Back to Journals » International Journal of Nanomedicine » Volume 21
Exosomes as Emerging Nanocarriers for Targeted Cancer Therapy
Authors Du Y, Liu D, Liu J, Yu J, Hao Z, Zhang M, Li J, Peng X ![]()
Received 28 November 2025
Accepted for publication 26 March 2026
Published 14 April 2026 Volume 2026:21 585042
DOI https://doi.org/10.2147/IJN.S585042
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Prakash Misra
Yao Du,1,* Di Liu,2,* Jiaxing Liu,1 Jinling Yu,1 Zhixiong Hao,1 Manru Zhang,1 Jianwen Li,3 Xueqiang Peng1,4,5
1Department of General Surgery, The Fourth Affiliated Hospital, China Medical University, Shenyang, 110032, People’s Republic of China; 2Department of Anesthesiology, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, 121001, People’s Republic of China; 3Department of General Surgery, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200025, People’s Republic of China; 4Group of Chronic Disease and Environmental Genomics, School of Public Health, China Medical University, Shenyang, 110122, People’s Republic of China; 5Shenyang Key Laboratory for Biomedical and Intelligent Mesh, Shenyang, Liaoning, 110032, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianwen Li, Department of General Surgery, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, 200025, People’s Republic of China, Email [email protected] Xueqiang Peng, Department of General Surgery, Fourth Affiliated Hospital, China Medical University, Shenyang, 110032, People’s Republic of China, Email [email protected]
Abstract: Exosomes are nanoscale lipid bilayer vesicles secreted by eukaryotic cells into biological fluids. As an important subtype of extracellular vesicles, they can mediate intercellular material exchange and signal transmission by carrying bioactive substances such as proteins, nucleic acids, and lipids, and participate in the maintenance of physiological homeostasis and the regulation of pathophysiological processes in the body. These biological characteristics make exosomes less likely to be recognized and cleared by the immune system after entering the human body, nor do they cause obvious immune rejection reactions, laying the foundation for their use as delivery carriers. Through engineering techniques, they can be modified, and nucleic acids, small molecule drugs, and other substances can be precisely encapsulated inside exosomes through artificial intervention, forming “exosome-treatment payload” complexes. These complexes can take advantage of the properties of exosomes to significantly enhance their permeability in human tissues and cells, easily cross biological barriers, and promote the enrichment of treatment payloads in specific sites such as tumor tissues, thereby effectively optimizing treatment outcomes. However, the current application of exosomes is still limited by low separation and purification efficiency, high preparation costs, and easy damage to vesicle structure, and the clinical transformation process needs to be accelerated. This review focuses on the latest progress in the research of exosome-targeted delivery platforms, combines existing exosome drug loading technologies, and analyzes the clinical application potential and core challenges of this delivery system in cancer treatment, aiming to provide research directions for the development and clinical transformation of exosome-mediated anti-tumor targeted therapy strategies.
Keywords: exosome, drug delivery system, cargo loading, cancer
Introduction
Cancer, as a significant global public health issue, maintains persistently high incidence and mortality rates.1 Although traditional cancer treatments (such as surgical resection, chemotherapy, and radiotherapy) are widely applied, they are often constrained by the complex physiological barriers of the human body, resulting in low drug bioavailability and inevitably causing severe toxic side effects in peripheral healthy tissues. To overcome these obstacles, drug delivery systems (DDS) refer to systems that comprehensively regulate the spatial, temporal, and dosing distribution of drugs or molecules within the organism, which have undergone rapid development over the past few decades. However, conventional synthetic delivery systems (including viral vectors, liposomes, and polymer nanoparticles) still face inherent limitations in clinical translation, such as high immunogenicity, short in vivo circulation half-life, and rapid clearance by the mononuclear phagocyte system (MPS).
Against this backdrop, exosomes (Exosomes) as an emerging natural delivery vehicle have garnered increasing attention. Since their formal nomenclature in 1985, exosomes have been redefined from initially being considered carriers of “cellular waste” to pivotal mediators regulating intercellular communication and remodeling the tumor microenvironment.2 As a significant component of extracellular vesicles (EVs), exosomes are widely present in various body fluids and naturally carry biomacromolecules such as proteins, nucleic acids, and lipids.3 With advancing research on exosomes, natural exosomes can be engineered into “engineered exosomes” through surface modification or content loading, thereby enhancing their targeting specificity and therapeutic efficacy, which opens new avenues for exosome applications.4–6 Compared to synthetic liposomes or viral vectors, exosomes exhibit unparalleled biological advantages: exceptional biocompatibility, minimal immunogenicity, and the ability to naturally traverse stringent biological barriers (eg., the blood-brain barrier). This enables more precise drug delivery to target cells, significantly improving the efficacy of tumor therapy. Consequently, utilizing exosomes as carriers for chemotherapeutic drug delivery may emerge as an effective anti-tumor strategy.7,8
Despite the increasing research on engineered exosomes, this field still faces a significant clinical translation gap. Current review literature predominantly focuses on describing the fundamental biogenesis of exosomes or their natural roles in the tumor microenvironment, while lacking in-depth and critical discussions on how to systematically overcome clinical application barriers through engineering approaches (such as severe off-target effects in vivo, heterogeneity in large-scale production, and long-term safety evaluation). To address this literature gap, this review systematically summarizes the latest advancements of exosomes as emerging nanocarriers for targeted cancer therapy. This article not only provides a detailed summary of advanced engineering strategies to enhance exosome targeting efficacy and drug loading (including macromolecular nucleic acids and gene-editing tools), but also highlights their advantages and disadvantages compared to other delivery systems, as well as their applications and challenges in clinical translation. The aim is to offer comprehensive theoretical support and forward-looking guidance for the clinical design and interdisciplinary translation of next-generation intelligent exosome-based drugs.
Exosome Composition, Biogenesis and Function
Exosomes are ubiquitous in biological fluids such as blood, saliva, urine, and cerebrospinal fluid (CSF).9 Depending on their compositional modifications, exosomes fall into two primary categories: naturally occurring and engineered variants. Natural exosomes are further differentiated based on their origin, distinguishing between animal- and plant-derived sources. Among animal-derived exosomes, subtypes include those shed by healthy tissues and those secreted by tumor cells.
Exosomes originate from the endosomal system and typically measure 30–150 nanometers in diameter. Their biogenesis commences with the invagination of the plasma membrane, forming early sorting endosomes (ESE).10 These early endosomes mature into late sorting endosomes (LSE) through the orchestration of the endosomal sorting complexes required for transport (ESCRT) machinery and associated regulatory proteins. Cargo trafficking between the Golgi apparatus and inward budding events generates multivesicular bodies (MVBs). MVBs may undergo two fates: fusion with lysosomes or autophagosomes for degradation, or merger with the plasma membrane to release extracellular vesicles as exosomes11,12 (Figure 1). The biogenesis of exosomes is not a unidirectional linear process, but rather a highly complex and finely regulated mechanism governed by multiple pathways with dynamic competitive and synergistic interactions. Currently, it is primarily classified into ESCRT-dependent and ESCRT-independent pathways, which are also the fundamental causes of exosomal cargo heterogeneity.13 In the ESCRT-dependent pathway, ESCRT-0, -I, -II, and-III complexes precisely sort specific protein cargoes (such as growth factor receptors) by recognizing ubiquitination tags and mediate their entry into multivesicular bodies (MVBs). The expression levels of HRS and TSG101 significantly influence the proteomic profiles and vesicle sizes of secreted exosomes, while ALIX not only assists in membrane cleavage but also directly determines the loading efficiency of specific transmembrane proteins such as MHC class II molecules. In contrast, the ESCRT-independent pathway primarily relies on lipid rafts (eg., ceramides, cholesterol) and four transmembrane proteins (eg., CD63, CD81, CD9). The ceramide production mediated by neutral sphingomyelinase (nSMase) and the lipid-mediated pathway driven by Rab31 GTPase both belong to this category. Notably, Rab31 GTPase specifically sorts cargo carrying epidermal growth factor receptor (EGFR) into CD63-positive exosomal subpopulations, whereas exosomes generated via the nSMase-dependent pathway are often enriched with specific lipids and RNA.14 It is noteworthy that previous studies often treated exosomes as homogeneous entities, masking functional differences among subpopulations mediated by distinct biological pathways. In reality, cells dynamically adjust the weighting of these two pathways in response to external stress conditions (eg., hypoxia, immune stress), secreting exosome subpopulations with high molecular heterogeneity and targeted specificity to meet diverse communication needs. This suggests that viewing exosomes as a single homogeneous population in future research and clinical applications presents significant limitations.
 |
Figure 1 Biogenesis of Exosomes. Exosomes originate from the endosomal system, beginning with the inward budding of the plasma membrane to form early sorting endosomes (ESEs). As ESEs mature into late sorting endosomes, they exchange cargo with the Golgi apparatus and subsequently undergo inward budding to generate intraluminal vesicles (ILVs), giving rise to multivesicular bodies (MVBs). During this stage, exosomal contents are selectively packaged. MVBs can either fuse with lysosomes or autophagosomes for degradation, or fuse with the plasma membrane to release the enclosed ILVs as exosomes into the extracellular space. Exosomes carry a wide array of bioactive molecules, including proteins and nucleic acids. Tetraspanins such as CD9, CD63, and CD81, which are enriched on the exosomal membrane, are commonly used as canonical markers for exosomes. |
The internalization mechanism of cells fundamentally determines the targeting efficiency and pharmacokinetic distribution of exosomes or nanocarriers in vivo.15 Among these,The endocytosis mediated by clathrin is the main pathway for receptor-ligand interactions.16 In highly heterogeneous malignant tumors (eg., triple-negative breast cancer, TNBC), surface-engineered exosomes (eg., targeting specific integrins or peptides) rely heavily on the CME pathway to enter target cells. The clinical value of this mechanism lies in its exceptional targeting specificity: it not only significantly enhances drug accumulation in complex tumor microenvironments but also precisely blocks metastasis-promoting communication between tumor cells and surrounding stromal cells (eg., lymphatic endothelial cells).17–19 Therefore, optimizing CME-based uptake strategies is a critical prerequisite for reducing systemic off-target toxicity and improving therapeutic outcomes such as inhibiting lymphatic metastasis.20 Unlike the high specificity of CME, phagocytosis provides a non-specific, bulk internalization pathway.21 Many oncogenic mutations (eg., Ras-driven tumors) significantly upregulate phagocytic activity to uptake extracellular macromolecular nutrients. Exosomes and biomimetic nanocarriers can effectively exploit this metabolic vulnerability of tumor cells to facilitate uptake.22,23 The high permeability of phagocytic vesicles not only exponentially increases the absolute uptake of intracellular drugs but also holds decisive clinical significance for enhancing the penetration efficiency of macromolecular chemotherapeutic agents into dense tumor tissues and overcoming drug resistance.24,25
The ability of exosomes to achieve efficient “endosomal escape” after being internalized by recipient cells, thereby releasing their carried nucleic acids or proteins into the cytoplasm to function, is a critical stage in exosome-mediated intercellular signaling. In early research on nanomedicine and engineered exosomes (eg., cationically modified exosomes, PEI-modified exosomes), the “proton sponge effect” was frequently used to explain the endosomal escape process. This mechanism posits that cationically modified exosomes absorb a large number of protons in the acidic endosomal environment, prompting chloride influx and ultimately leading to endosomal rupture through a sudden increase in osmotic pressure. However, the actual contribution and reproducibility of this theory in vivo have been highly controversial: substantial in vivo evidence indicates that relying solely on cation-mediated endosomal osmotic imbalance not only results in extremely low escape efficiency (typically <1%) but also readily induces severe irreversible cytotoxicity. Moreover, the complex buffering systems in vivo often prevent the proton sponge effect from reaching the threshold required for endosomal rupture, leaving most exosomal cargo trapped in lysosomes for degradation.26 Recent in vivo fluorescence tracing and membrane biophysical studies further confirm that the buffering capacity of endosomes under physiological conditions is insufficient to generate an osmotic gradient large enough to cause membrane rupture.27 In contrast, the efficient intracellular delivery mechanism of natural exosomes and the more rational escape pathways of engineered exosomes rely more on specific “membrane fusion” or direct membrane destabilization caused by localized charge interactions. Under the micro-acidic environment of late endosomes (pH 5.0–6.0), specific lipid conformations on the exosomal membrane undergo changes, or fusion proteins on its surface (such as viral-like envelope protein fragments) are activated, promoting direct fusion of the exosomal membrane with the endosomal boundary membrane. The reverse fusion process within multivesicular bodies (MVBs) is also a key physiological pathway for exosomes to evade lysosomal degradation and release their functional contents.28,29 Membrane fusion enables the carrier to directly release its contents into the cytoplasm of target cells, completely circumventing the capture by the endosome-lysosome system and avoiding the inactivation of a large number of drugs before they can exert their therapeutic effects.30–32 For therapeutic strategies that require direct action on intracellular targets or the nucleus, membrane fusion can enhance the absolute bioavailability of active substances by hundreds or thousands of times, making it the most ideal internalization strategy for achieving efficient immune microenvironment remodeling and improving long-term patient survival outcomes.33,34 Zhuo Y et al demonstrated that increasing the cholesterol content of the exosomal membrane promoted vesicle deformation and expanded membrane contact areas, significantly enhancing siRNA delivery into recipient cells.30 Kumari P et al further demonstrated that CD14—a member of the Transporters Associated with Antigen Processing and Inflammation (TAXI) protein family—functions as a critical mediator in lipopolysaccharide (LPS) uptake.Their findings suggested that LPS bound to exosomes fused more efficiently with endosomal membranes compared to free LPS, highlighting the unique role of exosomes in immune signal transduction.35
The biological importance of exosomes is rooted not only in their unique structure and composition but also in their functional versatility. Accumulating evidence indicates that exosomes, by transporting DNA, RNA, proteins, and metabolites, regulate cellular processes and disease progression.36,37 (Table 1). Advances in high-throughput sequencing and mass spectrometry have greatly expanded our understanding of exosome classification and functional diversity.
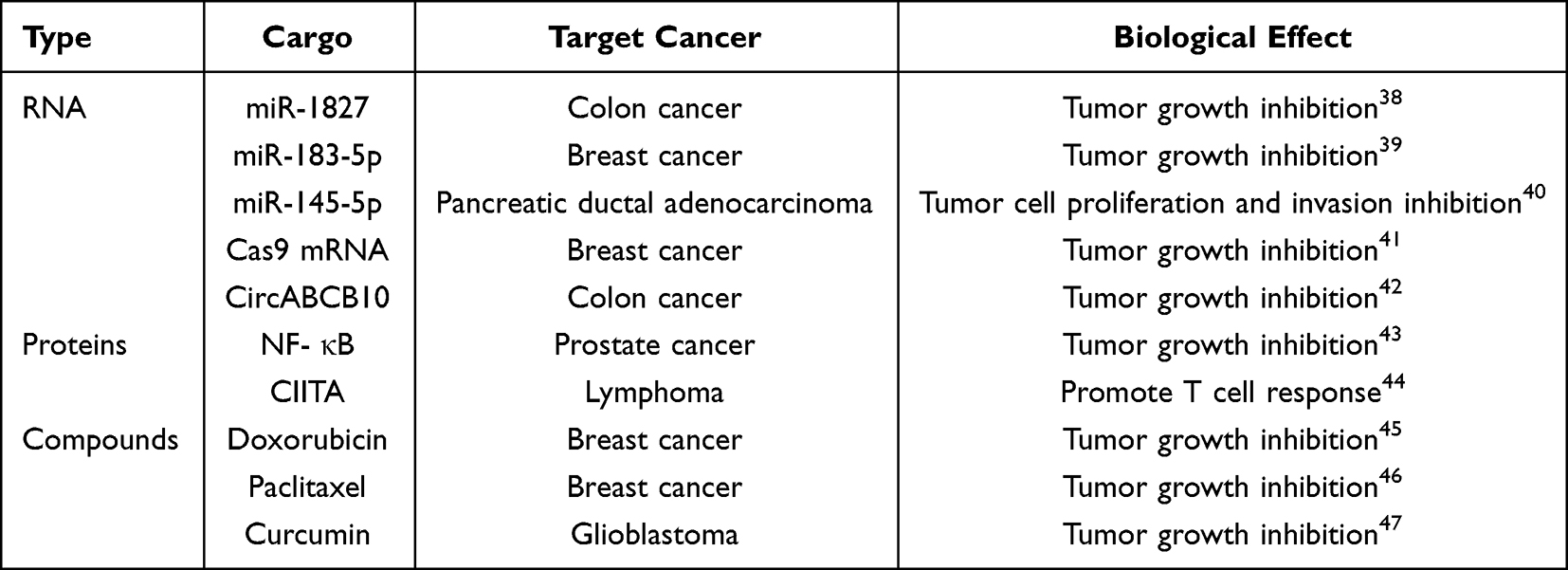 |
Table 1 Various Transport Substances of Exosome |
In cancer biology, exosomes accelerate tumor malignancy by remodeling the extracellular matrix (ECM) and driving tumor growth and dissemination.48 Moreover, these vesicles show promise as diagnostic tools for tracking tumor progression and enabling early clinical detection.11 Non-coding RNAs (eg., miRNAs, circRNAs) within exosomes exhibit exceptional stability in body fluids such as blood and urine due to protection by the lipid bilayer, making them highly promising biomarkers for liquid biopsy. Their diagnostic advantages have been confirmed in multiple large-scale clinical quantitative studies.49 In clinical applications for specific cancer types, different exosomal ncRNA combinations demonstrate superior diagnostic performance: For the early diagnosis of non-small cell lung cancer (NSCLC) and hepatocellular carcinoma (HCC), a clinical diagnostic model based on the combined detection of multiple exosomal miRNAs (eg., miR-1268b, miR-6075, miR-21) achieves a sensitivity of 95%–97.7% and specificity of 94.7%-99%, with the area under the receiver operating characteristic curve (AUC) remaining stable between 0.95 and 0.99, enabling precise screening for early-stage (Stage I) malignancies.50 In breast cancer diagnosis, the combination of miR-21 and miR-1246 in plasma exosomes demonstrates moderately high diagnostic efficacy, with an AUC of 0.73.51 Furthermore, circular RNAs (circRNAs) are emerging as next-generation diagnostic targets due to their exceptional nuclease resistance conferred by their closed-loop structure. Similarly, studies on exosomal circRNAs in blood and cerebrospinal fluid (CSF) have provided novel insights into the underlying mechanisms of neuropsychiatric disorders.52 These findings validate their practical translational value in precision medicine. Additionally, since the surface-specific antigens secreted by cells can reflect the properties of donor cells, researchers have identified multiple protein biomarkers in exosomes. These biomarkers not only facilitate the identification and isolation of exosomes but also hold promise as novel targets for disease diagnosis and treatment. These research achievements not only provide new perspectives for exosome studies but also lay a solid foundation for their application in disease diagnosis and therapy.
Isolation and Characterization of Exosomes
In exosome research, overcoming technical challenges remains a crucial task. Efficient, accurate isolation, purification, and identification of these nanoscale vesicles continues to be a significant hurdle. Exosome separation methods are typically based on properties like density, size, and antigenicity. However, the yield, purity, and physicochemical traits of isolated exosomes differ based on the chosen separation technique.53 Conventional approaches—including ultracentrifugation, density gradient separation, have been widely used to eliminate protein contaminants and cellular debris. Meanwhile, researchers are actively exploring novel technologies such as microfluidics, immunoaffinity capture, and nano-flow cytometry, which have demonstrated promising potential in recent years.54,55 To more intuitively demonstrate the advantages and disadvantages of the aforementioned separation techniques, we conducted a systematic comparative evaluation of the core performance metrics of different methods, detailing the key performance indicators and pros and cons of current mainstream exosome separation methods (Table 2). As previously mentioned, the high heterogeneity of exosomes and the limitations of extraction techniques remain significant technical challenges in current research. Traditional methods (such as ultracentrifugation, ultrafiltration, and precipitation) are widely applied but often involve trade-offs in terms of purity, recovery rate, or vesicle integrity. In contrast, emerging technologies (such as microfluidic chips and the EXODUS system) exhibit great potential for automated, high-throughput, and large-scale clinical applications in exosome separation.
 |
Table 2 Isolation Method of Exosome |
Following isolation and purification, due to the intrinsic heterogeneity of exosomes, detailed characterization and quantification through multiple approaches are necessary. These characterization methods are broadly categorized into two types: external (morphology and size analysis) and internal (membrane proteins, lipids, etc). Common techniques include transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), Western blotting of exosomal surface markers, tunable resistive pulse sensing (TRPS), and atomic force microscopy (AFM).65–67 These technologies provide comprehensive information on exosome morphology, size distribution, purity, concentration, and overall biophysical properties, which are critically important for advancing therapeutic and diagnostic applications, especially in oncology.68,69 In practice, the purity, quantity, and physicochemical properties of exosomes may vary depending on the separation conditions. Therefore, it is essential to comprehensively consider specific requirements and select appropriate separation and characterization methods.
Traditional Isolation Techniques
Ultracentrifugation is widely regarded as the benchmark method for exosome purification, encompassing both differential and density gradient-based protocols. Differential centrifugation achieves particle separation by manipulating centrifugal force and sedimentation time, while density gradient centrifugation leverages media to establish a density gradient for enhanced separation. Nonetheless, ultracentrifugation may cause exosome aggregation, contamination with lipoproteins, and potential membrane damage due to high-speed forces. Ultrafiltration utilizes size-selective membranes to fractionate exosomes from impurities, effectively excluding non-vesicular contaminants. Compared to ultracentrifugation, ultrafiltration offers improved purity, integrity, and shorter processing time; however, membrane adsorption can reduce exosome yield, and pore-induced platelet rupture remains a concern. SEC partitions particles according to size, allowing larger vesicles like exosomes to elute earlier while smaller molecules penetrate the pores. Despite its advantages, SEC can be time-consuming, costly, and may damage vesicle membranes. To overcome these constraints, researchers have engineered multiple commercial kits tailored to address these challenges. Polymer-based precipitation, typically utilizing polyethylene glycol (PEG) or salts to neutralize surface charges, aggregates exosomes for subsequent centrifugation-based recovery. While simple and scalable, precipitation methods often suffer from co-isolation of soluble proteins, resulting in lower purity and potential false positives.
Specific Affinity Techniques
Immunoaffinity capture relies on the binding of specific antibodies to exosome membrane proteins, providing exceptionally high specificity. Nevertheless, this approach requires stringent storage conditions, unsuitability for large-scale isolation, and the potential generation of interfering proteins.
Emerging High-Throughput Technologies
Microfluidics offer a high-throughput approach to exosome isolation, utilizing chips that separate vesicles based on size, density, and other properties. When biological fluids flow through microfluidic devices, microvortices enhance exosome interactions, while probes selectively capture vesicles, achieving >70% separation efficiency without compromising downstream analysis. Microfluidic technology serves as an emerging high-throughput approach that integrates principles such as electrophoresis, acoustics, or 3D porous structures, achieving separation efficiencies exceeding 70%. It offers advantages of being faster and more cost-effective with lower sample requirements, while also allowing exosomes to be stored in their native form, thereby retaining biological activity. Compared to traditional methods, microfluidic platforms are faster, more cost-effective, require smaller sample volumes, and better preserve exosome bioactivity.
Additionally, the EXODUS platform, an ultra-fast separation system, enables automatic, label-free exosome purification, demonstrating high efficiency and rapidity for large-scale clinical exosome enrichment from biofluids (eg., urine and plasma). Zhu Q et al utilized EXODUS to isolate urinary exosomes for lipidomic profiling of NAFLD and NASH patients, revealing potential biomarkers via machine learning analysis. EXODUS-based plasma exosome profiling has been applied to monitor dynamic treatment responses in esophageal squamous cell carcinoma. Overall, EXODUS combines efficiency, speed, and automation, making it well-suited for clinical and research-scale applications.
Advanced Characterization Technologies
Following isolation, advanced technologies such as Nano-Flow Cytometry and the ExoView system provide detailed, single-particle level characterization to address exosome heterogeneity. Nano-Flow Cytometry, utilizing light scattering and fluorescence, offers multidimensional analysis of size distribution, concentration, and surface markers detection (eg., the specific marker PODXL in stem cell-derived exosomes).70–72 The automated fluorescence analysis system ExoView, based on single-particle interferometric reflectance imaging sensor (SP-IRIS) technology, simultaneously performs counting, protein phenotyping, and cargo detection of exosome subpopulations in complex samples (eg., cerebrospinal fluid and plasma), providing a highly precise biomarker analysis tool for diagnostics in fields such as neurodegenerative diseases.73,74 In addition,Recent advancements in nanomaterial-peptide platforms have introduced novel strategies for exosome characterization. For instance, a smart sensing system integrating graphene oxide (GO) and a CD63-binding peptide (CP05) has been developed for the highly sensitive, dual-modal quantitative detection of tumor-derived exosomes in serum, significantly improving diagnostic accuracy.75
Preparation of Engineered Exosomes
Drug Loading
Genetic editing or chemical approaches can be employed to modify exosomes, thereby conferring novel functionalities upon these extracellular vesicles, increasing their potential in biomedical applications. To efficiently load drugs into exosomes, various techniques have been developed, including genetic engineering, chemical methods, physical techniques, and microfluidic systems. These methods allow drugs to be incorporated into exosomes without losing their biological activity.
The exosomal membrane structure serves a dual role: it shields encapsulated drugs from enzymatic hydrolysis and concurrently evades immune recognition and clearance, thereby conferring exceptional biocompatibility and stability to these extracellular vesicles.76,77 Leveraging their amphipathic nature, exosomes facilitate the targeted delivery of diverse therapeutic agents to recipient cells, rendering them highly effective carriers for drug delivery systems. Two primary strategies are employed for drug loading: pre-secretory (incorporation during exosome biogenesis) and post-secretory (loading into mature exosomes). Pre-secretory loading involves incubating the drug with cells before isolating the exosomes, allowing the cells to load the drug naturally into the exosomes.78 This method supports continuous production and avoids membrane degradation. Post-secretory loading involves adding the drug to isolated exosomes. Common methodologies for modifying exosomes include electroporation, repetitive freezing-thawing cycles, sonication, extrusion and surfactant disruption. Post-secretory loading is more efficient and gives better control over drug encapsulation, making it suitable for different drug delivery needs (Figure 2).
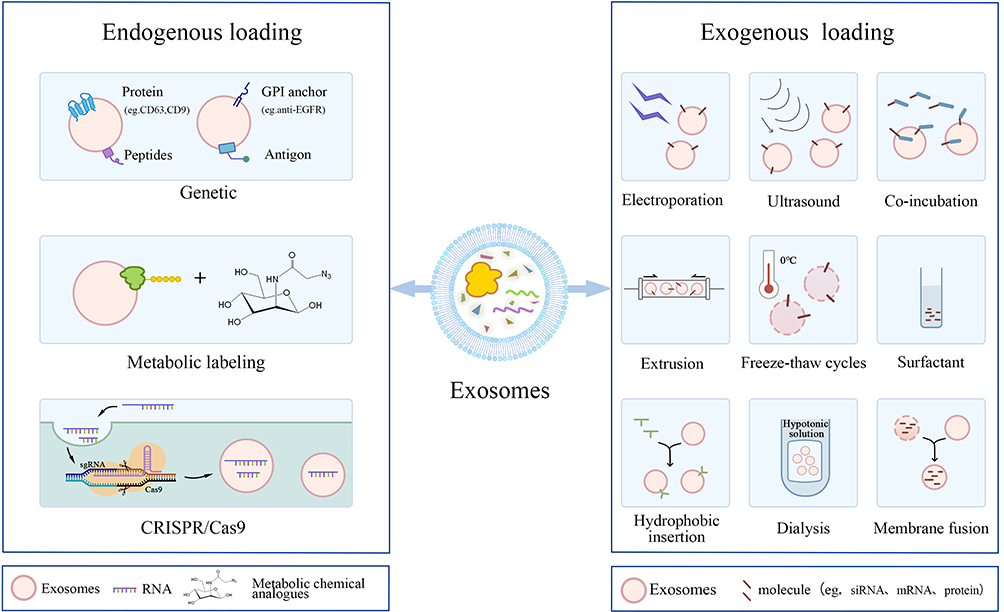 |
Figure 2 Drug Loading Methods for Exosomes. Drug loading into exosomes is generally categorized into two main approaches: pre-secretion loading and post-secretion loading. (Left) Pre-secretion loading involves introducing the drug into donor cells before exosome secretion, leveraging the cells’ endogenous mechanisms to incorporate the therapeutic agents into the exosomes. Common strategies include co-incubation of drugs with cells, gene transfection, and metabolic labeling et al (Right) Post-secretion loading is performed after exosomes have been isolated from the cells. Drugs are directly introduced into the purified exosomes using techniques such as electroporation, sonication, extrusion, freeze–thaw cycles, treatment with surfactants, hydrophobic insertion, and dialysis et al. These methods are adaptable to accommodate various drug delivery requirements. |
Co-Incubation
Recent research highlights that co-culturing hydrophobic drugs (eg., doxorubicin and paclitaxel) with parental cells facilitates their vesicular secretion via exosomes, thereby triggering cytotoxicity and suppressing tumor progression.79 For instance, He et al engineered NK cells to express interleukin-15 (IL-15), generating IL-15-enriched exosomes. Compared to unmodified NK-EVs, these cytokine-loaded vesicles exhibited superior tumor-targeting efficacy, resulting in elevated apoptosis rates among cancer cells.80 Pascucci et al incubated mesenchymal stem cells (MSC) with PTX before exosome isolation, resulting in significantly improved PTX delivery to cancer cells, enhancing cytotoxicity and apoptosis.81 The co-incubation method is simple, cost-effective, and works well for drugs that can easily pass through cell membranes. However, its efficacy may be reduced for other drugs, and the toxicity of the drug during the incubation process could have adverse effects.82
Transfection
Transfection is a technique for delivering therapeutic DNA, RNA, or proteins into parental cells, followed by the overexpression of these compounds in exosomes.83 For instance, to determine whether exosomes derived from adipose mesenchymal stem cells (AMSCs) could serve as effective vectors for delivering miR-199a-3p to enhance the chemotherapeutic efficacy of hepatocellular carcinoma, AMSCs were modified with miR-199a-3p via LV-199a infection (AMSC-199a). qRT-PCR validation confirmed a significant increase in miR-199a-3p expression in the treated cells (9.8 ± 1.1-fold), demonstrating that exosomes can directly deliver therapeutic agents to tumor cells.84 The transfection method offers advantages such as high reproducibility and operational simplicity, but it has low transfection efficiency and is highly dependent on cell viability.
Electroporation
Electroporation enhances the permeability of exosome membranes by applying an electric field, thereby loading hydrophilic small molecules such as amino acids and peptides. Yan et al utilized electroporation to encapsulate miR-31-5p in exosomes and quantified miR-31-5p levels from each sample. The results demonstrated that free miR-31-5p was almost completely degraded, whereas over 60% of the miR-31-5p loaded in exosomes remained stable This indicates that exosome-mediated miRNA delivery offers multiple advantages compared to direct miRNA delivery, including enhanced cellular uptake, resistance to miRNA enzymatic degradation, and improved miRNA bioavailability.85 However, electroporation may compromise the integrity of exosomes, thereby reducing their loading capacity.
Sonication
Translational studies demonstrate that ultrasound probes create transient pores on the exosome membrane, allowing hydrophilic small molecules to enter the exosomes. Xiang X et al loaded siRNA into exosomes via ultrasound treatment and calculated the encapsulation rate by the weight ratio of siRNA to loaded exosomes, achieving a drug loading efficiency as high as 24%.86 Yuan R et al loaded siHSP47 (EXO-siHSP47) into exosomes and improved the loading efficiency of siHSP47 in exosomes using ultrasound microfluidic methods, reaching 31.1%.87 Compared with methods such as electroporation, ultrasound treatment typically achieves higher drug loading efficiency. Kim MS et al measured the amount of paclitaxel loaded into exosomes using high-performance liquid chromatography (HPLC), with the highest loading efficiency of 28.3% achieved by ultrasound treatment, while incubation and electroporation yielded loading efficiencies of 1.4% and 5.3%, respectively.8
Membrane Fusion
Membrane fusion is the process by which two lipid bilayers approach and merge to form a stable structure. The drug is dissolved or dispersed in a buffer system and mixed with the exosome suspension. The mixture is then subjected to multiple extrusions through a lipid bilayer extruder, during which the complementary DNA strands from each vesicle are compressed together, triggering vesicle docking and membrane fusion to produce therapeutic exosomes. Xie et al fused siRNA-loaded liposomes targeting NFKBIZ with exosomes derived from corneal epithelial cells (CEC) to treat dry eye disease (DED), effectively delivering siRNA to corneal cells.88
Load Efficiency and Cargo Stability
Evaluating the encapsulation efficiency and cargo stability of exosomes is a crucial step in ensuring the effectiveness of therapeutic delivery systems. By quantitatively analyzing drug loading and stability, researchers can optimize the design of these platforms.
Regarding drug loading efficiency, post-secretion physical disruption methods (such as sonication, extrusion, and electroporation) generally exhibit significantly higher yields compared to passive incubation. As previously noted, sonication and microfluidic extrusion can boost the loading rates of hydrophobic or hydrophilic drugs to 28.3% and nearly 20%, respectively. However, electroporation may compromise the structural integrity of exosomes when loading certain macromolecular drugs, paradoxically reducing the final encapsulation efficiency. Pre-secretion loading strategies (eg., co-incubation), while highly convenient and cost-effective, typically yield extremely low loading efficiencies for most non-hydrophobic drugs due to the natural permeability constraints of cell membranes.
In terms of cargo and membrane stability, payload integrity relies on preserving the macromolecular conformation, biological activity, and therapeutic function under physiological conditions. In this regard, mild pre-secretory techniques (like co-incubation and transfection) are superior. Because they harness the natural vesicular secretion mechanisms, they maximally preserve the integrity of the exosomal lipid bilayer and the bioactivity of surface-targeting proteins. Conversely, while sonication and extrusion offer high loading rates, the intense mechanical shear forces, thermal effects, and electric field interference can cause irreversible membrane damage or lipid reorganization. Such structural instability not only accelerates premature drug leakage during in vivo circulation but may also trigger the abnormal aggregation and degradation of nucleic acids or protein macromolecules, ultimately compromising the controlled-release capabilities of the system.
As for the transition to industrial-scale production, co-incubation methods demonstrate exceptional scalability for large-batch processing, much like precipitation techniques. Electroporation also shows favorable batch-processing potential through high-throughput systems. Sonication, however, exhibits poor scalability for large-scale clinical-grade production due to equipment limitations, risks of localized overheating, and significant batch-to-batch variability. Notably, emerging microfluidic extrusion systems are increasingly viewed as the most promising solution for balancing high loading efficiency with outstanding scalability, thanks to their capacity for continuous, stable, and automated fluid control. Cargo release kinetics quantify the fraction of the drug liberated from exosomes over time, reflecting the loading capacity and sustained-release performance. Standard evaluation techniques include UV-Vis absorbance spectroscopy, fluorescent labeling, high-performance liquid chromatography (HPLC), and Tinopal assays. For instance, Chen et al engineered a graphene oxide nanocomposite and validated a 73% sustained cargo retention using UV-Vis spectrophotometry.89 Similarly, Sun et al encapsulated curcumin and utilized HPLC to correlate retention time with antioxidant efficacy.90 Additionally, the retention of exosome payloads is heavily regulated by multiple factors, including the hydrophobicity of drug and the exosome storage buffer.91,92
Ultimately, the selection of an exosome loading strategy requires a careful balance between encapsulation efficiency, structural preservation, and industrial scalability. Future advancements will likely rely on hybrid approaches or advanced microfluidic technologies to achieve the optimal combination of high therapeutic payload and robust clinical translation.
Construction of Delivery Systems
Exosomes demonstrate remarkable potential as therapeutic nanocarriers, leveraging their intrinsic biocompatibility and targeted payload translocation mechanisms to enhance bioavailability and mitigate off-target effects. First, exosomes are derived from the body itself, providing high biocompatibility, which reduces immunogenicity and toxicity during the drug delivery process. This characteristic makes exosomes highly promising for drug delivery applications. Second, exosomes have a distinct membrane structure that can effectively encapsulate drug molecules, reducing degradation and loss, thus improving drug bioavailability. Nevertheles, the transduction efficacy of exosomes is regulated by both the parent cells and the recipient cells, and the yield varies significantly depending on the source of the exosomes. Natural exosomes also have the drawback of poor targeting ability. To enhance drug delivery efficiency and minimize side effects, researchers are focusing on developing exosome delivery systems with high targeting specificity. By designing specific targeting molecules and functionalizing the surface of exosomes with targeting ligands, active agents achieve site-specific accumulation in cells or tissues via exosomal carriers, thereby augmenting the therapeutic efficacy of exosome-mediated drug delivery systems (Figure 3).
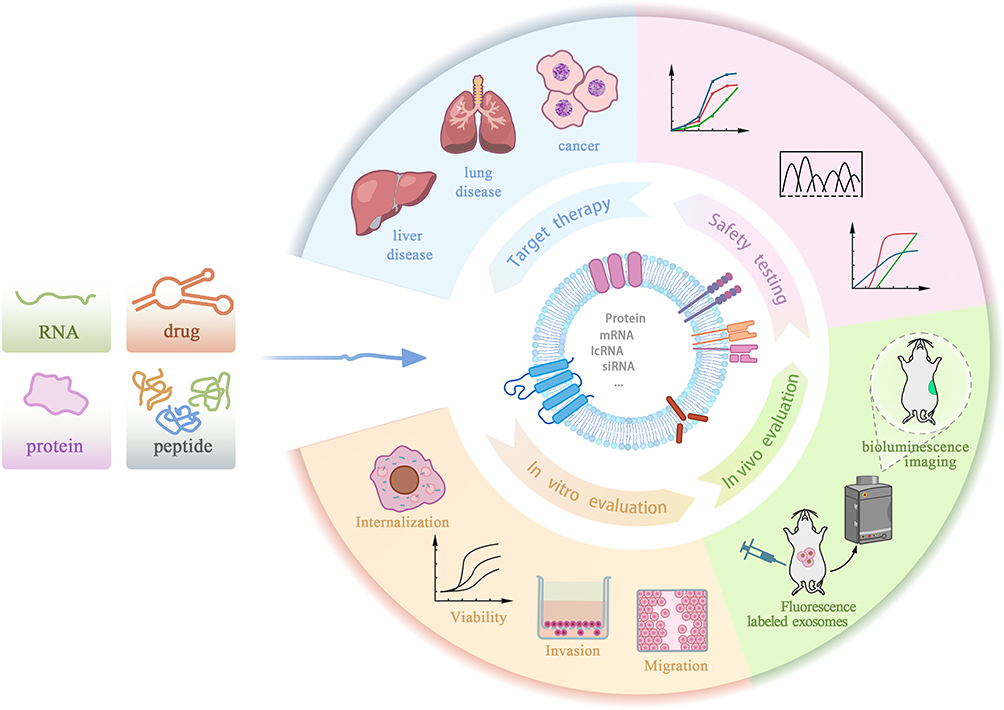 |
Figure 3 Validation and Application Workflow of Drug-Loaded Exosomes. Evaluation of the therapeutic efficacy and safety of engineered exosomes via in vitro, in vivo, and clinical analyses. ①In Vitro Evaluation: Exosome-mediated drug delivery efficiency is evaluated by measuring intracellular drug accumulation and assessing cell viability or functional changes post-treatment. ②In Vivo Evaluation: Disease models in mice are used to study the biodistribution and therapeutic efficacy of drug-loaded exosomes in a physiological setting. ③Safety Testing: The safety of engineered exosomes is assessed by examining drug loading efficiency, release kinetics, and potential cytotoxic or systemic adverse effects. ④Targeted Therapy: Engineered exosomes are utilized for targeted treatment of specific diseases, customized to the unique pathological features and therapeutic needs of each condition. |
Targeted Modification
The biodistribution of natural exosomes after intravenous injection exhibits high localization, with over 80% of systemically administered exosomes being rapidly cleared by the mononuclear phagocyte system (MPS), primarily accumulating in the liver, spleen, and lungs. This significant off-target uptake not only substantially reduces the effective dose delivered to the tumor site but may also induce potential hepatosplenotoxicity.The principle of targeted therapy is to precisely act on diseased cells or tissues through specific drugs or carriers, thereby minimizing damage to normal cells. Surface ligand engineering is a key strategy to overcome this natural distribution barrier. By employing methods such as chemical modification, physical techniques, or gene editing, specific targeting ligands—including antibodies, peptides, or other biomolecules—can be introduced on the outer layer of exosomes. These ligands direct exosomes to cells expressing the corresponding receptors, enabling precise targeting and delivery to specific cells.93–95 For example, the EGFR-targeting GE11 peptide can be conjugated to exosomes, enabling the selective delivery of tumor-suppressive miRNAs to EGFR-overexpressing breast carcinomas.96 Koh et al modified exosome surfaces with SIRP-α, designing exosomes with SIRP-α variants that interfere with the CD47-SIRP-α interaction between cancer cells and bone marrow-derived macrophages (BMDM), enhancing the macrophage-guided phagocytosis of tumor cells.97 Alternatively, ligands can be used to anchor proteins in cells, which are then incorporated into secreted exosomes. This targeted delivery reduces side effects on non-target cells and enhances therapeutic efficacy. For instance, Liu Y et al modified exosomes with a neuron-specific RVG peptide, which binds to acetylcholine receptors on neurons, enabling the brain-specific delivery of MOR siRNA, effectively inhibiting morphine relapse by downregulating MOR expression.98 Aptamers, short oligonucleotides that specifically recognize target molecules, were previously conjugated to cholesterol moieties on exosome surfaces. A bivalent RNA aptamer targeting PSMA enhanced PSMA-mediated endocytosis in prostate cancer cells, facilitating exosome uptake and promoting antitumor efficacy.99 Pi et al modified exosome surfaces with RNA ligands non-covalently attached to folic acid or cholesterol, altering the surface properties of exosomes to more effectively deliver siRNA and miRNA to tumor regions, thereby enhancing anti-tumor effects.100 Antibody fragments targeting unique cell epitopes are another common approach for exosome-based targeting. Wiklander OPB et al successfully modified exosome surfaces to incorporate Fc-specific antibodies, enabling Fc-EVs to target PD-L1-expressing cancer cells. When loaded with doxorubicin, these exosomes reduced tumor burden and extended survival in melanoma mouse models.101
Integrin-targeting peptides (eg., iRGD) or nanobodies targeting the epidermal growth factor receptor (EGFR) are modified onto the surface of exosomes. This surface engineering not only significantly enhances the specific uptake of exosomes in tumor parenchymal cells through receptor-ligand-mediated endocytosis, but more importantly, it competitively reduces nonspecific cell recognition by altering the surface charge and steric hindrance of exosomes.102 Therefore, ligand modification is crucial for reshaping global biodistribution and reducing off-target toxicity.103,104 Furthermore, when introducing targeting ligands, the potential impact of these exogenous modifications on immune system recognition is often overlooked. The physicochemical properties of surface ligands (eg., hydrophilicity, hydrophobicity, charge distribution) may be recognized by serum opsonins (such as complement proteins or immunoglobulins), thereby accelerating phagocytic clearance.105 To achieve active targeting while evading immune recognition, a dual surface engineering strategy has been developed. For example, tumor-targeting ligands are not only modified on the exosome surface but also co-expressed with the CD47 molecule. CD47, as a potent “don’t eat me” signal, binds to the SIRPα receptor on macrophage surfaces, activates SHP-1/2 phosphatase to emit inhibitory signals, and directly blocks phagocytosis.106 This strategy effectively prolongs the circulation half-life of target-expressed exosomes in the bloodstream, providing sufficient time windows for their penetration of vascular barriers into the tumor microenvironment. However, it must be noted that if ligand modification density is excessively high or cross-linking agents are immunogenic, acquired immune responses may still be induced. Therefore, future designs of target-expressed exosomes must strike an precise balance between “maximizing targeting affinity” and “minimizing immunogenicity”.107
Engineering Strategies: Targeted Efficacy, and Safety of Exosome Delivery
With ongoing advancements in biotechnology, various strategies for engineering exosomes have emerged. A major frontier in this domain is the incorporation of macromolecular nucleic acids and genome editing tools. Among them, gene editing techniques, particularly the CRISPR-Cas9 system, allow researchers to edit specific genes, enabling the manipulation of exosome composition, size, and function to meet specific therapeutic needs. However, loading large molecular cargos (such as Cas9 proteins, large mRNA transcripts, or plasmid DNA) presents significant challenges. Conventional physical encapsulation methods often suffer from low loading efficiency, exosome aggregation, or structural disruption of the lipid bilayer due to the sheer size of these macromolecules.108
To overcome these barriers, emerging membrane permeabilization and endogenous loading techniques have been rapidly developed. Advanced membrane permeabilization methods, such as microfluidic-assisted gentle electroporation or the use of mild chemical permeabilizing agents (eg., saponin), can transiently destabilize the exosomal membrane, facilitating the entry of large genome-editing tools while preserving vesicle integrity and cargo bioactivity.109 Alternatively, endogenous loading provides a highly efficient biological solution. By genetically engineering producer cells to co-express specific sorting motifs (such as viral RNA-binding proteins) or by fusing Cas9 to exosome-associated tetraspanins (eg., CD63), macromolecules can be actively recruited into vesicles during biogenesis. Consequently, Cas9 proteins are selectively isolated and incorporated into exosomes, forming complexes with sgRNA.110,111 These engineered sgRNA-Cas9-loaded exosomes provide a reliable and effective delivery method that addresses the limitations of traditional viral or lipid-based vectors.111–113 Kim et al showed that CRISPR/Cas9-engineered exosomes triggered apoptosis in ovarian cancer cells via PARP-1 suppression, highlighting the therapeutic potential of exosome-based CRISPR delivery. Emerging evidence also demonstrates that CRISPR/Cas9-engineered exosomes targeting PARP-1 markedly augmented the cisplatin responsiveness of SKOV3 ovarian cancer cells and suppressed tumor growth in murine xenograft models.114
Given the potent capabilities of these engineered exosomes—especially those carrying genome-editing tools or nanomaterials—evaluating their targeted efficacy and safety profile is a critical step to ensure their clinical applicability. The therapeutic potential and safety profiles of exosome-based delivery platforms must be systematically assessed via in vitro cellular assays and in vivo animal models. Jiang L et al engineered exosomes co-loaded with triptolide (TPL) and TRAIL for melanoma-targeted therapy, which effectively inhibited tumor growth while mitigating TPL-induced systemic toxicity, exhibiting excellent biocompatibility.115 Xie et al developed a machine learning surface-enhanced Raman scattering (SERS) platform to quantify HER2 expression dynamics in breast malignancy cells, enabling longitudinal monitoring of drug-loaded exosome efficacy.116 This technology captures temporal changes in cellular viability, supporting data-driven therapeutic decisions. Ultimately, as the development and clinical translation of engineered exosomes accelerate, ethical frameworks must be rigorously applied to safeguard participant welfare.
Comparison to Other Delivery Systems
Although engineered exosomes demonstrate unprecedented advantages in precision delivery, an objective side-by-side comparison of their pros and cons with widely applied traditional carriers (eg., liposomes, viral vectors, and polymers) can more clearly define their unique niche and current challenges in modern nanomedicine. Viral vectors exhibit high transfection efficiency and long-term gene expression capabilities. However, their clinical application is severely hampered by critical limitations, including high immunogenicity, limited cargo capacity, and the inherent risk of insertional mutagenesis. Synthetic nanocarriers such as liposomes and polymeric nanoparticles represent the most clinically advanced and commercially viable delivery systems. Liposomes, as one of the earliest lipid-based nanoparticle systems, effectively prevent drug dilution, degradation, or inactivation in the bloodstream by encapsulating the drugs. Similarly, polymeric nanoparticles (eg., PLGA) offer highly tunable properties and controlled drug release kinetics. Synthetic carriers, offer significant advantages from a manufacturing perspective, such as highly scalable production, precise control over physicochemical properties, and standardized drug loading capacities.Nevertheless, synthetic drug delivery approaches still encounter multiple hurdles, including inadequate specific targeting, harmful effects from the carriers, the ability to trigger immune responses, and restricted therapeutic effectiveness. For instance,the cationic lipids utilized in LNPs and the acidic degradation byproducts of polymeric materials can induce distinct cellular toxicity and elicit strong innate immune responses. Furthermore, synthetic nanoparticles are prone to rapid clearance by the mononuclear phagocyte system (MPS) and struggle to passively penetrate formidable biological barriers, such as the blood-brain barrier (BBB), without complex and often unstable surface modifications. In conclusion, the clinical application of lipid-based carriers remains challenging, due to issues like low bioavailability, toxicity, rapid clearance from the bloodstream, and the potential for immune responses.
In sharp contrast, exosomes—as naturally occurring vesicular carriers—completely circumvent the risk of mutagenesis and exhibit significantly lower immunogenicity, offering a much safer alternative for systemic administration. Compared to these synthetic counterparts, exosomes offer enhanced biocompatibility and lower immunogenicity. Exosomes, which stem from biological systems, inherently possess enhanced biocompatibility, profound ability to cross the BBB, and highly specific cellular tropism driven by their natural surface proteins (eg., tetraspanins and integrins).Once their delivery mission is accomplished, exosomes are naturally metabolized and eliminated from the body, thereby minimizing long-term accumulation toxicity.Moreover, because of their tiny size, they can effortlessly pass through blood vessel walls and the extracellular matrix. When compared with other delivery systems, exosomes trigger only minor immune reactions. They are also deemed to have the least tumorigenic potential. These characteristics make exosomes a highly promising option for targeted drug delivery.
However, despite these unique biological advantages, the exosome delivery system is not without significant limitations. First, unlike the highly reproducible and massive synthesis of LNPs, exosome production from cell cultures suffers from profoundly low yields, making clinical-scale manufacturing economically and technically daunting. Second, exosomes exhibit significant molecular and structural heterogeneity, heavily depending on the producer cell type and its physiological state.Third, while synthetic carriers easily encapsulate large therapeutic payloads during bottom-up self-assembly, introducing exogenous cargoes into pre-formed exosomes via physical or chemical methods (eg., electroporation) is remarkably inefficient and often compromises vesicle integrity. Finally, the lack of universally standardized, high-yield, and high-purity isolation techniques remains a formidable bottleneck, significantly impeding the translation of engineered exosomes from laboratory concepts to robust clinical therapeutics.Overcoming these manufacturing and standardization bottlenecks remains a critical hurdle before exosomes can fully outcompete mature synthetic delivery platforms in clinical settings (Table 3).
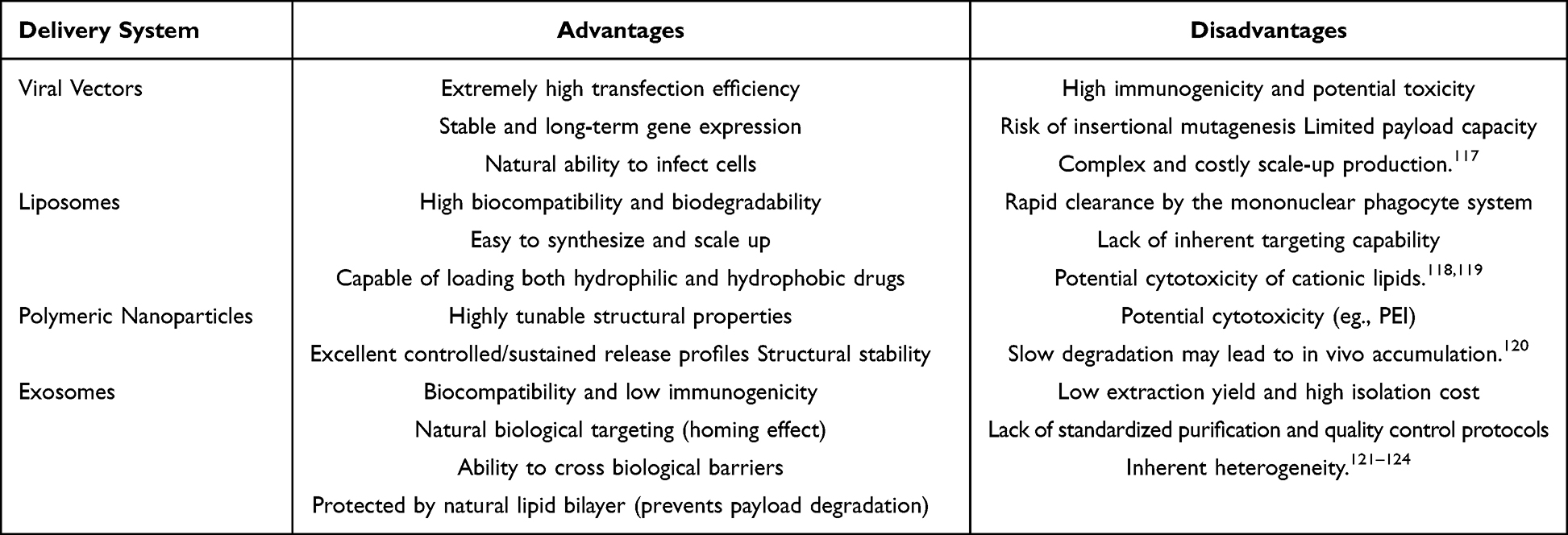 |
Table 3 Comparison of Drug Delivery Systems |
Applications in Cancer Therapy
Exosomes have shown tremendous potential in disease applications, particularly in cancer precision therapy and immunotherapy. They exhibit high heterogeneity in size and molecular composition, making them highly valuable for various applications (Figure 4).
 |
Figure 4 Exosome-Mediated Modulation of Cancer Immune Responses. Exosomes modulate the tumor immune microenvironment through multiple mechanisms, thereby influencing cancer progression and therapeutic responses. Exosomes can carry major MHC I and MHC II molecules as well as tumor-associated antigens (TAAs), which are either taken up by antigen-presenting cells (APCs), such as dendritic cells, or directly interact with T cells. This leads to the activation of CD8⁺ cytotoxic T lymphocytes via MHC I and CD4⁺ helper T cells via MHC II, thus inducing anti-tumor immune responses. The cargoes of exosomes are internalized by recipient cells or APCs through protein-mediated endocytosis or direct fusion with the plasma membrane, facilitating the release of their contents into the cytoplasm. |
Exosome-Based Therapeutic Delivery
Delivery systems centered around exosomes have showcased considerable promise within the realm of cancer treatment.These systems can encapsulate potent anticancer drugs and be administered through multiple routes, directly targeting cancer cells for specific killing while minimizing damage to normal tissues. Common administration methods contain intravenous infusion, subcutaneous shot, intraperitoneal administration, intratumoral infusion, nasal delivery, and oral administration.125–131 All these routes have been proven effective for delivering exosomal drug carriers to target tissues.However, despite the achievements of exosome-based delivery systems in preclinical models, the direct application of exosomes in clinical cancer treatment still faces severe challenges, with the most critical bottleneck being their poor in vivo targeted delivery efficiency and significant off-target effects.132,133 Firstly, the actual delivery efficiency of exosomes is severely constrained at the intracellular level by “endosomal entrapment”. Studies have shown that the majority of exosomes internalized by targeted cancer cells are trapped in endosomes and eventually degraded in lysosomes; the proportion of their carried active molecules (especially large RNA or proteins) successfully released into the cytoplasm to exert biological functions is extremely low, greatly reducing the actual therapeutic efficiency of the drugs.134 Secondly, off-target effects are the biggest challenge in systemic administration. After systemic administration (such as the most commonly used intravenous injection), the vast majority of natural exosomes are rapidly captured and cleared by the mononuclear phagocyte system (MPS), resulting in a high proportion of the administered dose being highly concentrated in non-target organs such as the liver (Kupffer cells), spleen, and lungs.104,135 This significant off-target effect not only greatly reduces the effective therapeutic concentration reaching the tumor site but also easily triggers systemic toxic side effects, such as exosomes loaded with high concentrations of potent chemotherapeutic drugs (such as DOX, PTX) potentially causing severe non-targeted toxic damage to these clearance organs. In addition, the dense extracellular matrix and high interstitial pressure in the tumor microenvironment (TME) further constitute physical barriers, severely limiting the penetration of exosomes into deep tumor tissues and resulting in overall low delivery efficiency.136 Therefore, relying solely on the passive targeting of natural exosomes is insufficient to meet the demands of clinical precision treatment. To overcome these translational barriers, advanced surface engineering techniques (such as modifying specific targeting ligands to increase tumor affinity, or introducing “do not eat me” signals like CD47 or surface-attached targeting peptides to evade macrophage phagocytosis and enhance tumor homing) can be utilized to reshape the in vivo biodistribution of exosomes, minimizing off-target risks while substantially improving the specific delivery efficiency to tumor sites.
In response to the urgent need to overcome the aforementioned delivery and off-target problems, researchers have developed a variety of exosome systems from specific sources or through modification. For instance, paclitaxel (PTX), a widely used anti-cancer drug, has shown remarkable efficacy, particularly against ovarian cancer, breast cancer, and non-small cell lung cancer. Nevertheless, its use in clinical settings is constrained by factors such as low water solubility, significant toxicity, and the development of drug resistance.Research findings indicate that by delivering PTX through exosomes secreted by M1-type macrophages, it is possible to effectively avoid non-specific clearance, enhance the accumulation of the drug in tumor tissues, activate the NF-κB pathway, thereby enhancing the tumor immune response, reducing the toxicity of PTX and improving its bioavailability.This dual - action mechanism not only bolsters the immune response against tumors but also mitigates the toxicity of PTX while enhancing its availability in the body.131 Kim et al showed that paclitaxel-loaded exosomes preferentially accumulated in tumor tissues, enabling targeted drug release while minimizing cytotoxicity and immunogenic responses.137 Moreover, when PTX is encapsulated within exosomes derived from cerebral endothelial cells, it gains the ability to effectively cross the cerebrovascular barrier and reach glioblastoma cells. This highlights the remarkable targeting abilities of such exosome - based PTX delivery systems.138 Wei et al found that exosomes loaded with doxorubicin (DOX) exhibited a significantly higher IC50 in cardiomyocyte H9C2 cells compared to free DOX, suggesting that exosomes as carriers can substantially mitigate the cardiac toxicity of DOX.139 Cisplatin is a commonly used chemotherapy drug. Research indicates that exosomes can deliver cisplatin in a way that prevents endosomal entrapment, reducing drug resistance. In addition, it has been demonstrated that exosomes obtained from M1 macrophages can work in a synergistic manner to boost the therapeutic outcomes of cisplatin in a mouse - based lung cancer model.140 Exosomes can also deliver antisense oligonucleotides, lncRNAs, shRNAs, and circRNAs to modulate gene expression and inhibit tumor progression. For example, exosome-delivered lncRNA can inhibit the progression of bladder cancer, while exosomes loaded with circRNA BTG, an anti-proliferative factor, can Inhibit the growth and spread of glioma cells.141,142 Targeted delivery by exosomes significantly reduces the chances of tumor recurrence and metastasis, paving the way for long-term cancer treatment.
In addition to cancer therapy, engineered exosomes show broad potential in other areas, such as neurodegenerative diseases and autoimmune disorders.143–145 Exosomes have the ability to traverse physiological barrier, transporting therapeutic substances straight to impaired neurons. This process subsequently facilitates neuronal repair and regeneration.Exosomes can cross the blood-brain barrier, delivering therapeutic molecules directly to damaged neurons, thus promoting neuronal repair and regeneration.146 For example, in Alzheimer’s disease (AD) research, one study demonstrated that engineered exosomes could enhance the delivery efficiency of antibodies, significantly reducing amyloid plaque deposition in animal models and improving cognitive function. This research not only provides new directions for Alzheimer’s disease therapy but also offers novel approaches for treating other neurological disorders.147
Exosomes from Diverse Cells in Immune Regulation
In the complex and challenging field of cancer therapy, the regulatory role of the immune system is increasingly gaining attention, with exosomes emerging as a critical intercellular communication mediator. Exosomes secreted by various cellular sources are enriched with bioactive molecules and significantly contribute to modulating cancer-related immune responses.
Exosomes originating from immune cells dynamically modulate the tumor-associated immune landscape. Intercellular signaling among immune cells, including T lymphocytes,macrophages, dendritic cells (DCs), and B lymphocytes. Immune cell-derived exosomes can Avoid elimination by the immune system, extending their retention time in peripheral circulation.130 CD4+T cell-derived exosomes markedly potentiate B cell activation in a dosage-sensitive fashion, subsequently elevating antibody synthesis directed against the hepatitis B surface antigen (HBsAg).148 Shaobo Ruan et al utilized near-UV LED light (365 nm) to stimulate dendritic cells to secrete exosomes, significantly improving the exosome production rate while maintaining good biocompatibility and immunogenicity. Immunological markers of exosomes, such as CD9 and CD63, were detected using ExoView, revealing that most exosomes expressed CD9, and the number of CD63+ exosomes increased. This suggests that light induction has minimal impact on immune marker expression, efficiently inducing the production of immunologically active exosomes, providing a foundation for exosome-based immunotherapies.149 Exosome-based delivery of MSC-derived exosomes has also been investigated for enhancing the treatment of acute myocardial infarction (AMI). Huang et al found that myocardial delivery of exosomes improved cardiac function, reduced infarct area, and promoted angiogenesis compared to treatment with either exosomes or MSCs alone.150
Exosomes originating from non-T cell sources also demonstrate multifaceted roles in tumor-associated immune modulation. Specifically, tumor-secreted exosomes can encapsulate particular antigens and RNA, thereby fostering anti-tumor immune responses. Conversely, these exosomes may induce T cell apoptosis, impede maturation of monocytes and establish a inflammatory cancer microenvironment.151 Whitehead et al identified tumor-derived exosomes capable of activating cytotoxic T lymphocytes (CTLs) and suppressing the progression of mastocytomas and breast carcinomas.152 Additionally, tumor-derived exosomes displaying HSP70/bag-4 on their surface can trigger natural killer cell-mediated granzyme B release, resulting in tumor cell apoptosis.153 In a separate study, Huang et al engineered exosomes derived from Luc cells by encapsulating immunogenic cell death (ICD) agents, which activated dendritic cells (DCs) and promoted CD8+ T cell expansion.154 Furthermore, exosomes derived from LPS-activated macrophages promote IFN-γ release in reaction to hepatitis B virus surface antigen in mice.148 Wang’s group employed exosomes secreted by M1 macrophages as a delivery platform, leading to 2.3-to 4.1-fold increase in inflammatory signaling molecule expression.155 Concurrently, these exosomes triggered the activation of CD8+ T lymphocytes and natural killer cells.This dual therapeutic approach of “drug killing + immune activation” not only achieved high concentrations of PTX in tumor tissues but also enhanced therapeutic effects by modulating the tumor microenvironment (TME). Exosomes derived from M2 macrophages can strongly induce endothelial cell proliferation and angiogenesis by activating the HIF1AN/HIF-1α/VEGFA signaling pathway.156 Exosomes secreted by mesenchymal stem cells (MSCs) are capable of immune modulation and tissue regeneration. They achieve this by stimulating angiogenesis-related mediators within the tumor microenvironment, which in turn facilitates tumor progression. Exosomes derived from breast cancer cells (eg., MDA-MB-231) have been demonstrated to aberrantly promote CD4+ T cell activation and upregulate pro-inflammatory cytokines, highlighting their potential as novel immunotherapeutic agents.157
Exosome-Based Vaccines
Exosomes have significant potential as vaccines in immunology. In contrast to conventional vaccines, exosomes exhibit multiple benefits: minimal toxicity and antigenicity, release antigens for immune activation, and high versatility.158 Exosomes are particularly advantageous for immunotherapy as they can transport antigens which stimulate cytotoxic T lymphocytes, enabling their use as autologous vaccine candidates. For example, dendritic cell-derived exosomes express MHC- I and II, which are capable of eliciting anti-tumor immune responses.DC-EXO vaccines show tremendous potential in metastatic melanoma and hepatocellular carcinoma.159 Compared to DC vaccines, which are rapidly cleared by T lymphocytes, DC-derived exosome vaccines have a relatively longer lifespan and have been tested in multiple Phase I clinical trials.160–162 To provide a more comprehensive and clear perspective on the clinical translation of exosome-based therapies, Table 4 summarizes representative ongoing and completed clinical trials globally.
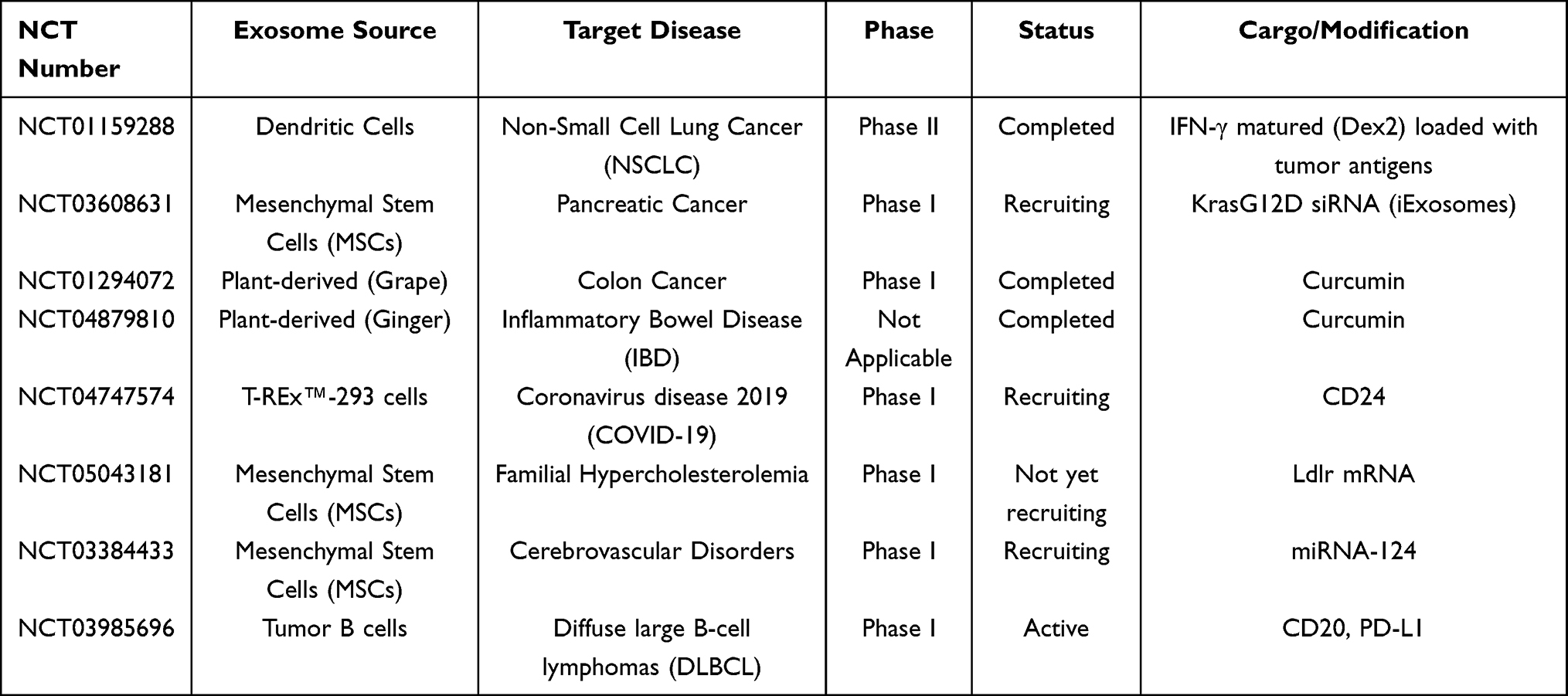 |
Table 4 Clinical Trials for Exosome-Based Cancer Therapies |
In contrast, tumor-secreted exosomes harbor numerous oncogenic microRNAs (miRNAs) that facilitate tumor growth and dissemination. Consequently, with continued refinement, exosome-based vaccines represent a promising innovation in cancer immunotherapy.163 Collectively, the application of exosomes as vaccines or delivery vehicles underscores their significant potential in immunology and therapeutic interventions, providing a secure and efficacious alternative to conventional vaccines.
Translational Challenges in Manufacturing and Safety
As study advances and technologies progress, engineered exosomes are expected to have a broader and deeper application in clinical fields. In the future, exosome-based delivery systems are likely to become an efficient and personalized therapeutic option, offering patients more treatment choices and better therapeutic outcomes. Despite exosomes offer superior drug-encapsulation potential, biocompatibility for delivery, the translation process from laboratory to clinic still faces significant challenges and stringent requirements in chemistry, manufacturing, and control (CMC).
From a regulatory and production perspective, achieving scalable, Good Manufacturing Practice (GMP)-compliant production remains a formidable bottleneck. Unlike synthetic nanoparticles, exosomes are intrinsically highly heterogeneous; their molecular cargo heavily depends on the dynamic metabolic state of the producer cells. This biological complexity results in significant batch-to-batch variability. Furthermore, isolating high-purity exosomes from large-scale bioreactors using current techniques (such as ultracentrifugation or size-exclusion chromatography) is costly and inefficient. More importantly, there is a critical regulatory void regarding standardized potency assays. Accurately defining the “active pharmaceutical ingredient” (API) equivalent of an exosome dose—and ensuring its functional consistency across different batches—remains a major unresolved challenge for quality control (QC).164–168
Equally critical are the safety concerns, particularly regarding the long-term biodistribution and potential pro-tumorigenic risks of specific exosome types. Although exosomes are generally considered highly biocompatible, their systemic administration often leads to rapid sequestration by the mononuclear phagocyte system (MPS), resulting in massive, unintended accumulation in the liver and spleen. Over time, this off-target biodistribution could trigger cumulative organ toxicity. Furthermore, when utilizing tumor-derived exosomes (eg., as vaccines), there are severe inherent risks. These vesicles may carry oncogenic mutations, pro-tumorigenic miRNAs, or immunosuppressive factors (such as PD-L1). Unintended horizontal gene transfer to healthy recipient cells could paradoxically promote pre-metastatic niche formation, accelerate tumor growth, or trigger unwanted neoangiogenesis.169–172 Therefore, meticulous molecular profiling and long-term in vivo safety monitoring are imperative before widespread clinical implementation.While advancing engineered exosome technology, ethical issues and regulatory challenges cannot be overlooked. How to promote the rapid translation of therapeutic exosomes while protecting patient rights is an urgent problem to be addressed in the field of precision medicine. Researchers and relevant institutions need to jointly establish corresponding standards and regulations to facilitate their healthy and orderly development.
Conclusions and Future Perspectives
We highlight the translational potential of exosome-based therapies, particularly their emerging applications as autologous vaccines and targeted nanocarriers in recent clinical trials for cancer treatment. We also explicitly address the challenges of off-target effects in vivo, technical barriers in macromolecular drug loading, regulatory and production issues, as well as long-term safety concerns of tumor-derived exosomes. In summary, exosomes, as a novel class of biological therapeutic carriers, have demonstrated significant potential in overcoming biological barriers and achieving targeted delivery, making them a research hotspot in the field of precision oncology. They serve a dual role as both precise diagnostic markers and multifunctional therapeutic vectors. Exosomes inherently possess the ability to cross biological barriers such as the blood-brain barrier, evade immune clearance, and precisely target receptor cells, offering unique advantages over traditional synthetic nanocarriers and viral vectors. Currently, this field is undergoing a critical transition phase, evolving from early phenomenological observations to standardized and regulated clinical applications.118,173
However, as previously mentioned, merely describing biological phenomena is insufficient to drive substantial breakthroughs in the field. To overcome current developmental bottlenecks, it is imperative to establish a standardized system for large-scale exosome production and systematically evaluate their long-term safety in various disease models. Concurrently, integrating cutting-edge technologies is essential for clinical translation: single-cell multi-omics and high-resolution spatial transcriptomics can deeply elucidate the intrinsic heterogeneity of exosome biosynthesis; artificial intelligence and machine learning algorithms can provide predictive modeling for exosome-target interactions, optimize the rational design of engineered vesicles, and lay the foundation for personalized medicine. To fully realize the translational value of exosome therapies, it is necessary to break through existing technological and biological barriers, shifting from single-discipline research to a paradigm of interdisciplinary collaboration. This requires deep integration of chemistry (surface functionalization), nanotechnology (efficient drug loading), oncology (analysis of tumor microenvironment interactions), and bioinformatics (AI-driven target screening). Only through unified multidisciplinary collaboration can exosomes be transformed from a promising biological phenomenon into standardized, safe, and efficient first-line clinical therapeutic carriers. Finally, it is essential to objectively acknowledge the inherent limitations of this review. Given the current explosive growth in exosome-related research, this review primarily focuses on oncology and engineered delivery systems, which may not comprehensively cover cutting-edge advancements in other therapeutic areas such as cardiovascular diseases and neurodegenerative disorders. Future systematic reviews and meta-analyses based on the MISEV standardized guidelines will provide critical support for further consolidating research conclusions and promoting standardized development in this field.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Natural Science Foundation of China (No.82303373), and Shenyang Young and Middle-aged Science and Technology Talents Support Program (RC230583).
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12–24. doi:10.3322/caac.21820
2. Johnstone RM, Adam M, Hammond JR, et al. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–9420. doi:10.1016/S0021-9258(18)48095-7
3. Valadi H, Ekström K, Bossios A, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–659. doi:10.1038/ncb1596
4. Alvarez-Erviti L, Seow Y, Yin H, et al. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
5. Rayamajhi S, Nguyen TDT, Marasini R, et al. Macrophage-derived exosome-mimetic hybrid vesicles for tumor targeted drug delivery. Acta Biomater. 2019;94:482–494. doi:10.1016/j.actbio.2019.05.054
6. Jamalkhah M, Asaadi Y, Azangou-Khyavy M, et al. MSC-derived exosomes carrying a cocktail of exogenous interfering RNAs an unprecedented therapy in era of COVID-19 outbreak. J Transl Med. 2021;19(1):164. doi:10.1186/s12967-021-02840-3
7. Wiklander OP, Nordin JZ, O’Loughlin A, et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4(1):26316. doi:10.3402/jev.v4.26316
8. Liu Z, Yan J, Tong L, et al. The role of exosomes from BALF in lung disease. J Cell Physiol. 2022;237(1):161–168. doi:10.1002/jcp.30553
9. Liao W, Du Y, Zhang C, et al. Exosomes: the next generation of endogenous nanomaterials for advanced drug delivery and therapy. Acta Biomater. 2019;86:1–14. doi:10.1016/j.actbio.2018.12.045
10. Ren J, He W, Zheng L, et al. From structures to functions: insights into exosomes as promising drug delivery vehicles. Biomater Sci. 2016;4(6):910–921. doi:10.1039/C5BM00583C
11. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
12. Mashouri L, Yousefi H, Aref AR, et al. Exosomes: composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol Cancer. 2019;18(1):75. doi:10.1186/s12943-019-0991-5
13. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30(1):255–289. doi:10.1146/annurev-cellbio-101512-122326
14. Wei D, Zhan W, Gao Y, et al. RAB31 marks and controls an ESCRT-independent exosome pathway. Cell Res. 2021;31(2):157–177. doi:10.1038/s41422-020-00409-1
15. Behzadi S, Serpooshan V, Tao W, et al. Cellular uptake of nanoparticles: journey inside the cell. Chem Soc Rev. 2017;46(14):4218–4244. doi:10.1039/C6CS00636A
16. Kaksonen M, Roux A. Mechanisms of clathrin-mediated endocytosis. Nat Rev Mol Cell Biol. 2018;19(5):313–326. doi:10.1038/nrm.2017.132
17. Vasave R, Bhawale R, Khan O, et al. Translational Potential of Surface Engineered Extracellular Vesicles as Theranostic Nanocarriers in Cancer Management: recent Developments from Bench to Bedside. ACS Appl Bio Mater. 2025;8(8):6647–6675. doi:10.1021/acsabm.5c00955
18. Johnson V, Vasu S, Kumar US, Kumar M. Surface-Engineered Extracellular Vesicles in Cancer Immunotherapy. Cancers. 2023;15(10):2838. doi:10.3390/cancers15102838
19. Hoshino A, Costa-Silva B, Shen TL, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527(7578):329–335. doi:10.1038/nature15756
20. McMahon HT, Boucrot E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nat Rev Mol Cell Biol. 2011;12(8):517–533. doi:10.1038/nrm3151
21. Tang D, Wang J, Kroemer G, Kang R. Targeting macropinocytosis for cancer therapy. Nat Rev Cancer. 2026;26(3):167–184. doi:10.1038/s41568-025-00892-x
22. Moon HR, Cho BK, Kang SH, Ryu JH, Kwon IC. Targeting and exploiting macropinocytosis in cancer therapy. J Control Release. 2025;385:113962. doi:10.1016/j.jconrel.2025.113962
23. Xu G, Zhang Q, Cheng R, Qu J, Li W. Survival strategies of cancer cells: the role of macropinocytosis in nutrient acquisition, metabolic reprogramming, and therapeutic targeting. Autophagy. 2025;21(4):693–718. doi:10.1080/15548627.2025.2452149
24. Zhao M, Zhou L, Zhai Y, Sun A, Shao G, Lin Q. Macropinocytosis: both a Target and a Tool for Cancer Therapy. Biomolecules. 2025;15(7):936. doi:10.3390/biom15070936
25. Li C, Liu Y, Liu C, et al. AGER-dependent macropinocytosis drives resistance to KRAS-G12D-targeted therapy in advanced pancreatic cancer. Sci Transl Med. 2025;17(783):eadp4986. doi:10.1126/scitranslmed.adp4986
26. Vermeulen LMP, De Smedt SC, Remaut K, Braeckmans K. The proton sponge hypothesis: fable or fact? Eur J Pharm Biopharm. 2018;129:184–190. doi:10.1016/j.ejpb.2018.05.034
27. Wojnilowicz M, Glab A, Bertucci A, Caruso F, Cavalieri F. Super-resolution Imaging of Proton Sponge-Triggered Rupture of Endosomes and Cytosolic Release of Small Interfering RNA. ACS Nano. 2019;13(1):187–202. doi:10.1021/acsnano.8b05151
28. Bonsergent E, Grisard E, Buchrieser J, Schwartz O, Théry C, Lavieu G. Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells. Nat Commun. 2021;12(1):1864. doi:10.1038/s41467-021-22126-y
29. Vermeulen LMP, Brans T, Samal SK, et al. Endosomal Size and Membrane Leakiness Influence Proton Sponge-Based Rupture of Endosomal Vesicles. ACS Nano. 2018;12(3):2332–2345. doi:10.1021/acsnano.7b07583
30. Zhuo Y, Luo Z, Zhu Z, et al. Direct cytosolic delivery of siRNA via cell membrane fusion using cholesterol-enriched exosomes. Nat Nanotechnol. 2024;19(12):1858–1868. doi:10.1038/s41565-024-01785-0
31. Ou X, Zhou J, Xie HY, Nie W. Fusogenic Lipid Nanovesicles as Multifunctional Immunomodulatory Platforms for Precision Solid Tumor Therapy. Small. 2025;21(27):e2503134. doi:10.1002/smll.202503134
32. Doherty GJ, McMahon HT. Mechanisms of endocytosis. Annu Rev Biochem. 2009;78(1):857–902. doi:10.1146/annurev.biochem.78.081307.110540
33. Chen YF, Zhou SK, Jian QH, et al. A Dual-Fusogenic Virus-like Vector Enables Direct Cytosolic Delivery of mRNA Vaccines to Dendritic Cells. ACS Nano. 2026;20(4):3551–3564. doi:10.1021/acsnano.5c16068
34. Meng Y, Lin Y, Luo Z, et al. Fusion Peptide-Incorporated Lipid Nanoparticles Boost Endosomal Escape and Enhance Cytosolic mRNA Delivery. Adv Mater. 2026;38(17):e15130. doi:10.1002/adma.202515130
35. Kumari P, Vasudevan SO, Russo AJ, et al. Host extracellular vesicles confer cytosolic access to systemic LPS licensing non-canonical inflammasome sensing and pyroptosis. Nat Cell Biol. 2023;25(12):1860–1872. doi:10.1038/s41556-023-01269-8
36. Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88(1):487–514. doi:10.1146/annurev-biochem-013118-111902
37. Zhang L, Yu D. Exosomes in cancer development, metastasis, and immunity. Biochim Biophys Acta Rev Cancer. 2019;1871(2):455–468. doi:10.1016/j.bbcan.2019.04.004
38. Chen J, Li Z, Yue C, et al. Human umbilical cord mesenchymal stem cell-derived exosomes carrying miR-1827 downregulate SUCNR1 to inhibit macrophage M2 polarization and prevent colorectal liver metastasis. Apoptosis. 2023;28(3–4):549–565. doi:10.1007/s10495-022-01798-x
39. Guo J, Duan Z, Zhang C, et al. Mouse 4T1 Breast Cancer Cell-Derived Exosomes Induce Proinflammatory Cytokine Production in Macrophages via miR-183. J Immunol. 2020;205(10):2916–2925. doi:10.4049/jimmunol.1901104
40. Ding Y, Cao F, Sun H, et al. Exosomes derived from human umbilical cord mesenchymal stromal cells deliver exogenous miR-145-5p to inhibit pancreatic ductal adenocarcinoma progression. Cancer Lett. 2019;442:351–361. doi:10.1016/j.canlet.2018.10.039
41. Usman WM, Pham TC, Kwok YY, et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun. 2018;9(1):2359. doi:10.1038/s41467-018-04791-8
42. Xie Y, Liu JB, Li JM, Zhang C, Lu CX, Wen ZJ. Effects of silencing circRNA ABCB10 expression on biological properties of colorectal cancer cells. Zhonghua Zhong Liu Za Zhi. 2021;43(4):449–456. doi:10.3760/cma.j.cn112152-20200116-00040
43. Morel KL, Hamid AA, Clohessy JG, Pandell N, Ellis L, Sweeney CJ. NF-κB Blockade with Oral Administration of Dimethylaminoparthenolide (DMAPT), Delays Prostate Cancer Resistance to Androgen Receptor (AR) Inhibition and Inhibits AR Variants. Mol Cancer Res. 2021;19(7):1137–1145. doi:10.1158/1541-7786.MCR-21-0099
44. Accolla RS, Ramia E, Tedeschi A, Forlani G. CIITA-Driven MHC Class II Expressing Tumor Cells as Antigen Presenting Cell Performers: toward the Construction of an Optimal Anti-tumor Vaccine. Front Immunol. 2019;10:1806. doi:10.3389/fimmu.2019.01806
45. Wang X, Chen T, Li C, et al. CircRNA-CREIT inhibits stress granule assembly and overcomes doxorubicin resistance in TNBC by destabilizing PKR. J Hematol Oncol. 2022;15(1):122. doi:10.1186/s13045-022-01345-w
46. Chen C, Shen M, Wan X, et al. Activated T cell-derived exosomes for targeted delivery of AXL-siRNA loaded paclitaxel-poly-L-lysine prodrug to overcome drug resistance in triple-negative breast cancer. Chem Eng J. 2023;468:143454. doi:10.1016/j.cej.2023.143454
47. Mousavi SM, Hosseindoost S, Mahdian SMA, et al. Exosomes released from U87 glioma cells treated with curcumin and/or temozolomide produce apoptosis in naive U87 cells. Pathol Res Pract. 2023;245:154427. doi:10.1016/j.prp.2023.154427
48. Poggio M, Hu T, Pai CC, et al. Suppression of Exosomal PD-L1 Induces Systemic Anti-tumor Immunity and Memory. Cell. 2019;177(2):414–427.e13. doi:10.1016/j.cell.2019.02.016
49. Srivastava A, Moxley K, Ruskin R, Dhanasekaran DN, Zhao YD, Ramesh R. A Non-invasive Liquid Biopsy Screening of Urine-Derived Exosomes for miRNAs as Biomarkers in Endometrial Cancer Patients. AAPS J. 2018;20(5):82. doi:10.1208/s12248-018-0220-y
50. Asakura K, Kadota T, Matsuzaki J, et al. A miRNA-based diagnostic model predicts resectable lung cancer in humans with high accuracy. Commun Biol. 2020;3(1):134. doi:10.1038/s42003-020-0863-y
51. Hannafon BN, Trigoso YD, Calloway CL, et al. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016;18(1):90. doi:10.1186/s13058-016-0753-x
52. Vahabi A, Rezaie J, Hassanpour M, et al. Tumor Cells-derived exosomal CircRNAs: novel cancer drivers, molecular mechanisms, and clinical opportunities. Biochem Pharmacol. 2022;200:115038. doi:10.1016/j.bcp.2022.115038
53. Zhuo CJ, Hou WH, Jiang DG, et al. Circular RNAs in early brain development and their influence and clinical significance in neuropsychiatric disorders. Neural Regen Res. 2020;15(5):817–823. doi:10.4103/1673-5374.268969
54. Sidhom K, Obi PO, Saleem A. A Review of Exosomal Isolation Methods: is Size Exclusion Chromatography the Best Option? Int J Mol Sci. 2020;21(18):6466. doi:10.3390/ijms21186466
55. Nie W, Wu G, Zhang J, et al. Responsive Exosome Nano-bioconjugates for Synergistic Cancer Therapy. Angew Chem Int Ed Engl. 2020;59(5):2018–2022. doi:10.1002/anie.201912524
56. Jiang Z, Liu G, Li J. Recent Progress on the Isolation and Detection Methods of Exosomes. Chem Asian J. 2020;15(23):3973–3982. doi:10.1002/asia.202000873
57. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jconrel.2022.05.027
58. Linares R, Tan S, Gounou C, et al. High-speed centrifugation induces aggregation of extracellular vesicles. J Extracell Vesicles. 2015;4(1):29509. doi:10.3402/jev.v4.29509
59. Witwer KW, Buzás EI, Bemis LT, et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J Extracell Vesicles. 2013;2(1):10. doi:10.3402/jev.v2i0.20360
60. Yin Y, Liu B, Cao Y, et al. Colorectal Cancer-Derived Small Extracellular Vesicles Promote Tumor Immune Evasion by Upregulating PD-L1 Expression in Tumor-Associated Macrophages. Adv Sci. 2022;9(9):2102620. doi:10.1002/advs.202102620
61. Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in Exosome Isolation Techniques. Theranostics. 2017;7(3):789–804. doi:10.7150/thno.18133
62. Han S, Xu Y, Sun J, et al. Isolation and analysis of extracellular vesicles in a Morpho butterfly wing-integrated microvortex biochip. Biosens Bioelectron. 2020;154:112073. doi:10.1016/j.bios.2020.112073
63. Chen C, Skog J, Hsu CH, et al. Microfluidic isolation and transcriptome analysis of serum microvesicles. Lab Chip. 2010;10(4):505–511. doi:10.1039/B916199F
64. Chen Y, Zhu Q, Cheng L, et al. Exosome detection via the ultrafast-isolation system: EXODUS. Nat Methods. 2021;18(2):212–218. doi:10.1038/s41592-020-01034-x
65. Zhu Q, Li H, Ao Z, et al. Lipidomic identification of urinary extracellular vesicles for non-alcoholic steatohepatitis diagnosis. J Nanobiotechnology. 2022;20(1):349. doi:10.1186/s12951-022-01540-4
66. Gardiner C, Ferreira YJ, Dragovic RA, et al. Extracellular vesicle sizing and enumeration by nanoparticle tracking analysis. J Extracell Vesicles. 2013;2(1):10. doi:10.3402/jev.v2i0.19671
67. Kurian TK, Banik S, Gopal D, et al. Elucidating Methods for Isolation and Quantification of Exosomes: a Review. Mol Biotechnol. 2021;63(4):249–266. doi:10.1007/s12033-021-00300-3
68. Ashcroft BA, de Sonneville J, Yuana Y, et al. Determination of the size distribution of blood microparticles directly in plasma using atomic force microscopy and microfluidics. Biomed Microdevices. 2012;14(4):641–649. doi:10.1007/s10544-012-9642-y
69. Sonbhadra S, Mehak, Pandey LM. Biogenesis, Isolation, and Detection of Exosomes and Their Potential in Therapeutics and Diagnostics. Biosensors. 2023;13(8):802. doi:10.3390/bios13080802
70. Saad MG, Beyenal H, Dong WJ. Exosomes as Powerful Engines in Cancer: isolation, Characterization and Detection Techniques. Biosensors. 2021;11(12):518. doi:10.3390/bios11120518
71. Zhang C, Guo Z, Jing Z. Prediction of Response to Chemoradiotherapy by Dynamic Changes of Circulating Exosome Levels in Patients with Esophageal Squamous Cell Carcinoma. Int J Nanomed. 2024;19:1351–1362. doi:10.2147/IJN.S440684
72. Liu H, Tian Y, Xue C, et al. Analysis of extracellular vesicle DNA at the single-vesicle level by nano-flow cytometry. J Extracell Vesicles. 2022;11(4). doi:10.1002/jev2.12206.
73. Rao D, Lu H, Wang X, et al. Tissue-derived exosome proteomics identifies promising diagnostic biomarkers for esophageal cancer. eLife. 2023;12:1.
74. Visan KS, Lobb RJ, Ham S, et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J Extracell Vesicles. 2022;11(9). doi:10.1002/jev2.12266.
75. You Y, Zhang Z, Sultana N, et al. ATP1A3 as a target for isolating neuron-specific extracellular vesicles from human brain and biofluids. Sci Adv. 2023;9(37). doi:10.1126/sciadv.adi3647.
76. Lu JY, Guo Z, Huang WT, et al. Peptide-graphene logic sensing system for dual-mode detection of exosomes, molecular information processing and protection. Talanta. 2024;267:125261. doi:10.1016/j.talanta.2023.125261
77. Cully M. Exosome-based candidates move into the clinic. Nat Rev Drug Discov. 2021;20(1):6–7. doi:10.1038/d41573-020-00220-y
78. Jin Y, Ma L, Zhang W, et al. Extracellular signals regulate the biogenesis of extracellular vesicles. Biol Res. 2022;55(1):35. doi:10.1186/s40659-022-00405-2
79. Li MY, Liu LZ, Dong M. Progress on pivotal role and application of exosome in lung cancer carcinogenesis, diagnosis, therapy and prognosis. Mol Cancer. 2021;20(1):22. doi:10.1186/s12943-021-01312-y
80. Lee J, Lee H, Goh U, et al. Cellular Engineering with Membrane Fusogenic Liposomes to Produce Functionalized Extracellular Vesicles. ACS Appl Mater Interfaces. 2016;8(11):6790–6795. doi:10.1021/acsami.6b01315
81. Pascucci L, Coccè V, Bonomi A, et al. Paclitaxel is incorporated by mesenchymal stromal cells and released in exosomes that inhibit in vitro tumor growth: a new approach for drug delivery. J Control Release. 2014;192:262–270. doi:10.1016/j.jconrel.2014.07.042
82. Zhu L, Kalimuthu S, Oh JM, et al. Enhancement of antitumor potency of extracellular vesicles derived from natural killer cells by IL-15 priming. Biomaterials. 2019;190-191:38–50. doi:10.1016/j.biomaterials.2018.10.034
83. Liu Q, Li D, Pan X, Liang Y. Targeted therapy using engineered extracellular vesicles: principles and strategies for membrane modification. J Nanobiotechnology. 2023;21(1):334. doi:10.1186/s12951-023-02081-0
84. Ha D, Yang N, Nadithe V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: current perspectives and future challenges. Acta Pharm Sin B. 2016;6(4):287–296. doi:10.1016/j.apsb.2016.02.001
85. Lou G, Chen L, Xia C, et al. MiR-199a-modified exosomes from adipose tissue-derived mesenchymal stem cells improve hepatocellular carcinoma chemosensitivity through mTOR pathway. J Exp Clin Cancer Res. 2020;39(1):4. doi:10.1186/s13046-019-1512-5
86. Yan C, Chen J, Wang C, et al. Milk exosomes-mediated miR-31-5p delivery accelerates diabetic wound healing through promoting angiogenesis. Drug Deliv. 2022;29(1):214–228. doi:10.1080/10717544.2021.2023699
87. Xiang X, Chen J, Jiang T, et al. Milk-derived exosomes carrying siRNA-KEAP1 promote diabetic wound healing by improving oxidative stress. Drug Deliv Transl Res. 2023;13(9):2286–2296. doi:10.1007/s13346-023-01306-x
88. Yuan R, Mu Z, Zhang H, et al. Ultrasonic Microfluidic Method Used for siHSP47 Loaded in Human Embryonic Kidney Cell-Derived Exosomes for Inhibiting TGF-β1 Induced Fibroblast Differentiation and Migration. Int J Mol Sci. 2025;26(1):382. doi:10.3390/ijms26010382
89. Chen Q, Che C, Liu J, et al. Construction of an exosome-functionalized graphene oxide based composite bionic smart drug delivery system and its anticancer activity. Nanotechnology. 2022;33(17):10. doi:10.1088/1361-6528/ac49bf
90. Sun D, Zhuang X, Xiang X, et al. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol Ther. 2010;18(9):1606–1614. doi:10.1038/mt.2010.105
91. Fuhrmann G, Serio A, Mazo M, Nair R, Stevens MM. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J Control Release. 2015;205:35–44. doi:10.1016/j.jconrel.2014.11.029
92. Mishra A, Singh P, Qayoom I, et al. Current strategies in tailoring methods for engineered exosomes and future avenues in biomedical applications. J Mater Chem B. 2021;9(32):6281–6309. doi:10.1039/D1TB01088C
93. Tianjiao G, Ling D, Liu M. Mengyang L.et al.Preservation of extracellular vesicles for drug delivery: a comparative evaluation of storage buffers. J Drug Delivery Sci Technol. 2025;107:106850. doi:10.1016/j.jddst.2025.106850
94. Koh E, Lee EJ, Nam GH, et al. Exosome-SIRPα, a CD47 blockade increases cancer cell phagocytosis. Biomaterials. 2017;121:121–129. doi:10.1016/j.biomaterials.2017.01.004
95. Pei W, Zhang Y, Zhu X, et al. Multitargeted Immunomodulatory Therapy for Viral Myocarditis by Engineered Extracellular Vesicles. ACS Nano. 2024;18(4):2782–2799. doi:10.1021/acsnano.3c05847
96. Yuan H, Chen Y, Hu Y, et al. Disulfide bond-driven nanoassembly of lipophilic epirubicin prodrugs for breast cancer therapy. J. Pharm. Investig. 2025;55:889–902.
97. Ohno S, Takanashi M, Sudo K, et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther. 2013;21(1):185–191. doi:10.1038/mt.2012.180
98. Liu Y, Li D, Liu Z, et al. Targeted exosome-mediated delivery of opioid receptor Mu siRNA for the treatment of morphine relapse. Sci Rep. 2015;5(1):17543. doi:10.1038/srep17543
99. Razlansari M, Jafarinejad S, Rahdar A, et al. Development and classification of RNA aptamers for therapeutic purposes: an updated review with emphasis on cancer. Mol Cell Biochem. 2023;478(7):1573–1598. doi:10.1007/s11010-022-04614-x
100. Pi F, Binzel DW, Lee TJ, et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat Nanotechnol. 2018;13(1):82–89. doi:10.1038/s41565-017-0012-z
101. Wiklander OPB, Mamand DR, Mohammad DK, et al. Antibody-displaying extracellular vesicles for targeted cancer therapy. Nat Biomed Eng. 2024;8(11):1453–1468. doi:10.1038/s41551-024-01214-6
102. Huang P, Wang L, Li Q, et al. Combinatorial treatment of acute myocardial infarction using stem cells and their derived exosomes resulted in improved heart performance. Stem Cell Res Ther. 2019;10(1):300. doi:10.1186/s13287-019-1353-3
103. Sarkar R, Biswas S, Ghosh R, et al. Exosome-sheathed porous silica nanoparticle-mediated co-delivery of 3,3’-diindolylmethane and doxorubicin attenuates cancer stem cell-driven EMT in triple negative breast cancer. J Nanobiotechnology. 2024;22(1):285. doi:10.1186/s12951-024-02518-0
104. Tenchov R, Sasso JM, Wang X, et al. Exosomes─Nature’s Lipid Nanoparticles, a Rising Star in Drug Delivery and Diagnostics. ACS Nano. 2022;16(11):17802–17846. doi:10.1021/acsnano.2c08774
105. Lin Y, Cheng Q, Wei T. Surface engineering of lipid nanoparticles: targeted nucleic acid delivery and beyond. Biophys Rep. 2023;9(5):255–278. doi:10.52601/bpr.2023.230022
106. Aggarwal P, Hall JB, McLeland CB, Dobrovolskaia MA, McNeil SE. Nanoparticle interaction with plasma proteins as it relates to particle biodistribution, biocompatibility and therapeutic efficacy. Adv Drug Deliv Rev. 2009;61(6):428–437. doi:10.1016/j.addr.2009.03.009
107. Kamerkar S, LeBleu VS, Sugimoto H, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546(7659):498–503. doi:10.1038/nature22341
108. Farooq MA, Johnston APR, Trevaskis NL. Impact of nanoparticle properties on immune cell interactions in the lymph node. Acta Biomater. 2025;193:65–82. doi:10.1016/j.actbio.2024.12.039
109. Yim N, Ryu SW, Choi K, et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein-protein interaction module. Nat Commun. 2016;7(1):12277. doi:10.1038/ncomms12277
110. Familtseva A, Jeremic N, Tyagi SC. Exosomes: cell-created drug delivery systems. Mol Cell Biochem. 2019;459(1–2):1–6. doi:10.1007/s11010-019-03545-4
111. Gee P, Lung MSY, Okuzaki Y, et al. Extracellular nanovesicles for packaging of CRISPR-Cas9 protein and sgRNA to induce therapeutic exon skipping. Nat Commun. 2020;11(1):1334. doi:10.1038/s41467-020-14957-y
112. Kojima R, Bojar D, Rizzi G, et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat Commun. 2018;9(1):1305. doi:10.1038/s41467-018-03733-8
113. Aslan C, Zolbanin NM, Faraji F, Jafari R. Exosomes for CRISPR-Cas9 Delivery: the Cutting Edge in Genome Editing. Mol Biotechnol. 2024;66(11):3092–3116. doi:10.1007/s12033-023-00932-7
114. Kim SM, Yang Y, Oh SJ, et al. Cancer-derived exosomes as a delivery platform of CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J Control Release. 2017;266:8–16. doi:10.1016/j.jconrel.2017.09.013
115. Jia G, Han Y, An Y, et al. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials. 2018;178:302–316. doi:10.1016/j.biomaterials.2018.06.029
116. Shaikh S, Younis M, Yingying S, et al. Bleomycin loaded exosomes enhanced antitumor therapeutic efficacy and reduced toxicity. Life Sci. 2023;330:121977. doi:10.1016/j.lfs.2023.121977
117. Tenchov R, Bird R, Curtze AE, Zhou Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano. 2021;15(11):16982–17015. doi:10.1021/acsnano.1c04996
118. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16(7):748–759. doi:10.1038/s41565-021-00931-2
119. Mitchell MJ, Billingsley MM, Haley RM, et al. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
120. Gregoriadis G. Liposomes in Drug Delivery: how It All Happened. Pharmaceutics. 2016;8(2):19. doi:10.3390/pharmaceutics8020019
121. Evers MJW, van de Wakker SI, de Groot EM, et al. Functional siRNA Delivery by Extracellular Vesicle-Liposome Hybrid Nanoparticles. Adv Healthc Mater. 2022;11(5):e2101202. doi:10.1002/adhm.202101202
122. Bradley JA, Bolton EM, Pedersen RA. Stem cell medicine encounters the immune system. Nat Rev Immunol. 2002;2(11):859–871. doi:10.1038/nri934
123. Dougherty JA, Kumar N, Noor M, et al. Extracellular Vesicles Released by Human Induced-Pluripotent Stem Cell-Derived Cardiomyocytes Promote Angiogenesis. Front Physiol. 2018;9:1794. doi:10.3389/fphys.2018.01794
124. Zhao L, Gu C, Gan Y, Shao L, Chen H, Zhu H. Exosome-mediated siRNA delivery to suppress postoperative breast cancer metastasis. J Control Release. 2020;318:1–15. doi:10.1016/j.jconrel.2019.12.005
125. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
126. Tian Y, Li S, Song J, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials. 2014;35(7):2383–2390. doi:10.1016/j.biomaterials.2013.11.083
127. Yang G, Chen Q, Wen D, et al. A Therapeutic Microneedle Patch Made from Hair-Derived Keratin for Promoting Hair Regrowth. ACS Nano. 2019;13(4):4354–4360. doi:10.1021/acsnano.8b09573
128. Sheller-Miller S, Choi K, Choi C, Menon R. Cyclic-recombinase-reporter mouse model to determine exosome communication and function during pregnancy. Am J Obstet Gynecol. 2019;221(5):
129. Mizrak A, Bolukbasi MF, Ozdener GB, et al. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol Ther. 2013;21(1):101–108. doi:10.1038/mt.2012.161
130. Sterzenbach U, Putz U, Low LH, et al. Engineered Exosomes as Vehicles for Biologically Active Proteins. Mol Ther. 2017;25(6):1269–1278. doi:10.1016/j.ymthe.2017.03.030
131. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
132. Wang P, Wang H, Huang Q, et al. Exosomes from M1-Polarized Macrophages Enhance Paclitaxel Antitumor Activity by Activating Macrophages-Mediated Inflammation. Theranostics. 2019;9(6):1714–1727. doi:10.7150/thno.30716
133. Wiklander OPB, Brennan MÁ, Lötvall J, et al. Advances in therapeutic applications of extracellular vesicles. Sci Transl Med. 2019;11(492):eaav8521. doi:10.1126/scitranslmed.aav8521
134. Mentkowski KI, Snitzer JD, Rusnak S, Lang JK. Therapeutic Potential of Engineered Extracellular Vesicles. AAPS J. 2018;20(3):50. doi:10.1208/s12248-018-0211-z
135. Joshi BS, de Beer MA, Giepmans BNG, Zuhorn IS. Endocytosis of Extracellular Vesicles and Release of Their Cargo from Endosomes. ACS Nano. 2020;14(4):4444–4455. doi:10.1021/acsnano.9b10033
136. Du W, Chen C, Liu Y, et al. A combined “eat me/don’t eat me” strategy based on exosome for acute liver injury treatment. Cell Rep Med. 2025;6(4):102033. doi:10.1016/j.xcrm.2025.102033
137. Liang G, Zhu Y, Ali DJ, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnology. 2020;18(1):10. doi:10.1186/s12951-019-0563-2
138. Ye Y, Zhang X, Xie F, et al. An engineered exosome for delivering sgRNA:Cas9 ribonucleoprotein complex and genome editing in recipient cells. Biomater Sci. 2020;8(10):2966–2976. doi:10.1039/D0BM00427H
139. Wei H, Chen J, Wang S, et al. A Nanodrug Consisting Of Doxorubicin And Exosome Derived From Mesenchymal Stem Cells For Osteosarcoma Treatment In Vitro. Int J Nanomed. 2019;14:8603–8610. doi:10.2147/IJN.S218988
140. Zhou G, Gu Y, Zhu Z, et al. Exosome Mediated Cytosolic Cisplatin Delivery Through Clathrin-Independent Endocytosis and Enhanced Anti-cancer Effect via Avoiding Endosome Trapping in Cisplatin-Resistant Ovarian Cancer. Front Med Lausanne. 2022;9:810761. doi:10.3389/fmed.2022.810761
141. Zheng R, Du M, Wang X, et al. Exosome-transmitted long non-coding RNA PTENP1 suppresses bladder cancer progression. Mol Cancer. 2018;17(1):143. doi:10.1186/s12943-018-0880-3
142. Shi L, Cao Y, Yuan W, Guo J, Sun G. Exosomal circRNA BTG2 derived from RBP-J overexpressed-macrophages inhibits glioma progression via miR-25-3p/PTEN. Cell Death Dis. 2022;13(5):506. doi:10.1038/s41419-022-04908-4
143. Hu X, Ning X, Zhao Q, et al. Islet-1 Mesenchymal Stem Cells-Derived Exosome-Incorporated Angiogenin-1 Hydrogel for Enhanced Acute Myocardial Infarction Therapy. ACS Appl Mater Interfaces. 2022;14(32):36289–36303. doi:10.1021/acsami.2c04686
144. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med. 2011;365(23):2205–2219. doi:10.1056/NEJMra1004965
145. You DG, Lim GT, Kwon S, et al. Metabolically engineered stem cell-derived exosomes to regulate macrophage heterogeneity in rheumatoid arthritis. Sci Adv. 2021;7(23):eabe0083. doi:10.1126/sciadv.abe0083
146. Qu M, Lin Q, Huang L, et al. Dopamine-loaded blood exosomes targeted to brain for better treatment of Parkinson’s disease. J Control Release. 2018;287:156–166. doi:10.1016/j.jconrel.2018.08.035
147. Passeri E, Elkhoury K, Morsink M, et al. Alzheimer’s Disease: treatment Strategies and Their Limitations. Int J Mol Sci. 2022;23(22):13954. doi:10.3390/ijms232213954
148. Sanderson RD, Bandari SK, Vlodavsky I. Proteases and glycosidases on the surface of exosomes: newly discovered mechanisms for extracellular remodeling. Matrix Biol. 2019;75-76:160–169. doi:10.1016/j.matbio.2017.10.007
149. Jesus S, Soares E, Cruz MT, Borges O. Exosomes as adjuvants for the recombinant hepatitis B antigen: first report. Eur J Pharm Biopharm. 2018;133:1–11. doi:10.1016/j.ejpb.2018.09.029
150. Lu J, Wu J, Xie F, et al. CD4+ T Cell-Released Extracellular Vesicles Potentiate the Efficacy of the HBsAg Vaccine by Enhancing B Cell Responses. Adv Sci. 2019;6(23):1802219. doi:10.1002/advs.201802219
151. Kim MS, Haney MJ, Zhao Y, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–664. doi:10.1016/j.nano.2015.10.012
152. Taylor DD, Gercel-Taylor C. Exosomes/microvesicles: mediators of cancer-associated immunosuppressive microenvironments. Semin Immunopathol. 2011;33(5):441–454. doi:10.1007/s00281-010-0234-8
153. Ruan S, Erwin N, He M. Light-induced high-efficient cellular production of immune functional extracellular vesicles. J Extracell Vesicles. 2022;11(3):e12194. doi:10.1002/jev2.12194
154. Wesselhoeft RA, Kowalski PS, Parker-Hale FC, et al. RNA Circularization Diminishes Immunogenicity and Can Extend Translation Duration In Vivo. Mol Cell. 2019;74(3):508–520.e4. doi:10.1016/j.molcel.2019.02.015
155. Esteves M, Abreu R, Fernandes H, et al. MicroRNA-124-3p-enriched small extracellular vesicles as a therapeutic approach for Parkinson’s disease. Mol Ther. 2022;30(10):3176–3192. doi:10.1016/j.ymthe.2022.06.003
156. Zhu Q, Ling X, Yang Y, et al. Embryonic Stem Cells-Derived Exosomes Endowed with Targeting Properties as Chemotherapeutics Delivery Vehicles for Glioblastoma Therapy. Adv Sci. 2019;6(6):1801899. doi:10.1002/advs.201801899
157. Luo G, Zhou Z, Cao Z, et al. M2 macrophage-derived exosomes induce angiogenesis and increase skin flap survival through HIF1AN/HIF-1α/VEGFA control. Arch Biochem Biophys. 2024;751:109822. doi:10.1016/j.abb.2023.109822
158. Mustafa O, Alper TO, Hulya O, et al. Immunomodulatory Effects of MDA-MB-231-derived Exosome Mimetic Nanovesicles on CD4+ T Cell Line. Eurasian J Med Oncol. 2024;8(1):40–48.
159. Huang Z, Zhang Y, Zhou J, Zhang Y. Urinary Exosomal miR-193a Can Be a Potential Biomarker for the Diagnosis of Primary Focal Segmental Glomerulosclerosis in Children. Biomed Res Int. 2017;2017:7298160. doi:10.1155/2017/7298160
160. Santos P, Almeida F. Exosome-Based Vaccines: history, Current State, and Clinical Trials. Front Immunol. 2021;12:711565. doi:10.3389/fimmu.2021.711565
161. Lu Z, Zuo B, Jing R, et al. Dendritic cell-derived exosomes elicit tumor regression in autochthonous hepatocellular carcinoma mouse models. J Hepatol. 2017;67(4):739–748. doi:10.1016/j.jhep.2017.05.019
162. Dai S, Wei D, Wu Z, et al. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol Ther. 2008;16(4):782–790. doi:10.1038/mt.2008.1
163. Bourderioux M, Nguyen-Khoa T, Chhuon C, et al. A new workflow for proteomic analysis of urinary exosomes and assessment in cystinuria patients. J Proteome Res. 2015;14(1):567–577. doi:10.1021/pr501003q
164. Rezaie J, Feghhi M, Etemadi T. A review on exosomes application in clinical trials: perspective, questions, and challenges. Cell Commun Signal. 2022;20(1):145. doi:10.1186/s12964-022-00959-4
165. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
166. Cheng CA. Before Translating Extracellular Vesicles into Personalized Diagnostics and Therapeutics: what We Could Do. Mol Pharm. 2024;21(6):2625–2636. doi:10.1021/acs.molpharmaceut.4c00185
167. Gimona M, Brizzi MF, Choo ABH, et al. Critical considerations for the development of potency tests for therapeutic applications of mesenchymal stromal cell-derived small extracellular vesicles. Cytotherapy. 2021;23(5):373–380. doi:10.1016/j.jcyt.2021.01.001
168. Tan TT, Toh WS, Lai RC, Lim SK. Practical considerations in transforming MSC therapy for neurological diseases from cell to EV. Exp Neurol. 2022;349:113953. doi:10.1016/j.expneurol.2021.113953
169. Witwer KW, Van Balkom BWM, Bruno S, et al. Defining mesenchymal stromal cell (MSC)-derived small extracellular vesicles for therapeutic applications. J Extracell Vesicles. 2019;8(1):1609206. doi:10.1080/20013078.2019.1609206
170. Becker A, Thakur BK, Weiss JM, et al. Extracellular Vesicles in Cancer: cell-to-Cell Mediators of Metastasis. Cancer Cell. 2016;30(6):836–848. doi:10.1016/j.ccell.2016.10.009
171. Dos Anjos Pultz B, Andrés Cordero da Luz F, Socorro Faria S, et al. The multifaceted role of extracellular vesicles in metastasis: priming the soil for seeding. Int, J, Cancer. 2017;140(11):2397–2407. doi:10.1002/ijc.30595
172. Xu R, Rai A, Chen M, et al. Extracellular vesicles in cancer-implications for future improvements in cancer care. Nat Rev Clin Oncol. 2018;15(10):617–638. doi:10.1038/s41571-018-0036-9
173. Clancy JW, D’Souza-Schorey C. Tumor-Derived Extracellular Vesicles: multifunctional Entities in the Tumor Microenvironment. Annu Rev Pathol. 2023;18(1):205–229. doi:10.1146/annurev-pathmechdis-031521-022116
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Current Advances in Nanocarriers for Cancer Therapy
Zeinali R, Zaeifi D, Zolfaghari-Moghaddam SY, Paul MK, Biazar E
International Journal of Nanomedicine 2025, 20:12217-12262
Published Date: 7 October 2025