Back to Journals » International Journal of Nanomedicine » Volume 20
Exosomes-Based Nanotherapeutic Strategies: An Important Approach for Spinal Cord Injury Repair
Authors Ju C, Dong H, Liu R, Wang X, Xu R, Hu H, Hao D
Received 11 May 2025
Accepted for publication 15 August 2025
Published 27 August 2025 Volume 2025:20 Pages 10407—10431
DOI https://doi.org/10.2147/IJN.S539673
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Krishna Nune
Cheng Ju,1,2,* Hui Dong,1,2,* Renfeng Liu,1,2,* Xuan Wang,1,2 Ruiqing Xu,1,2 Huimin Hu,1,2 Dingjun Hao1,2
1Department of Spine Surgery, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China; 2Shaanxi Key Laboratory of Spine Bionic Treatment, Honghui Hospital, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dingjun Hao, Email [email protected] Huimin Hu, Email [email protected]
Abstract: The repair and functional regeneration of spinal cord injury (SCI) remains a major challenge and focal point in regenerative medicine. Following SCI significant inflammation and neuronal damage occur. Conventional drug therapies often fail to precisely target the injured areas and cannot cross the blood-spinal cord barrier, severely limiting therapeutic efficacy. Therefore, precision therapeutics are crucial to improve the prognosis of SCI patients. In recent years, exosomes have gained widespread attention as natural delivery vehicles due to their low immunogenicity, high biocompatibility, and efficient delivery capabilities. Exosomes can effectively cross cell membranes and target specific cells, playing an important role in intercellular signaling. This makes them highly promising for precision therapies in SCI. By engineering exosomes for targeted delivery, new strategies can be developed for drug delivery, gene therapy, and personalized treatment after SCI. We aimed to review the biological functions of exosomes derived from different cell sources and discuss the role in tissue repair following SCI. Additionally, we explore the prospects and potential of exosomes in clinical SCI applications, to provide valuable research insights to improve functional recovery and long-term health management for SCI patients in the future.
Keywords: spinal cord injury, exosomes, nanodelivery, inflammation, nerve injury, non-coding RNA
Introduction
Spinal cord injury (SCI) results in immediate damage to neurons and axons, triggering a cascade of secondary pathological events.1 These events lead to sensory, motor, and autonomic dysfunction below the level of injury. Furthermore, high-level SCI may cause respiratory difficulties.2 SCI can be classified into complete and incomplete injuries based on the location and severity of the damage. Complete SCI results in total loss of function below the level of injury, leaving patients unable to perceive or control the parts of the body below the injury site. In contrast, patients with incomplete SCI may retain some function below the injury level, maintaining the ability to perceive certain sensations or partial motor capabilities.3 Currently, the incidence of new SCI cases continues to rise globally, imposing a significant economic burden on patients and society.4 The prognosis of SCI depends on the severity of the injury and the extent of preserved function. The pathological process of SCI can be divided into primary and secondary injuries. Primary injury is an instantaneous and irreversible process caused by initial mechanical compression, while secondary injury involves inflammatory infiltration and neuronal apoptosis, which can persistently expand the lesion area, further exacerbating the progression of SCI.5 Surgical intervention and hormone therapy remain the primary approaches during the acute phase following SCI.6 However, despite advancements in the treatment of SCI, the limited regenerative capacity of the spinal cord means that there is currently no fully effective treatment capable of completely restoring spinal cord function, and clinical outcomes remain poor. Long-term health management for patients poses significant challenges. Consequently, the search for safe, effective, and sustainable long-term treatment methods has become a crucial area of research.
The Main Pathological Processes of Secondary Injury
Damage to Blood Vessels, Ischemia, and Hypoxia After SCI
The blood supply to the spinal cord is disrupted during the early stages of primary injury. Due to excessive hemorrhage and neurogenic shock, patients with SCI often experience impaired perfusion and ischemia at the lesion site. After the blood vessels are damaged, the activation of multiple pathways can lead to systemic damage, causing a significant increase in extracellular glutamate levels, triggering excitotoxicity.7 Additionally, blood perfusion in the tissue can increase the permeability of the blood-spinal cord barrier (BSCB), promote the production of reactive oxygen species (ROS), and induce inflammation. Direct alterations to the BSCB endothelial cells can also facilitate the infiltration of immune mediators, leading to edema.8
The Inflammatory Response After SCI
Inflammatory response is a critical component of secondary injury, with inflammation infiltration dependent on the time course of injury and the activation state of immune cells.9 Following SCI, astrocytes, resident microglia, endothelial cells, and peripherally derived immune cells secrete harmful products, mediating activation of the immune system and the inflammatory response. Some studies indicate that inflammatory cytokines are upregulated within hours after injury, subsequently triggering the infiltration of exogenous and local cells.10 Moreover, these infiltrating immune cells can further produce and secrete other chemokines and cytokines that regulate the immune response, creating a detrimental inflammatory cycle.10 During the initial phase of inflammation (0–2 days), astrocytes, blood-derived neutrophils, and microglia are recruited to the injured area. Approximately 3 days post-injury, the second phase begins, during which circulating phagocytes and lymphocytes are recruited to the injury site. Activated macrophages, microglia, and other cells can stimulate T lymphocytes, thereby contributing to immune protection.11 CD4+ helper T lymphocytes promote the generation of B lymphocytes by producing various factors.12 The inflammatory response triggered by these cells plays a critical role in the progression of SCI.
Cell Death After SCI
Cell death is a critical process in secondary injury. Furthermore, as a result of mechanical damage caused by primary injury, the death of neurons and glial cells continues throughout the acute and subacute stages of injury.13 Necrosis and apoptosis are two important mechanisms of cell death.14 Necrosis is triggered by various factors, including the accumulation of blood-derived toxic substances, increased glutamate, ion imbalance, and excessive ATP depletion.15 Apoptosis is a programmed cell death process that can occur through extrinsic and intrinsic pathways based on the triggering mechanisms. The extrinsic pathway is activated by triggering death receptors, while the intrinsic pathway is regulated through the dynamic coordination of intracellular apoptosis-related proteins.16 Apoptosis typically occurs in areas distant from the injury site. In rat models of SCI, apoptosis can occur as early as 4 hours post-injury and peaks at 7 days.17 Therefore, modulating the necroptosis pathway appears to be a strategy for neuroprotection following SCI.
Scar Formation After SCI
Following SCI, astrocytes, pericytes, and fibroblasts are activated, promoting the development of glial/fibrotic scars. The upregulation of TGF-β enhances astrocyte activation and the subsequent formation of glial scar borders, as well as the deposition of fibronectin and laminin.18 Additionally, the transcription factor STAT3 plays a crucial role in establishing the glial scar border, which helps isolate infiltrating cells from entering the lesion core.19 However, the role of scars is complex. Glial scars have beneficial effects, such as repairing the damage to the BSCB caused by inflammation and isolating toxic substances from the injury site. Astrocytes simultaneously support axonal growth.20 Conversely, persistent scar formation induced by astrocytes impedes the regeneration and repair process of the spinal cord during the subacute and chronic stages of injury.11 Secretory products like proteoglycans produced by activated astrocytes can also interact with various harmful molecules and exert detrimental effects.21 The neural plasticity of mammals has been increasingly supported by numerous studies. When reactive astrocytes are transplanted into a naive spinal cord, they revert to a naive astrocyte state. Similarly, when they are transplanted into an injury site, they transform into scar-forming astrocytes.22 This indicates that the environment determines the phenotype of astrocytes, thereby influencing the inhibitory functions mediated by glial scars. In summary, SCI can be caused by a multitude of factors including traffic accidents, falls from height, violent injury, and sports injury. After SCI, a series of pathological changes occur in the injured area, leading to abnormal responses in multiple body systems (Figure 1, All figures were drawn by Figdraw).
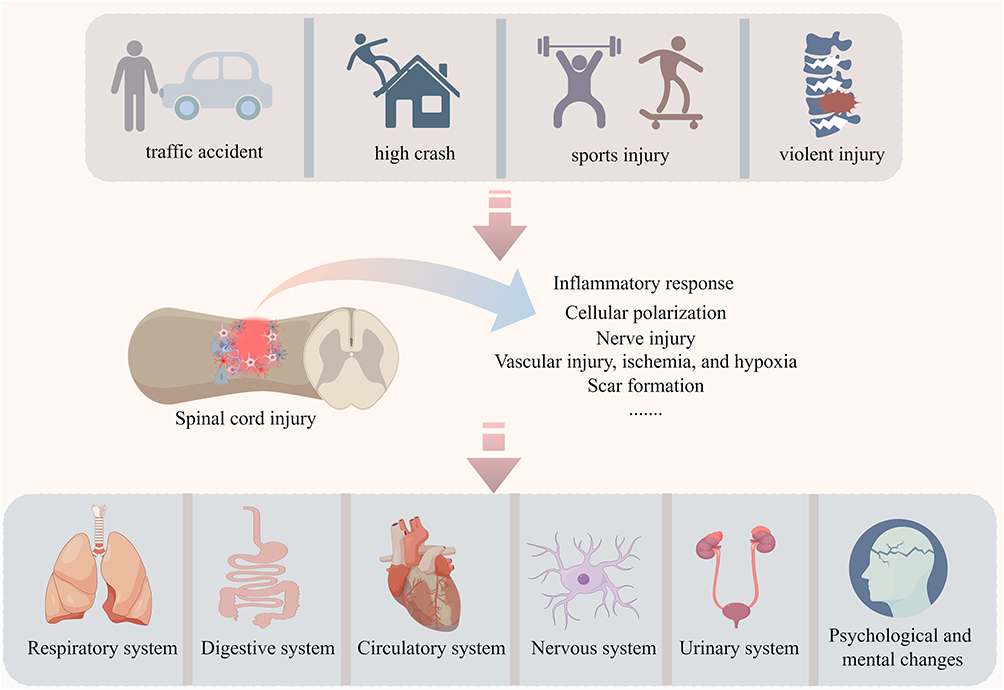 |
Figure 1 SCI can be caused by various factors such as traffic accident, high crash, violent injury, and sports injury. After SCI, the injured area undergoes a series of pathological changes, including inflammatory responses, cell polarization, neural damage, vascular injury, ischemia, hypoxia, and scar formation. Additionally, SCI can trigger abnormal reactions and alterations in multiple systems, including the respiratory, digestive, circulatory, nervous, and urinary systems, as well as psychological and mental health. |
Exosomes as Acellular Therapy
The pathophysiological mechanisms following SCI are complex, involving the participation of various cells in different pathological processes within the injury zone.23 Moreover, the functions of these cells and their signaling communication vary across different stages of injury. With advancements in medical technology, emerging therapies have brought potential hope for the treatment of SCI. Exosomes, leveraging their natural biological properties, hold significant potential in terms of safety, targeting ability, drug delivery efficiency, and functional diversity, particularly in the fields of precision medicine and personalized therapy.24 As naturally occurring nanoscale vesicles derived from cells, exosomes carry a variety of proteins, lipids, nucleic acids, and bioactive small molecules, enabling intercellular communication and material exchange.25 Extracellular vesicles (EVs) are membranous vesicles commonly found in a variety of body fluids and cells.26 Typical EVs can be divided into three types: microvesicles, exosomes, and apoptotic bodies.27 Studies have focused on the two main types of EVs (exosomes and microvesicles), and the release of these vesicles plays an important role in signal communication and molecular transmission between cells.28 Exosomes (approximately 50–100 nm in diameter) have been reported as acellular structures and are formed by the invagination of the membrane into endosomes, followed by the formation of multivesicular bodies, which fuse with the plasma membrane, releasing EVs called exosomes.29 Exosomes are widely distributed in the human body, and can be obtained from bodily fluid systems such as blood, urine, sweat, saliva, cerebrospinal fluid, and milk. The most commonly used method to obtain them is ultracentrifugation.30 Exosomes are composed of many types of proteins, lipids and nucleic acids. The proteins include major histocompatibility complex (MHC)-II, integrin, differentiation cluster, four transmembrane proteins and heat shock protein (Hsp). Lipids mainly include sphingomyelin and cholesterol. Nucleic acids include miRNA, circRNA, mRNA and lncRNA.31 The source, extraction, and structure of exosomes are described in Figure 2. Exosomes can achieve intercellular communication and material exchange functions, and their main mechanism of action is through receptor ligand binding, direct membrane fusion, and endocytosis. Exosomes are also closely related to the progression of various diseases, including cancer, immune diseases, and neurological diseases.32–34 Exosomes also lack growth and proliferation ability, have low tumor risk, high membrane transport efficiency, and can cross the BSCB to reach the injured site of the spinal cord. Therefore, exosomes may have great potential and advantages in the treatment of SCI. The research progress of exosomes in SCI treatment can be regarded from three viewpoints: the source of exosomes, the non-coding RNA in exosomes and the engineering modification of exosomes. Exosomes strongly associated with the progression of various diseases, including cancer, immune disorders, and neurological diseases. Following SCI, exosomes play a pivotal role in promoting neuroprotection, alleviating inflammatory responses, and enhancing the regenerative capacity of neurons. Additionally, exosomes can modulate the activity of immune cells such as glial cells and macrophages, thereby contributing to the improvement of the post-SCI immune microenvironment and reducing the occurrence of secondary injuries.
 |
Figure 2 The origin, extraction, and structure of exosomes. |
Local ischemia after SCI is a critical factor that hampers neural repair. Exosomes, carrying various small bioactive molecules, can enhance vascular stability and reduce the disruption of the BSCB. They also promote endothelial cell migration and angiogenesis, which are essential for restoring blood supply and supporting tissue regeneration. Moreover, exosomes exhibit high biocompatibility and low immunogenicity, allowing them to avoid the adverse effects commonly associated with traditional drugs and therapies. Their natural lipid bilayer structure enables exosomes to maintain stability in vivo and effectively cross the blood–brain barrier (BBB), a property crucial for treating neurological conditions like SCI. This appears to offer new strategies for the treatment of SCI.
The Function of Exosomes From Different Sources in SCI Diseases
Schwann Cell Derived Exosomes
In recent years, increasing evidence has demonstrated a connection between mitochondrial dysfunction and acute central nervous system (CNS) injury.35 Therefore, maintaining normal mitochondrial function is crucial for neuronal cell stability. Schwann cells (SCs) contribute positively to nerve repair by clearing myelin sheath and axonal debris, as well as facilitating axonal growth and proliferation.36 Recently study indicated that SCs-Exo can stimulate mitochondrial autophagy mediated by the AMPK pathway, thereby reducing oxidative stress, inflammation, and cell necrotic apoptosis.37 Pan et al showed that SCs-Exo suppress suppress the expression of EGFR and inhibit the Akt/mTOR signaling pathway, thereby enhancing autophagy levels and exerting a protective effect on SCI.38 Furthermore, SCs-Exo treatment inhibits the occurrence of M1 polarization in LPS-stimulated bone marrow-derived macrophages (BMDMs). Additionally, a composite hydrogel patch prepared with SCs-Exo and methylprednisolone can promote the transition of M1 macrophages to the M2 phenotype, suppress neuronal apoptosis, and enhance neuronal survival.39,40 The delivery of SCs-Exo to astrocytes can increase TLR2 expression and reduce CSPG deposition in the injured area. Intravenous injection of SCs-Exo effectively decreases scar formation following SCI.41,42 In terms of angiogenesis, SCs-Exo can be internalized by bEnd.3 cells and promote angiogenesis via integrin-β1.43
Human Umbilical Cord Mesenchymal Stem Cells Derived Exosomes
Umbilical cord mesenchymal stem cells (UCMSCs) versatile stem cells found in neonatal umbilical cord tissue, are capable of differentiating into various types of tissue cells with high differentiation potential. UCMSCs hold significant clinical promise in tissue engineering applications, including the regeneration of bone, cartilage, muscle, tendon, ligament, nerve, liver, endothelium, and myocardium tissues.44,45 However, the function of hUCMSCs derived exosomes (hUCMSCs-Exo) in SCI remains unclear. Exosomes from hUCMSCs have great therapeutic potential in improving neuronal recovery after SCI. hUCMSCs-Exo decreased the activation of microglia and astrocytes in mice, and increased the motor function of SCI mice.46 In in vitro and in vivo experiments, hUCMSCs-Exo effectively reduced the protein and mRNA expression levels of neurotoxic astrocyte markers C3 and Lcn2. Further mechanistic studies revealed that miR-146a-5p, by binding to IRAK1 and TRAF6, inhibits the expression of target genes, which is a crucial pathway through which hUCMSCs-Exo promote the recovery of motor function in mice.47 In neurons, hUCMSCs-Exo- miR-146b inhibits the activation of the NF- κB pathway by targeting TLR4, which can significantly alleviate the inflammatory injury to PC12 cells.48 In addition, hUCMSCs-Exo can promote the transformation of M1 macrophages into M2 macrophages by downregulating inflammatory cytokines such as TNF-α, MIP-1α, IL-6, and IFN-γ, as well as reducing ROS production. This process inhibits microglial apoptosis, thereby alleviating the inflammatory environment in the injured area.49,50 Isolating CD146+ CD271+ umbilical cord mesenchymal stem cells and obtaining hUCMSCs-Exo can reduce endothelial cell permeability, promote tight cell junctions, and enhance angiogenesis and BSCB integrity in the injured area following SCI.51 In summary, current findings indicate that hUCMSCs exert beneficial effects on inflammatory response, vascular regeneration, and neural recovery following SCI.
Endothelial Progenitor Cells Derived Exosomes
Endothelial progenitor cells (EPCs) are mainly derived from bone marrow and have the potential of multi-differentiation and stem cell-specific self-renewal. Bone marrow-derived EPCs promote the regeneration of blood vessels and nerve axons, migrate to organs or tissues with pathological damage, and promote tissue recovery.52,53 EPCs-Exo can be absorbed by BMDM in vitro and polarized towards an anti-inflammatory phenotype. In vivo experiments, intravenous injection of EPCs-Exo significantly reduced the proportion of M1 macrophages in the injured area and promoted an increase in M2 macrophages. Mechanistically, exosomes-derived miR-222-3p inhibits SOCS3 and activates the downstream JAK2/STAT3 pathway, thereby improving motor function recovery in mice.54
Human Placental Mesenchymal Stem Cells Derived Exosomes
As reported by Zhou et al, exosomes derived from human placental mesenchymal stem cells (hPMSCs-Exo) can activate endogenous neural progenitor cells (NPCs), induce neural differentiation, and improve hindlimb motor function. Additionally, exosomes therapy promotes the recovery of urinary function by restoring neurogenic bladder function and reducing urinary retention in mice with SCI. This process may involve activation of MEK/ERK/CREB signaling pathways.55 Conversely, the combined application of exosomes and hyperbaric oxygen promotes recovery after SCI in rats, and administration of hPMSCs-Exo promotes tube formation and migration of human umbilical vein endothelial cells (HUVECs).56,57
Neurons and Neural Stem Cells Derived Exosomes
As neuronal synaptic function differentiates, the capacity for neural regeneration declines with age. Neuron-derived exosomes (Neurons-Exo) can enhance the activity of adjacent neurons, promote neuronal differentiation and the maturation of oligodendrocyte precursor cells (OPCs), and inhibit the toxic activation of A1 astrocytes and M1 microglia.58 An miRNA array analysis revealed that miR-124-3p is the most enriched in neuron-derived exosomes, and MYH9 was identified as a downstream target gene of miR-124-3p. The miR-124-3p-MYH9 axis reduces neuroinflammation by modulating the PI3K/AKT/NF-κB signaling pathway, thereby playing a role in SCI repair.59 Neural stem cells derived exosomes (NSCs-Exo) derived VEGF-A promotes the angiogenic activity of SCMECs and the recovery of neurological function in mice.60 In addition, NSCs-Exo-miR-374-5p activates neuronal autophagy response by targeting STK-4, thereby exerting a protective function against SCI.61 Three-dimensional (3D)-printed hydrogel patches loaded with GFR+ NSCs-Exo can activate the autophagy pathway, promote neural functional recovery, and facilitate neurite regeneration.62
Microvascular Endothelial Cells Derived Exosomes
Microvascular endothelial cells are important cells that maintain the BSCB and angiogenesis.63,64 The inflammatory microenvironment may impair the survival of microvascular endothelial cells. Exosomes derived from endothelial cells (ECs-Exo) have demonstrated significant potential in promoting nerve regeneration during the repair of peripheral nerve injuries.65 In SCI, ECs-Exo can promote the M2 polarization of microglia/macrophages and improve LPS-induced mitochondrial damage. In vivo injection experiments have demonstrated effective promotion of BMS scores and muscle strength recovery in mice. Through transcriptome sequencing and mass spectrometry analysis, exosomes-derived USP13, as a deubiquitinase, regulates microglial polarization and mitochondrial function by stabilizing IκBα activity.66 Hypoxic stimulation can increase the levels of angiogenic growth factors in cells, such as hypoxia-inducible factor-1α (HIF-1α), fibroblast growth factor, and transforming growth factor-β1. Exosomes derived from hypoxically treated HUVECs have been shown to exhibit significant pro-angiogenic effects following SCI.67 Studies have shown that macrophages are the primary cells that take up exosomes derived from ECs Once inside macrophages, ECs-exo can promote the polarization of macrophages into the M2 subtype.68 Conversely, pericytes are mesenchymal cells located outside vascular endothelial cells, surrounding the basement membrane of capillaries and small venules. Pericytes are not only closely related to vascular growth, stability, and repair but also play a role in neuroprotection, neural repair, and the regulation of inflammatory responses. In the CNS, pericytes, together with neurons, astrocytes, and vascular endothelial cells, form the neurovascular unit, which is crucial to maintaining the integrity of the BBB. Research indicates that ECs can uptake exosomes derived from pericytes, thereby enhancing endothelial cell integrity and promoting the restoration of the BSCB.69
Induced Pluripotent Stem Cells Derived Exosomes
Induced pluripotent stem cells (iPSCs) can control the self-renewal, proliferation, differentiation, apoptosis and metabolism of stem cells.70 Injection of iPSCs-Exo in vivo can improve the M2 phenotypic transformation of M1 macrophages and promote motor recovery and neuronal regeneration in rats. Mechanically, miR-199b-5p was significantly expressed in iPSCs-Exo, and high expression of miR-199b-5p targeted HGf, inhibiting the activation of the PI3K signaling pathway, which is a key pathway for iPSCs-Exo to promote SCI repair.71 In addition, exosomes derived from induced pluripotent stem cell-derived neural stem cells (iPSC-NSCs) can inhibit microglia/macrophage pyroptosis and alleviate inflammatory responses.72
Treg Cell Derived Exosomes
Regulatory T cells (Tregs) are a subset of T cells with significant immunosuppressive functions, characterized by the expression of Foxp3, CD25, and CD4. Treg cells are crucial to maintain peripheral tolerance, prevent autoimmunity, and limit chronic inflammatory diseases.73,74 Research has found that Treg cell infiltration and microglial cell apoptosis increase simultaneously after SCI. Diphtheria toxininjection of Foxp3 selectively consumes Treg cells, resulting in extensive microglial death in mice, as well as significant alterations to foot imprinting and BMS scores. The injection of Treg cells into the tail vein of mice alleviates the scorched death process of microglia and the loss of motor function. Bioinformatics analysis showed that miR-709 was significantly enriched in the exosomes secreted by Treg cells. Through online database prediction and experimental verification, it was clarified that the miR-709-NKAP axis was an important mechanism for the function of Treg-Exo.74 In terms of angiogenesis, Treg-Exo can restore the integrity of the BSCB after SCI and reduce its permeability.75
Macrophage/Microglial Derived Exosomes
M1 macrophages exacerbate the progression of SCI by releasing various pro-inflammatory factors.76 Studies have shown that M1-derived exosomes treatment significantly increases ROS levels and mitochondrial fragmentation in bEnd.3 cells, and the M1-Exo-miR-155/SOCS6/p65 axis impairs mitochondrial function in bEnd.3 cells. When M1-Exo is injected in vivo, the recovery of motor function post-SCI is significantly hindered, as evidenced by BMS scores, footprint analysis, electrophysiological tests, and swimming tests. EB extravasation tests reveal that M1-Exo induces endothelial-mesenchymal transition (EndoMT) and aggravates the disruption of the BSCB.77 In contrast, M2 macrophages release anti-inflammatory factors that promote the repair of damaged spinal cord tissue. Treatment with M2 macrophage-derived exosomes enhances angiogenesis and neurogenesis following SCI, reduces tissue damage, and improves functional recovery. This process is associated with the activation of the HIF-1/VEGF signaling pathway.78 Similarly, peripheral macrophage-derived exosomes (PM-Exo) enhance autophagy and promote the polarization of anti-inflammatory microglia by inhibiting the PI3K/AKT/mTOR pathway, which is crucial for neuroprotection following SCI.79
Microglia derived exosomes: The anti-inflammatory microglia (M2 microglia) have been shown to reduce inflammation and promote spinal cord recovery. However, the role of exosomes derived from M2 microglia in SCI therapy is unclear. Through a series of experiments, Zhou et al found that miR-672-5p is the key miRNA for the function of M2-Exo. By targeting AIM2 to inhibit ASC/ caspase-1 signal pathway, thus to reduce neuronal scorch death, promote axonal regeneration and the recovery of functional behavior in mice.80 M2-Exo may exert this neuroprotective effect by inhibiting the NF-κB pathway, thereby reducing A1 astrocyte activation and enhancing neuronal survival and axonal growth.81 LPS-induced activated microglial exosomes were collected and co-cultured with astrocytes. The results showed that the combination of exosomes-miR-145-5p and Smad3 could regulate the proliferation of astrocytes, which was closely related to scar formation after SCI.82
BMSCs Derived Exosomes
BMSCs are a type of cell with the potential to differentiate into bone, cartilage, fat, nerve, and muscle cells. Their therapeutic effects in neurological and inflammatory diseases have been widely reported.83 Obtaining BMSCs-Exo for treatment can upregulate macrophage receptors with collagen structure (MARCO) in macrophages, enhance macrophage phagocytic ability, and promote neurological recovery after SCI.84 It is worth mentioning that BMSCs-Exo is used to educate macrophages to promote angiogenesis and axonal growth after SCI.85 In addition to the effects on macrophages, BMSCs-Exo can inhibit the activation of A1 astrocytes and reduce glial scar formation.86 The overexpressed neurotrophic factor (NGF) in BMSCs-Exo can stimulate the differentiation of NSCs and promote axonal regeneration of neurons.87,88 Additionally, BMSC-Exo inhibits BSCB damage and pericyte pyrogenesis while maintaining BSCB integrity by regulating the TIMP2/MMP signal pathway and the NOD1 signal pathway.89,90
Olfactory Ensheathing Cells Derived Exosomes
Transplantation of olfactory ensheathing cells (OECs) has been shown to play a beneficial role in SCI by the regulation of neuronal inflammation, neuronal survival and promoting angiogenesis.91,92 To further verify whether OECs-Exo are an important carrier for the function of OECs, extracted OECs-Exo were delivered to microglia and found that it can inhibit NF-κB and c-Jun in vitro and reverse the M1 polarization of microglia. In vivo injection of OECs-Exo reduced neuronal death and promoted axonal survival after SCI.93 This suggests that exosomes may serve as signaling molecules for OECs in the treatment of SCI.
Adipose Derived Multipotent Adult Stem Cells Derived Exosomes
Mesenchymal stem cells extracted from human epidural adipose (hEpiAD-MSC) tissue, typically obtained during posterior lumbar decompression surgery, have shown therapeutic potential. ADMSC-Exo can inhibit the expression of inflammatory factors in spinal cord tissue and M1 microglia while promoting the expression of M2 microglia.94 After extraction and identification, hEpiAD-MSC-derived exosomes (hEpiAD-MSC-Exo) were injected into SCI rats through tail vein. The results showed that compared with the injury group, hEpiAD-MSC-Exo treatment significantly decreased the expression of inflammatory cytokines such as IL-1β, IL-2 and TNF- α in the injured area, and promoted the improvement of neurotrophic related factors and BBB score in rats.95 Conversely, the exosomes were obtained after anoxic treatment of AD-MSC. It was found that Hypo-exo significantly reduced neuronal apoptosis and cavity formation in the injured site of OGD model. Further microRNA sequencing analysis of exosomes shows that miR-499a-5p targeting JNK3/c-jun apoptosis signal pathway is the key mechanism.96
Dental Pulp Stem Cells Derived Exosomes
Dental pulp stem cells (DPSCs) are usually extracted from discarded teeth.97 Like the spinal cord, DPSCs also originates from the neural crest. DPSCs can differentiate into a variety of cells, such as neurons and glial cells, in vitro and in vivo.98 The transplantation of hDPSCs can improve motor nerve function in rat SCI model. DPSCs can also produce neurotrophic factors to support nerve cell survival.98 DPSCs-Exo has stronger immunomodulatory effects than BMSCs-Exo. Therefore, the exosomes derived from DPSCs may be an excellent candidate for the treatment of SCI.97,99 Furthermore, increased levels of ROS can promote the pro-inflammatory polarization of M1 macrophages. However, after treatment with exosomes derived from DPSCs both in vitro and in vivo, the exosomes are able to inhibit the polarization of M1 macrophages and alleviate inflammation and neuronal damage through the ROS-MAPK-NF-κB p65 signaling pathway.100 This study shows the potential hope of DPSC-Exo in SCI repair.
Here, we reviewed the biological functions of exosomes from different cell sources in SCI. Studies indicated that exosomes have diverse functions (Figure 3). Currently, most research still focuses on the roles of exosomes-derived non-coding RNAs in the mechanisms of inflammation, neuroregeneration, angiogenesis, and functional recovery in SCI. This highlights the potential clinical value of non-coding RNAs as key molecules that should not be overlooked in SCI repair. Therefore, we have provided a detailed explanation of the regulatory mechanisms of exosomes-derived non-coding RNAs in SCI.
 |
Figure 3 The biological functions of exosomes derived from different cell sources. |
Exosomes Derived Non-Coding RNA
Although over 80% of the genome can be transcribed, only approximately 2% of the genome continues to be translated into proteins after transcription. Most remaining genomic sequences are transcribed into non-coding RNAs.101 Non-coding RNAs do not directly participate in protein synthesis, but they influence many physiological and pathological processes in organisms by regulating gene expression, cellular structure, signal transduction, and other aspects. Therapeutic strategies targeting miRNA, circRNA, or lncRNA are becoming part of precision medicine.
miRNA derived from exosomes: miRNA, also known as microRNA, is a short RNA molecule that is not involved in coding. It has a size of approximately 21–25 nucleotides and has the ability to attach to either the 3 “UTR or 5” UTR area of the target gene. This attachment leads to the downregulation of gene expression after transcription.102 Many different targeted mRNAs can be combined with the same miRNA. Conversely, the same mRNA can also be targeted by different miRNA.103 miRNA can regulate gene expression and participate in the occurrence and development of many pathological conditions such as cancer, cardiovascular disease, endocrine system disease and nervous system disease.104–107 In SCI disease, miRNA from multiple exosomes sources plays an important role in the progression of SCI by combining with target genes. For example, BMCS- Exo-miR-9-5p promotes FGF2 expression by inhibiting HDAC5-mediated deacetylation, thereby inhibiting LPS-induced apoptosis, inflammation and ER stress in PC12 cells.108 The exosomes were obtained from hypoxia-treated MSCs (HExos). It was found that HExos-miR-216a-5p could promote the transition from M1 phenotype to M2 phenotype of macrophages in vivo and in vitro by targeting TLR4.109 Microglia-Exo-miRNA-151-3p regulates neuronal apoptosis and axonal regeneration by regulating p53/p21/CDK1 signal pathway.110 In addition, Exo-miRNA plays an important role in promoting anti-inflammatory macrophage polarization, neural stem cell proliferation and angiogenesis, maintaining SC repair-related phenotype and slowing cell pyrogenesis (all related Exo-miRNA are listed in Table 1). This indicates that Exo-miRNA can serve as an important molecule in SCI repair.
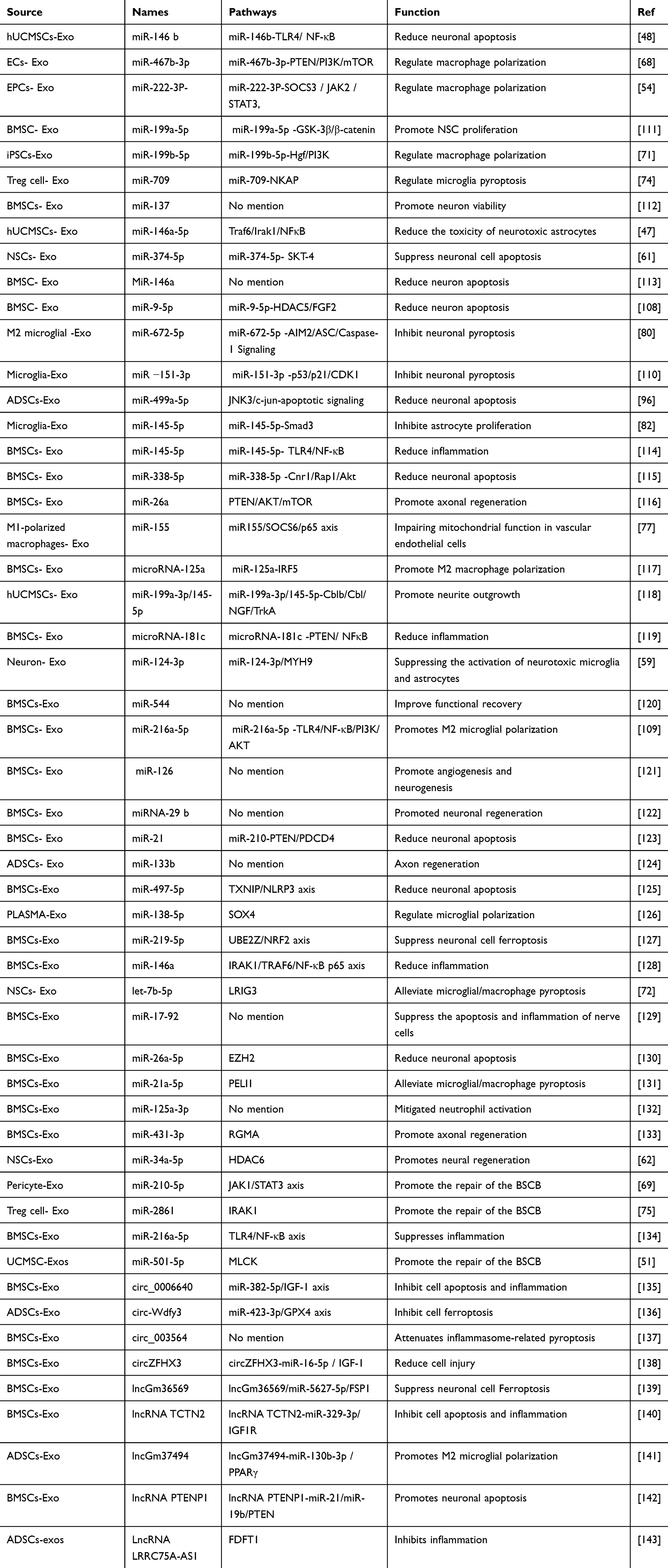 |
Table 1 The Mechanism of Non-Coding RNA Derived From Exosomes in Regulating SCI |
lncRNA derived from exosomes: long non coding RNA (lncRNA) with a length not exceeding 200 nucleotides and does not encode any peptides or proteins. lncRNA can regulate genes at three levels: epigenetic, transcriptional and post-transcriptional.144 These lncRNA are involved in the progression of many diseases.145,146 The functional mechanisms of lncRNA include decoying lncRNA, guiding lncRNA, scaffold lncRNA, stabilizing lncRNA, and competitive endogenous lncRNA (ceRNA). Among them, ceRNA is the classical mechanism by which lncRNAs function.147 The ceRNA hypothesis means that RNA transcripts with the same miRNA reaction element can compete with each other and bind to the same miRNA, and hinder the binding of miRNA to the target gene through sponge adsorption.148 In SCI studies, LncGm37494 was significantly expressed in the exosomes of hypoxic treated adipose mesenchymal stem cells. Upregulation of Exo-lncGm37494 can inhibit miR-130b-3p and promote PPARγ expression and phenotype transformation of microglia from M1 to M2.141 Transfection of knockdown lncRNA PTENP1 exosomes in PC12 cells can promote the upregulation of miR-21/miR-19b and down-regulate the expression of PTEN to inhibit neuronal apoptosis.142 Moreover, Exo-lncRNA TCTN2/miR-329-3p/IGF1R inhibits LPS-induced apoptosis, inflammation and oxidative stress, while the lncGm36569-miR-5627-5p/FSP1 pathway inhibits iron degradation.139,140
circRNA derived from exosomes: Aside from lncRNA, circRNA, as an important member of the non-coding family, functions in SCI repair by competing with miRNA. As reported by Tian et al, Exo-circZFHX3 regulated the expression of IGF-1 by adsorbing miR-16-5p, which increases the viability of BV-2 cells treated with LPS and inhibits apoptosis, inflammation and oxidative stress in vitro.138 In an in vivo experiment, Exo-circZFHX3 treatment was shown to reduce the area of SCI damage and promote the functional recovery of mice.149 In another report, Exo-circle003564 can reduce the process of neuronal pyroptosis, but the specific mechanism remains to be elucidated.137 Here, we elucidate the impact of non-coding RNAs derived from exosoms on SCI, and summarize the sources and functions of all non-coding RNAs in Table 1.
A number of studies focus on exosomes-derived non-coding RNAs, suggesting that the ncRNAs carried by exosomes play important biological roles in intercellular communication, regulation of gene expression, cell differentiation and apoptosis, as well as epigenetic repair (Figure 4). This provides an explanation for the possible mechanisms of exosomes functions and lays a favorable foundation for the future development of effective molecular drugs targeting SCI repair.
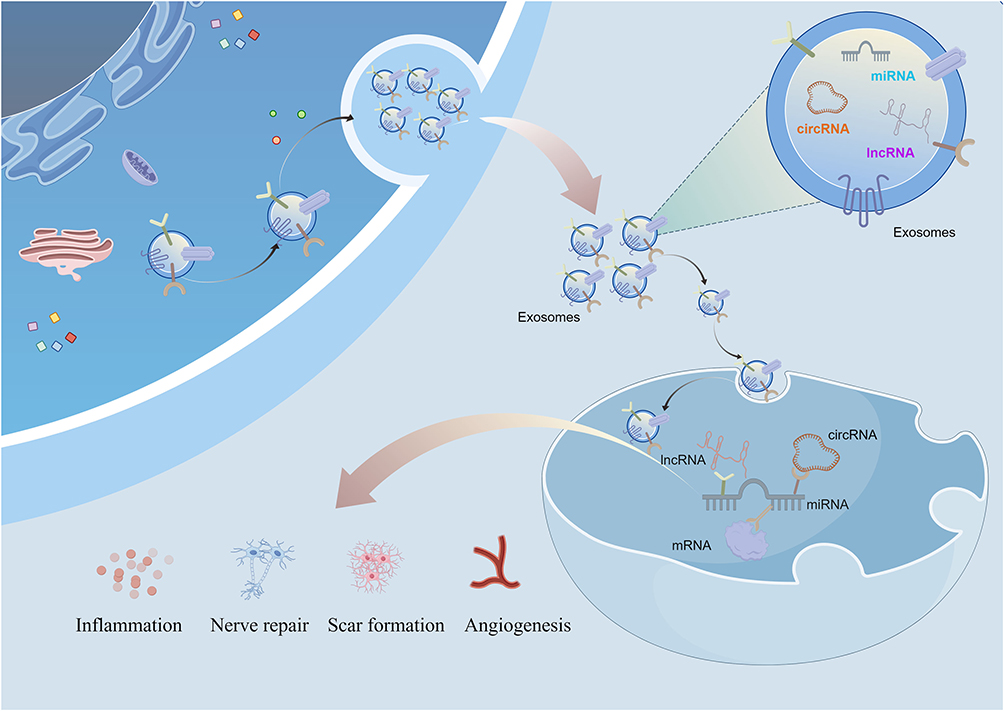 |
Figure 4 Exosomes-derived lncRNA, circRNA, and miRNA can regulate the inflammatory response, nerve damage, scar formation, and angiogenesis after SCI through the classical ceRNA mechanism. |
Engineering Modification of Exosomes
Engineering modifications of exosomes can enhance their stability and persistence in circulation (making them less prone to degradation or clearance), thereby prolonging their action time. Through engineering modifications, specific ligands (such as antibodies, peptides, or aptamers) can be introduced on the surface of exosomes, enabling them to precisely target specific cells or tissues, thereby improving therapeutic efficacy and reducing off-target effects. In addition, exosomes can be designed to carry multiple functional molecules (such as proteins, RNAs, and drugs), providing integrated therapeutic functions (Figure 5).
 |
Figure 5 Hydrogel-loaded exosomes for sustained release or exosomes carrying various functional molecules (such as proteins, RNA, and drugs) exert repair functions in SCI. |
Hydrogel Loaded Exosomes
As a biomaterial with good biocompatibility and controllability, hydrogel is considered to be the best choice for loading exosomes.150 The composition of hydrogel is similar to that of extracellular matrix (ECM), which provides a favorable culture environment for cells and is usually used as a scaffold for the treatment of SCI cystic cavity.151 The degradability and biocompatibility of different hydrogels are different. It is very important to select appropriate hydrogels or mix a variety of hydrogels according to different proportions for the sustained release of exosomes and to maintain activity. Spinal cord implantation of exosomes requires viscoelastic 3D materials and adhesive surface characteristics. A PPFLMLLLKGSTR peptide modified HA hydrogel (pGel) has been proposed to comprehensively regulate the SCI microenvironment by implanting exosomes and supplementing ECM in the interstitial space of transverse lesions. pGel improves 3D adhesion and retention of hMSC-derived exosomes. Exosomes are continuously released from the 3D pGel for approximately11 days, and more than 90% of the EVs are finally released smoothly. pGel simulates ECM function in vitro, and has persistent retention and delivery functions for hMSC-Exo. Furthermore, Exo-pGel implantation effectively restores motor function and reduced inflammatory markers in SCI rats. Examination of the urinary system has revealed that Exo-pGel has a significant function in the recovery of the urinary system.152 In 3D gelatin methacrylate hydrogel (GelMA), exosomes can be uniformly distributed on the hydrogel skeleton, while GelMA hydrogel shows a slow degradation rate and can release exosomes continuously for more than 14 days. GelMA loaded with exosomes was injected into damaged lesions, promoting the differentiation and extension of neurons in vitro, and alleviating the glial scar in the damaged lesions.153
The biocompatibility of hydrogels is crucial for loading exosomes. Fiber gel has good safety and tissue adaptability in clinical application. Therefore, the UCMSCs-Exo and the BMSCs-Exo coated with fibrin gel are injected into the spinal cord transected area, which matches the mechanical strength of the spinal cord tissue and can effectively reduce oxidation and inflammatory microenvironment, promote the protection of nerve and urinary tissue, differentiation of OPCs and oligodendrocyte production in vivo.154,155
3D-printed scaffolds are widely used in modern manufacturing, healthcare, engineering, and other fields, offering advantages such as personalized customization and complex manufacturing capabilities. The application of 3D-printed hydrogels loaded with plant-derived exosomes in SCI has shown positive effects in alleviating inflammatory responses and promoting nerve regeneration.156 Compared with simple hydrogel loading, hydrogel combined with microneedle array patch can effectively reduce inflammation and glial scar caused by SCI by loading 3D cultured MSCs-Exo. In composite hydrogel, ubiquitin thioesterase otulin (OTULIN) derived from M2-Exos was embedded in light-cured mixed hydrogel (Gel+HA-NB) to form a local drug delivery system. The physicochemical properties show that the sol gel after 37 °C UV exposure provides a basis for local administration after SCI, and this porous structure hydrogel has good biocompatibility and biodegradability. M2-Exo-OTULIN injection in vivo had a positive effect on angiogenic activity.157
Significantly, conductive hydrogel scaffolds have suitable swelling and mechanical properties, as well as excellent electrical conductivity similar to natural nerve tissue. Photocrosslinked gelatin methacrylate and polypyrrole (PPy) hydrogel constitute a double network conductive (GMP) hydrogel loaded with exosomes. The network has natural cell binding motifs such as Arg-Gly-Asp, which allow cells to grow in it. Covalent binding does not affect the structure and biological activity of the exosomes, and ensures that the exosomes is released slowly and continuously in the early stage of implantation. Exosomes were fixed into a GMP hydrogel network, which increased NSC compatibility of conductive hydrogel and promoted oligodendrocyte differentiation. The mechanism study shows that conductive hydrogels may regulate NSC differentiation and axonal regeneration through PTEN/PI3K/AKT/mTOR pathway.158 The implantation of conductive hydrogels can trigger immune responses, and the persistent presence of conductive components may exacerbate early inflammation following acute SCI. Loading M2-Exos into conductive hydrogels can reduce post-injury inflammatory responses, promote neuronal survival, and mitigate the formation of glial scars.159 In summary, conductive hydrogels that mimic the electrical transport properties of natural nerve tissue are highly beneficial for the repair of SCI.
Gene Modified Exosomes Therapy
Gene therapy has shown good therapeutic effects in various diseases. As a signal carrier for intercellular communication, exosomes can freely pass through the BSCB without causing immune rejection, and can carry various molecules for substance delivery, thereby achieving widespread intercellular signaling communication.160 Different genes can be loaded into exosomes to achieve therapeutic purposes. Engineered iPSCs-Exo loaded with miR-23b, miR-21-5p, and miR-199b-5p can alleviate inflammation in neurons stimulated by LPS and IFN-γ, significantly promoting the recovery of SCI.161 It is reported that miR-21 can regulate astrocyte polarization, glial scar formation and axonal growth after SCI.162 To effectively achieve the specificity and precision of miR-21 in the treatment of SCI, miR-21 was transiently overexpressed in CBD-Lamp2b-293T cells to obtain CBD-LP-Exo enriched with miR-21. The functional biomaterial scaffold of CBD-Lamp2b-miR-21- exosomes collagen-I was constructed by genetic engineering, and the scaffold was implanted into the body for SCI repair. Scaffolds promoted the continuous release of miR-21 into cells, inhibited apoptosis, improved neuronal survival and reduced the production of glial scars at SCI sites, showing excellent performance in SCI repair.163
Netrin-1 is one of the earliest recognized axon guiding factors, which can initiate axon growth and branch formation, promoting the migration and survival of nerve cells.164 Chemically modified information RNA (modRNA) refers to mRNA synthesized in vitro using chemically modified bases, which is further transferred to target cells through liposomes or nanoparticles for protein translation and expression.165 Using exosomes as carriers, loading modRNAmodNetrin−1 encoding netrin-1 effectively alleviated LPS induced PC12 cell inflammation and necrosis, and tail vein injection of Exo-modRNAmodNetrin−1 promoted nerve recovery.166 In addition, miR-138-5p-modified UCMSC-Exo loaded in thermosensitive hydrogel can reduce neuronal apoptosis in vivo by inhibiting intracellular ROS levels through the Nrf2-keap1 signaling pathway.167
Protein Modification of Exosomes
Although exosomes have shown a good therapeutic effect after SCI, the exosomes delivered to the injured area still lacks targeting. Usually, after being delivered to the damaged area, exosomes can enter various cells for signal transmission, resulting in a lack of specificity and effectiveness in treatment. Therefore, adding specific recognition peptides to the surface of exosomes can increase the targeting of exosomes delivery and accelerate the progress of SCI repair. However, lack of site targeting may lead to different therapeutic responses. Exosomes from autologous plasma (AP-Exo) have been used as functional cargo carriers and growth promoting peptides (ILP and ISP) and targeted peptides were loaded through CP05. Biological scaffolds based on AP-Exo can target and deliver AP-Exo through the BSCB, homing them to the SCI injury site, entering neurons in the damaged spinal cord, and promoting axonal regeneration.168
Collagen, as the main component of the extracellular matrix, is superior to other materials in promoting cell adhesion and growth.169,170 Among them, linearly ordered collagen scaffold can guide the growth of nerve processes along its fibers and reduce the deposition of scar-related components in the injured site.171 The author of that study designed a new type of bio-specificity peptide that can effectively connect the collagen scaffold with mesenchymal stem cells derived exosomes (MExo). In vivo implantation of MExos can promote the migration of more NSCs to the center of the damaged area, which improves SCI recovery.172
M2 macrophage-Exo have shown good anti-inflammatory effects in vitro and in vivo. Le-Lys-Val-Ala-Val (IKVAV) is the core functional peptide of laminin, which contributes to the survival, proliferation and differentiation of NSCs.173 Viral macrophage inflammatory protein II (vMIP-II) is a broad-spectrum chemokine receptor-binding peptide. M2 macrophage-derived Exos modified with the vMIP-II-Lamp2b gene can target chemokine receptors highly expressed in injured spinal cord, inhibit chemokine signaling, reduce the production of pro-inflammatory factors, and regulate microglia/macrophage polarization to promote SCI repair.174 Therefore, it is of great significance to prepare nano-reagents with both anti-inflammatory and nerve-targeted repair as ideal therapeutic drugs for SCI. IKVAV peptides were coupled to the surface of the M2 macrophage-Exo by copper-free cyclo-addition reaction and tail vein injection of exosomes showed that this new nano drug has excellent effects in promoting M2 macrophage polarization and NSCs differentiation.175
Exosomes Loaded with Small Molecule Drugs
Drug therapy is an important means to deal with SCI. Drug treatment is traditionally via oral or intravenous infusion; however, when the drug passes through the blood circulation, because of the BSCB, it is difficult for the drug to be delivered to the damaged tissue, which greatly reduces the effectiveness of the drug. Therefore, it is critical to find a suitable drug carrier and deliver the drug to the injured area in an efficient and orderly manner. Because of their acellular structure, exosomes have the advantage of natural drug loading, so it has been regarded as the best drug carrier, and can be loaded with targeted drugs for specific diseases. PTEN is a well-known tumor suppressor, which has been identified as a key regulator of neuronal apoptosis and inflammation. Effectively knocking down the expression of PTEN can promote SCI repair.176 In the study of Guo et al, it was found that intranasal delivery MSCs-Exo migrated through chemotaxis and had high affinity for neurons. PTEN-siRNA reagent was loaded into MSC-Exo (Exo-siPTEN) in a certain proportion to treat dorsal root ganglion (DRG) neurons. Compared with neurons treated with control siRNA, DRG neurons exhibited more complex, branched, and elongated morphology after 1 day of Exo-siPTEN treatment. This indicates that Exo-siPTEN can promote the growth of DRG neurons in vitro. In vivo experiments, nasal ExoPTEN treatment can induce motor, sensory, and bladder function recovery in SCI mice.177
Berberine originates from the traditional Chinese medicine Huanglian and has various pharmacological effects, including anti-inflammatory, antioxidant, antibacterial, and neuroprotective effects.178,179 However, the short half-life and low efficiency of crossing the BBB seriously affect the pharmacological effects of berberine. To overcome these shortcomings, berberine was loaded Ber into M2-Exo and -the -barrier penetration effect of exosomes was used to deliver drugs to SCI damaged areas for treatment. Experiments show that Exos- Berberine can reduce the expression of inflammatory factors such as TNF-α, IL-1β and IL-6 and neuronal apoptosis, and promote the recovery of motor function.180
FTY720 is a functional antagonist of sphingosine 1-phosphate receptor-1 (S1P1) and has immunomodulatory function.181 However, although the application of FTY720 has a positive effect on SCI, systemic administration easily causes serious adverse reactions.182 Therefore, NSCs-Exos were used as a carrier for FTY720 for SCI repair. The results showed that compared with the simple FTY720 reagent group, tail vein injection of NSCs-Exo-loaded FTY720 had a better effect on improving endothelial cell function, relieving apoptosis and spinal cord edema, and promoting the recovery of motor function in mice.183 AM1241 is a selective CB2 receptor agonist that plays an important role in anti-inflammatory, neuroprotective, and analgesic functions. It has broad application prospects in the treatment of neurological diseases and chronic pain.184 Exosomes-loaded AM1241 can activate the Nrf2/HO-1 signaling pathway, thereby reducing neuronal inflammation, oxidative stress, and apoptosis. It also contributes to promoting neural stem cell differentiation and motor function recovery.185 Although the delivery system formed by loading small molecule drugs, proteins, and genes through EVs has shown certain therapeutic effects in SCI, exosomes still face issues as carriers such as poor targeting, short in vivo maintenance time, and off target. Further engineering transformation of exosomes is a necessary exploration for the development of exosomes therapy.
Exploration of Future Applications of Exosomes in SCI
Non-Invasive Prognostic Markers
Patients with SCI often experience poor overall health and require long-term, repeated monitoring to assess their recovery status. Exosomes, which can be obtained from various body fluids such as blood, urine, and saliva, offer a rich source of biomarkers.186 Compared to traditional biopsy or tissue sampling, the collection of exosomes is simpler, safer, and lower risk. Therefore, for high-risk or older adult patients, non-invasive exosomes-based detection provides a convenient and effective means of prognostic evaluation. Exosomes contain a variety of molecular information, including proteins, mRNA, miRNA, and DNA, which are determined by the state and function of their parent cells. As a result, these molecules exhibit high sensitivity and specificity, accurately and comprehensively reflecting cellular states and indicators such as inflammation, neural recovery, muscle strength, and sensory recovery.187 By monitoring the dynamic changes in these biomarkers, treatment plans can be adjusted in a timely manner. Thus, exosomes can provide comprehensive and precise disease information, and the detection of these markers holds great potential for long-term monitoring and treatment efficacy evaluation in SCI patients.
The Diversified Administration Methods of Exosomes
Exosomes are naturally occurring nanoparticles with good biocompatibility and low immunogenicity. As an ideal drug delivery system, exosomes offer various administration methods, including intravenous injection, local administration, and oral administration.
Intravenous injection: Due to their ability to cross the BBB and be taken up by various cells, intravenous injection is a common method for exosomes-based drug delivery. Through intravenous injection, exosomes can rapidly enter the bloodstream and be transported to the injured area to exert their therapeutic effects. Additionally, exosomes can remain stable in the bloodstream and circulate for an extended period, making intravenous injection a promising approach for clinical treatment. However, most exosomes are metabolized by the liver and kidneys after intravenous injection, and only a small fraction of exosomes reach the injured area to exert their therapeutic effect. This significantly reduces the effective therapeutic dose and may lead to systemic side effects. Therefore, intravenous injection requires close monitoring of the patient’s overall condition.
Local administration: Through local administration, exosomes and the small molecules they carry can reach higher concentrations at the site of injury. By targeting macrophages, microglia, astrocytes, and neurons, the precision and efficacy of local treatment can be increased, maximizing the therapeutic effect at the local site. Additionally, local administration significantly reduces the distribution of the drug in the systemic circulation, minimizing metabolism and clearance by organs such as the liver and kidneys, thus avoiding systemic side effects. This also reduces the damage to healthy tissues and improves patient tolerance. When exosomes are combined with local delivery systems (such as hydrogels and scaffolds), sustained release and long-lasting effects of the drug can be achieved. This reduces the frequency of administration and enhances patient compliance. Therefore, local exosomes delivery can lower the difficulty of treatment and improve patient acceptance. With the continuous development and maturation of exosomes technology and local delivery systems, their application prospects in clinical treatment after SCI will become even more extensive.
Oral administration: Oral administration is a non-invasive method that avoids the discomfort and potential complications associated with other delivery methods, such as injections. Patients can self-administer medications at home, enhancing the convenience of treatment, which is particularly important for the long-term management of chronic SCI. Additionally, the production, transportation, and use of oral medications are generally cost-effective, potentially reducing the overall medical expenses for patients. However, oral administration also faces challenges similar to intravenous delivery, such as systemic reactions. Moreover, the digestive system may affect the activity and stability of exosomes, posing a potential limitation.
Currently, the diversified administration methods of exosomes hold promise to meet the clinical application demands. In the long term, the suitability of exosomes for both short-term and long-term use in SCI patients remains uncertain. Whether exosomes should be adopted as a long-term strategy for SCI patients requires further exploration through extensive clinical prospective studies.
Exosomes Therapy in Personalized Medicine Model
The personalized medicine model for exosomes therapy is a new medical approach that is patient-centered and customizes treatment plans based on individual differences. In personalized medicine, treatment not only considers the patient’s condition but also factors such as immune status, environmental influences, and lifestyle, to create tailored treatment strategies. By leveraging the unique properties of exosomes and integrating biotechnology and data analysis, highly personalized treatment plans are one of the optimal choices in the treatment of SCI. Treatment can involve extracting exosomes from the patient’s cells (such as mesenchymal stem cells, immune cells, or tumor cells) to ensure biocompatibility and low immunogenicity, achieving personalized sourcing of exosomes. Moreover, based on the patient’s genetic characteristics, the extent of the injury, and systemic response status, artificial intelligence technology can be used to optimize the design of exosomes and treatment plans. This involves surface targeting modifications and loading specific therapeutic molecules (such as miRNA, siRNA, proteins, or drugs) to design personalized local delivery systems, enabling highly precise treatment for the patient. In long-term health management, by monitoring the dynamic changes in exosomes in the patient’s body fluids, it is possible to track treatment outcomes or adverse reactions, assess the effectiveness of treatment, and enable dynamic personalized therapy. This provides a new strategy for the development of precision medicine following SCI.
However, the rehabilitation process for SCI patients is slow. By combining exosomes therapy with exercise training, neural plasticity can be effectively enhanced. Tailoring the preparation and delivery of exosomes based on the patient’s condition, along with dynamic safety assessments, holds promise to achieve long-term health management in these patients. However, the long-term effects and safety of exosomes therapy still need attention. First, the exosomes delivery system may accumulate in the body, and the long-term effects could be influenced by factors such as drug release control, exosomes stability, and cellular uptake. Second, immune response issues related to exosomes still exist, especially with prolonged use, which may trigger immune rejection. Additionally, exosomes may carry harmful substances or gene-editing tools, leading to off-target effects or the risk of tumor formation, particularly when they fail to precisely target the intended cells during delivery. When crossing the BBB, exosomes may introduce potential neurotoxicity. Therefore, the long-term safety and efficacy of exosomes still need further evaluation and optimization to ensure their broad clinical application.
Challenges and Considerations in Exosomes Therapy
Despite the immense therapeutic potential of exosomes therapy, there are still numerous challenges and difficulties in its clinical translation. Exosomes commonly used in SCI treatment research are primarily derived from mesenchymal stem cells, peripheral blood, platelets, and other sources. While these sources offer certain functional advantages, the large-scale production of exosomes remains a major obstacle to their clinical application. Additionally, exosomes derived from different cell sources exhibit significant heterogeneity, and there is a lack of unified screening standards, leading to substantial batch-to-batch variability. Furthermore, there is no universally accepted optimal method for exosomes administration. Exosomes may produce different therapeutic effects in various experimental models and individuals, which is closely related to factors such as the patient’s immune status, injury severity, exosomes source, and dosage. Moreover, existing animal experimental results are difficult to fully extrapolate to clinical research, and long-term use of exosomes may raise concerns regarding potential tumorigenicity, immune dysregulation, and other issues, for which systematic studies are still lacking.
To address the challenges in exosomes applications, we believe efforts should be made to promote automated and standardized purification processes, develop new technologies to improve purity and stability, and establish rigorous industry standards and clinical guidelines. The development of engineered exosomes, coupled with imaging technologies (such as fluorescent tracing), can be used to monitor exosomes distribution within the body, enabling precise treatment. The development of personalized treatment strategies should also be pursued, selecting exosomes from specific sources based on the type of SCI and the clinical symptoms of the patient. Long-term toxicological assessments and gene expression profiling should be carried out to monitor the potential side effects of exosomes, clarifying their distribution, metabolism, and clearance pathways in the body. In conclusion, the standardization of exosomes applications holds great promise for clinical use in SCI.
Summary
The current status of SCI treatment and the various functions of exosomes derived from different cell sources in SCI have been summarized. Exosomes have significant therapeutic effects on the inflammation response, cell polarization, scar formation, angiogenesis, neuroregeneration, and axon formation following SCI. Exosomes play a crucial role in promoting SCI repair. Among them, various non-coding RNAs (such as miRNA, lncRNA, circRNA) in exosomes are key components that perform important biological functions and play a critical role in elucidating the specific mechanisms of exosomes. Additionally, researchers have made extensive efforts in engineering exosomes, including genetic modification, protein modification, hydrogel loading, and drug loading. These engineered modifications not only effectively enhance the long-term therapeutic effects of exosomes but also provide new directions and strategies for SCI treatment through novel nano-delivery systems formed by exosomes. Finally, we highlight the importance of exosomes as prognostic biomarkers and the diversification of exosomes delivery methods, and explore the value of personalized treatment in the future management of SCI (Figure 6).
 |
Figure 6 Exploring the significance of exosomes as prognostic biomarkers, diversified exosomes delivery methods, and personalized treatment in the future management of SCI. |
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (No. 82402989), Xi’an Municipal Health Commission Science and Technology Project (2025ms11).
Disclosure
The authors declare no competing interests in this work.
References
1. Ropper AE, Ropper AH. Acute spinal cord compression. N Engl J Med. 2017;376(14):1358–1369. doi:10.1056/NEJMra1516539
2. Singh A, Tetreault L, Kalsi-Ryan S, et al. Global prevalence and incidence of traumatic spinal cord injury. Clin Epidemiol. 2014;6:309–331. doi:10.2147/CLEP.S68889
3. Hu X, Xu W, Ren Y, et al. Spinal cord injury: molecular mechanisms and therapeutic interventions. Signal Transduct Target Ther. 2023;8(1):245. doi:10.1038/s41392-023-01477-6
4. Feigin VL, Nichols E, Alam T. Global, regional, and national burden of neurological disorders, 1990-2016: a systematic analysis for the global burden of disease study 2016. Lancet Neurol. 2019;18(5):459–480. doi:10.1016/S1474-4422(18)30499-X
5. Sekhon LH, Fehlings MG. Epidemiology, demographics, and pathophysiology of acute spinal cord injury. Spine. 2001;26(24 Suppl):S2–12. doi:10.1097/00007632-200112151-00002
6. Ju C, Liu R, Ma Y, et al. Targeted microbiota dysbiosis repair: an important approach to health management after spinal cord injury. Ageing Res Rev. 2025;104:102648. doi:10.1016/j.arr.2024.102648
7. Anwar MA, Shehabi TSA, Eid AH. Inflammogenesis of secondary spinal cord injury. Front Cell Neurosci. 2016;10:98. doi:10.3389/fncel.2016.00098
8. Campos J, Silva NA, Salgado AJ. Nutritional interventions for spinal cord injury: preclinical efficacy and molecular mechanisms. Nutr Rev. 2022;80(5):1206–1221. doi:10.1093/nutrit/nuab068
9. Miron VE, Franklin RJ. Macrophages and CNS remyelination. J Neurochem. 2014;130(2):165–171. doi:10.1111/jnc.12705
10. Donnelly DJ, Popovich PG. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp Neurol. 2008;209(2):378–388. doi:10.1016/j.expneurol.2007.06.009
11. Freyermuth-Trujillo X, Segura-Uribe JJ, Salgado-Ceballos H, et al. Inflammation: a target for treatment in spinal cord injury. Cells. 2022;11(17):2692 doi:10.3390/cells11172692.
12. Kigerl KA, Gensel JC, Ankeny DP, et al. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J Neurosci. 2009;29(43):13435–13444. doi:10.1523/JNEUROSCI.3257-09.2009
13. Hu X, Xu Y, Zhang H, et al. Role of necroptosis in traumatic brain and spinal cord injuries. J Adv Res. 2022;40:125–134. doi:10.1016/j.jare.2021.12.002
14. Shi Z, Yuan S, Shi L, et al. Programmed cell death in spinal cord injury pathogenesis and therapy. Cell Prolif. 2021;54(3):e12992. doi:10.1111/cpr.12992
15. Beattie MS. Inflammation and apoptosis: linked therapeutic targets in spinal cord injury. Trends Mol Med. 2004;10(12):580–583. doi:10.1016/j.molmed.2004.10.006
16. Al Mamun A, Wu Y, Monalisa I, et al. Role of pyroptosis in spinal cord injury and its therapeutic implications. J Adv Res. 2021;28:97–109. doi:10.1016/j.jare.2020.08.004
17. Hu X, Chen H, Xu H, et al. Role of pyroptosis in traumatic brain and spinal cord injuries. Int J Biol Sci. 2020;16(12):2042–2050. doi:10.7150/ijbs.45467
18. East E, Golding JP, Phillips JB. A versatile 3D culture model facilitates monitoring of astrocytes undergoing reactive gliosis. J Tissue Eng Regen Med. 2009;3(8):634–646. doi:10.1002/term.209
19. Tran AP, Warren PM, Silver J. The biology of regeneration failure and success after spinal cord injury. Physiol Rev. 2018;98(2):881–917. doi:10.1152/physrev.00017.2017
20. Anderson MA, Burda JE, Ren Y, et al. Astrocyte scar formation aids central nervous system axon regeneration. Nature. 2016;532(7598):195–200. doi:10.1038/nature17623
21. Wiese S, Karus M, Faissner A. Astrocytes as a source for extracellular matrix molecules and cytokines. Front Pharmacol. 2012;3:120. doi:10.3389/fphar.2012.00120
22. Hara M, Kobayakawa K, Ohkawa Y, et al. Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury. Nat Med. 2017;23(7):818–828. doi:10.1038/nm.4354
23. Rowland JW, Hawryluk GWJ, Kwon B, et al. Current status of acute spinal cord injury pathophysiology and emerging therapies: promise on the horizon. Neurosurg Focus. 2008;25(5):E2. doi:10.3171/FOC.2008.25.11.E2
24. Kostyusheva A, Romano E, Yan N, et al. Breaking barriers in targeted therapy: advancing exosome isolation, engineering, and imaging. Adv Drug Deliv Rev. 2025;218:115522. doi:10.1016/j.addr.2025.115522
25. Saleem M, Shahzad KA, Marryum M, et al. Exosome-based therapies for inflammatory disorders: a review of recent advances. Stem Cell Res Ther. 2024;15(1):477. doi:10.1186/s13287-024-04107-2
26. Sun Q, Chang H, Wang H, et al. Regulatory roles of extracellular vesicles in pregnancy complications. J Adv Res. 2025. doi:10.1016/j.jare.2025.02.010
27. Théry C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2(8):569–579. doi:10.1038/nri855
28. Simons M, Raposo G. Exosomes--vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21(4):575–581. doi:10.1016/j.ceb.2009.03.007
29. Deng S, Cao H, Cui X, et al. Optimization of exosome-based cell-free strategies to enhance endogenous cell functions in tissue regeneration. Acta Biomater. 2023;171:68–84. doi:10.1016/j.actbio.2023.09.023
30. Martins TS, Vaz M, Henriques AG. A review on comparative studies addressing exosome isolation methods from body fluids. Anal Bioanal Chem. 2023;415(7):1239–1263. doi:10.1007/s00216-022-04174-5
31. Gurunathan S, Kang M-H, Jeyaraj M, et al. Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cells. 2019;8(4):307. doi:10.3390/cells8040307
32. Khan NA, Asim M, Biswas KH, et al. Exosome nanovesicles as potential biomarkers and immune checkpoint signaling modulators in lung cancer microenvironment: recent advances and emerging concepts. J Exp Clin Cancer Res. 2023;42(1):221. doi:10.1186/s13046-023-02753-7
33. Mattingly J, Li Y, Bihl JC, et al. The promise of exosome applications in treating central nervous system diseases. CNS Neurosci Ther. 2021;27(12):1437–1445. doi:10.1111/cns.13743
34. Zhang S, Duan Z, Liu F, et al. The impact of exosomes derived from distinct sources on rheumatoid arthritis. Front Immunol. 2023;14:1240747. doi:10.3389/fimmu.2023.1240747
35. Anjum A, Yazid MD, Fauzi Daud M, et al. Spinal cord injury: pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int J Mol Sci. 2020;21(20):7533. doi:10.3390/ijms21207533
36. Fu H, Hu D, Chen J, et al. Repair of the injured spinal cord by schwann cell transplantation. Front Neurosci. 2022;16:800513. doi:10.3389/fnins.2022.800513
37. Xu B, Zhou Z, Fang J, et al. Exosomes derived from schwann cells alleviate mitochondrial dysfunction and necroptosis after spinal cord injury via AMPK signaling pathway-mediated mitophagy. Free Radic Biol Med. 2023;208:319–333. doi:10.1016/j.freeradbiomed.2023.08.026
38. Pan D, Zhu S, Zhang W, et al. Autophagy induced by schwann cell-derived exosomes promotes recovery after spinal cord injury in rats. Biotechnol Lett. 2022;44(1):129–142. doi:10.1007/s10529-021-03198-8
39. Ren J, Zhu B, Gu G, et al. Schwann cell-derived exosomes containing MFG-E8 modify macrophage/microglial polarization for attenuating inflammation via the SOCS3/STAT3 pathway after spinal cord injury. Cell Death Dis. 2023;14(1):70. doi:10.1038/s41419-023-05607-4
40. Zhu B, Gu G, Ren J, et al. Schwann cell-derived exosomes and methylprednisolone composite patch for spinal cord injury repair. ACS Nano. 2023;17(22):22928–22943. doi:10.1021/acsnano.3c08046
41. Pan D, Li Y, Yang F, et al. Increasing toll-like receptor 2 on astrocytes induced by schwann cell-derived exosomes promotes recovery by inhibiting CSPGs deposition after spinal cord injury. J Neuroinflammation. 2021;18(1):172. doi:10.1186/s12974-021-02215-x
42. Zhu S, Ma H, Hou M, et al. Schwann cell-derived exosomes induced axon growth after spinal cord injury by decreasing PTP-σ activation on CSPGs via the Rho/ROCK pathway. Neurochem Res. 2024;49(8):2120–2130. doi:10.1007/s11064-024-04166-0
43. Huang JH, Chen Y-N, He H, et al. Schwann cells-derived exosomes promote functional recovery after spinal cord injury by promoting angiogenesis. Front Cell Neurosci. 2022;16:1077071. doi:10.3389/fncel.2022.1077071
44. Hoang DM, Pham PT, Bach TQ, et al. Stem cell-based therapy for human diseases. Signal Transduct Target Ther. 2022;7(1):272. doi:10.1038/s41392-022-01134-4
45. Zhang Y, Zhuang H, Ren X, et al. Therapeutic effects of different intervention forms of human umbilical cord mesenchymal stem cells in the treatment of osteoarthritis. Front Cell Dev Biol. 2023;11:1246504. doi:10.3389/fcell.2023.1246504
46. Kang J, Guo Y. Human umbilical cord mesenchymal stem cells derived exosomes promote neurological function recovery in a rat spinal cord injury model. Neurochem Res. 2022;47(6):1532–1540. doi:10.1007/s11064-022-03545-9
47. Lai X, Wang Y, wang X, et al. miR-146a-5p-modified hUCMSC-derived exosomes facilitate spinal cord function recovery by targeting neurotoxic astrocytes. Stem Cell Res Ther. 2022;13(1):487. doi:10.1186/s13287-022-03116-3
48. Wang X, Yang Y, Li W, et al. Umbilical mesenchymal stem cell-derived exosomes promote spinal cord functional recovery through the miR-146b/TLR4 -mediated NF-κB p65 signaling pathway in rats. Biochem Biophys Rep. 2023;35:101497. doi:10.1016/j.bbrep.2023.101497
49. Luan Z, Liu J, Li M, et al. Exosomes derived from umbilical cord-mesenchymal stem cells inhibit the NF-κB/MAPK signaling pathway and reduce the inflammatory response to promote recovery from spinal cord injury. J Orthop Surg Res. 2024;19(1):184. doi:10.1186/s13018-024-04651-w
50. Sun G, Li G, Li D, et al. hucMSC derived exosomes promote functional recovery in spinal cord injury mice via attenuating inflammation. Mater Sci Eng C Mater Biol Appl. 2018;89:194–204. doi:10.1016/j.msec.2018.04.006
51. Xie Y, Sun Y, Liu Y, et al. Targeted delivery of RGD-CD146 + CD271 + human umbilical cord mesenchymal stem cell-derived exosomes promotes blood–spinal cord barrier repair after spinal cord injury. ACS Nano. 2023;17(18):18008–18024. doi:10.1021/acsnano.3c04423
52. Shi X, Seidle KA, Simms KJ, et al. Endothelial progenitor cells in the host defense response. Pharmacol Ther. 2023;241:108315. doi:10.1016/j.pharmthera.2022.108315
53. Zhao YH, Yuan B, Chen J, et al. Endothelial progenitor cells: therapeutic perspective for ischemic stroke. CNS Neurosci Ther. 2013;19(2):67–75. doi:10.1111/cns.12040
54. Yuan F, Peng W, Yang Y, et al. Endothelial progenitor cell-derived exosomes promote anti-inflammatory macrophages via SOCS3/JAK2/STAT3 axis and improve the outcome of spinal cord injury. J Neuroinflammation. 2023;20(1):156. doi:10.1186/s12974-023-02833-7
55. Zhou W, Silva M, Feng C, et al. Exosomes derived from human placental mesenchymal stem cells enhanced the recovery of spinal cord injury by activating endogenous neurogenesis. Stem Cell Res Ther. 2021;12(1):174. doi:10.1186/s13287-021-02248-2
56. Cheshmi H, Mohammadi H, Akbari M, et al. Human placental mesenchymal stem cell-derived exosomes in combination with hyperbaric oxygen synergistically promote recovery after spinal cord injury in rats. Neurotox Res. 2023;41(5):431–445. doi:10.1007/s12640-023-00649-0
57. Zhang C, Zhang C, Xu Y, et al. Exosomes derived from human placenta-derived mesenchymal stem cells improve neurologic function by promoting angiogenesis after spinal cord injury. Neurosci Lett. 2020;739:135399. doi:10.1016/j.neulet.2020.135399
58. Xu Y, Zhu Z-H, Xu X, et al. Neuron-derived exosomes promote the recovery of spinal cord injury by modulating Nerve cells in the cellular microenvironment of the lesion area. Mol Neurobiol. 2023;60(8):4502–4516. doi:10.1007/s12035-023-03341-8
59. Jiang D, Gong F, Ge X, et al. Neuron-derived exosomes-transmitted miR-124-3p protect traumatically injured spinal cord by suppressing the activation of neurotoxic microglia and astrocytes. J Nanobiotechnology. 2020;18(1):105. doi:10.1186/s12951-020-00665-8
60. Zhong D, Cao Y, Li C-J, et al. Neural stem cell-derived exosomes facilitate spinal cord functional recovery after injury by promoting angiogenesis. Exp Biol Med. 2020;245(1):54–65. doi:10.1177/1535370219895491
61. Zhang L, Han P. Neural stem cell-derived exosomes suppress neuronal cell apoptosis by activating autophagy via miR-374-5p/STK-4 axis in spinal cord injury. J Musculoskelet Neuronal Interact. 2022;22(3):411–421.
62. Qin T, Li C, Xu Y, et al. Local delivery of EGFR(+)NSCs-derived exosomes promotes neural regeneration post spinal cord injury via miR-34a-5p/HDAC6 pathway. Bioact Mater. 2024;33:424–443. doi:10.1016/j.bioactmat.2023.11.013
63. Chen F, Zhan J, Liu M, et al. FGF2 alleviates microvascular ischemia-reperfusion injury by KLF2-mediated ferroptosis inhibition and antioxidant responses. Int J Biol Sci. 2023;19(13):4340–4359. doi:10.7150/ijbs.85692
64. Tanaka S, Portilla D, Okusa MD. Role of perivascular cells in kidney homeostasis, inflammation, repair and fibrosis. Nat Rev Nephrol. 2023;19(11):721–732. doi:10.1038/s41581-023-00752-7
65. Huang J, Zhang G, Li S, et al. Endothelial cell-derived exosomes boost and maintain repair-related phenotypes of schwann cells via miR199-5p to promote nerve regeneration. J Nanobiotechnology. 2023;21(1):10. doi:10.1186/s12951-023-01767-9
66. Ge X, Zhou Z, Yang S, et al. Exosomal USP13 derived from microvascular endothelial cells regulates immune microenvironment and improves functional recovery after spinal cord injury by stabilizing IκBα. Cell Biosci. 2023;13(1):55. doi:10.1186/s13578-023-01011-9
67. Li L, Mu J, Zhang Y, et al. Stimulation by exosomes from hypoxia preconditioned human umbilical vein endothelial cells facilitates mesenchymal stem cells angiogenic function for spinal cord repair. ACS Nano. 2022;16(7):10811–10823. doi:10.1021/acsnano.2c02898
68. Peng W, Xie Y, Luo Z, et al. UTX deletion promotes M2 macrophage polarization by epigenetically regulating endothelial cell-macrophage crosstalk after spinal cord injury. J Nanobiotechnology. 2023;21(1):225. doi:10.1186/s12951-023-01986-0
69. Gao P, Yi J, Chen W, et al. Pericyte-derived exosomal miR-210 improves mitochondrial function and inhibits lipid peroxidation in vascular endothelial cells after traumatic spinal cord injury by activating JAK1/STAT3 signaling pathway. J Nanobiotechnology. 2023;21(1):452. doi:10.1186/s12951-023-02110-y
70. Shamsian A, Sahebnasagh R, Norouzy A, et al. Cancer cells as a new source of induced pluripotent stem cells. Stem Cell Res Ther. 2022;13(1):459. doi:10.1186/s13287-022-03145-y
71. Li J, Jing Y, Bai F, et al. Induced pluripotent stem cells as natural biofactories for exosomes carrying miR-199b-5p in the treatment of spinal cord injury. Front Pharmacol. 2022;13:1078761. doi:10.3389/fphar.2022.1078761
72. Liu J, Kong G, Lu C, et al. IPSC-NSCs-derived exosomal let-7b-5p improves motor function after spinal cord Injury by modulating microglial/macrophage pyroptosis. J Nanobiotechnology. 2024;22(1):403. doi:10.1186/s12951-024-02697-w
73. Ramanan D, Pratama A, Zhu Y, et al. Regulatory T cells in the face of the intestinal microbiota. Nat Rev Immunol. 2023;23(11):749–762. doi:10.1038/s41577-023-00890-w
74. Xiong W, Li C, Kong G, et al. Treg cell-derived exosomes miR-709 attenuates microglia pyroptosis and promotes motor function recovery after spinal cord injury. J Nanobiotechnology. 2022;20(1):529. doi:10.1186/s12951-022-01724-y
75. Kong G, Xiong W, Li C, et al. Treg cells-derived exosomes promote blood-spinal cord barrier repair and motor function recovery after spinal cord injury by delivering miR-2861. J Nanobiotechnology. 2023;21(1):364. doi:10.1186/s12951-023-02089-6
76. Kroner A, Greenhalgh A, Zarruk J, et al. TNF and increased intracellular iron alter macrophage polarization to a detrimental M1 phenotype in the injured spinal cord. Neuron. 2014;83(5):1098–1116. doi:10.1016/j.neuron.2014.07.027
77. Ge X, Tang P, Rong Y, et al. Exosomal miR-155 from M1-polarized macrophages promotes EndoMT and impairs mitochondrial function via activating NF-κB signaling pathway in vascular endothelial cells after traumatic spinal cord injury. Redox Biol. 2021;41:101932. doi:10.1016/j.redox.2021.101932
78. Huang JH, He H, Chen Y-N, et al. Exosomes derived from M2 Macrophages Improve Angiogenesis and Functional Recovery after Spinal Cord Injury through HIF-1α/VEGF Axis. Brain Sci. 2022;12(10):1322. doi:10.3390/brainsci12101322
79. Zhang B, Lin F, Dong J, et al. Peripheral macrophage-derived exosomes promote repair after spinal cord injury by inducing local anti-inflammatory type microglial polarization via increasing autophagy. Int J Biol Sci. 2021;17(5):1339–1352. doi:10.7150/ijbs.54302
80. Zhou Z, Li C, Bao T, et al. Exosome-shuttled miR-672-5p from anti-inflammatory microglia repair traumatic spinal cord injury by inhibiting AIM2/ASC/caspase-1 signaling pathway mediated neuronal pyroptosis. J Neurotrauma. 2022;39(15–16):1057–1074. doi:10.1089/neu.2021.0464
81. Zhang J, Hu D, Li L, et al. M2 microglia-derived exosomes promote spinal cord injury recovery in mice by alleviating A1 astrocyte activation. Mol Neurobiol. 2024;61(9):7009–7025. doi:10.1007/s12035-024-04026-6
82. Ye Y, Hao J, Hong Z, et al. Downregulation of MicroRNA-145-5p in activated microglial exosomes promotes astrocyte proliferation by removal of smad3 inhibition. Neurochem Res. 2022;47(2):382–393. doi:10.1007/s11064-021-03446-3
83. Calabrese EJ. Hormesis and bone marrow stem cells: enhancing cell proliferation, differentiation and resilience to inflammatory stress. Chem Biol Interact. 2022;351:109730. doi:10.1016/j.cbi.2021.109730
84. Sheng X, Zhao J, Li M, et al. Bone marrow mesenchymal stem cell-derived exosomes accelerate functional recovery after spinal cord injury by promoting the phagocytosis of macrophages to clean myelin debris. Front Cell Dev Biol. 2021;9:772205. doi:10.3389/fcell.2021.772205
85. Li C, Qin T, Zhao J, et al. Bone marrow mesenchymal stem cell-derived exosome-educated macrophages promote functional healing after spinal cord injury. Front Cell Neurosci. 2021;15:725573. doi:10.3389/fncel.2021.725573
86. Wu H, Wang Q, Liao Y, et al. MSC-derived exosomes deliver ZBTB4 to mediate transcriptional repression of ITIH3 in astrocytes in spinal cord injury. Brain Res Bull. 2024;212:110954. doi:10.1016/j.brainresbull.2024.110954
87. Li S, Liao X, He Y, et al. Exosomes derived from NGF-overexpressing bone marrow mesenchymal stem cell sheet promote spinal cord injury repair in a mouse model. Neurochem Int. 2022;157:105339. doi:10.1016/j.neuint.2022.105339
88. Liu W, Wang Y, Gong F, et al. Exosomes derived from bone mesenchymal stem cells repair traumatic spinal cord injury by suppressing the activation of A1 neurotoxic reactive astrocytes. J Neurotrauma. 2019;36(3):469–484. doi:10.1089/neu.2018.5835
89. Xin W, Qiang S, Jianing D, et al. Human bone marrow mesenchymal stem cell-derived exosomes attenuate blood-spinal cord barrier disruption via the TIMP2/MMP pathway after acute spinal cord injury. Mol Neurobiol. 2021;58(12):6490–6504. doi:10.1007/s12035-021-02565-w
90. Zhou Y, Wen -L-L, Li Y-F, et al. Exosomes derived from bone marrow mesenchymal stem cells protect the injured spinal cord by inhibiting pericyte pyroptosis. Neural Regen Res. 2022;17(1):194–202. doi:10.4103/1673-5374.314323
91. Jiang C, Wang X, Jiang Y, et al. The anti-inflammation property of olfactory ensheathing cells in neural regeneration after spinal cord injury. Mol Neurobiol. 2022;59(10):6447–6459. doi:10.1007/s12035-022-02983-4
92. Jiang Y, Guo J, Tang X, et al. The immunological roles of olfactory ensheathing cells in the treatment of spinal cord injury. Front Immunol. 2022;13:881162. doi:10.3389/fimmu.2022.881162
93. Fan H, Chen Z, Tang H-B, et al. Exosomes derived from olfactory ensheathing cells provided neuroprotection for spinal cord injury by switching the phenotype of macrophages/microglia. Bioeng Transl Med. 2022;7(2):e10287. doi:10.1002/btm2.10287
94. Luo Y, He Y-Z, Wang Y-F, et al. Adipose-derived mesenchymal stem cell exosomes ameliorate spinal cord injury in rats by activating the Nrf2/HO-1 pathway and regulating microglial polarization. Folia Neuropathol. 2023;61(3):326–335. doi:10.5114/fn.2023.130455
95. Sung SE, Seo M-S, Kim Y-I, et al. Human epidural AD-MSC exosomes improve function recovery after spinal cord injury in rats. Biomedicines. 2022;10(3):678. doi:10.3390/biomedicines10030678
96. Liang Y, Wu J-H, Zhu J-H, et al. Exosomes secreted by hypoxia-pre-conditioned adipose-derived mesenchymal stem cells reduce neuronal apoptosis in rats with spinal cord injury. J Neurotrauma. 2022;39(9–10):701–714. doi:10.1089/neu.2021.0290
97. Li S, Luo L, He Y, et al. Dental pulp stem cell-derived exosomes alleviate cerebral ischaemia-reperfusion injury through suppressing inflammatory response. Cell Prolif. 2021;54(8):e13093. doi:10.1111/cpr.13093
98. Luo L, He Y, Wang X, et al. Potential roles of dental pulp stem cells in neural regeneration and repair. Stem Cells Int. 2018;2018:1731289. doi:10.1155/2018/1731289
99. Ji L, Bao L, Gu Z, et al. Comparison of immunomodulatory properties of exosomes derived from bone marrow mesenchymal stem cells and dental pulp stem cells. Immunol Res. 2019;67(4):432–442. doi:10.1007/s12026-019-09088-6
100. Liu C, Hu F, Jiao G, et al. Dental pulp stem cell-derived exosomes suppress M1 macrophage polarization through the ROS-MAPK-NFκB P65 signaling pathway after spinal cord injury. J Nanobiotechnology. 2022;20(1):65 doi:10.1186/s12951-022-01273-4.
101. Ju C, Liu R, Zhang Y-W, et al. Mesenchymal stem cell-associated lncRNA in osteogenic differentiation. Biomed Pharmacother. 2019;115:108912. doi:10.1016/j.biopha.2019.108912
102. Silvestro S, Mazzon E. MiRNAs as promising translational strategies for neuronal repair and regeneration in spinal cord injury. Cells. 2022;11(14):2177. doi:10.3390/cells11142177
103. Connerty P, Ahadi A, Hutvagner G. RNA binding proteins in the miRNA pathway. Int J Mol Sci. 2015;17(1):31. doi:10.3390/ijms17010031
104. Chandrasekera D, Katare R. Exosomal microRNAs in diabetic heart disease. Cardiovasc Diabetol. 2022;21(1):122 doi:10.1186/s12933-022-01544-2.
105. Cui Y, Qi Y, Ding L, et al. miRNA dosage control in development and human disease. Trends Cell Biol. 2024;34(1):31–47.
106. Kontidou E, Collado A, Pernow J, et al. Erythrocyte-derived microRNAs: emerging players in cardiovascular and metabolic disease. Arterioscler Thromb Vasc Biol. 2023;43(5):628–636. doi:10.1161/ATVBAHA.123.319027
107. Zhu Z, Huang X, Du M, et al. Recent advances in the role of miRNAs in post-traumatic stress disorder and traumatic brain injury. Mol Psychiatry. 2023;28(7):2630–2644. doi:10.1038/s41380-023-02126-8
108. He X, Zhang J, Guo Y, et al. Exosomal miR-9-5p derived from BMSCs alleviates apoptosis, inflammation and endoplasmic reticulum stress in spinal cord injury by regulating the HDAC5/FGF2 axis. Mol Immunol. 2022;145:97–108. doi:10.1016/j.molimm.2022.03.007
109. Liu W, Rong Y, Wang J, et al. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J Neuroinflammation. 2020;17(1):47. doi:10.1186/s12974-020-1726-7
110. Li C, Qin T, Liu Y, et al. Microglia-derived exosomal microRNA-151-3p enhances functional healing after spinal cord injury by attenuating neuronal apoptosis via regulating the p53/p21/CDK1 signaling pathway. Front Cell Dev Biol. 2021;9:783017. doi:10.3389/fcell.2021.783017
111. Yang Y, Li Y, Zhang S, et al. miR-199a-5p from bone marrow mesenchymal stem cell exosomes promotes the proliferation of neural stem cells by targeting GSK-3β. Acta Biochim Biophys Sin. 2023;55(5):783–794. doi:10.3724/abbs.2023024
112. Shao Y, Wang Q, Liu L, et al. Alleviation of spinal cord injury by MicroRNA 137-overexpressing bone marrow mesenchymal stem cell-derived exosomes. Tohoku J Exp Med. 2023;259(3):237–246. doi:10.1620/tjem.2022.J118
113. Shao Y, Wang Q, Liu L, et al. Exosomes from microRNA 146a overexpressed bone marrow mesenchymal stem cells protect against spinal cord injury in rats. J Orthop Sci. 2023;28(5):1149–1156. doi:10.1016/j.jos.2022.07.013
114. Jiang Z, Zhang J. Mesenchymal stem cell-derived exosomes containing miR-145-5p reduce inflammation in spinal cord injury by regulating the TLR4/NF-κB signaling pathway. Cell Cycle. 2021;20(10):993–1009. doi:10.1080/15384101.2021.1919825
115. Zhang A, Bai Z, Yi W, et al. Overexpression of miR-338-5p in exosomes derived from mesenchymal stromal cells provides neuroprotective effects by the Cnr1/Rap1/Akt pathway after spinal cord injury in rats. Neurosci Lett. 2021;761:136124. doi:10.1016/j.neulet.2021.136124
116. Chen Y, Tian Z, He L, et al. Exosomes derived from miR-26a-modified MSCs promote axonal regeneration via the PTEN/AKT/mTOR pathway following spinal cord injury. Stem Cell Res Ther. 2021;12(1):224. doi:10.1186/s13287-021-02282-0
117. Chang Q, Hao Y, Wang Y, et al. Bone marrow mesenchymal stem cell-derived exosomal microRNA-125a promotes M2 macrophage polarization in spinal cord injury by downregulating IRF5. Brain Res Bull. 2021;170:199–210. doi:10.1016/j.brainresbull.2021.02.015
118. Wang Y, Lai X, Wu D, et al. Umbilical mesenchymal stem cell-derived exosomes facilitate spinal cord functional recovery through the miR-199a-3p/145-5p-mediated NGF/TrkA signaling pathway in rats. Stem Cell Res Ther. 2021;12(1):117. doi:10.1186/s13287-021-02148-5
119. Zhang M, Wang L, Huang S, et al. Exosomes with high level of miR-181c from bone marrow-derived mesenchymal stem cells inhibit inflammation and apoptosis to alleviate spinal cord injury. J Mol Histol. 2021;52(2):301–311. doi:10.1007/s10735-020-09950-0
120. Li C, Li X, Zhao B, et al. Exosomes derived from miR-544-modified mesenchymal stem cells promote recovery after spinal cord injury. Arch Physiol Biochem. 2020;126(4):369–375. doi:10.1080/13813455.2019.1691601
121. Huang JH, Xu Y, Yin X-M, et al. Exosomes derived from miR-126-modified MSCs promote angiogenesis and neurogenesis and attenuate apoptosis after spinal cord injury in rats. Neuroscience. 2020;424:133–145. doi:10.1016/j.neuroscience.2019.10.043
122. Yu T, Zhao C, Hou S, et al. Exosomes secreted from miRNA-29b-modified mesenchymal stem cells repaired spinal cord injury in rats. Braz J Med Biol Res. 2019;52(12):e8735. doi:10.1590/1414-431x20198735
123. Kang J, Li Z, Zhi Z, et al. MiR-21 derived from the exosomes of MSCs regulates the death and differentiation of neurons in patients with spinal cord injury. Gene Ther. 2019;26(12):491–503. doi:10.1038/s41434-019-0101-8
124. Ren ZW, Zhou J-G, Xiong Z-K, et al. Effect of exosomes derived from MiR-133b-modified ADSCs on the recovery of neurological function after SCI. Eur Rev Med Pharmacol Sci. 2019;23(1):52–60. doi:10.26355/eurrev_201901_16747
125. Xu J, Zhang J, Liu Q, et al. Bone marrow mesenchymal stem cells-derived exosomes promote spinal cord injury repair through the miR-497-5p/TXNIP/NLRP3 axis. J Mol Histol. 2024;56(1):16. doi:10.1007/s10735-024-10289-z
126. Chen H, Sun H, Yang Y, et al. Engineered melatonin-pretreated plasma exosomes repair traumatic spinal cord injury by regulating miR-138-5p/SOX4 axis mediated microglia polarization. J Orthop Translat. 2024;49:230–245. doi:10.1016/j.jot.2024.09.007
127. Dong J, Gong Z, Bi H, et al. BMSC-derived exosomal miR-219-5p alleviates ferroptosis in neuronal cells caused by spinal cord injury via the UBE2Z/NRF2 pathway. Neuroscience. 2024;556:73–85. doi:10.1016/j.neuroscience.2024.06.011
128. Tang Q, Chen W, Ke H, et al. Optical imaging detection of extracellular vesicles of miR-146 modified bone marrow mesenchymal stem cells promoting spinal cord injury repair. SLAS Technol. 2024;29(4):100172. doi:10.1016/j.slast.2024.100172
129. Wang W, Yao F, Xing H, et al. Exosomal miR-17-92 cluster from BMSCs alleviates apoptosis and inflammation in spinal cord injury. Biochem Genet. 2025;63(4):3363–3377. doi:10.1007/s10528-024-10876-5
130. Chen M, Lin Y, Guo W, et al. BMSC-derived exosomes carrying miR-26a-5p ameliorate spinal cord injury via negatively regulating EZH2 and activating the BDNF-TrkB-CREB signaling. Mol Neurobiol. 2024;61(10):8156–8174. doi:10.1007/s12035-024-04082-y
131. Gu J, Wu J, Wang C, et al. BMSCs-derived exosomes inhibit macrophage/microglia pyroptosis by increasing autophagy through the miR-21a-5p/PELI1 axis in spinal cord injury. Aging. 2024;16(6):5184–5206. doi:10.18632/aging.205638
132. Morishima Y, Kawabori M, Yamazaki K, et al. Intravenous administration of mesenchymal stem cell-derived exosome alleviates spinal cord injury by regulating neutrophil extracellular trap formation through exosomal miR-125a-3p. Int J Mol Sci. 2024;25(4):2406. doi:10.3390/ijms25042406
133. Sun Y, Liu Q, Qin Y, et al. Exosomes derived from CD271 + CD56 + bone marrow mesenchymal stem cell subpopoulation identified by single-cell RNA sequencing promote axon regeneration after spinal cord injury. Theranostics. 2024;14(2):510–527. doi:10.7150/thno.89008
134. Xue H, Ran B, Li J, et al. Bone marrow mesenchymal stem cell exosomes-derived microRNA-216a-5p on locomotor performance, neuronal injury, and microglia inflammation in spinal cord injury. Front Cell Dev Biol. 2023;11:1227440. doi:10.3389/fcell.2023.1227440
135. Yang D, Wei H, Sheng Y, et al. Circ_0006640 transferred by bone marrow-mesenchymal stem cell-exosomes suppresses lipopolysaccharide-induced apoptotic, inflammatory and oxidative injury in spinal cord injury. J Orthop Surg Res. 2024;19(1):50. doi:10.1186/s13018-023-04523-9
136. Shao M, Ye S, Chen Y, et al. Exosomes from hypoxic ADSCs ameliorate neuronal damage post spinal cord injury through circ-Wdfy3 delivery and inhibition of ferroptosis. Neurochem Int. 2024;177:105759. doi:10.1016/j.neuint.2024.105759
137. Zhao Y, Chen Y, Wang Z, et al. Bone marrow mesenchymal stem cell exosome attenuates inflammasome-related pyroptosis via delivering circ_003564 to improve the recovery of spinal cord injury. Mol Neurobiol. 2022;59(11):6771–6789. doi:10.1007/s12035-022-03006-y
138. Tian F, Yang J, Xia R. Exosomes secreted from circZFHX3-modified mesenchymal stem cells repaired spinal cord injury through mir-16-5p/IGF-1 in mice. Neurochem Res. 2022;47(7):2076–2089. doi:10.1007/s11064-022-03607-y
139. Shao C, Chen Y, Yang T, et al. Mesenchymal stem cell derived exosomes suppress neuronal cell ferroptosis via lncGm36569/miR-5627-5p/FSP1 axis in acute spinal cord injury. Stem Cell Rev Rep. 2022;18(3):1127–1142 doi:10.1007/s12015-022-10327-x.
140. Liu J, Lin M, Qiao F, et al. Exosomes derived from lncRNA TCTN2-modified mesenchymal stem cells improve spinal cord injury by miR-329-3p/IGF1R axis. J Mol Neurosci. 2022;72(3):482–495. doi:10.1007/s12031-021-01914-7
141. Shao M, Jin M, Xu S, et al. Exosomes from long noncoding RNA-Gm37494-ADSCs repair spinal cord injury via shifting microglial M1/M2 polarization. Inflammation. 2020;43(4):1536–1547. doi:10.1007/s10753-020-01230-z
142. Wang Z, Song Y, Han X, et al. Long noncoding RNA PTENP1 affects the recovery of spinal cord injury by regulating the expression of miR-19b and miR-21. J Cell Physiol. 2020;235(4):3634–3645. doi:10.1002/jcp.29253
143. Xing X, Xu P, Xing X, et al. Effects of ADSC-derived exosome LRRC75A-AS1 on anti-inflammatory function after SCI. Appl Biochem Biotechnol. 2024;196(9):5920–5935. doi:10.1007/s12010-023-04836-9
144. Ivanov KI, Samuilova OV, Zamyatnin AA. The emerging roles of long noncoding RNAs in lymphatic vascular development and disease. Cell Mol Life Sci. 2023;80(8):197. doi:10.1007/s00018-023-04842-4
145. Li W, Wang YY, Xiao L, et al. Mysterious long noncoding RNAs and their relationships to human disease. Front Mol Biosci. 2022;9:950408. doi:10.3389/fmolb.2022.950408
146. Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways. Cancer Cell. 2016;29(4):452–463. doi:10.1016/j.ccell.2016.03.010
147. Sanchez-Mejias A, Tay Y. Competing endogenous RNA networks: tying the essential knots for cancer biology and therapeutics. J Hematol Oncol. 2015;8(1):30. doi:10.1186/s13045-015-0129-1
148. Marques TM, Gama-Carvalho M. Network approaches to study endogenous RNA competition and its impact on tissue-specific microRNA functions. Biomolecules. 2022;12(2):332. doi:10.3390/biom12020332
149. Tian F, Yang J, Xia R. Exosomes secreted from circZFHX3-modified mesenchymal stem cells repaired spinal cord injury through mir-16-5p/IGF-1 in mice. Neurochem Res. 2022;47(7):2076–2089 doi:10.1007/s11064-022-03607-y.
150. Zou Z, Li H, Xu G, et al. Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023;18:4751–4778. doi:10.2147/IJN.S417422
151. Morwood AJ, El-Karim IA, Clarke SA, et al. The role of extracellular matrix (ECM) adhesion motifs in functionalised hydrogels. Molecules. 2023;28(12):4616. doi:10.3390/molecules28124616
152. Li L, Zhang Y, Mu J, et al. Transplantation of human mesenchymal stem-cell-derived exosomes immobilized in an adhesive hydrogel for effective treatment of spinal cord injury. Nano Lett. 2020;20(6):4298–4305. doi:10.1021/acs.nanolett.0c00929
153. Cheng J, Chen Z, Liu C, et al. Bone mesenchymal stem cell-derived exosome-loaded injectable hydrogel for minimally invasive treatment of spinal cord injury. Nanomedicine. 2021;16(18):1567–1579. doi:10.2217/nnm-2021-0025
154. He X, Yang L, Dong K, et al. Biocompatible exosome-modified fibrin gel accelerates the recovery of spinal cord injury by VGF-mediated oligodendrogenesis. J Nanobiotechnology. 2022;20(1):360. doi:10.1186/s12951-022-01541-3
155. Mu J, Wu J, Cao J, et al. Rapid and effective treatment of traumatic spinal cord injury using stem cell derived exosomes. Asian J Pharm Sci. 2021;16(6):806–815. doi:10.1016/j.ajps.2021.10.002
156. Wang Q, Liu K, Cao X, et al. Plant-derived exosomes extracted from Lycium barbarum L. loaded with isoliquiritigenin to promote spinal cord injury repair based on 3D printed bionic scaffold. Bioeng Transl Med. 2024;9(4):e10646. doi:10.1002/btm2.10646
157. Luo Z, Peng W, Xu Y, et al. Exosomal OTULIN from M2 macrophages promotes the recovery of spinal cord injuries via stimulating Wnt/β-catenin pathway-mediated vascular regeneration. Acta Biomater. 2021;136:519–532. doi:10.1016/j.actbio.2021.09.026
158. Fan L, Liu C, Chen X, et al. Exosomes-loaded electroconductive hydrogel synergistically promotes tissue repair after spinal cord injury via immunoregulation and enhancement of myelinated axon growth. Adv Sci. 2022;9(13):e2105586. doi:10.1002/advs.202105586
159. Guan P, Fan L, Zhu Z, et al. M2 microglia-derived exosome-loaded electroconductive hydrogel for enhancing neurological recovery after spinal cord injury. J Nanobiotechnology. 2024;22(1):8. doi:10.1186/s12951-023-02255-w
160. Wang C, Xu M, Fan Q, et al. Therapeutic potential of exosome-based personalized delivery platform in chronic inflammatory diseases. Asian J Pharm Sci. 2023;18(1):100772. doi:10.1016/j.ajps.2022.100772
161. Abbas A, Huang X, Ullah A, et al. Enhanced spinal cord repair using bioengineered induced pluripotent stem cell-derived exosomes loaded with miRNA. Mol Med. 2024;30(1):168. doi:10.1186/s10020-024-00940-6
162. Malvandi AM, Rastegar-moghaddam SH, Ebrahimzadeh-Bideskan S, et al. Targeting miR-21 in spinal cord injuries: a game-changer? Mol Med. 2022;28(1):118. doi:10.1186/s10020-022-00546-w
163. Liu X, Zhang L, Xu Z, et al. A functionalized collagen-I scaffold delivers microRNA 21-loaded exosomes for spinal cord injury repair. Acta Biomater. 2022;154:385–400. doi:10.1016/j.actbio.2022.10.027
164. Spilman PR, Corset V, Gorostiza O, et al. Netrin-1 interrupts amyloid-β amplification, increases sAβPPα in vitro and in vivo, and improves cognition in a mouse model of Alzheimer’s disease. J Alzheimers Dis. 2016;52(1):223–242. doi:10.3233/JAD-151046
165. Hadas Y, Katz MG, Bridges CR, et al. Modified mRNA as a therapeutic tool to induce cardiac regeneration in ischemic heart disease. Wiley Interdiscip Rev Syst Biol Med. 2017;9(1). doi:10.1002/wsbm.1367
166. Lu X, Xu G, Lin Z, et al. Engineered exosomes enriched in netrin-1 modRNA promote axonal growth in spinal cord injury by attenuating inflammation and pyroptosis. Biomater Res. 2023;27(1):3. doi:10.1186/s40824-023-00339-0
167. Xiao Y, Hu X, Jiang P, et al. Thermos-responsive hydrogel system encapsulated engineered exosomes attenuate inflammation and oxidative damage in acute spinal cord injury. Front Bioeng Biotechnol. 2023;11:1216878. doi:10.3389/fbioe.2023.1216878
168. Ran N, Li W, Zhang R, et al. Autologous exosome facilitates load and target delivery of bioactive peptides to repair spinal cord injury. Bioact Mater. 2023;25:766–782. doi:10.1016/j.bioactmat.2022.07.002
169. Han Q, Jin W, Xiao Z, et al. The promotion of neural regeneration in an extreme rat spinal cord injury model using a collagen scaffold containing a collagen binding neuroprotective protein and an EGFR neutralizing antibody. Biomaterials. 2010;31(35):9212–9220. doi:10.1016/j.biomaterials.2010.08.040
170. Li X, Dai J. Bridging the gap with functional collagen scaffolds: tuning endogenous neural stem cells for severe spinal cord injury repair. Biomater Sci. 2018;6(2):265–271. doi:10.1039/C7BM00974G
171. Fan C, Li X, Xiao Z, et al. A modified collagen scaffold facilitates endogenous neurogenesis for acute spinal cord injury repair. Acta Biomater. 2017;51:304–316. doi:10.1016/j.actbio.2017.01.009
172. Zhang L, Fan C, Hao W, et al. NSCs migration promoted and drug delivered exosomes-collagen scaffold via a bio-specific peptide for one-step spinal cord injury repair. Adv Healthc Mater. 2021;10(8):e2001896. doi:10.1002/adhm.202001896
173. Sahab Negah S, Khooei A, Samini F, et al. Laminin-derived Ile-Lys-Val-ala-Val: a promising bioactive peptide in neural tissue engineering in traumatic brain injury. Cell Tissue Res. 2018;371(2):223–236. doi:10.1007/s00441-017-2717-6
174. Fu GQ, Wang -Y-Y, Xu Y-M, et al. Exosomes derived from vMIP-II-Lamp2b gene-modified M2 cells provide neuroprotection by targeting the injured spinal cord, inhibiting chemokine signals and modulating microglia/macrophage polarization in mice. Exp Neurol. 2024;377:114784. doi:10.1016/j.expneurol.2024.114784
175. Zeng J, Gu C, Sun Y, et al. Engineering of M2 macrophages-derived exosomes via click chemistry for spinal cord injury repair. Adv Healthc Mater. 2023;12(11):e2203391. doi:10.1002/adhm.202203391
176. Stewart AN, Kumari R, Bailey WM, et al. PTEN knockout using retrogradely transported AAVs transiently restores locomotor abilities in both acute and chronic spinal cord injury. Exp Neurol. 2023;368:114502. doi:10.1016/j.expneurol.2023.114502
177. Guo S, Perets N, Betzer O, et al. Intranasal delivery of mesenchymal stem cell derived exosomes loaded with phosphatase and tensin homolog siRNA repairs complete spinal cord injury. ACS Nano. 2019;13(9):10015–10028. doi:10.1021/acsnano.9b01892
178. Ma X, Zhang T, Luo Z, et al. Functional nano-vector boost anti-atherosclerosis efficacy of berberine in Apoe ((-/-)) mice. Acta Pharm Sin B. 2020;10(9):1769–1783. doi:10.1016/j.apsb.2020.03.005
179. Tian X, Wang P, Li T, et al. Self-assembled natural phytochemicals for synergistically antibacterial application from the enlightenment of traditional Chinese medicine combination. Acta Pharm Sin B. 2020;10(9):1784–1795. doi:10.1016/j.apsb.2019.12.014
180. Gao ZS, Zhang C-J, Xia N, et al. Berberine-loaded M2 macrophage-derived exosomes for spinal cord injury therapy. Acta Biomater. 2021;126:211–223. doi:10.1016/j.actbio.2021.03.018
181. Liu Y, Yang C-L, Yang B, et al. Prophylactic administration of fingolimod (FTY720) ameliorated experimental autoimmune myasthenia gravis by reducing the number of dendritic cells, follicular T helper cells and antibody-secreting cells. Int Immunopharmacol. 2021;96:107511. doi:10.1016/j.intimp.2021.107511
182. Li Y, Chen Y, Hu X, et al. Fingolimod (FTY720) hinders interferon-γ-mediated fibrotic scar formation and facilitates neurological recovery after spinal cord injury. J Neurotrauma. 2023;40(23–24):2580–2595. doi:10.1089/neu.2022.0387
183. Chen J, Zhang C, Li S, et al. Exosomes derived from nerve stem cells loaded with FTY720 promote the recovery after spinal cord injury in rats by PTEN/AKT signal pathway. J Immunol Res. 2021;2021:8100298. doi:10.1155/2021/8100298
184. Curto‐Reyes V, Llames S, Hidalgo A, et al. Spinal and peripheral analgesic effects of the CB 2 cannabinoid receptor agonist AM1241 in two models of bone cancer-induced pain. Br J Pharmacol. 2010;160(3):561–573. doi:10.1111/j.1476-5381.2009.00629.x
185. Shaikh II, Bhandari R, Singh S, et al. Therapeutic potential of EVs loaded with CB2 receptor agonist in spinal cord injury via the Nrf2/HO-1 pathway. Redox Rep. 2024;29(1):2420572. doi:10.1080/13510002.2024.2420572
186. Xu C, Jiang C, Li Z, et al. Exosome nanovesicles: biomarkers and new strategies for treatment of human diseases. MedComm. 2020;5(8):e660. doi:10.1002/mco2.660
187. Tang H, Yu D, Zhang J, et al. The new advance of exosome-based liquid biopsy for cancer diagnosis. J Nanobiotechnology. 2024;22(1):610. doi:10.1186/s12951-024-02863-0
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Activation of Neuroinflammation via mTOR Pathway is Disparately Regulated by Differential Target Multiplexed and Traditional Low-Rate Spinal Cord Stimulation in a Neuropathic Pain Model
Tilley DM, Vallejo R, Vetri F, Platt DC, Cedeno DL
Journal of Pain Research 2022, 15:2857-2866
Published Date: 13 September 2022
Long-Term Effects of Severe Burns on the Kidneys: Research Advances and Potential Therapeutic Approaches
Yang G, Tan L, Yao H, Xiong Z, Wu J, Huang X
Journal of Inflammation Research 2023, 16:1905-1921
Published Date: 1 May 2023
Exosomes as Vehicles for Noncoding RNA in Modulating Inflammation: A Promising Regulatory Approach for Ischemic Stroke and Myocardial Infarction
Lai Z, Ye T, Zhang M, Mu Y
Journal of Inflammation Research 2024, 17:7485-7501
Published Date: 21 October 2024
Exploration of Hub Genes and Immune Cell Infiltration Characteristics Associated With Spinal Cord Injury in Mice
Chen W, Zhang Q, Zhang Z, Ding Y, Zhang F, Chen G
Journal of Inflammation Research 2025, 18:2613-2628
Published Date: 20 February 2025
Tailored Extracellular Vesicles from Dental Stem Cells: Advances in Specific Modifications for Enhanced Therapeutic Applications
Liu X, Li Z, Fu J, Wang R, He J, Yao J, Ye Q, He Y
International Journal of Nanomedicine 2025, 20:8327-8341
Published Date: 26 June 2025