19–s40. doi:10.2337/dc23-S002
4. Ye W, Luo C, Huang J, Li C, Liu Z, Liu F. Gestational diabetes mellitus and adverse pregnancy outcomes: systematic review and meta-analysis. BMJ. 2022;377:e067946. doi:10.1136/bmj-2021-067946
5. Metzger BE, Lowe LP, Dyer AR, et al. Hyperglycemia and adverse pregnancy outcomes. N Engl J Med. 2008;358(19):1991–2002.
6. Moon JH, Jang HC. Gestational diabetes mellitus: diagnostic approaches and maternal-offspring complications. Diabetes Metab J. 2022;46(1):3–14. doi:10.4093/dmj.2021.0335
7. Sweeting A, Wong J, Murphy HR, Ross GP. A clinical update on gestational diabetes mellitus. Endocr Rev. 2022;43(5):763–793. doi:10.1210/endrev/bnac003
8. Zehravi M, Maqbool M, Ara I. Correlation between obesity, gestational diabetes mellitus, and pregnancy outcomes: an overview. Int J Adolesce Med Health. 2021;33(6):339–345. doi:10.1515/ijamh-2021-0058
9. Monteiro LJ, Norman JE, Rice GE, Illanes SE. Fetal programming and gestational diabetes mellitus. Placenta. 2016;48(Suppl 1):S54–s60. doi:10.1016/j.placenta.2015.11.015
10. Munoz-Islas E, Elizondo-Martinez CE, Gutierrez-Lopez M, et al. Effect of experimental gestational diabetes mellitus on mechanical sensitivity, capsaicin-induced pain behaviors and hind paw glabrous skin innervation of male and female mouse offspring. J Pain Res. 2021;14:1573–1585. doi:10.2147/JPR.S313467
11. Campos Lima T, Lemes JBP, Capop T, de Lima LB, Monteiro da Cruz Lotufo C. Altered morphology and function of the peripheral nociceptive system in the offspring of diabetic rats. Int J Dev Neurosci. 2020;80(4):267–275. doi:10.1002/jdn.10023
12. Pogatzki EM, Raja SN. A mouse model of incisional pain. Anesthesiology. 2003;99(4):1023–1027. doi:10.1097/00000542-200310000-00041
13. Brennan TJ, Zahn PK, Pogatzki-Zahn EM. Mechanisms of incisional pain. Anesthesiol Clin North Ame. 2005;23(1):1–20. doi:10.1016/j.atc.2004.11.009
14. Govindarajah V, Sakabe M, Good S, et al. Gestational diabetes in mice induces hematopoietic memory that affects the long-term health of the offspring. J Clin Invest. 2024;134(2). doi:10.1172/JCI169730
15. Bridge-Comer PE, Vickers MH, Ferraro S, Pagnon A, Reynolds CM, Sigaudo-Roussel D. Maternal intake of either fructose or the artificial sweetener acesulfame-k results in differential and sex-specific alterations in markers of skin inflammation and wound healing responsiveness in mouse offspring: a pilot study. Nutrients. 2023;15(11):2534. doi:10.3390/nu15112534
16. Hokke SN, Armitage JA, Puelles VG, et al. Altered ureteric branching morphogenesis and nephron endowment in offspring of diabetic and insulin-treated pregnancy. PLoS One. 2013;8(3):e58243. doi:10.1371/journal.pone.0058243
17. Muñoz-Islas E, Santiago-SanMartin ED, Mendoza-Sánchez E, et al. Long-term effects of gestational diabetes mellitus on the pancreas of female mouse offspring. World J Diabetes. 2024;15(4):758–768. doi:10.4239/wjd.v15.i4.758
18. SAGARPA. NOM-062-ZOO-1999 Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Diario Oficial de la Federación México. 1999.
19. National Research Council. Guide for the Care and Use of Laboratory Animals.
20. Muñoz-Islas E, Vargas-Balderas DI, Hernandez I, Vazquez-Mora JA, Acosta-González RI, Jiménez-Andrade JM. Long-term effects of streptozotocin-induced gestational diabetes mellitus on mechanical sensitivity and intraepidermal nerve fibers in female and male mice offspring. Neurosci lett. 2023;812:137402. doi:10.1016/j.neulet.2023.137402
21. Huang H, Apaijai N, Thonusin C, et al. Mothers with obesity and gestational diabetes did not induce brain pathologies or premature brain aging in their adolescent and early adult offspring in rats. Neuroscience. 2025;568:454–464. doi:10.1016/j.neuroscience.2025.01.056
22. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53(1):55–63. doi:10.1016/0165-0270(94)90144-9
23. Pham VM, Tu NH, Katano T, et al. Impaired peripheral nerve regeneration in type-2 diabetic mouse model. Eur J Neurosci. 2018;47(2):126–139. doi:10.1111/ejn.13771
24. Arora V, Morado-Urbina CE, Gwak YS, et al. Systemic administration of a β2-adrenergic receptor agonist reduces mechanical allodynia and suppresses the immune response to surgery in a rat model of persistent post-incisional hypersensitivity. Molecular Pain. 2021;17:1744806921997206. doi:10.1177/1744806921997206
25. Huerta-Cervantes M, Pena-Montes DJ, Montoya-Perez R, et al. Gestational diabetes triggers oxidative stress in hippocampus and cerebral cortex and cognitive behavior modifications in rat offspring: age- and sex-dependent effects. Nutrients. 2020;12(2):376. doi:10.3390/nu12020376
26. Guida F, De Gregorio D, Palazzo E, et al. Behavioral, biochemical and electrophysiological changes in spared nerve injury model of neuropathic pain. Int J Mole Sci. 2020;21(9):3396. doi:10.3390/ijms21093396
27. Loredo-Pérez AA, Montalvo-Blanco CE, Hernández-González LI, et al. High-fat diet exacerbates pain-like behaviors and periarticular bone loss in mice with CFA-induced knee arthritis. Obesity. 2016;24(5):1106–1115. doi:10.1002/oby.21485
28. Cheng HT, Dauch JR, Hayes JM, Hong Y, Feldman EL. Nerve growth factor mediates mechanical allodynia in a mouse model of type 2 diabetes. J Neuropathol Exp Neurol. 2009;68(11):1229–1243. doi:10.1097/NEN.0b013e3181bef710
29. Chartier SR, Thompson ML, Longo G, Fealk MN, Majuta LA, Mantyh PW. Exuberant sprouting of sensory and sympathetic nerve fibers in nonhealed bone fractures and the generation and maintenance of chronic skeletal pain. Pain. 2014;155(11):2323–2336. doi:10.1016/j.pain.2014.08.026
30. Di Marco E, Marchisio PC, Bondanza S, Franzi AT, Cancedda R, De Luca M. Growth-regulated synthesis and secretion of biologically active nerve growth factor by human keratinocytes. J Biol Chem. 1991;266(32):21718–21722. doi:10.1016/S0021-9258(18)54695-0
31. Evans L, Andrew D, Robinson P, Boissonade F, Loescher A. Increased cutaneous NGF and CGRP-labelled trkA-positive intra-epidermal nerve fibres in rat diabetic skin. Neurosci Lett. 2012;506(1):59–63. doi:10.1016/j.neulet.2011.10.049
32. Kawamoto K, Matsuda H. Nerve growth factor and wound healing. Prog Brain Res. 2004;146:369–384.
33. Lewin GR, Ritter AM, Mendell LM. Nerve growth factor-induced hyperalgesia in the neonatal and adult rat. J Neurosci. 1993;13(5):2136–2148. doi:10.1523/JNEUROSCI.13-05-02136.1993
34. Boutzios G, Livadas S, Piperi C, et al. Polycystic ovary syndrome offspring display increased oxidative stress markers comparable to gestational diabetes offspring. Fertil Steril. 2013;99(3):943–950. doi:10.1016/j.fertnstert.2012.10.050
35. Roszer T. Inflammation as death or life signal in diabetic fracture healing. Inflamm Res. 2011;60(1):3–10. doi:10.1007/s00011-010-0246-9
36. Witko-Sarsat V, Gausson V, Nguyen A-T, et al. AOPP-induced activation of human neutrophil and monocyte oxidative metabolism: a potential target for N-acetylcysteine treatment in dialysis patients. Kidney Int. 2003;64(1):82–91. doi:10.1046/j.1523-1755.2003.00044.x
37. Steiling H, Munz B, Werner S, Brauchle M. Different types of ROS-scavenging enzymes are expressed during cutaneous wound repair. Exp Cell Res. 1999;247(2):484–494. doi:10.1006/excr.1998.4366
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Effect of a Local Anesthetic Injection Kit on Pain Relief and Postoperative Recovery After Transumbilical Single-Incision Laparoscopic Cholecystectomy
Yang N, Tao QY, Niu JY, Sun H, He Y, Hou YB, Luo H, Zhang Z, Yu JM
Journal of Pain Research 2023, 16:2791-2801
Published Date: 11 August 2023
Predictive Clinical Factors of Pain-Related Quality of Recovery Following Elective Gastrointestinal and Hepato-Pancreato-Biliary Surgery: An Observational Study in Norway
Bergestuen L, Hagen M, Kisa S
Journal of Pain Research 2025, 18:47-59
Published Date: 8 January 2025